1. Introduction: Neural Representations and Their (Dis)Contents
Representations figure prominently in several human affairs. Human beings routinely use representational artifacts like maps to navigate their environments. Maps represent the terrain to be traversed, for an agent capable of reading it and of leveraging the information that it contains to guide their behavior. It is quite uncontroversial to claim that human beings consciously and deliberately engage in intellectual tasks, such as theorizing about causes and effects—which entails the ability to mentally think about situations and states of affairs. Most of us can see in our mind’s eye situations past, possible, and fictive, via the imagination and mental imagery, which are traditionally cast in terms of representational abilities.
In the cognitive sciences, neurosciences, and the philosophy of mind, the concept of representation has been used to try and explain naturalistically how the fundamental property of ‘aboutness’ or ‘intentionality’ emerges in living systems [
1]. Indeed, living creatures must interact with the world in which they are embedded, and must distinguish environmental features and other organisms that are relevant for their survival, from those that are not. Living creatures act as if they had beliefs about the world, about its structure and its denizens, which guide their decision-making processes, especially with respect to the generation of adaptive action. This property of aboutness is thus a foundational one for any system that must make
probabilistic inferences to support their
decision-making in an uncertain world, which are central to the special issue to which this paper contributes.
In this context, to provide a naturalistic explanation is to explain some phenomenon by appealing to physical processes [
2]. The strategy deployed by cognitive science has been to naturalize intentionality by postulating the existence of physical structures internal to the agent that carry, encode, or otherwise bear semantic content; classical accounts include Fodor [
3], Millikan [
4,
5].
In the sciences that study the mind and in philosophy, representations are typically defined as some internal vehicles—
neural representations—that carry semantic content [
6]. Representations are thus physical structures that are internal to an organism; typically, states and processes unfolding in their brains, which carry or encode representational content. The epistemic role played by neural representations is to explain how creatures are able to engage with relevant features of their environment and plan situationally appropriate, adaptive behavior [
6,
7]. The semantic content of a representation is what the representation is about, that towards which it stands in an intentional relation—that “in virtue of what they represent what they do, or get to be ‘about’ what they are about” ([
8], p. 2390). The problem of specifying the nature and origins of semantic content is known as the hard problem of content [
9,
10].
There are several well-accepted constraints for the appropriateness of representational explanations: such an account should (1) cohere broadly with the actual practices that are used in computational cognitive science research; (2) allow for misrepresentation, i.e., the representation must be able to “get it wrong”; (3) provide the principled method for attributing of determinate contents to specific states or structures (typically internal to the system), and finally (4) be naturalistic, meaning that the account of semantic content does not itself appeal to semantic terms when defining how the representational capacity is realized by the physical system, on pain of circularity in reasoning [
11,
12].
A fundamental and thorny question is whether there is some construct of representation that not only applies in all cases (i.e., to the construct of neural representations and to our more familiar, deliberate, everyday representational activities), but that also really explains the intentional relation between living creatures and their environment. An equally thorny issue involves the ontological and epistemological status of neural representations: Do such things really exist? Do they have explanatory value?
The aim of this paper is twofold. First, we aim to determine whether or not the construct of neural representations plays an explanatory role in an increasingly popular framework for the study of action and cognition in living systems, namely, the variational free-energy principle and its corollary process theory, active inference. Second, if the postulation of neural representations is warranted under the free-energy principle, we aim to assess which of the available philosophical positions about the ontological and epistemological status of representations is most appropriate for the construct under this framework. Since the issue to be determined cannot merely be decided by appeal to formal frameworks, we first discuss the issue of representationalism.
In the remainder of this first section, we review the issues surrounding representationalism. In the second section, we present and motivate the view of the brain under the free-energy principle, as a self-organizing nonequilibrium steady-state enshrouded by a statistical boundary (called a Markov blanket). In the following section, we consider non-realist accounts of neural representation: a deflationary account, according to which the contents of neural representations are mathematical, and a fictionalist account, according to which representations are scientifically useful fictions. In the fourth section, we propose to combine aspects of both these accounts, yielding a nuanced realist account that defines semantic contents of representations formally—what one might call a deflationary, organism-centered fictionalist interpretation of neural representations. We argue that even pursuing a minimalist, deflationary account of the content of neural representations under the free-energy principles licenses an appeal to a robust kind of semantic content, the kind at stake in the ‘aboutness’ or intentionality of cognitive systems. The ensuing position coheres broadly with, but rests on distinct assumptions from, the realist one.
1.1. The Faces of Representationalism: Realism and Non-Realism
In the philosophy of mind, there are, roughly speaking, two main ways to think about the ontological status of the neural representation construct, which have implications for the available epistemological positions. One is
realism about neural representations. This view combines two positions: ontologically, that neural representations really exist (typically, that they are physically instantiated in the brain); epistemologically, that they are scientifically useful postulates as well [
6,
13].
Non-realist positions also are available, which do not share all of these assumptions. Non-realists are either agnostic about the reality of neural representations or explicitly reject the assumption.
Anti-realism says that neural representations do not exist. Several varieties of non-realism are available, which have different epistemological implications.
Eliminationism is the anti-realist view that the construct of neural representation should be eliminated from scientific practice [
14,
15].
Instrumentalism or
fictionalism is the non-realist view that argues that neural representations are useful fictions: they are a scientifically useful way of describing the world [
11,
12,
16,
17,
18,
19].
To get clear on which of these positions is most appropriate, it is useful to review the different versions of the neural representation construct. The classical view of neural representations casts them as symbolic structures that are realized by brain states and that are manipulated by rule-governed processes. This follows from the computational theory of mind [
3], according to which cognition is the rule-governed manipulation of symbol-like, internal cognitive structures (i.e., neural representations). In these classical accounts, the content of a representation is determined either by appealing to an innate stock of concepts and mechanisms that ensure the accuracy and objectivity of what is represented [
3,
20]; by accounting for contents through the actions of a biological proper function [
21,
22,
23]; or by referring to the phenomenal content of our first-person experience of things in the world [
24,
25]. All such accounts have in common a construct of neural representation as an internal symbol (or type) that gets instantiated (or tokened) in the appropriate circumstances; what varies is how the appropriateness condition gets implemented. There are also non-representational versions of the computational theory of mind, which will not concern us here; see Miłkowski [
26]; Piccinini [
27].
Motivated by parallel distributed processing, connectionist models of neural representations disagree with proponents of the classical approach over the nature of the representational vehicle; but agree that the brain harnesses internal cognitive structures that act as the vehicles for content [
28,
29]. Rather than discrete symbolic structures, the connectionist argues that neural representations are distributed representations; that is, that they accomplish their function of representing states of affairs in terms of joint configurations of their activity.
Today, the most popular (and in our view, the most compelling) representationalist accounts are of the connectionist type. They cast neural representations as
structural representations. On this account, neural representations are able to represent their target domain (i.e., to encode semantic content about their target domain) because their neural vehicles encode
exploitable structural similarities shared with the target domain [
8,
30,
31,
32]. On this account, representations function much like maps: they recapitulate the high-order structural features of that domain, for example, its statistical properties; also see [
33]. More specifically, structural representations encode information in a format that the organism can exploit to guide its behavior, that can afford the detection of errors (i.e., that affords misrepresentation), and that can be used for ‘offline’ navigation [
8,
32,
34,
35,
36]. Structural representations operate iconically, via a process “in which the structure of internal representations in the brain come to replicate the structure of the generative process by which sensory input impinges upon it” ([
31], p. 1962).
1.2. Towards Anti-Realism: Deficiencies of the Realist View
Most research in the cognitive sciences and neurosciences tacitly operate on
realist assumptions about neural representation and design experiments that aim at explaining how our experience of, and action in, the world is mediated by structured bodies of knowledge that are encoded in the networks of the brain. The existence and explanatory value of neural representations is a basic premise of almost any psychology textbook. For instance, one can read in
The MIT Encyclopedia of the Cognitive Sciences: “Psychology is the science that investigates the
representation and
processing of information” ([
37], p. xl, emphasis added).
Although
realism about mental representation is the default mode of operation in most of the cognitive sciences and neurosciences, it is not the consensus position. The motivation for adopting an
anti-realist,
anti-representationalist approach comes from the observation that, despite enormous efforts and scientific investment, representations have yet to be naturalized [
6,
9,
38,
39,
40,
41,
42]. This is notably because extant attempts to articulate a theory of neural representation have so far failed to provide a naturalistic theory of semantic content that does not presuppose the very intentional relation and representational content that it seeks to explain, securing point (4) discussed above. Thus, scholars such as Ramsey [
43] call for caution about the use of the construct. They argue that often, the appeal to semantic content is a philosophical gloss that does not add any explanatory value:
“The roles provided by commonsense psychology are those that distinguish different types of mental representations. What we need and what is not provided by commonsense psychology is, more generally,
the sort of physical condition that makes something a representational state, period. In functional terms, we would like to know what different types of representations perhaps have in common, qua representation.
Neither commonsense psychology nor computationalism tells us much about the sort causal/physical conditions that bestow upon brain states the functional role of representing (at least not directly).” ([
43], p. 6, emphasis added)
In the literature, it is quite common to see the selective responsiveness of neural tissue to a given stimulus described as the representation or encoding of stimuli. This conceptualization is adopted in the study of perception, particularly to highlight cellular specialization in detecting certain features of the perceptual object [
44,
45,
46,
47,
48]; in the study of memory [
49,
50] and motor activity [
51,
52,
53] as well. However, as Ramsey [
6] notes, response-selectivity by itself cannot make a physical state a representation. Many physical states have response-selectivity but are not representations. For instance, the states of one’s skin vary with the weather (e.g., it gets darker the more it is exposed to the sun), but we would not (intuitively) count one’s skin as a representation of the sun or weather.
We find approaches in the biological sciences and in the neuroscience less committed to the view of cognition as a representational process taking place within the boundaries of the brain [
54,
55,
56,
57,
58]. These views include perceptual and motor control theory [
59,
60]; robotics [
61]; cybernetics [
62,
63,
64,
65]; and, arguably, the free-energy principle and active inference [
56,
57,
66]. These accounts, which often hail from embodied and enactive approaches in cognitive science [
14,
67,
68,
69,
70], converge on the idea that the primary aim of cognition is not internally reconstructing proxies for the structure of a hidden world, but rather to adapt to and act in an environment.
In this setting, Sprevak [
17] suggests that there appear to be two options. We can take a hard-headed
realist assumption that the naturalization of neural representations will eventually succeed. (Even though no account has succeeded so far.) Alternatively,
non-realism (eliminativism and fictionalism) downplays the value of representation talk in cognitive science. Non-realists observe it is not possible to define the content of a neuronal representation without conflating it already with the cognitive process: We borrow our semantics (and thereby, the content of the representation) from our scientific practice. More precisely, if the supposed representational content cannot be determined without appealing to what we know about the cognitive activity itself, then it is the cognitive activity that has explanatory power. Conversely, if the explanation in terms of the cognitive activity suffices without appealing to an experimenter-imposed semantics, then there is no reason to postulate representational content. This may motivate restraining the use of neural representation use to an “informal gloss” [
11,
16].
The cost of eliminating the construct of neural representation altogether is that it requires a painful revision of the mainstream representational paradigm in cognitive science. Indeed, we typically appeal to neural representations to explain goal-directed, probabilistic inference, and decision-making. To abandon this posit leaves us with the obligation to abandon some of the most powerful explanatory tools that we can use. Is this legitimate?
1.3. Representations Under the Free-Energy Principle?
The object of this paper is more precisely the status of neural representations in Bayesian neuroscience, known as the free-energy principle and active inference. In Bayesian neuroscience, the brain is cast as a statistical organ or inference engine that minimizes its uncertainty about the state of the world. In this family of theories, the brain is depicted as doing its predictive work by drawing on probabilistic knowledge about its environment to explain the likely causes of the sensory signals with which it is bombarded [
36,
64,
71,
72]—and to act in ways that bring about its preferred or expected sensory states [
73,
74].
Some Bayesian approaches based on predictive coding algorithms for describing canonical microcircuits in the cortex have strong representationalist commitments [
8,
31,
32,
34,
35,
36,
73,
75]. These accounts argue that the Bayesian brain entails the postulation of structural representations. Thus, neural representations are taken to be internal, map-like structures that are instantiated in the networks of the brain and that encode exploitable information.
Several recent papers have discussed whether a realist interpretation of neural representations is warranted under the free-energy principle [
8,
32,
34,
35,
56]. Besides a few notable exceptions [
76], few papers have sought to evaluate the variety of non-realist arguments in light of the free-energy principle. In this paper, we will argue that taking seriously two forms of the non-realist position (the deflationary and fictionalist views) leads back to a nuanced form of realism that is apt to provide a naturalistic basis for the study of intentionality.
Why concern ourselves with the ontological and epistemological status of representations? From our point of view, one main reason to do so is that one’s position with regards to the status of representations has implications for research in computational neuroscience; and to determine which structures play the role of representation, and how they carry their semantic contents, is crucial for the practice of neuroscience. One salient example is that of motor representations and motor commands in the human brain [
77,
78]. Representationalist frameworks in computational neuroscience assume that there exist structures in the brain that represent motor tasks. Optimal control theory is one of the more popular frameworks for modelling that borrows such assumptions. This approach is underwritten by strong assumptions about the nature of the models and signals that the brain processes in motor control. The hypothesis in optimal control is that motor representations are brain structures that encode explicit instructions to perform a task and that are specified in terms of intrinsic coordinates (i.e., in terms of the contraction and stretching of muscle fibres). However, optimal control theoretic constructs have been criticized and empirical evidence is lacking for explicit instruction-like motor commands in the brain. For discussion, see Hipolito et al. [
77]. Our framework offers an alternative characterization of representational capacities, which are not premised on instruction-like motor commands. Instead, we characterize the representational capacity as underwritten by an ontology of fictive states, and on a process of active inference that realizes preferences about sensory data. Our deflationary perspective on representational capacities does away with the problematic representational posits of optimal control, while also shedding light on how semantic contents are acquired through histories of active inference.
2. The Free-Energy Principle and Active Inference: From Information Geometry to the Physics of Phenotypes
2.1. State Spaces, Nonequilibrium Dynamics, and Bears (Oh My)
Our paper will focus on a prominent Bayesian theory of action and cognition, the variational free-energy principle, and its corollary, active inference [
66,
79]. The free-energy principle starts with the observation that biological systems like living creatures have a phenotype. Living organisms maintain their phenotypic integrity and resist the tendency towards thermodynamic equilibrium with their ambient surroundings—that is usually dictated by the fluctuation theorems that generalize the second law of thermodynamics [
80,
81]. Living creatures do so by upper bounding the entropy (the dispersion or spread) of their constituent states. To get a better handle on this, in this section, we introduce two formal notions: the state space and nonequilibrium steady states. A brief technical treatment of what follows can be found in the
Appendix A and Glossary of terms and expressions.
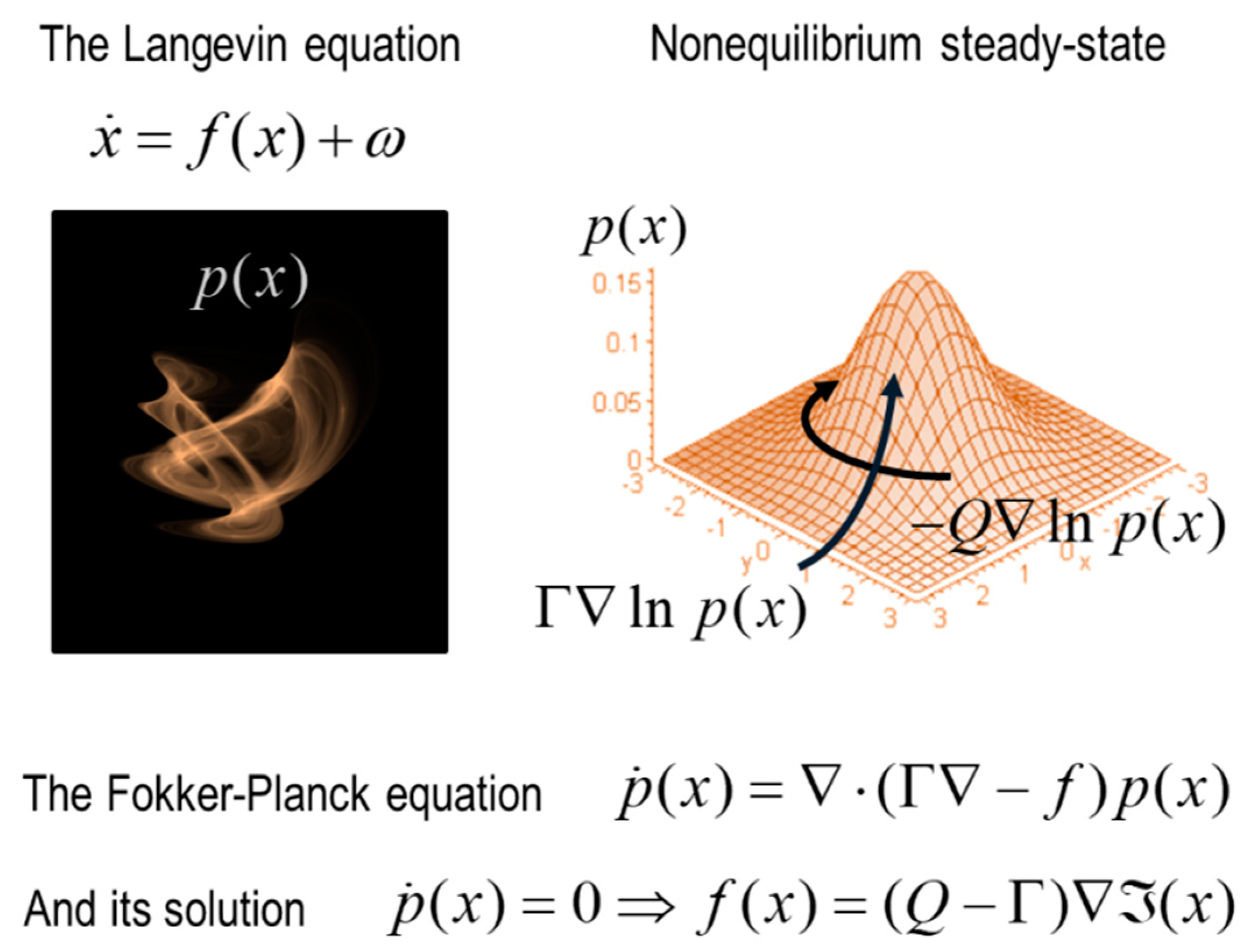
In physics, equilibrium and nonequilibrium are distinguished by the end-state towards which a dynamics evolves. Equilibrium dynamics resolve when all the energy gradients have been consumed; at which the point system is at thermodynamic equilibrium with its environment. For living creatures, thermodynamic equilibrium is death. Living creatures are open systems that remain far from equilibrium. How can we model this using formal resources?
The state or phase space formalism comes from dynamical systems theory and allows us to get a formal grip on the predicament of living systems. A state or phase space is an abstract space that allows us to model the time evolution of a system in terms of all the possible states in which it can find itself. To construct a state space, we identify all the relevant quantities that can change in the system (i.e., all the relevant variables) and then plot each variable on a dimension in an abstract space. This space is called a state space. Every dimension of this space corresponds to a variable in the system; such that a point in this space corresponds to a complete instantaneous specification of the system, since we assign a value to every variable of the system; i.e., we assign a position to the system along every dimension. A trajectory in this space, in turn, corresponds to the flow of the states of the system over time.
The state space formalism allows us to describe the time evolution of a system implicitly by depicting trajectories through state space. This turns out to be crucial. If we draw a probability density over all the states that a system can find itself, those combinations of states with the highest probability, to which the system returns periodically, are known as a pullback attractor [
66,
82]. We can associate the states (i.e., the regions of this space) in which a creature finds itself most of the time with its
phenotypic states.
The probability density that describes the system at its nonequilibrium steady state (i.e., its phenotypic states) is aptly called the nonequilibrium steady state density. Such a probabilistic description of the system’s dynamics can be interpreted in two mutually consistent and complementary ways. First, the system can be described in terms of the flow of the system’s states—that are subject to random fluctuations—in which case, we can formulate the flow in terms of a path integral formulation, as a path of least action. Equivalently, we can describe the nonequilibrium steady-state in terms of the probability of finding the system in some state when sampling at any random time. These two descriptions are linked mathematically by the fact that at nonequilibrium steady-state, the flow is the solution to something called the Fokker Planck equation that describes the density dynamics. This dual interpretation will play a crucial role later.
2.2. Markov Blankets and the Dynamics of Living Systems
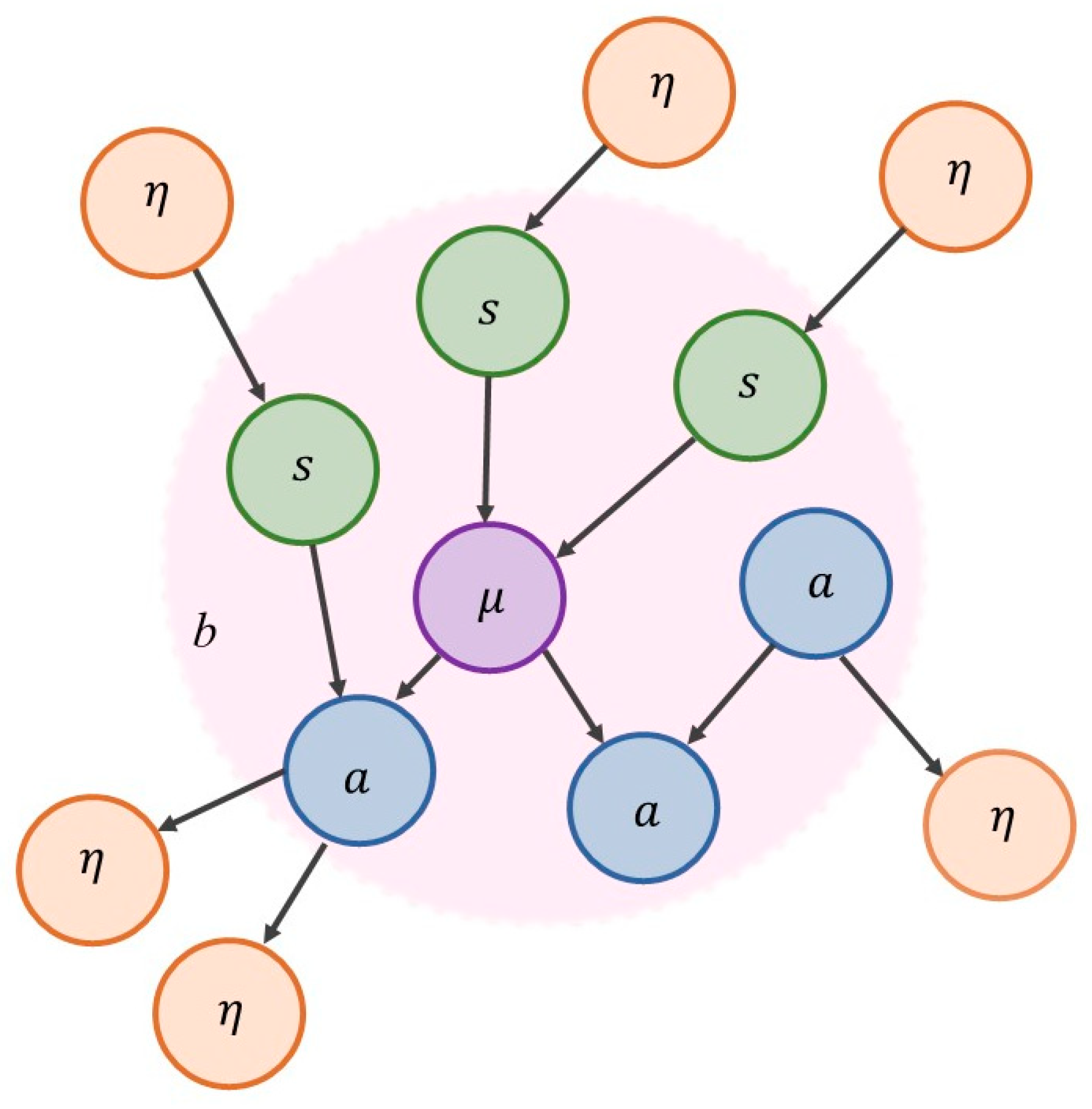
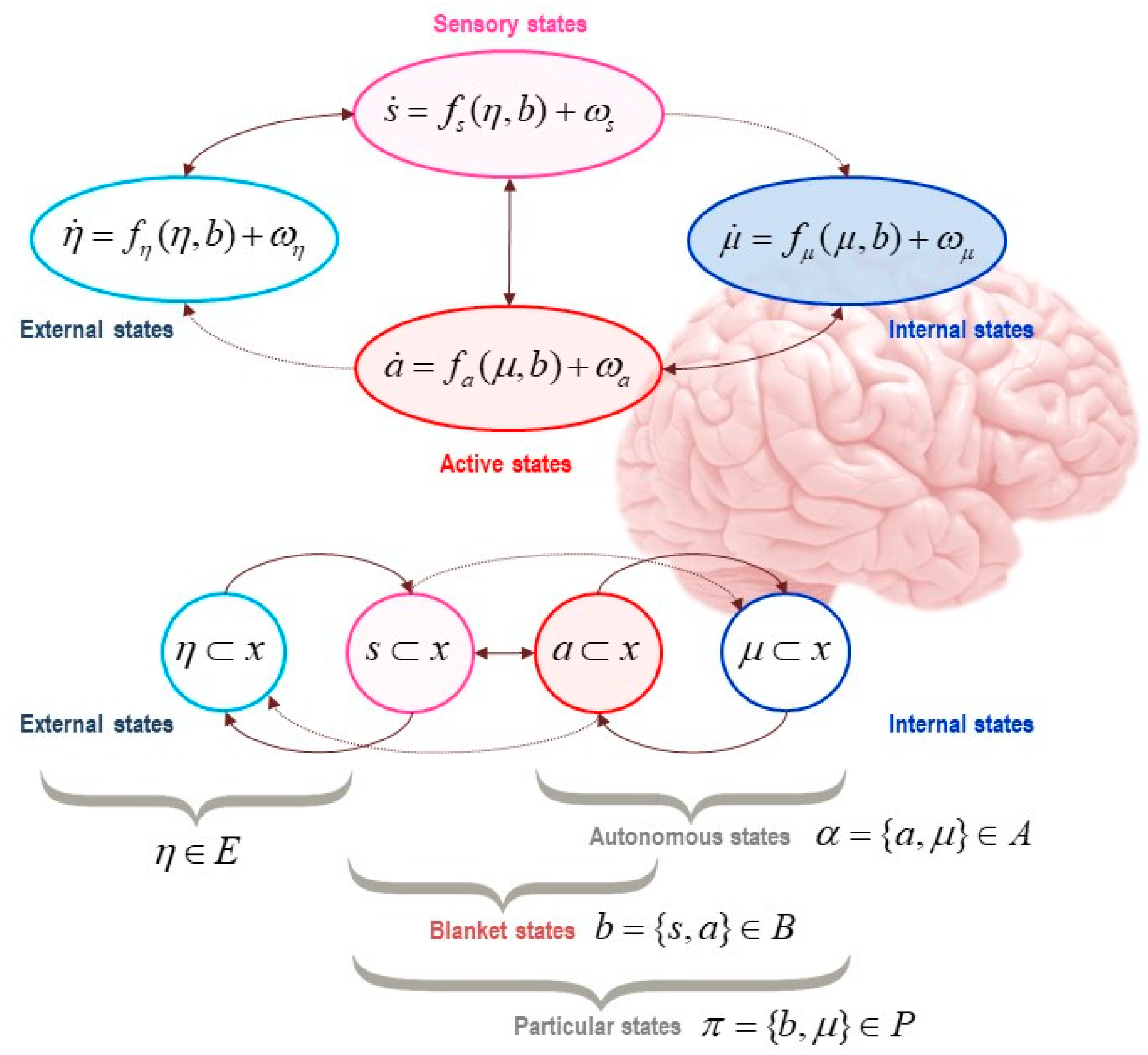
The free-energy principle builds on the dynamic systems theoretic approach, which concerns the time evolution of systems, but now augmented with considerations about the statistical properties and the measurability of those systems. The free-energy principle allows us to describe the flow of a system’s states in one of two mathematically equivalent ways, statistical and dynamical—an equivalence that is warranted by virtue of the conditional independence entailed by the presence of a statistical boundary (called a Markov blanket) and the existence of the system in a regime of steady (phenotypic) states far from equilibrium.
For a system to exist (separately from the rest of the universe), it must be endowed with a degree of independence from its embedding environment. A Markov blanket is a set of variables or states that we use to stipulatively individuate a system in terms of what is part of it (internal states, denoted
μ), and what is not (external states, denoted
η). The Markov blanket itself is defined as those states that mediate interactions between the system and its embedding environment (active and sensory states, denoted
a and
s). The Markov blanket is defined by the absence of certain connections: internal states do not cause sensory states, and external states do not cause active states. See
Figure 1 and
Figure 2.
The presence of a Markov blanket induces a conditional independence between internal and external variables. The key word here is ‘conditional’: internal and external are not really independent of one another—they just appear to be so if we discount their dependencies via the blanket states.
2.3. Information Geometries and the Physics of Sentient Systems
The core intuition behind the free-energy principle is that if a system is endowed with a phenotype (i.e., a nonequilibrium steady state density) and has a Markov blanket, then there are two ways of describing the flow of a system’s states that turn out to be equivalent: one rooted in the state space description of the system that is formulated in terms of the flow or dynamics of the internal states; the other rooted in a statistical interpretation of the same flow. The presence of a Markov blanket in such a system ensures that both descriptions are coincidentally true or conjugate to each other [
66,
84].
This description of the system—in terms of movement in internal phase space—is the system’s ‘intrinsic’ information geometry, which is closely related to measure theory and statistical thermodynamics. Measure theory is a field of mathematics about how we can systematically assign a number to subsets of a given set, where a measure or metric consists precisely in such an assignment; for example, a probability measure assigns a probability value systematically to elements of a subset. We can think of measures as capturing something about size, or distances in an abstract space. Generally, an arbitrary set of points that compose a state space does not have a measure, and so no associated notion of distance can be defined for that space. However, one can equip a space with a metric, usually in the form of a matrix that describes how ‘far’ one has moved as a function of small changes in position.
In Euclidean geometry, this metric is just an identity matrix, i.e., if I move 100 m along some particular direction, then I will have moved a total of 100 m. This is not true, e.g., of movement on a sphere: the planet Earth, for instance, has a circumference of approximately 40,000 km; if I move (approximately) 40,000 km in the same direction on Earth, I will not have moved at all in total relative to the Earth’s surface (since I will be back where I started).
This notion of metric plays a special role when dealing with sufficient statistics and statistical manifolds. Sufficient statistics are the minimal set of numbers that are needed to reconstruct or parameterize a probability distribution (which contains an infinite number of points). For normal or Gaussian distributions, these numbers are the mean and variance. A statistical manifold is a space in which the coordinates are the sufficient statistics of a family of probabilities densities. For instance, the sufficient statistics of a Gaussian distribution are its mean and variance, giving a two-dimensional statistical manifold or state space. Given any position on this manifold, it is possible to reconstruct the probability density (which assigns a probability to an infinite number of points). Trajectories on a statistical manifold correspond to changes in the shape (i.e., the value of the parameters) of the associated probability density.
Information geometry, in turn, is the field of mathematics dealing with measures or metrics on a statistical manifold (information manifolds, typically endowed with a Fisher or Riemannian information metric). In other words, information geometry allows us to define a metric for probability distributions (or probability densities for continuous variables); that is, we can talk about distances between probability densities. Furthermore, if we associate probability densities with probabilistic beliefs, we now have a naturalized way of talking about the distance between beliefs.
With all this in place, we can appreciate what the free-energy principle brings to the table. Starting from our state space description of the system, we can define a metric that allows us to speak about distances between probabilistic configurations of a system’s internal states. Such a geometry is ‘intrinsic’ because it describes the structure of a system’s possible configurations in a manner that only refers to internal states themselves (rather than the external states of the environment). The free-energy principle says that so long as the system at hand is equipped with a nonequilibrium steady state density and a Markov blanket is in play, then an additional—and mathematically conjugate—information geometry can be defined [
66,
84]. These two information geometries can take the form of the following (Fisher information) metrics, where the sufficient statistics correspond to expected thermodynamic states and internal states, respectively:
This licenses a spectacular observation: namely, that internal states can be interpreted in terms of their extrinsic geometry, i.e., as parameterising a probability density over external states. This simple fact is a natural consequence of the conditional independences that define the Markov blanket. Put simply, for every blanket state (i.e., joint sensory and active state) there is a conditional probability density over internal and external states. Crucially, these are conditionally independent, by definition, given the blanket state in question. This means that for every expected internal state, given the blanket state, there must be a conditional probability density over external states. In turn, this means that the expected internal state is a statistical manifold—equipped with an extrinsic information geometry. This extrinsic information geometry describes the distance among probabilistic beliefs about external states, which are parameterized by the expected internal states. In other words, expected internal states constitute the sufficient statistics of beliefs about external states.
2.4. Phenotypes: A Tale of Two Densities
Essentially, the free-energy principle is a story about two probability densities [
56]. The first is the nonequilibrium steady state density itself, which harnesses the statistical structure of the phenotype. The second,
variational density, is parameterized or embodied by the internal states of the system. We have seen that internal states constitute the sufficient statistics of probabilistic beliefs about external states. Another way of looking at this is to say that internal states encode a probability distribution—the variational density—over the states of the external world that are generating sensory (and active) states. Technically, that a system will evolve towards—and maintain—its nonequilibrium steady state means that it minimizes the discrepancy between the variational density that it embodies and the probability density over external states, given blanket states [
66,
84]. This discrepancy is the variational free-energy, and the steady state flow that underwrites nonequilibrium steady-state becomes a gradient flow on variational free-energy. In other words, it will look as if internal states are trying to optimize their posterior beliefs about external states. When cast in terms of movement in the extrinsic geometry, one can interpret existential behavior in terms of Bayesian belief updating. This all follows because the expected internal states parameterize a conditional or Bayesian belief about external states.
Anthropomorphically, the system does not ‘know’ in what state it is in, but it will look as if it is inferring the state of its external milieu ‘out there’, based on prior beliefs and its current sensory states. The state of the external world is thus never directly perceived, it is instead something that the organism infers and brings about actively through interactions with the world. In this sense, the implicit inference is enactive, in the pragmatist sense of being for action [
56,
57,
85]. When the organism interacts with the world, it perturbs external states and consequent sensory states. These sensory impressions couple back to internal states that attune to the world around them. Put otherwise, the organism engages in a form of Bayesian inference (i.e., active inference), with respect to the state of its ecological environment, based on the situated interaction.
Why inference? The free-energy principle says that living things exist in virtue of gradient flows on an information theoretic quantity called
surprisal. This is the solution to the Fokker Planck equation that furnishes nonequilibrium steady-state. Crucially, the dynamics of internal (and active) states at nonequilibrium steady-state can be cast equivalently as flowing down surprisal or free-energy gradients. Free-energy scores the atypicality of sensory (and active) states, given a (generative) model of how those data was generated [
66,
86,
87].
The equivalence between surprisal and free-energy rests upon the fact that the (expected) internal states parameterize beliefs about external states. When this variational density corresponds to the density over external states, conditioned on blanket states, surprisal and free-energy are the same. Crucially, free-energy is a functional (i.e., function of the function) of the variational density and an implicit generative model. The generative model is just the nonequilibrium steady-state density over external and blanket states. And can be regarded as a description of how external states generate blanket states.
In what follows, we will treat the generative model as an implicit attribute of any nonequilibrium steady-state that possesses a Markov blanket. The generative model is implicit because the only thing needed to describe self-organization and belief updating are the free-energy gradients. This means that the free-energy and its generative model are not evaluated or realised explicitly. This is sometimes referred to as
entailing a generative model. [
56,
88]. In short, the free-energy is a functional of two densities, the generative model and the variational density encoded by internal states.
For people familiar with information theory, surprisal is also known as self-information, where the long-term average of self-information is entropy. This means that nonequilibrium steady-state flows counter the entropy production due to random fluctuations. In turn, this means that the kind of inference implicit in the flow of autonomous states (namely, internal and active states) underwrites the existential imperative to maintain a steady-state far from equilibrium.
What the free-energy principle says then, is that so long as a Markov blanket is in play at nonequilibrium steady state density, gradient flows on surprisal (a function of states) are equivalent to gradient flows on free-energy (a function of sufficient statistics), where the sufficient statistics parameterize probability distributions over—or beliefs about—external states. This echoes the reasoning above about the conjugate information geometries, in terms of dynamics and in terms of statistics. Gradient flow here just means that the autonomous states (i.e., internal and active states; see
Figure 2) flow down variational free-energy gradients. And this, in turn, is just another way of talking about
action and perception. This scheme, naturalizing action and perception as the gradient flow of active and internal states (respectively) on variational free-energy, is known as
active inference, the corollary process theory of the free-energy principle.
The main take-home message is that the free-energy principle casts the phenotype in two complementary ways: as a flow of states at nonequilibrium steady state (described via an intrinsic information geometry) and a flow that entails belief updating (described via an extrinsic information geometry). In virtue of the active states, the apparent role of internal states is not merely to infer the causes of sensory data, but to generate appropriate patterns of interaction. This means that internal states could parameterize beliefs about the consequences of action, and facilitate the consequences of action for beliefs [
66]. Generative models are thus not designed to merely do the interpretative work of determining the true state of the world, they cover the consequences of acting on worldly states.
2.5. Living Models: A Mechanistic View on Goal-Directed, Probabilistic Inference and Decision-Making Under the Free-Energy Principle
With this setup in place, we are in a position to appreciate how generative models allow organisms to engage in goal-directed, probabilistic inference, and decision-making under the free-energy principle. The free-energy principle is often cast as the claim that living systems
just are generative models of their environment [
56,
76,
89,
90,
91,
92]. We can now make sense of this seemingly enigmatic claim. The free-energy principle says that organisms leverage the statistical structure of their acting bodies to remain in their phenotypic states, where that typical structure is interpreted probabilistically, as a joint distribution over all systemic states. Decision-making about what to do next is then based on a probabilistic inference about “what I must be doing, on the assumption that I am a free-energy minimizing creature.”
Hitherto, we have considered inference as an emergent property of self-organization to nonequilibrium steady-state. The ensuing Bayesian mechanics is licensed by an equivalence between surprisal and variational free-energy. In what follows, we can make a further move and describe creatures or particles in terms of the generative model that defines free-energy. Once we have the free-energy, we know the gradients. Once we have the gradients, we know the gradient flow. Once we have the gradient flow, we can naturalize any embodied exchange with the environment.
A typical generative model is depicted in
Figure 3. Here, hidden states (
η) correspond to the external states that are hidden from the internal states behind the Markov blanket. The crucial thing to appreciate is that beliefs about hidden states correspond to a
hypothesis—that the organism embodies—about the causes of its sensations. These hidden states arguably have all the properties that would make them the content of structural representations [
8,
31,
32,
34,
35,
56,
93]. The hidden states of the generative model are parameterized by the internal states of the system (e.g., the brain) and encode exploitable information about external states that guide adaptive behavior. We will return to this point later.
To summarize, the presence of a Markov blanket at nonequilibrium steady state allows us to associate a living particle or creature with its internal and blanket states. The flow of the internal states acquires a dual aspect, described by conjugate intrinsic and extrinsic information geometries. These geometries inherit naturally from the Markovian structure of the partition. The intrinsic information geometry describes the thermodynamic behavior of internal states (e.g., neuronal dynamics). However, the internal states also are equipped with an extrinsic geometry, which pertains to the probability distributions over, or beliefs about, external states that are parameterized by internal states. A gradient flow of active and internal states on free-energy (action and perception), effectively implements active inference; namely, inferring external states and planning what to do next. This completes our technical review of the free-energy principle.
4. A Variational Semantics: From Generative Models to Deflated Semantic Content
4.1. A Deflationary Account of Content Under the Free-Energy Principle
In this last section, we combine elements of the deflationary and the instrumentalist accounts of neural representations to propose a kind of organism-centered fictionalism or instrumentalism. We expand upon the ecological component of deflated mathematical content, which we argue leads to a naturalistic theory of intentionality: a formal theory of semantic content based on the free-energy principle. The key to formulating a robust mathematical theory of semantic content, one capable of naturalizing intentionality, is to notice that the free-energy principle essentially tells a story about the mutual attunement between a system and its environment.
In our view, the deflationary view of representational contents downplays the role of the fifth, ecological component of the computational theory proper. We argue that formulating the ecological component using the resources of the free-energy principle allows us to salvage the intentionality of semantic content—and thereby recover a robust conception of content tied to the domain to which it is intentionally related (or about)—without appealing to the artificial intentional gloss of cognitive content. The resulting view is of a semantics that emerges naturally from the fact that the system we are considering is equipped with a dual information geometry of states and beliefs.
The formalism that underwrites the free-energy principle licenses a crucial observation: namely, that the mathematical structures and processes in play are defined over a state space and, implicitly, over an associated belief space or statistical manifold [
93]. The mathematical framework of the state space formalism means that the system’s dynamics are defined over states of the system; and that as a consequence of the associated extrinsic information geometry, we can always associate a
semantics to this intrinsic description.
This semantics comes from the ‘beliefs’ built into the extrinsic information geometry. The term ‘belief’ is used in the sense of ‘belief propagation’ and ‘Bayesian belief updating’, which are just ways of talking about probability distributions or densities. ‘Beliefs’ in Bayesian terms are posteriors and priors, corresponding to the probability distributions (a world of possible states) that are shaped by physically realized states (i.e., the internal states that parameterize distributions over external states). In general, although we use the term ‘beliefs’ to describe the probability densities defined over external states, it is generally recognized that these densities are not themselves the same as propositional beliefs. In brief, propositional beliefs have truth conditions; that is, they are the kind of thing that can be true or false [
15]. The probability densities at play here are not of this kind; they represent the manner in which variables covary. This does not imply truth-conditionality, which means that they are non-propositional.
It is often noted that one does not obtain semantic content from mere systematic covariation [
6,
15,
108]. However, this argument can be defeated by noting that, under the free-energy principle, for any living system, there is an implicit semantics at play that is baked into the system’s dynamics. Importantly, that is just saying that the system’s internal dynamics have a probabilistic aspect (and extrinsic information geometry) that connects it to an embedding system. Via the Markovian partition, we can always associate this trajectory of states on internal (statistical) manifold to a semantics—a formal semantics that falls out of the systemic dynamics and that can be characterized purely mathematically.
Thus, we obtain a bona fide formal semantics from interactions between the system and its context, as well as the histories of environmental interactions between an organism and its niche. What follows from our account is a somewhat more ‘realistic’ deflationary position, a weakly deflationary position according to which the content of a representation is indeed the mathematical function that it realizes, but where this computational theory proper entails an implicit semantics.
4.2. From a Computational Theory Proper to a Formal Semantics
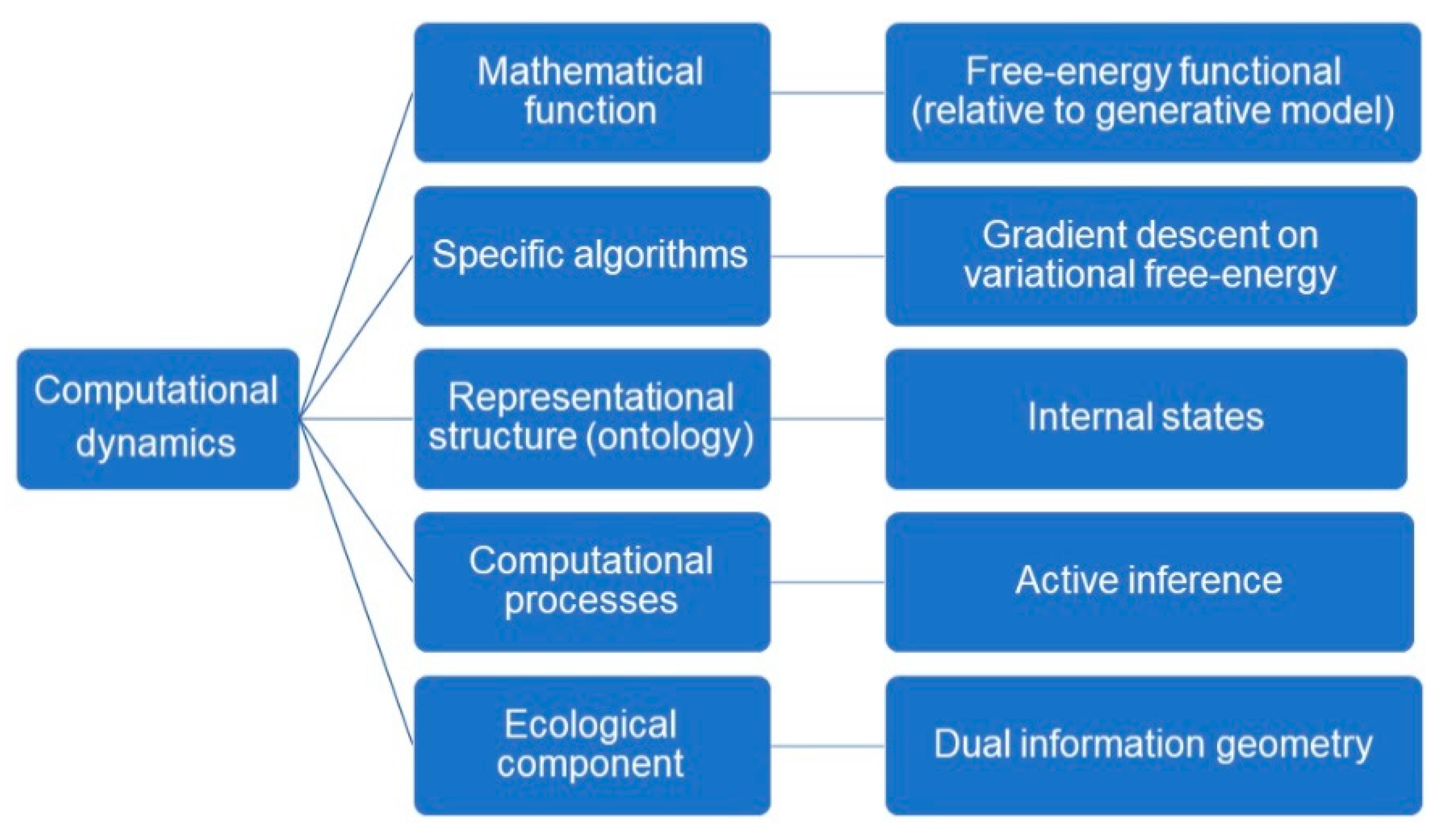
Let us now take stock. We have retained the general description of representational content from the deflationary account. We now use this deflationary model to specify the computational theory proper that leads to a formal semantics via the free-energy principle.
The computational theory proper has five components, which we can map to elements of the free-energy formulation. (1) Under the free-energy principle, the mathematical function that is realized by the (gradient flows of the) cognitive system is a free-energy functional that measures the divergence between the posterior and variational densities. (2) The specific algorithms that the system uses to compute this function is gradient descent on variational free-energy. (3) Echoing the literature on structural representations, the representational structures that are maintained and updated by the mechanism are the internal states of the system. (4) The computational processes that are defined over these representational structures, and which update and maintain them, are implemented as active inference. (5) Finally, the ecological component is provided by the dual information geometry.
Figure 4 and
Figure 5 reformulates Egan’s [
11] account deflationary account of content in light of the free-energy principle. We amend the deflationary account to highlight that it provides us with a fully naturalistic, mathematical account of the origin of semantic content, in terms of a calculus of beliefs and intentions that is the counterpart of the intrinsic description of the flow of internal states. Note that the external states that figure in the generative model implicitly define the
ecological component—and this, purely mathematically. This overcomes the problem of
naturalizing intentionality by appealing purely to well understood mathematical and physical processes and properties.
4.3. Phenotypic Representations? Ontologies?
Our last move is to leverage the fictionalist account to nuance the claim that the account just rehearsed vindicates the neural representations construct. This nuance is on two counts: with regards to the terms ‘neural’ and ‘representations’.
First, the term ‘neural’ should be replaced by ‘phenotypic’, to reflect the expanded realization base of the vehicles of deflated, mathematical content under the free-energy principle. The spirit of deflated neural representationalism is vindicated by the free-energy principle. We can indeed assign mathematical content to structures that are internal to an agent, which come to encode or carry semantic content when the right ecological conditions are in play (thanks to the dual aspect information geometries of living systems). However, the internal states that play this role are very far from classical representations in the symbolic or connectionist traditions. They essentially comprise all internal states of a system and so are not strictly neural. This has the consequence that the neural representationalist intuition is vindicated by its traditional adversary, the embodied-enactive approach to cognition: if there is anything like structural representations under the free-energy principle, they correspond to the system’s bodily states and are harnessed, maintained, and updated through histories of adaptive action.
Second, with respect to the term ‘representation’, we note that under the free-energy principle, the deflated representational structures may best be understood as the ontology that system brings to bear in understanding its environment; that is, the set of hypotheses or categories that it uses to parse the flow of its sensory states.
Heuristically, we say that the free-energy principle licenses the claim that the system believes that this or that environmental factor is causing its sensory impressions. In light of the discussion above, it appears more accurate to say that, when it is in its usual ecologically valid operating conditions, a system equipped with such a partition that exists at nonequilibrium steady state will act in a way that looks as if it has an intentional relation with some features of its environment. We now know what this “as if” character amounts to: it refers to the duality of information geometries and thereby the duality of possible descriptions (in terms of a flow towards nonequilibrium steady state and in terms of belief updating under a generative model).
The free-energy principle descends from a modeling strategy called generative modeling. In this scheme, we write down alternative probabilistic models of the process that caused our data, and we score the probability of each model as a function of how well it explains the variance in our data. This score is the variational free-energy. Crucially, as discussed in the previous section, these models are
hypotheses about the structure of the process that caused our data. The hidden or latent (c.f., external) states in these models are essentially guesses about the causes of sensory data. Crucially, they need not reflect the existence of anything in reality [
66,
84,
109]. This is a subtle but important point. The external states exist only relative to the generative model and accompanying phenotype, and only play a role so long as they subtend the generation of adaptive, contextually appropriate behavior.
What is being described by the formalism that underwrites the free-energy principle, then, is less a story about how an internal reconstruction of the external world is constructed in the brain, as traditional (symbolic and connectionist) accounts of neural representations would have it. What is at stake is more like the
ontology with which a system is equipped [
110]. An ontology, in this sense, is the set of semantic structures (a large part of which are learned through experience from immersion in specific contexts) that a living creature uses, implicitly or explicitly, to parse and order the flow of its sensory states [
111,
112,
113,
114].
We are effectively combining the deflationary account and the fictionalist or instrumentalist account to provide an interpretation that might be called organism-centered functionalism. The organism’s phenotype—its moving and acting body—is a nonequilibrium steady state density that can be interpreted as a manifold towards which the flow of system’s states settles on average and over time; and also as a joint probability distribution over all the variables of the system. The organism’s behavior is driven by these density dynamics: by the tendency to settle towards its nonequilibrium steady state density, which is implemented as a gradient descent on variational free-energy (a.k.a. active inference).
This is just another way of saying that the actions selected by the organism are driven by the statistical structure of its phenotype and interactions with the environment. The organism leverages its own statistical structure in driving its action selection. This dovetails with embodied-enactive approaches to cognition and effectively constitutes a new take on morphological, developmental, and evolutionary computation [
115,
116] and knowledge-driven skillful action [
77].
While our aim here was to explore the consequences of non-realist approaches to cognition, we note that the pragmatist interpretation that we propose may be compatible with certain realist, structural representationalist accounts, where content is determined by functional isomorphism, thereby illuminating the role of structural representations using a form of functional role semantics [
8,
32,
81]. These accounts are explicitly developed in a direction that does not require the represented system to actually exist in reality, which coheres with the account of ontologies just presented. The free-energy principle warrants the claim that there are phenotypic states that carry semantic content; here, we articulated a computational theory proper for such mathematical semantics. What we call these states, at the end of the day, may be a matter of preference.
5. Conclusions
In this paper, we aimed to assess whether the construct of neural representations plays an explanatory role under the variational free-energy principle, and to determine which philosophical position about the ontological and epistemological status of the representations construct is most appropriate for that theory. We examined non-realist approaches, rather than the more commonly discussed realist ones. We started by a deflationary account of mental representation, according to which the explanatorily relevant contents of neural representations are mathematical, and a fictionalist account, according to which representations are scientifically useful fictions. We hope to have shown that under the free-energy principles, even quite minimalist, deflationary accounts of the kind of content carried by neural representations warrant an appeal to a semantic content, which echoes (while being distinct from) the realist position. We hope to have shown that, by drawing on a modified fictionalist account, the formal semantics derived from the free-energy principle can provide us with an explanation of the aboutness or intentionality of living systems.
Much hangs philosophically on what it means to represent some target domain, specifically in terms of the relation between mental states and the physical states that realize them. The relations between the free-energy principle and classical positions in the philosophy of mind (e.g., physicalist monism, dual-aspect monism, and Cartesian dualism) have been explored at length elsewhere [
84]. We will only comment on which of these seems to cohere most with our account. Briefly, of the philosophical perspectives on the relation between mental and physical states, ours here is most consonant with
functionalism and the concept of
multiple realization that it entails. Functionalism is the view that the features that characterized mental states are not the intrinsic features of that state, but rather the functional (e.g., input-output) relations between that state and other states of the system [
117,
118]. Multiple realization is the view that the same (mental) macrostate state can be realized variously by different configurations of (physical) microstates, so long as they implement the appropriate functional (e.g., computational) relations [
118]. The proximity between our view and functionalism is based on the technical detail of how the semantic content is realized under the free-energy principle, via the assignment of fictive external states to internal states implied by the dual information geometry of the free-energy principle. As discussed above, the free-energy principle licences the claim that, for every blanket state, we can identify an
average of internal states that we can associate with the parameters of a probability density over (fictive) external states. The crucial thing to note is that it is the
average of internal states that can be so associated. This means that the probability density over external states can be realized by an equivalence class of internal states, which end of parameterizing the same belief.
An outstanding issue is whether the framework on offer is able to account for the issue of
misrepresentation. Any candidate representational structure must at least in principle be able to misrepresent the state of affairs that it represents. This has long been a sticking point in the discussion on representation [
119]. In a nutshell, because misrepresentation is possible (e.g., recognizing one object as another that it is not), an account of representation needs to allow for misrepresentation while also specifying what makes the representation about one object versus another—if it can be induced by both objects [
3,
120]. It has been pointed out, in our view correctly, that the information theoretic measures used under the free-energy principle are measures of mere covariance, which are insufficient to account for misrepresentation [
108]. We think that this picture is incomplete. A future direction for more fully addressing this issue begins by noting that variational free-energy scores the degree to which sensory data conforms to hypotheses about what caused it. Variational free-energy is a not just any measure of information, but instead measures the discrepancy between the current sensory data and the sensory data
expected under some ontology or hypothesis, which lends it an irreducibly semantic aspect. High free-energy indicates that the hypothesis does not ‘explain’ the data, or that some other hypothesis would fare better. Thus, the model of semantics on offer might be able to account for misrepresentation and the search for alternative hypotheses.
Our primary concern in this paper was to show that phenotypic states can come to acquire semantic contents of a deflationary (mathematical) sort via active inference. Of note is that this semantic content is not equivalent to the kind of
propositional content that is at play in language use, nor does our account explain the manners in which human agents use language and narrative to fashion and remake themselves as agents [
121,
122]. While this issue is, at least arguably, beyond the scope of this paper, we generally agree with the claim that the move from semantic to propositional content requires that agents engage in specific kinds of content-involving practices. These are practices like truth-telling [
9,
15] and story-telling [
123], which build greatly upon the basic representational or semantic capacity of agents by enabling more elaborate, storied, forms of self-access that would not be possible without language.
Where does this leave us with regard to our initial questions? The upshot of our discussion is that, under the free-energy principle, there indeed are structures internal to an organism that act as the bearers of semantic content. These structures can be specified mathematically in terms of computational theory proper, as held by the deflationary account. However, by virtue of the dual information geometries in play under the free-energy principle, this purely mathematical account comes with an implicit semantics: the set of hypotheses about underlying causal factors (or the ontology) with which the system parses and makes sense of its sensory stream. This might be seen as vindicating the structural representationalist account discussed in the introduction, albeit with a critical twist: those structures that bear content are not merely neural representations, but indeed phenotypic representations (if they are representations at all), for it is all the internal states of an organism, given the Markovian partition, that bear this content.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}





