
TAS1R3 and TAS2R38 Polymorphisms Affect Sweet Taste Perception: An Observational Study on Healthy and Obese Subjects


 ,
,  , , ,
, , , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Population
2.2. Taste Test
2.3. DNA Isolation and Genotyping
2.4. Statistical Analysis
3. Results
3.1. Subject Characteristics and SNP Genotyping
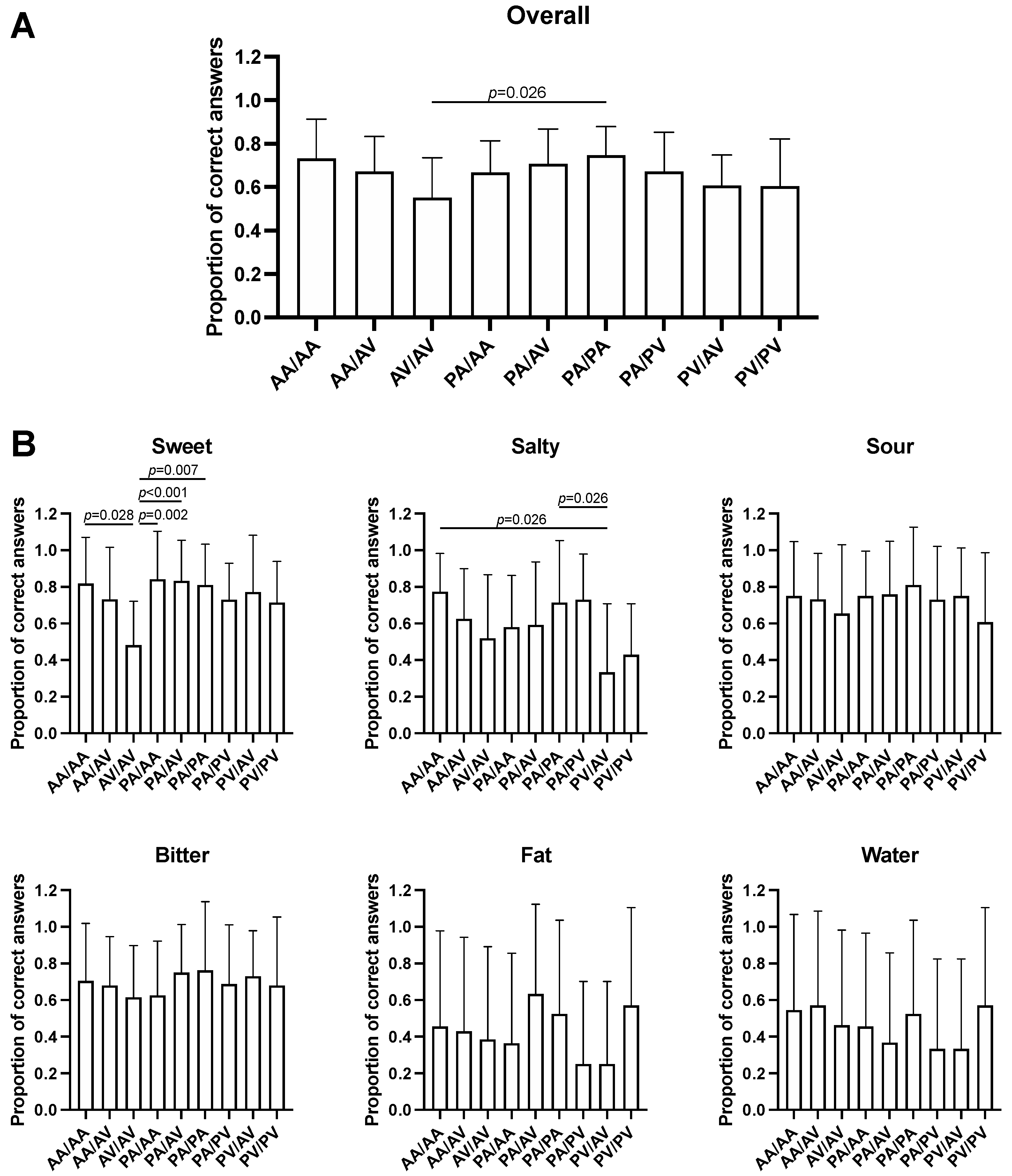
3.2. Association between TAS2R38 SNPs and Taste Recognition
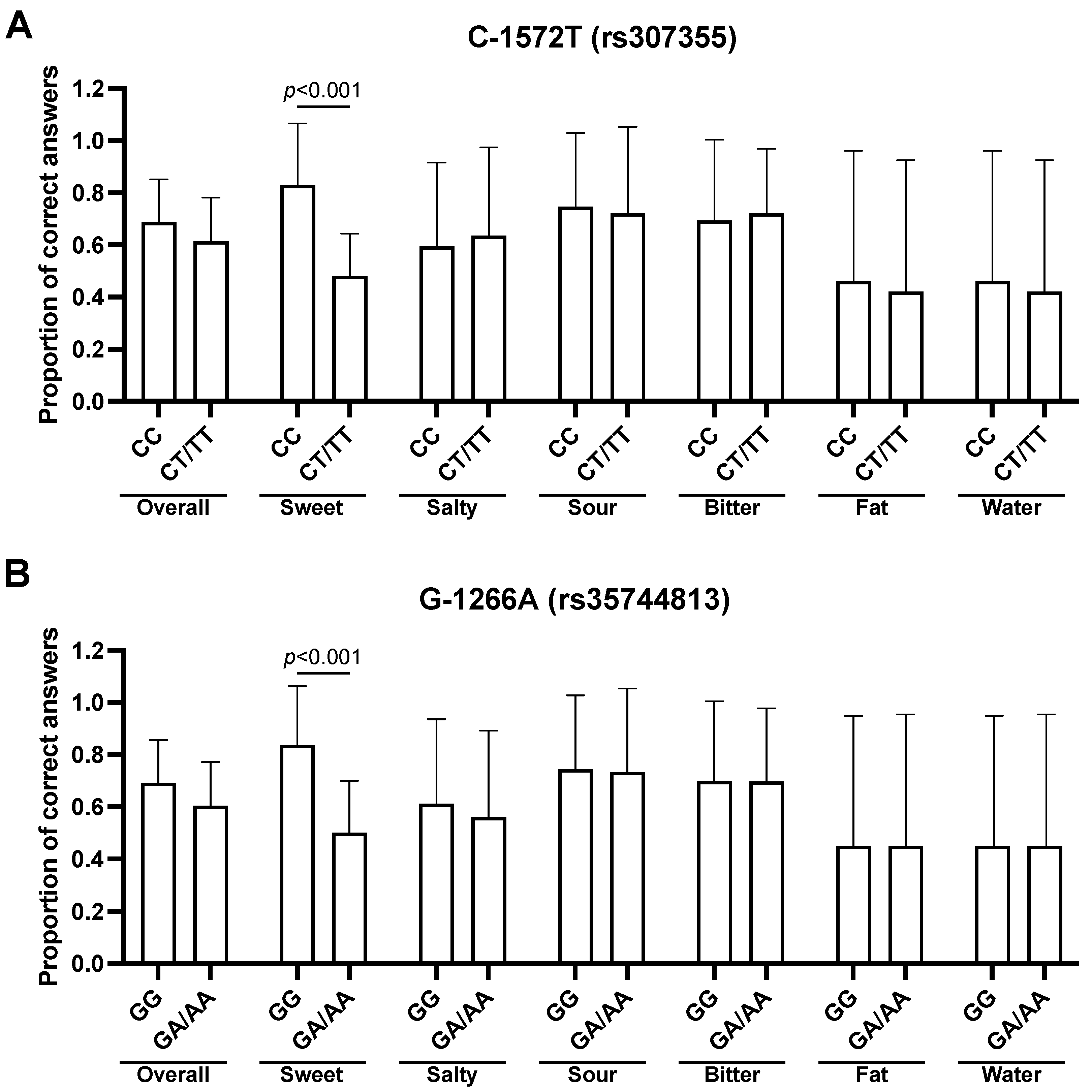
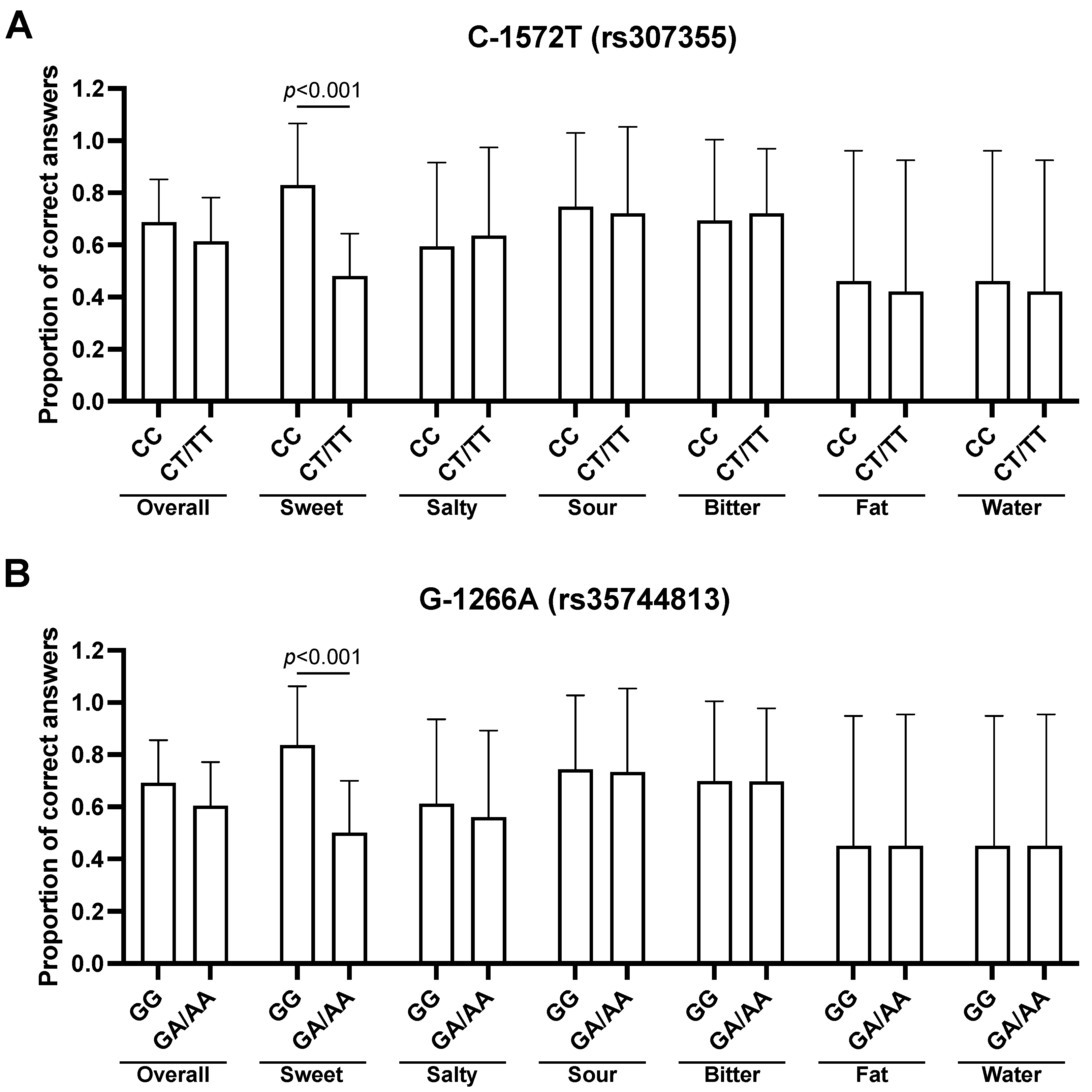
3.3. Association between TAS1R3 SNPs and Taste Recognition
3.4. Effect of Overweight, Obesity, and Taste Receptor SNPs on Taste Recognition
3.5. Multivariate Analysis for the Effects of TAS2R38 and TAS1R3 SNPs on Sweet Taste Recognition
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Jung, R.T. Obesity as a disease. Br. Med. Bull. 1997, 53, 307–321. [Google Scholar] [CrossRef]
- Pineda, E.; Sanchez-Romero, L.M.; Brown, M.; Jaccard, A.; Jewell, J.; Galea, G.; Webber, L.; Breda, J. Forecasting Future Trends in Obesity across Europe: The Value of Improving Surveillance. Obes. Facts 2018, 11, 360–371. [Google Scholar] [CrossRef] [PubMed]
- Boesveldt, S.; de Graaf, K. The Differential Role of Smell and Taste For Eating Behavior. Perception 2017, 46, 307–319. [Google Scholar] [CrossRef] [PubMed]
- Hayes, J.E.; Feeney, E.L.; Allen, A.L. Do polymorphisms in chemosensory genes matter for human ingestive behavior? Food Qual. Prefer. 2013, 30, 202–216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feeney, E.; O’Brien, S.; Scannell, A.; Markey, A.; Gibney, E.R. Genetic variation in taste perception: Does it have a role in healthy eating? Proc. Nutr. Soc. 2010, 70, 135–143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duffy, V.B. Variation in oral sensation: Implications for diet and health. Curr. Opin. Gastroenterol. 2007, 23, 171–177. [Google Scholar] [CrossRef]
- Roper, S.D.; Chaudhari, N. Taste buds: Cells, signals and synapses. Nat. Rev. Neurosci. 2017, 18, 485–497. [Google Scholar] [CrossRef]
- Reed, D.R.; Tanaka, T.; McDaniel, A.H. Diverse tastes: Genetics of sweet and bitter perception. Physiol. Behav. 2006, 88, 215–226. [Google Scholar] [CrossRef] [Green Version]
- Drayna, D. Human Taste Genetics. Annu. Rev. Genom. Hum. Genet. 2005, 6, 217–235. [Google Scholar] [CrossRef]
- Kim, U.-K.; Jorgenson, E.; Coon, H.; Leppert, M.; Risch, N.; Drayna, D. Positional Cloning of the Human Quantitative Trait Locus Underlying Taste Sensitivity to Phenylthiocarbamide. Science 2003, 299, 1221–1225. [Google Scholar] [CrossRef]
- Wooding, S.; Gunn, H.; Ramos, P.; Thalmann, S.; Xing, C.; Meyerhof, W. Genetics and Bitter Taste Responses to Goitrin, a Plant Toxin Found in Vegetables. Chem. Senses 2010, 35, 685–692. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hayes, J.E.; Bartoshuk, L.M.; Kidd, J.R.; Duffy, V.B. Supertasting and PROP Bitterness Depends on More Than the TAS2R38 Gene. Chem. Sens. 2008, 33, 255–265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bufe, B.; Breslin, P.A.S.; Kuhn, C.; Reed, D.R.; Tharp, C.D.; Slack, J.P.; Kim, U.-K.; Drayna, D.; Meyerhof, W. The molecular basis of individual differences in phenylthiocarbamide and propylthiouracil bitterness perception. Curr. Biol. 2005, 15, 322–327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yeomans, M.R.; Vi, C.; Mohammed, N.; Armitage, R.M. Re-evaluating how sweet-liking and PROP-tasting are related. Physiol. Behav. 2022, 246, 113702. [Google Scholar] [CrossRef] [PubMed]
- Diószegi, J.; Llanaj, E.; Ádány, R. Genetic Background of Taste Perception, Taste Preferences, and Its Nutritional Implications: A Systematic Review. Front. Genet. 2019, 10, 1272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Q.; Kraft, M.; Shen, Y.; MacFie, H.; Ford, R. Sweet Liking Status and PROP Taster Status impact emotional response to sweetened beverage. Food Qual. Prefer. 2019, 75, 133–144. [Google Scholar] [CrossRef]
- Reed, D.R.; Zhu, G.; Breslin, P.A.; Duke, F.F.; Henders, A.K.; Campbell, M.; Montgomery, G.W.; Medland, S.E.; Martin, N.G.; Wright, M.J. The perception of quinine taste intensity is associated with common genetic variants in a bitter receptor cluster on chromosome 12. Hum. Mol. Genet. 2010, 19, 4278–4285. [Google Scholar] [CrossRef] [Green Version]
- Lipchock, S.V.; Mennella, J.A.; Spielman, A.I.; Reed, D.R. Human bitter perception correlates with bitter receptor messenger RNA expression in taste cells. Am. J. Clin. Nutr. 2013, 98, 1136–1143. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Staszewski, L.; Xu, H.; Durick, K.; Zoller, M.; Adler, E. Human receptors for sweet and umami taste. Proc. Natl. Acad. Sci. USA 2002, 99, 4692–4696. [Google Scholar] [CrossRef] [Green Version]
- Chandrashekar, J.; Hoon, M.A.; Ryba, N.J.P.; Zuker, C.S. The receptors and cells for mammalian taste. Nature 2006, 444, 288–294. [Google Scholar] [CrossRef]
- Fushan, A.A.; Simons, C.T.; Slack, J.P.; Manichaikul, A.; Drayna, D. Allelic polymorphism within the TAS1R3 promoter is as-sociated with human taste sensitivity to sucrose. Curr. Biol. 2009, 19, 1288–1293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hwang, L.-D.; Lin, C.; Gharahkhani, P.; Cuellar-Partida, G.; Ong, J.-S.; An, J.; Gordon, S.D.; Zhu, G.; MacGregor, S.; Lawlor, D.A.; et al. New insight into human sweet taste: A genome-wide association study of the perception and intake of sweet substances. Am. J. Clin. Nutr. 2019, 109, 1724–1737. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Landis, B.N.; Welge-Luessen, A.; Brämerson, A.; Bende, M.; Mueller, C.A.; Nordin, S.; Hummel, T. “Taste Strips”—A rapid, lateralized, gustatory bedside identification test based on impregnated filter papers. J. Neurol. 2009, 256, 242–248. [Google Scholar] [CrossRef] [Green Version]
- Pugnaloni, S.; Alia, S.; Mancini, M.; Santoro, V.; Di Paolo, A.; Rabini, R.A.; Fiorini, R.; Sabbatinelli, J.; Fabri, M.; Mazzanti, L.; et al. A Study on the Relationship between Type 2 Diabetes and Taste Function in Patients with Good Glycemic Control. Nutrients 2020, 12, 1112. [Google Scholar] [CrossRef] [PubMed]
- Pugnaloni, S.; Vignini, A.; Borroni, F.; Sabbatinelli, J.; Alia, S.; Fabri, M.; Taus, M.; Mazzanti, L.; Berardi, R. Modifications of taste sensitivity in cancer patients: A method for the evaluations of dysgeusia. Support. Care Cancer 2020, 28, 1173–1181. [Google Scholar] [CrossRef] [PubMed]
- Vignini, A.; Borroni, F.; Sabbatinelli, J.; Pugnaloni, S.; Alia, S.; Taus, M.; Ferrante, L.; Mazzanti, L.; Fabri, M. General Decrease of Taste Sensitivity Is Related to Increase of BMI: A Simple Method to Monitor Eating Behavior. Dis. Markers 2019, 2019, 2978026. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Athanasiadou, E.; Kyrkou, C.; Fotiou, M.; Tsakoumaki, F.; Dimitropoulou, A.; Polychroniadou, E.; Menexes, G.; Athanasiadis, A.P.; Biliaderis, C.G.; Michaelidou, A.-M. Development and Validation of a Mediterranean Oriented Culture-Specific Semi-Quantitative Food Frequency Questionnaire. Nutrients 2016, 8, 522. [Google Scholar] [CrossRef] [Green Version]
- The Diabetes Prevention Program Research Group. Relationship of body size and shape to the development of diabetes in the diabetes prevention program. Obesity 2006, 14, 2107–2117. [Google Scholar] [CrossRef] [Green Version]
- Costanzo, A.; Orellana, L.; Nowson, C.; Duesing, K.; Keast, R. Fat Taste Sensitivity Is Associated with Short-Term and Habitual Fat Intake. Nutrients 2017, 9, 781. [Google Scholar] [CrossRef] [Green Version]
- Stephens, M.; Donnelly, P. A Comparison of Bayesian Methods for Haplotype Reconstruction from Population Genotype Data. Am. J. Hum. Genet. 2003, 73, 1162–1169. [Google Scholar] [CrossRef] [Green Version]
- Pronin, A.N.; Xu, H.; Tang, H.; Zhang, L.; Li, Q.; Li, X. Specific Alleles of Bitter Receptor Genes Influence Human Sensitivity to the Bitterness of Aloin and Saccharin. Curr. Biol. 2007, 17, 1403–1408. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dias, A.G.; Eny, K.M.; Cockburn, M.; Chiu, W.; Nielsen, D.; Duizer, L.; El-Sohemy, A. Variation in the TAS1R2 Gene, Sweet Taste Perception and Intake of Sugars. J. Nutr. Nutr. 2015, 8, 81–90. [Google Scholar] [CrossRef] [PubMed]
- Fushan, A.A.; Simons, C.T.; Slack, J.P.; Drayna, D. Association between common variation in genes encoding sweet taste sig-naling components and human sucrose perception. Chem. Sen. 2010, 35, 579–592. [Google Scholar] [CrossRef] [PubMed]
- Skrandies, W.; Zschieschang, R. Olfactory and gustatory functions and its relation to body weight. Physiol. Behav. 2015, 142, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Hwang, L.-D.; Cuellar-Partida, G.; Ong, J.-S.; Breslin, P.A.S.; Reed, D.R.; MacGregor, S.; Gharahkhani, P.; Martin, N.G.; Rentería, M.E. Sweet Taste Perception is Associated with Body Mass Index at the Phenotypic and Genotypic Level. Twin Res. Hum. Genet. 2016, 19, 465–471. [Google Scholar] [CrossRef] [Green Version]
- Anderson, G.H. Sugars, sweetness, and food intake. Am. J. Clin. Nutr. 1995, 62, 195S–201S. [Google Scholar] [CrossRef]
- Joseph, P.V.; Reed, D.R.; Mennella, J.A. Individual Differences Among Children in Sucrose Detection Thresholds: Relationship With Age, Gender, and Bitter Taste Genotype. Nurs. Res. 2016, 65, 3–12. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
| Stimulus | Substance | Concentration (g/mL) |
|---|---|---|
| Sweet | Sucrose | −0.05 |
| −0.1 | ||
| −0.2 | ||
| −0.4 | ||
| Salty | Sodium Chloride | −0.016 |
| −0.04 | ||
| −0.1 | ||
| −0.25 | ||
| Bitter | Quinine hydrochloride | −0.0004 |
| −0.0009 | ||
| −0.0024 | ||
| −0.006 | ||
| Sour | Citric acid | −0.05 |
| −0.09 | ||
| −0.165 | ||
| −0.3 | ||
| Fat | Rape oil | Pure |
| Neutral | Deionized water | Pure |
| SNP | Primer Fw (5’-3’) | Primer Rv (5’-3’) | Control Primer (5’-3’) |
|---|---|---|---|
| TAS2R38 A49P (rs713598) | GGTGGCAACCAGGTCTTTAG | CACAATCACTGTTGCTCAGTGC /CACAATCACTGTTGCTCAGTGG | GATGGCTTGGTAGCTGTGGT |
| TAS2R38 V262A (rs1726866) | CTTCTTTGTGATATCATCCTGTGT/ CTTCTTTGTGATATCATCCTGTGC | TGTGGTCGGCTCTTACCTTC | GGAAGGCACATGAGGACAAT |
| TAS1R3 C-1572T (rs307355) | AATGTGCAGGTGCCAGTTG | ACATGGTACACGCAAAGCG /ACATGGTACACGCAAAGCA | CACGGCACACACAATACACA |
| TAS1R3 G-1266A (rs35744813) | TGTGAGGGACACACACTACCA | ATGTATGCTGTGCACGTGC /ATGTATGCTGTGCACGTGT | GTGCCGTTTCCGTGTGTATT |
| OW/OB (n = 85) | NW (n = 57) | p | |
|---|---|---|---|
| BMI (Kg/m2) | 31 ± 4.6 | 22 ± 2.3 | <0.001 |
| Gender (F/M) | 52/33 | 41/16 | 0.186 |
| Age (years) | 45 ± 3 | 48 ± 4.2 | 0.857 |
| Total Cholesterol (mg/dL) | 194 ± 15 | 186 ± 13 | 0.081 |
| HDL-Cholesterol (mg/dL) | 41 ± 3 | 45 ± 4 | 0.072 |
| LDL-Cholesterol (mg/dL) | 124 ± 9 | 119 ± 8 | 0.068 |
| Triglycerides (mg/dL) | 141 ± 11 | 92 ± 7 | 0.061 |
| TAS2R38 A49P-V262A haplotype frequencies | |||
| PA (CC) | 0.29 | 0.39 | |
| PV (CT) | 0.19 | 0.16 | |
| AA (GC) | 0.26 | 0.22 | |
| AV (GT) | 0.25 | 0.24 | 0.434 |
| TAS1R3 C-1572T (rs307355) allelic frequencies | |||
| C | 0.86 | 0.94 | |
| T | 0.14 | 0.06 | 0.888 |
| TAS1R3 G-1266A (rs35744813) allelic frequencies | |||
| G | 0.85 | 0.90 | |
| A | 0.15 | 0.10 | 0.370 |
| Stimuli | OW/OB (n = 85) | NW (n = 57) | p |
|---|---|---|---|
| Sweet | 0.747 (0.278) | 0.797 (0.227) | 1 |
| Salty | 0.569 (0.311) | 0.647 (0.341) | 1 |
| Bitter | 0.658 (0.300) | 0.759 (0.289) | 0.336 |
| Sour | 0.682 (0.309) | 0.828 (0.240) | 0.022 |
| Fat | 0.310 (0.465) | 0.660 (0.479) | <0.001 |
| Neutral | 0.390 (0.491) | 0.530 (0.503) | 0.709 |
| Overall | 0.629 (0.180) | 0.740 (0.122) | <0.001 |
| Source of Variation | df | Mean Square | F | p | Partial eta Squared | Observed Power |
|---|---|---|---|---|---|---|
| Age | 1 | 0.057 | 1.745 | 0.189 | 0.014 | 0.259 |
| BMI | 1 | 0.030 | 0.929 | 0.337 | 0.007 | 0.160 |
| Gender | 1 | 0.024 | 0.731 | 0.394 | 0.006 | 0.136 |
| TAS2R38 haplotype | 8 | 0.117 | 3.041 | 0.004 | 0.160 | 2.007 |
| TAS1R3 C-1572T | 1 | 0.254 | 7.752 | 0.006 | 0.059 | 0.789 |
| TAS1R3 G-1266A | 1 | 0.260 | 7.915 | 0.006 | 0.060 | 0.797 |
| Error | 128 | 0.039 | ||||
| SNP | n | Adjusted Mean | SEM | p |
|---|---|---|---|---|
| Model 1—TAS2R38 diplotype | ||||
| AA/AA | 11 | 0.861 | 0.076 | Ref. |
| AA/AV | 14 | 0.727 | 0.066 | 0.187 |
| AV/AV | 13 | 0.478 | 0.068 | <0.001 |
| PA/AA | 22 | 0.852 | 0.052 | 0.928 |
| PA/AV | 30 | 0.842 | 0.046 | 0.837 |
| PA/PA | 21 | 0.790 | 0.056 | 0.458 |
| PA/PV | 12 | 0.757 | 0.072 | 0.315 |
| PV/AV | 12 | 0.794 | 0.072 | 0.518 |
| PV/PV | 7 | 0.730 | 0.094 | 0.275 |
| Model 2—TAS1R3 SNPs | ||||
| TAS1R3 C-1572T (rs307355) | ||||
| CC | 118 | 0.698 | 0.028 | Ref. |
| CT/TT | 24 | 0.490 | 0.042 | 0.001 |
| TAS1R3 G-1266A (rs35744813) | ||||
| GG | 113 | 0.685 | 0.034 | Ref. |
| GA/AA | 29 | 0.502 | 0.038 | 0.001 |
| Model 3—Combined | ||||
| TAS2R38 diplotype | ||||
| AA/AA | 11 | 0.683 | 0.0648 | Ref. |
| AA/AV | 14 | 0.608 | 0.0551 | 0.365 |
| AV/AV | 13 | 0.392 | 0.0560 | < 0.001 |
| PA/AA | 22 | 0.690 | 0.0463 | 0.923 |
| PA/AV | 30 | 0.690 | 0.0414 | 0.918 |
| PA/PA | 21 | 0.662 | 0.0481 | 0.790 |
| PA/PV | 12 | 0.638 | 0.0598 | 0.590 |
| PV/AV | 12 | 0.642 | 0.0604 | 0.625 |
| PV/PV | 7 | 0.598 | 0.0780 | 0.381 |
| TAS1R3 C-1572T (rs307355) | ||||
| CC | 118 | 0.625 | 0.035 | Ref. |
| CT/TT | 24 | 0.465 | 0.046 | 0.006 |
| TAS1R3 G-1266A (rs35744813) | ||||
| GG | 113 | 0.626 | 0.039 | Ref. |
| GA/AA | 29 | 0.464 | 0.042 | 0.006 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cecati, M.; Vignini, A.; Borroni, F.; Pugnaloni, S.; Alia, S.; Sabbatinelli, J.; Nicolai, G.; Taus, M.; Santarelli, A.; Fabri, M.; et al. TAS1R3 and TAS2R38 Polymorphisms Affect Sweet Taste Perception: An Observational Study on Healthy and Obese Subjects. Nutrients 2022, 14, 1711. https://doi.org/10.3390/nu14091711
Cecati M, Vignini A, Borroni F, Pugnaloni S, Alia S, Sabbatinelli J, Nicolai G, Taus M, Santarelli A, Fabri M, et al. TAS1R3 and TAS2R38 Polymorphisms Affect Sweet Taste Perception: An Observational Study on Healthy and Obese Subjects. Nutrients. 2022; 14(9):1711. https://doi.org/10.3390/nu14091711
Chicago/Turabian StyleCecati, Monia, Arianna Vignini, Francesca Borroni, Sofia Pugnaloni, Sonila Alia, Jacopo Sabbatinelli, Giulia Nicolai, Marina Taus, Andrea Santarelli, Mara Fabri, and et al. 2022. "TAS1R3 and TAS2R38 Polymorphisms Affect Sweet Taste Perception: An Observational Study on Healthy and Obese Subjects" Nutrients 14, no. 9: 1711. https://doi.org/10.3390/nu14091711
APA StyleCecati, M., Vignini, A., Borroni, F., Pugnaloni, S., Alia, S., Sabbatinelli, J., Nicolai, G., Taus, M., Santarelli, A., Fabri, M., Mazzanti, L., & Emanuelli, M. (2022). TAS1R3 and TAS2R38 Polymorphisms Affect Sweet Taste Perception: An Observational Study on Healthy and Obese Subjects. Nutrients, 14(9), 1711. https://doi.org/10.3390/nu14091711