
Clinical and Microbiological Features of Fulminant Haemolysis Caused by Clostridium perfringens Bacteraemia: Unknown Pathogenesis


Abstract
1. Introduction
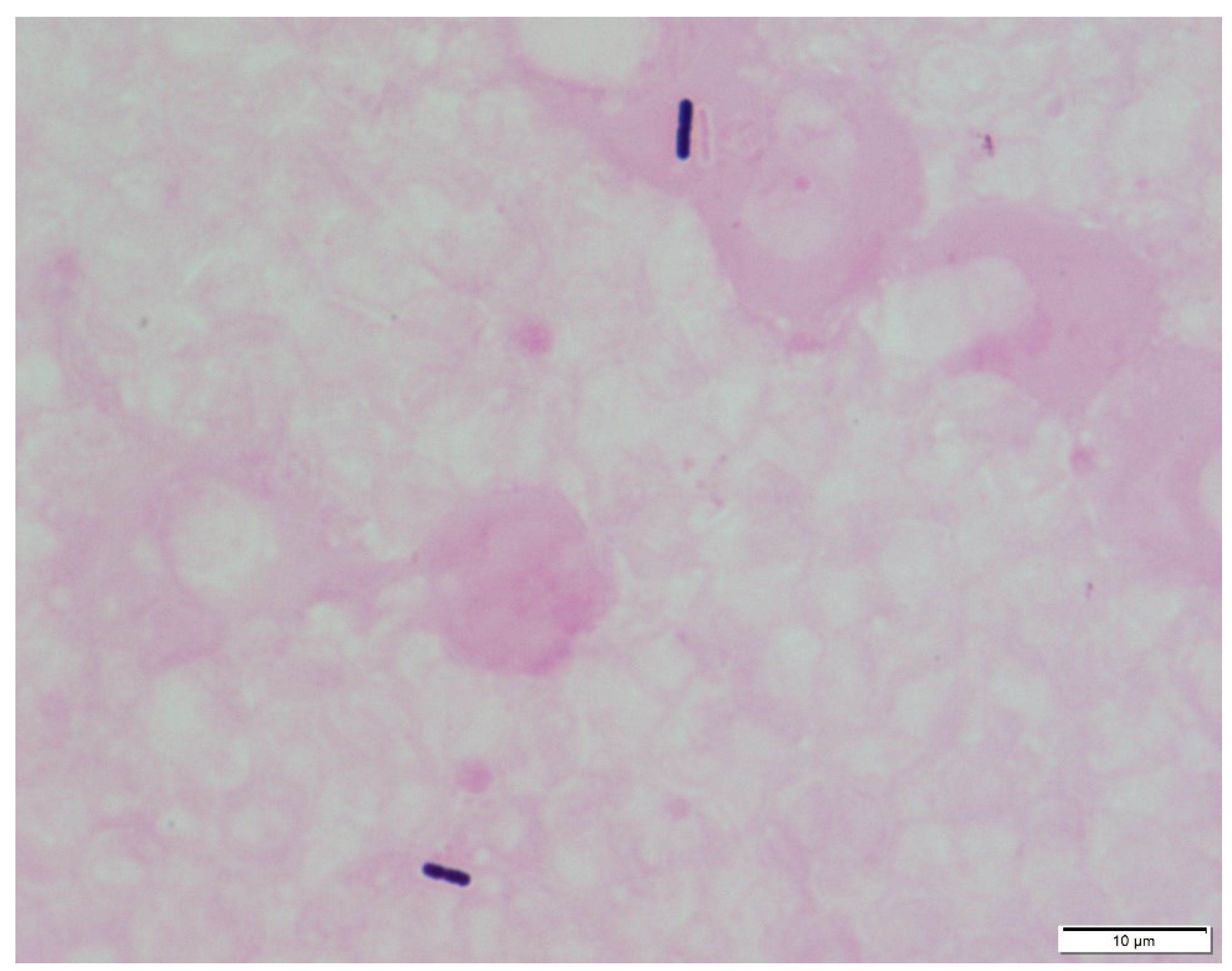
2. Clostridium Perfringens
3. C. perfringens Infection with MIH
Epidemiology
4. Pathogenic Factors
5. Symptoms and Laboratory Findings
6. Diagnosis with/without Bacterial Culture
7. State-of-the-Art Treatment and Prognosis
8. Inflammatory Foci or Bacterial Translocation Preceding C. pefringens Invasion Pathways
9. Evolutionary Significance of Bacterial Haemolysis
10. Involvement of Haemolysis in Pathophysiology of C. Perfringens MIH and Potential for Iron/Haem Scavenging Therapy
11. Local or Systemic Modulation of Immune Function by Genus Clostridium and How to Control Them
12. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Morel, G.; Mulier, G.; Ghrenassia, E.; Abdel-Nabey, M.; Tandjaoui, Y.; Kouatchet, A.; Platon, L.; Pène, F.; Moreau, A.S.; Seguin, A.; et al. Non-C. difficile Clostridioides bacteremia in intensive care patients, France. Emerg. Infect. Dis. 2021, 27, 1840–1849. [Google Scholar] [CrossRef]
- Van Bunderen, C.C.; Bomers, M.K.; Wesdorp, E.; Peerbooms, P.; Veenstra, J. Clostridium perfringens septicaemia with massive intravascular haemolysis: A case report and review of the literature. Neth. J. Med. 2010, 68, 343–346. [Google Scholar]
- Simon, T.G.; Bradley, J.; Jones, A.; Carino, G. Massive intravascular hemolysis from Clostridium perfringens septicemia: A review. J. Intensive Care Med. 2014, 29, 327–333. [Google Scholar] [CrossRef]
- Liu, F.; Xue, S.; Zhang, Y.; Yang, J.; Hu, J.; Li, D.; Ma, X.; Wang, J. Clostridium perfringens sepsis in three patients with acute leukemia and review of the literature. Int. J. Hematol. 2021, 113, 508–517. [Google Scholar] [CrossRef]
- Suzaki, A.; Komine-Aizawa, S.; Nishiyama, H.; Hayakawa, S. Massive intravascular hemolysis is an important factor in Clostridium perfringens-induced bacteremia. Intern. Emerg. Med. 2022, 17, 1959–1967. [Google Scholar] [CrossRef]
- Woittiez, N.J.C.; Van Prehn, J.; Van Immerseel, F.; Goossens, E.; Bauer, M.P.; Ramspek, C.L.; Slangen, R.M.E.; Purmer, I.M.; Ludikhuize, J. Toxinotype A Clostridium perfringens causing septicaemia with intravascular haemolysis: Two cases and review of the literature. Int. J. Infect. Dis. 2022, 115, 224–228. [Google Scholar] [CrossRef]
- Bauer, M.; Gerlach, H.; Vogelmann, T.; Preissing, F.; Stiefel, J.; Adam, D. Mortality in sepsis and septic shock in Europe, North America and Australia between 2009 and 2019-results from a systematic review and meta-analysis. Crit. Care 2020, 24, 239. [Google Scholar] [CrossRef]
- Kern, W.V.; Rieg, S. Burden of bacterial bloodstream infection-a brief update on epidemiology and significance of multidrug-resistant pathogens. Clin. Microbiol. Infect. 2020, 26, 151–157. [Google Scholar] [CrossRef]
- Adams, B.N.; Lekovic, J.P.; Robinson, S. Clostridium perfringens sepsis following a molar pregnancy. Am. J. Obstet. Gynecol. 2014, 210, e13–e14. [Google Scholar] [CrossRef]
- Kiu, R.; Hall, L.J. An update on the human and animal enteric pathogen Clostridium perfringens. Emerg. Microbes Infect. 2018, 7, 141. [Google Scholar] [CrossRef]
- Uzal, F.A.; Freedman, J.C.; Shrestha, A.; Theoret, J.R.; Garcia, J.; Awad, M.M.; Adams, V.; Moore, R.J.; Rood, J.I.; McClane, B.A. Towards an understanding of the role of Clostridium perfringens toxins in human and animal disease. Future Microbiol. 2014, 9, 361–377. [Google Scholar] [CrossRef]
- Rechner, P.M.; Agger, W.A.; Mruz, K.; Cogbill, T.H. Clinical features of clostridial bacteremia: A review from a rural area. Clin. Infect. Dis. 2001, 33, 349–353. [Google Scholar] [CrossRef]
- Stabler, S.; Titécat, M.; Duployez, C.; Wallet, F.; Loïez, C.; Bortolotti, P.; Faure, E.; Faure, K.; Kipnis, E.; Dessein, R.; et al. Clinical relevance of Clostridium bacteremia: An 8-year retrospective study. Anaerobe 2020, 63, 102202. [Google Scholar] [CrossRef]
- Leal, J.; Gregson, D.B.; Ross, T.; Church, D.L.; Laupland, K.B. Epidemiology of Clostridium species bacteremia in Calgary, Canada, 2000–2006. J. Infect. 2008, 57, 198–203. [Google Scholar] [CrossRef]
- Martín, S.; Pérez, A.; Aldecoa, C. Sepsis and immunosenescence in the elderly patient: A review. Front. Med. 2017, 4, 20. [Google Scholar] [CrossRef]
- Dhaliwal, G.; Cornett, P.A.; Tierney, L.M., Jr. Hemolytic anemia. Am. Fam. Phys. 2004, 69, 2599–2606. [Google Scholar]
- Ou, T.Y.; Chuang, C.Y.; Chen, C.D.; Cheng, C.Y. Therapeutic plasma exchange in the treatment of complicated Plasmodium falciparum malaria: A case report. J. Clin. Apher. 2018, 33, 419–422. [Google Scholar] [CrossRef]
- Setty, S.; Khalil, Z.; Schori, P.; Azar, M.; Ferrieri, P. Babesiosis: Two atypical cases from Minnesota and a review. Am. J. Clin. Pathol. 2003, 120, 554–559. [Google Scholar] [CrossRef]
- Jakharia, N.; Hossain, A.; Luethy, P.; Riedel, D.J. 48-year-old male with febrile neutropenia and massive hemolysis. Clin. Infect. Dis. 2019, 69, 2193–2194. [Google Scholar] [CrossRef]
- Yang, C.C.; Hsu, P.C.; Chang, H.J.; Cheng, C.W.; Lee, M.H. Clinical significance and outcomes of Clostridium perfringens bacteremia—A 10-year experience at a tertiary care hospital. Int. J. Infect. Dis. 2013, 17, e955–e960. [Google Scholar] [CrossRef]
- Shindo, Y.; Dobashi, Y.; Sakai, T.; Monma, C.; Miyatani, H.; Yoshida, Y. Epidemiological and pathobiological profiles of Clostridium perfringens infections: Review of consecutive series of 33 cases over a 13-year period. Int. J. Clin. Exp. Pathol. 2015, 8, 569–577. [Google Scholar] [PubMed]
- Haddy, R.I.; Nadkarni, D.D.; Mann, B.L.; Little, D.R.; Domers, T.D.; Clover, R.D.; Silvers, M.J. Clostridial bacteremia in the community hospital. Scand. J. Infect. Dis. 2000, 32, 27–30. [Google Scholar] [CrossRef] [PubMed]
- Sakaue, M.; Ota, K.; Nakamura, E.; Nitta, M.; Oka, M.; Oishi, Y.; Sano, Y.; Yonogi, S.; Takasu, A. Type A fuluminant Clostridium perfringens sepsis indicated RBC/Hb discrepancy; a case report. BMC Infect. Dis. 2019, 19, 719. [Google Scholar] [CrossRef]
- Hashiba, M.; Tomino, A.; Takenaka, N.; Hattori, T.; Kano, H.; Tsuda, M.; Takeyama, N. Clostridium perfringens infection in a febrile patient with severe hemolytic anemia. Am. J. Case Rep. 2016, 17, 219–223. [Google Scholar] [CrossRef]
- Wild, W.; Bormann, F.; Sweiti, H.; Tamimi, N.; Pichulek, D.; Divo, M.; Dörr, P.; Schwarzbach, M. Clostridium perfringens septicemia and a bleeding ulcer of a jejunal interposition: A case report and short review of the literature. Case Rep. Med. 2018, 2018, 4278904. [Google Scholar] [CrossRef] [PubMed]
- Suzaki, A.; Ohtani, K.; Komine-Aizawa, S.; Matsumoto, A.; Kamiya, S.; Hayakawa, S. Pathogenic characterization of Clostridium perfringens strains isolated from patients with massive intravascular hemolysis. Front. Microbiol. 2021, 12, 713509. [Google Scholar] [CrossRef]
- Ohtani, K.; Hirakawa, H.; Tashiro, K.; Yoshizawa, S.; Kuhara, S.; Shimizu, T. Identification of a two-component VirR/VirS regulon in Clostridium perfringens. Anaerobe 2010, 16, 258–264. [Google Scholar] [CrossRef]
- Popoff, M.R. Clostridial pore-forming toxins: Powerful virulence factors. Anaerobe 2014, 30, 220–238. [Google Scholar] [CrossRef]
- Bryant, A.E.; Bergstrom, R.; Zimmerman, G.A.; Salyer, J.L.; Hill, H.R.; Tweten, R.K.; Sato, H.; Stevens, D.L. Clostridium perfringens invasiveness is enhanced by effects of theta toxin upon PMNL structure and function: The roles of leukocytotoxicity and expression of CD11/CD18 adherence glycoprotein. FEMS Immunol. Med. Microbiol. 1993, 7, 321–336. [Google Scholar] [CrossRef]
- Awad, M.M.; Ellemor, D.M.; Boyd, R.L.; Emmins, J.J.; Rood, J.I. Synergistic effects of α-toxin and perfringolysin O in Clostridium perfringens-mediated gas gangrene. Infect. Immun. 2001, 69, 7904–7910. [Google Scholar] [CrossRef]
- Verherstraeten, S.; Goossens, E.; Valgaeren, B.; Pardon, B.; Timbermont, L.; Haesebrouck, F.; Ducatelle, R.; Deprez, P.; Wade, K.R.; Tweten, R.; et al. Perfringolysin O: The underrated Clostridium perfringens toxin? Toxins 2015, 7, 1702–1721. [Google Scholar] [CrossRef] [PubMed]
- Verherstraeten, S.; Goossens, E.; Valgaeren, B.; Pardon, B.; Timbermont, L.; Vermeulen, K.; Schauvliege, S.; Haesebrouck, F.; Ducatelle, R.; Deprez, P.; et al. The synergistic necrohemorrhagic action of Clostridium perfringens perfringolysin and alpha toxin in the bovine intestine and against bovine endothelial cells. Vet. Res. 2013, 44, 45. [Google Scholar] [CrossRef] [PubMed]
- Chinen, K. Sudden death caused by Clostridium perfringens sepsis presenting as massive intravascular hemolysis. Autops. Case Rep. 2020, 10, e2020185. [Google Scholar] [CrossRef]
- Kang, S.; Tanaka, T.; Kishimoto, T. Therapeutic uses of anti-interleukin-6 receptor antibody. Int. Immunol. 2014, 27, 21–29. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, R.; Makino, Y.; Chiba, F.; Motomura, A.; Inokuchi, G.; Yajima, D.; Iwase, H. Fatal Clostridium perfringens septicemia suggested by postmortem computed tomography: A medico-legal autopsy case report. Forensic Sci. Int. 2015, 253, e4–e9. [Google Scholar] [CrossRef]
- Uppal, A.; Hymes, K.; Schwartz, D.R. A 61-year-old-man with massive intravascular hemolysis. Chest 2009, 136, 1424–1427. [Google Scholar] [CrossRef]
- Yan, J.; Li, S.; Li, S. The role of the liver in sepsis. Int. Rev. Immunol. 2014, 33, 498–510. [Google Scholar] [CrossRef]
- Woźnica, E.A.; Inglot, M.; Woźnica, R.K.; Łysenko, L. Liver dysfunction in sepsis. Adv. Clin. Exp. Med. 2018, 27, 547–551. [Google Scholar] [CrossRef]
- Ng, H.; Lam, S.M.; Shum, H.P.; Yan, W.W. Clostridium perfringens liver abscess with massive haemolysis. Hong Kong Med. J. 2010, 16, 310–312. [Google Scholar]
- Cécilia, R.; Baptiste, V.; Benjamin, C.; Virginie, H.; Guillaume, V.; Philippe, R.; Matthieu, B. Acute hemolysis in the emergency department: Think about Clostridium perfringens! Case Rep. Emerg. Med. 2013, 2013, 948071. [Google Scholar] [CrossRef]
- Ohtani, S.; Watanabe, N.; Kawata, M.; Harada, K.; Himei, M.; Murakami, K. Massive intravascular hemolysis in a patient infected by Clostridium perfringens. Acta. Med. Okayama 2006, 60, 357–360. [Google Scholar] [CrossRef] [PubMed]
- McArthur, H.L.; Dalal, B.I.; Kollmannsberger, C. Intravascular hemolysis as a complication of Clostridium perfringens sepsis. J. Clin. Oncol. 2006, 24, 2387–2388. [Google Scholar] [CrossRef] [PubMed]
- Kumar, V. Toll-like receptors in sepsis-associated cytokine storm and their endogenous negative regulators as future immunomodulatory targets. Int. Immunopharmacol. 2020, 89 Pt B, 107087. [Google Scholar] [CrossRef]
- Hackett, S.P.; Stevens, D.L. Streptococcal toxic shock syndrome: Synthesis of tumor necrosis factor and interleukin-1 by monocytes stimulated with pyrogenic exotoxin A and streptolysin O. J. Infect. Dis. 1992, 165, 879–888. [Google Scholar] [CrossRef] [PubMed]
- Ikejima, T.; Dinarello, C.A.; Gill, D.M.; Wolf, S.M. Induction of human interleukin-1 by a product of Staphylococcus aureus associated with toxic shock syndrome. J. Clin. Investig. 1984, 73, 1312–1320. [Google Scholar] [CrossRef] [PubMed]
- Sarvari, K.P.; Vasas, B.; Kiss, I.; Lazar, A.; Horvath, I.; Simon, M.; Peto, Z.; Urban, E. Fatal Clostridium perfringens sepsis due to emphysematous gastritis and literature review. Anaerobe 2016, 40, 31–34. [Google Scholar] [CrossRef] [PubMed]
- Plowden, J.; Renshaw-Hoelscher, M.; Engleman, C.; Katz, J.; Sambhara, S. Innate immunity in aging: Impact on macrophage funcion. Aging Cell 2004, 3, 161–167. [Google Scholar] [CrossRef]
- Zhang, J.M.; An, J. Cytokines, infammation, and pain. Int. Anesthesiol. Clin. 2007, 45, 27–37. [Google Scholar] [CrossRef]
- Yang, Y.; Li, Q.; Tan, F.; Zhang, J.; Zhu, W. Mechanism of IL-8-induced acute lung injury through pulmonary surfactant proteins A and B. Exp. Ther. Med. 2020, 19, 287–293. [Google Scholar] [CrossRef]
- Aisiku, I.P.; Yamal, J.M.; Doshi, P.; Benoit, J.S.; Gopinath, S.; Goodman, J.C.; Robertson, C.S. Plasma cytokines IL-6, IL-8, and IL-10 are associated with the development of acute respiratory distress syndrome in patients with severe traumatic brain injury. Crit. Care 2016, 20, 288. [Google Scholar] [CrossRef]
- Fujikawa, H.; Araki, M. Clostridium perfringens septicemia with massive intravascular hemolysis. Intern. Med. 2020, 59, 591. [Google Scholar] [CrossRef] [PubMed]
- Johnson, S. Antibody responses to clostridial infection in humans. Clin. Infect. Dis. 1997, 25 (Suppl. S2), S173–S177. [Google Scholar] [CrossRef] [PubMed]
- McSwain, B.; Sawyers, J.L.M.; Lawler, R., Jr. Clostridial infections of the abdominal wall: Review of 10 cases. Ann. Surg. 1966, 163, 859–865. [Google Scholar] [CrossRef]
- Richard, O.K.; Grahofer, A.; Nathues, H.; Posthaus, H. Vaccination against Clostridium perfringens type C enteritis in pigs: A field study using an adapted vaccination scheme. Porcine Health Manag. 2019, 5, 20. [Google Scholar] [CrossRef] [PubMed]
- Thapa, R.; Ray, S.; Keyel, P.A. Interaction of macrophages and cholesterol-dependent cytolysins: The impact on immune response and cellular survival. Toxins 2020, 12, 531. [Google Scholar] [CrossRef]
- Vinchi, F.; Sparla, R.; Passos, S.T.; Sharma, R.; Vance, S.Z.; Zreid, H.S.; Juaidi, H.; Manwani, D.; Yazdanbakhsh, K.; Nandi, V.; et al. Vasculo-toxic and pro-inflammatory action of unbound haemoglobin, haem and iron in transfusion-dependent patients with haemolytic anaemias. Br. J. Haematol. 2021, 193, 637–658. [Google Scholar] [CrossRef]
- Vinchi, F.; Tolosano, E. Therapeutic approaches to limit hemolysis-driven endothelial dysfunction: Scavenging free heme to preserve vasculature homeostasis. Oxid. Med. Cell. Longev. 2013, 2013, 396527. [Google Scholar] [CrossRef]
- Chiabrando, D.; Vinchi, F.; Fiorito, V.; Mercurio, S.; Tolosano, E. Heme in pathophysiology: A matter of scavenging, metabolism and trafficking across cell membranes. Front. Pharmacol. 2014, 5, 61. [Google Scholar] [CrossRef]
- Belcher, J.D.; Mahaseth, H.; Welch, T.E.; Otterbein, L.E.; Hebbel, R.P.; Vercellotti, G.M. Heme oxygenase-1 is a modulator of inflammation and vaso-occlusion in transgenic sickle mice. J. Clin. Investig. 2006, 116, 808–816. [Google Scholar] [CrossRef]
- Smith, A.; McCulloh, R.J. Hemopexin and haptoglobin: Allies against heme toxicity from hemoglobin not contenders. Front. Physiol. 2015, 6, 187. [Google Scholar] [CrossRef]
- Liu, Y.; Pal, M.; Bao, W.; Shi, P.A.; Lobo, C.A.; An, X.; Manwani, D.; Zhong, H.; Yazdanbakhsh, K. Type I interferon is induced by hemolysis and drives antibody-mediated erythrophagocytosis in sickle cell disease. Blood 2021, 138, 1162–1171. [Google Scholar] [CrossRef] [PubMed]
- Atarashi, K.; Tanoue, T.; Shima, T.; Imaoka, A.; Kuwahara, T.; Momose, Y.; Cheng, G.; Yamasaki, S.; Saito, T.; Ohba, Y.; et al. Induction of colonic regulatory T cells by indigenous Clostridium species. Science 2011, 331, 337–341. [Google Scholar] [CrossRef] [PubMed]
- Ishii, W.; Komine-Aizawa, S.; Takano, C.; Fujita, Y.; Morioka, I.; Hayakawa, S. Relationship between the fecal microbiota and depression and anxiety in pediatric patients with orthostatic intolerance. Prim. Care Companion CNS Disord. 2019, 21, 25523. [Google Scholar] [CrossRef] [PubMed]
- Nagpal, R.; Tsuji, H.; Takahashi, T.; Nomoto, K.; Kawashima, K.; Nagata, S.; Yamashiro, Y. Gut dysbiosis following C-section instigates higher colonisation of toxigenic Clostridium perfringens in infants. Benef. Microbes 2017, 8, 353–365. [Google Scholar] [CrossRef]
- Yadav, J.P.; Kaur, S.; Dhaka, P.; Vijay, D.; Bedi, J.S. Prevalence, molecular characterization, and antimicrobial resistance profile of Clostridium perfringens from India: A scoping review. Anaerobe 2022, 77, 102639. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| By Decade | Reported Cases |
|---|---|
| 1951–1960 | 3 |
| 1961–1970 | 2 |
| 1971–1980 | 2 |
| 1981–1990 | 8 |
| 1991–2000 | 15 |
| 2001–2010 | 30 |
| 2011–2020 | 37 |
| 2021–2022 | 18 |
| Total | 115 |
| Author | Year | Age | Sex | Origin Infection | Positive Culture | Survival | Toxin | Toxinotype | ||
|---|---|---|---|---|---|---|---|---|---|---|
| CPA | CPE | |||||||||
| 1 | Bätge | 1992 | 61 | M | Liver ab-scess | Blood | Yes | NR | NR | |
| 2 | Ifthikaruddin | 1992 | 54 | F | Unknown | Blood | No | NR | NR | |
| 3 | Hübl | 1993 | 84 | F | Intestinal | Blood | No | + | NR | |
| 4 | Rogstad | 1993 | 61 | M | Micro liver abscess | Blood, liver | No | NR | NR | |
| 5 | Clarke | 1994 | 53 | F | Necrotising enteritis | Blood, peritoneal fluid | Yes | NR | NR | |
| 6 | Meyerhoff | 1995 | 66 | F | Unknown | Blood | No | NR | NR | |
| 7 | Gutiérrez | 1995 | 74 | M | Micro liver abscess | Blood | No | NR | NR | |
| 8 | Singh | 1996 | 73 | F | Unknown | Blood | No | NR | NR | |
| 9 | Bush | 1996 | 58 | F | Biliary | Blood | Yes | NR | NR | |
| 10 | Jones | 1996 | 66 | F | Liver abscess | Blood, liver abscess | No | NR | NR | |
| 11 | Pun | 1996 | 47 | M | Cholangitis | Blood | No | NR | NR | |
| 12 | Singer | 1997 | 55 | F | Unknown | Blood | No | NR | NR | |
| 13 | Alvarez | 1999 | 77 | F | Abdominal | Blood | No | NR | NR | |
| 14 | Thomas | 1999 | 73 | M | Cholecystitis | Blood | Yes | NR | NR | |
| 15 | Eckel | 2000 | 65 | F | Liver abscess | Blood | Yes | NR | NR | |
| 16 | Kreidl | 2002 | 80 | M | Liver abscess | Blood, liver abscess | No | NR | NR | |
| 17 | Barrett | 2002 | NR | F | Septic abortion | Blood | No | NR | NR | |
| 18 | Jimenez | 2002 | 79 | M | Unknown | Blood | No | NR | NR | |
| 19 | Halpin | 2002 | 29 | F | Post-caesarean endometritis | Blood | Yes | NR | NR | |
| 20 | Hamoda | 2002 | 39 | F | Post-amniocentesis endometritis | Blood | Yes | NR | NR | |
| 21 | Ikegami | 2004 | 67 | M | Acute pancreatitis | Pancreas | Yes | NR | NR | |
| 22 | Vaiopoulos | 2004 | 74 | M | Intestinal and biliary | Blood | No | NR | NR | |
| 23 | Solis | 2004 | 50 | M | Hepatic gas gangrene | Donor liver | No | NR | NR | |
| 24 | Rodriguez | 2005 | 57 | M | Biliary | Blood | No | NR | NR | |
| 25 | Pirrotta | 2005 | 50 | M | Unknown | Blood, stool | No | NR | NR | |
| 26 | Au | 2005 | 65 | M | Liver abscess | NR | No | NR | NR | |
| 27 | McArthur | 2006 | 49 | M | Abdominal | Blood | No | NR | NR | |
| 28 | Daly | 2006 | 80 | M | Liver abscess | Blood | No | NR | NR | |
| 29 | Kwon | 2006 | 71 | F | Unknown | Blood | No | NR | NR | |
| 30 | Loran | 2006 | 69 | F | Liver abscess | NR | No | NR | NR | |
| 31 | Ohtani | 2006 | 78 | M | Liver abscess | Blood, liver abscess | No | NR | NR | |
| 32 | Eigneberger | 2006 | 60 | M | Liver abscess | Liver (Gram stain) | No | NR | NR | |
| 33 | Poulou | 2007 | 74 | M | Unknown | Blood | No | Lecithinase | NR | |
| 34 | Kapoor | 2007 | 58 | M | Unknown | Blood | No | NR | NR | |
| 35 | Poon | 2007 | 64 | F | Hepatobiliary | Blood | No | NR | NR | |
| 36 | Nadisauskiene | 2008 | 31 | F | Post-caesarean endometritis | Blood | No | NR | NR | |
| 37 | Egyed | 2008 | 39 | F | Unknown | Blood | Yes | NR | NR | |
| 38 | Hess | 2008 | 81 | M | Diverticulitis | Blood, brain, heart, spleen | No | NR | NR | |
| 39 | Boyd | 2009 | 46 | M | Cholecystitis | Blood | No | NR | NR | |
| 40 | Uppal | 2009 | 61 | M | Unknown | Blood | No | NR | NR | |
| 41 | Merino | 2010 | 83 | F | Liver abscess | Blood | No | NR | NR | |
| 42 | Ng | 2010 | 61 | F | Liver abscess | Blood | Yes | NR | NR | |
| 43 | Rajendran | 2010 | 58 | M | Liver abscess | Blood, liver abscess, gall bladder | Yes | NR | NR | |
| 44 | Bunderen | 2010 | 74 | M | Cholangitis | Blood | Yes | NR | NR | |
| 45 | Bryant | 2010 | 60 | F | Uterus | Blood, intrauterine | Yes | NR | NR | |
| 46 | Stroumsa | 2011 | 41 | F | Uterine myoma | Blood | Yes | NR | NR | |
| 47 | Qandeel | 2012 | 59 | M | Liver abscess (post-laparoscopic cholecystectomy) | Blood | Yes | NR | NR | |
| 48 | Watt | 2012 | 52 | M | Pan-enteritis | Blood | Yes | NR | NR | |
| 49 | Law | 2012 | 50 | F | Liver abscess | Blood | No | NR | NR | |
| 50 | Okon | 2013 | 71 | M | Unknown | Blood, CSF | No | NR | NR | |
| 51 | Cécilia | 2013 | 64 | M | Unknown | Blood | No | NR | NR | |
| 52 | Dutton | 2013 | 66 | M | NR | Blood | No | NR | NR | |
| 53 | Kitterer | 2014 | 71 | M | Liver abscess | Blood | No | NR | NR | |
| 54 | Kurasawa | 2014 | 65 | M | Liver abscess | Blood | No | NR | NR | |
| 55 | Renaudon-Smith | 2014 | 37 | M | Liver abscess | Blood | Yes | NR | NR | |
| 56 | Simon | 2014 | 79 | F | Unknown | Blood | No | NR | NR | |
| 57 | Shindo | 2015 | 73 | F | Liver abscess | Liver abscess | No | + | − | A |
| 58 | Khan | 2015 | 77 | M | Cholecystitis, liver abscess | Liver (Gram stain) | No | NR | NR | |
| 59 | Cochrane | 2015 | 65 | F | Emphysematous cholecystitis | Blood | Yes | NR | NR | |
| 60 | Yamaguchi | 2015 | 80–89 | F | Unknown | Bile, pleural effusions | No | NR | NR | |
| 61 | Li | 2015 | 71 | M | Liver abscess (post-TACE) | Blood | Yes | NR | NR | |
| 62 | Medrano-Juarez | 2016 | 32 | M | Unknown | Blood | Yes | NR | NR | |
| 63 | Lim | 2016 | 58 | M | Liver abscess | Blood | No | NR | NR | |
| 64 | Hashiba | 2016 | 82 | M | Liver abscess, emphysematous cholecystitis | Blood | No | + | − | A |
| 65 | Sarvari | 2016 | 76 | F | Emphysematous gastritis | Intestine subcutaneous tissue | No | NR | NR | |
| 66 | Carretero | 2016 | 65 | M | Liver abscess | Blood, liver abscess | Yes | NR | NR | |
| 67 | Kent | 2017 | 74 | F | Enteritis | Blood | No | NR | NR | |
| 68 | Kukul | 2017 | 17 | M | Gastrointestinal tract | Quadratus muscle | No | NR | NR | |
| 69 | Balan | 2017 | 71 | F | Unknown | Blood | No | NR | NR | |
| 70 | Ewing | 2017 | 53 | F | Necrotising fasciitis | Wound | No | NR | NR | |
| 71 | Shibazaki | 2018 | 68 | F | Liver abscess | Blood | No | NR | NR | |
| 72 | Wild | 2018 | 81 | F | Unknown | Blood | No | + | − | A |
| 73 | Gelonch | 2018 | 66 | M | Liver abscess | NR | No | NR | NR | |
| 74 | Gelonch | 2018 | 63 | M | Liver abscess | NR | No | NR | NR | |
| 75 | Uojima | 2019 | 83 | M | Liver abscess (post-TACE) | Liver abscess | No | NR | NR | |
| 76 | Sakaue | 2019 | 76 | M | Liver abscess | Blood | No | + | − | A |
| 77 | Kawakami | 2020 | 83 | M | Pelvic abscess | Blood, intraabdominal samples | No | NR | NR | |
| 78 | Fujikawa | 2020 | 77 | F | Liver abscess | Blood | No | NR | NR | |
| 79 | Chinen | 2020 | 80 | F | Liver abscess | Blood, liver abscess | No | NR | NR | |
| 80 | Smit | 2020 | 61 | M | Liver abscess | Blood | No | NR | NR | |
| 81 | Smit | 2020 | 71 | F | Unknown | Blood | No | NR | NR | |
| 82 | Koubaissi | 2020 | 50 | M | Abdominal | Blood | No | NR | NR | |
| 83 | Poletti | 2021 | 64 | F | Unknown | Blood | No | NR | NR | |
| 84 | Liu | 2021 | 21 | M | Intestine | Blood | Yes | NR | NR | |
| 85 | Liu | 2021 | 42 | M | Intestine | Blood | No | NR | NR | |
| 86 | Fukui | 2021 | 69 | M | Unknown | Blood | No | + | − | A |
| 87 | Olds | 2021 | 85 | F | Liver abscess | Blood | No | NR | NR | |
| 88 | Bibi | 2021 | 77 | M | Cholecystitis (Post ERCP) | Blood | No | NR | NR | |
| 89 | Guo | 2021 | 62 | M | Hepatoma (post-microwave ablation) | Blood | No | NR | NR | |
| 90 | Woittiez | 2021 | 65 | M | Gangrenous cholecystitis | Blood, liver abscess | No | + | − | A |
| 91 | Woittiez | 2021 | 69 | M | Liver meta? (post-microwave ablation) | Blood | No | + | − | A |
| 92 | Takahashi | 2022 | 70 | M | Liver abscess | Blood, liver abscess | Yes | NR | NR | |
| 93 | Wong | 2022 | 80 | M | Liver abscess | Blood | Yes | NR | NR | |
| 94 | Kohya | 2022 | 60 | M | Perforation in ascending colon cancer | Blood | No | + | + | F |
| 95 | Suzaki | 2022 | 69 | M | Enteritis | Blood | No | + | − | A |
| 96 | Suzaki | 2022 | 65 | F | Cholecystitis | Blood | No | + | − | A |
| 97 | Suzaki | 2022 | 68 | F | Ovarian tumour | Blood | No | + | − | A |
| 98 | Suzaki | 2022 | 46 | M | Trauma | Blood | No | + | + | F |
| 99 | Suzaki | 2022 | 72 | M | Unknown | Blood | No | + | − | A |
| 100 | Suzaki | 2022 | 58 | M | Unknown | Blood | No | + | − | A |
| MIH (n = 6) | w/o MIH (n = 54) | ||
|---|---|---|---|
| Median age | 66.5 (46–72 years) | 77.0 (46–72 years) | p = 0.017 |
| Loss of consciousness | 6/6 (100%) | 19/54 (35.2%) | p = 0.004 |
| Severe pain at the onset | 4/6 (66.7%) | 10/54 (18.5%) | p = 0.008 |
| Shock at onset | 3/6 (50%) | 3/54 (5.6%) | p = 0.010 |
| Haematuria | 2/6 (33.3%) | 1/54 (1.9%) | p = 0.024 |
| GAS formation | 3/6 (50%) | 4/54 (7.4%) | p = 0.017 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Suzaki, A.; Hayakawa, S. Clinical and Microbiological Features of Fulminant Haemolysis Caused by Clostridium perfringens Bacteraemia: Unknown Pathogenesis. Microorganisms 2023, 11, 824. https://doi.org/10.3390/microorganisms11040824
Suzaki A, Hayakawa S. Clinical and Microbiological Features of Fulminant Haemolysis Caused by Clostridium perfringens Bacteraemia: Unknown Pathogenesis. Microorganisms. 2023; 11(4):824. https://doi.org/10.3390/microorganisms11040824
Chicago/Turabian StyleSuzaki, Ai, and Satoshi Hayakawa. 2023. "Clinical and Microbiological Features of Fulminant Haemolysis Caused by Clostridium perfringens Bacteraemia: Unknown Pathogenesis" Microorganisms 11, no. 4: 824. https://doi.org/10.3390/microorganisms11040824
APA StyleSuzaki, A., & Hayakawa, S. (2023). Clinical and Microbiological Features of Fulminant Haemolysis Caused by Clostridium perfringens Bacteraemia: Unknown Pathogenesis. Microorganisms, 11(4), 824. https://doi.org/10.3390/microorganisms11040824