
Light Microscopy and Proteomic Patterns of Ovulation in Cervical Mucus

 , and
, and
Abstract
1. Introduction
1.1. Menstrual Cycle and Fertile Window
1.2. Cervical Mucus
1.2.1. Biophysics of the Cervical Mucus
1.2.2. Biochemistry of the Cervical Mucus
1.3. Cervical Mucus: Functions
1.4. Cervical Mucus Proteomics in Fertile Women
2. Materials and Methods
3. Results
3.1. Light Microscope
3.1.1. Identification and Description of the Different Types of CM
- L mucus
- S mucus
- P mucus
- G mucus
3.1.2. Classification of the Samples According to the Phase of the Cycle
3.2. Proteomic Analysis
3.2.1. Proteins Identified
3.2.2. Identification of Peaking Proteins and the Biological Processes in Which They Participate
- Small proline-rich protein 3 (SPRR3), cornifin-B (SPRR1B), keratin, and type II cytoskeletal 6A (KRT6A) are involved in keratinization;
- Cystatin-B (CSTB) and serine protease inhibitor Kazal-type 5 (SPINK5) are protease inhibitors;
- Heat shock protein beta-1 (HSPB1) participates in the biological process of host–virus interaction;
- Immunoglobulin kappa constant (IGKC), immunoglobulin heavy constant gamma 1 (IGHG1), polymeric immunoglobulin receptor (PIGR), and protein S100-A8 (S100A8) participate in immunity;
- Serotransferrin (TF) participates in transport.
3.2.3. Bioinformatic Analysis of Molecular Functions
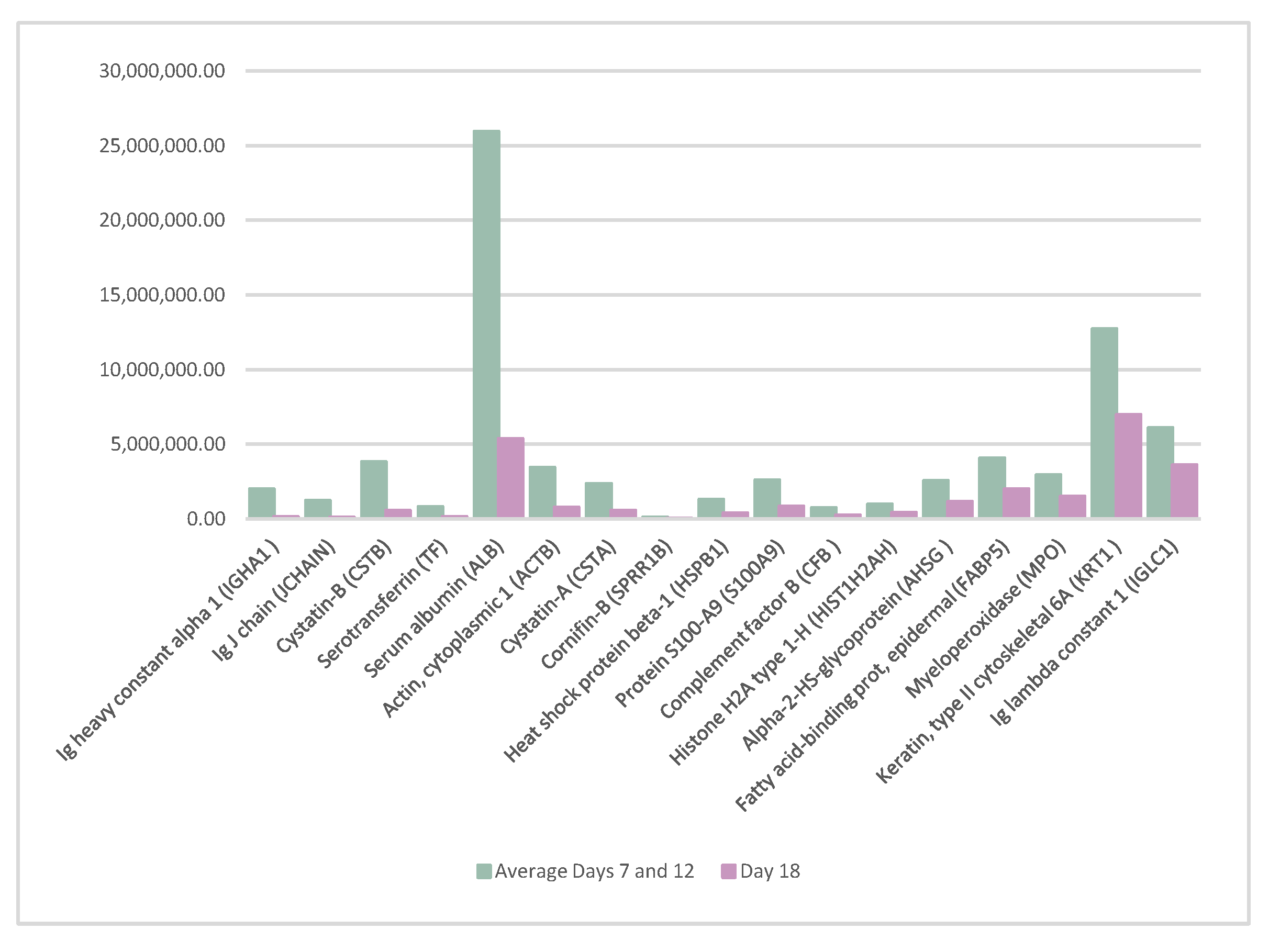
3.2.4. Variability in Protein Abundance
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Stanford, J.B.; White, G.L.; Hatasaka, H. Timing intercourse to achieve pregnancy: Current evidence. Obstet. Gynecol. 2002, 100, 1333–1341. [Google Scholar] [CrossRef] [PubMed]
- Dunson, D.B.; Colombo, B.; Baird, D.D. Changes with age in the level and duration of fertility in the menstrual cycle. Hum. Reprod. 2002, 17, 1399–1403. [Google Scholar] [CrossRef] [PubMed]
- Mihm, M.; Gangooly, S.; Muttukrishna, S. The normal menstrual cycle in women. Anim. Reprod. Sci. 2011, 124, 229–236. [Google Scholar] [CrossRef] [PubMed]
- Colombo, B.; Masarotto, G. Daily Fecundability. Demogr. Res. 2000, 6, 3–5. [Google Scholar] [CrossRef]
- Bigelow, J.L.; Dunson, D.B.; Stanford, J.B.; Ecochard, R.; Gnoth, C.; Colombo, B. Mucus observations in the fertile window: A better predictor of conception than timing of intercourse. Hum. Reprod. 2004, 19, 889–892. [Google Scholar] [CrossRef] [PubMed]
- Wilcox, A.J.; Dunson, D.; Baird, D.D. The timing of the “fertile window” in the menstrual cycle: Day specific estimates from a prospective study. BMJ 2000, 321, 1259–1262. [Google Scholar] [CrossRef]
- Fehring, R.J.; Schneider, M.; Raviele, K. Variability in the phases of the menstrual cycle. J. Obstet. Gynecol. Neonatal Nurs. 2006, 35, 376–384. [Google Scholar] [CrossRef]
- Treloar, A.E.; Boynton, R.E.; Behn, B.G.; Brown, B.W. Variation of the human menstrual cycle through reproductive life. Int. J. Fertil. 1967, 12, 77–126. [Google Scholar] [CrossRef]
- Lenton, E.A.; Landgren, B.; Sexton, L. Normal variation in the length of the luteal phase of the menstrual cycle: Identification of the short luteal phase. BJOG Int. J. Obstet. Gynaecol. 1984, 91, 685–689. [Google Scholar] [CrossRef]
- Bull, J.R.; Rowland, S.P.; Scherwitzl, E.B.; Scherwitzl, R.; Danielsson, K.G.; Harper, J. Real- world menstrual cycle characteristics of more than 600,000 menstrual cycles. NPJ Digit. Med. 2019, 2, 83. [Google Scholar] [CrossRef]
- Chandra, A.; Martinez, G.M.; Mosher, W.D.; Abma, J.C.; Jones, J. Fertility, Family Planning, and Reproductive Health of U.S. Women: Data from the 2002 National Survey of Family Growth; American Psychological Association: Washington, DC, USA, 2005; Volume 23, pp. 1–160. [Google Scholar]
- Costa Figueiredo, M.; Huynh, T.; Takei, A.; Epstein, D.A.; Chen, Y. Goals, life events, and transitions: Examining fertility apps for holistic health tracking. JAMIA Open 2021, 4, ooab013. [Google Scholar] [CrossRef] [PubMed]
- Moghissi, K.S.; Syner, F.N.; Evans, T.N. A composite picture of the menstrual cycle. Am. J. Obstet. Gynecol. 1972, 114, 405–418. [Google Scholar] [CrossRef]
- Wolf, D.P.; Blasco, L.; Khan, M.A.; Litt, M. Human cervical mucus. IV. Viscoelasticity and sperm penetrability during the ovulatory menstrual cycle. Fertil. Steril. 1978, 30, 163–169. [Google Scholar] [CrossRef]
- Morales, P.; Roco, M.; Vigil, P. Human cervical mucus: Relationship between biochemical characteristics and ability to allow migration of spermatozoa. Hum. Reprod. 1993, 8, 78–83. [Google Scholar] [CrossRef] [PubMed]
- Adamopoulos, D.A.; Kapolla, N.; Abrahamian, A.; Dessypris, A.; Nicopoulou, S.; Giannacodemos, G. Sex steroids in cervical mucus of spontaneous or induced ovulatory cycles. Steroids 2000, 65, 1–7. [Google Scholar] [CrossRef]
- Daunter, B. Biochemical and functional structural aspects of human cervical mucus. Scanning Electron Microsc. 1984, 1, 343–358. [Google Scholar]
- Lundberg, O. Abstracts of the 34rd annual meeting of the european society of human reproduction and embryology. Hum. Reprod. 2018, 33, i1–i541. [Google Scholar]
- Elstein, M.; Moghissi, K.S.; Borth, R. Cervical mucus in human reproduction. WHO Colloquium. In Proceedings of the WHO Expanded Programme of Research, Development, and Research Training in Human Reproduction, Geneva, Switzerland, 18–20 September 1972; Scriptor: Copenhagen, Denmark, 1973. [Google Scholar]
- Odeblad, E. Biophysical Techniques of Assessing Cervical Mucus and Microstructure of Cervical Epithelium; World Health Organization Colloquium: Geneva, Switzerland, 1972. [Google Scholar]
- Odeblad, E. Recent research on cervical mucus. In Proceedings of the III International Symposium about Advances in Natural Family Planning, Málaga, Spain; October 1992. [Google Scholar]
- Odeblad, E.; Ingelman-Sundberg, A.; Hallström, L.; Höglund, A.; Lepänen, U.; Lisspers, K.; Perenyi, E.; Rudolfsson-Asberg, K.; Sahlin, K.; Lindström-Sjögren, C.; et al. The biophysical properties of the cervical-vaginal secretions. Int. Rev. Nat. Fam. Plann. 1983, 7, 1–56. [Google Scholar]
- Odeblad, E. Cervical mucus and their functions. J.-Ir. Coll. Physicians Surg. 1997, 26, 27–32. [Google Scholar]
- Menarguez, M.; Pastor, L.M.; Odeblad, E. Morphological characterization of different human cervical mucus types using light and scanning electron microscopy. Hum. Reprod. 2003, 18, 1782–1789. [Google Scholar] [CrossRef]
- Elstein, M. Cervical mucus: Its physiological role and clinical significance. Mucus Health Dis. 1982, 144, 301–318. [Google Scholar]
- Yudin, A.I.; Hanson, F.W.; Katz, D.F. Human cervical mucus and its interaction with sperm: A fine-structural view. Biol. Reprod. 1989, 40, 661–671. [Google Scholar] [CrossRef] [PubMed]
- Gipson, I.K.; Spurr-Michaud, S.; Moccia, R.; Zhan, Q.; Toribara, N.; Ho, S.B.; Gargiulo, A.R.; Hill, J.A., 3rd. MUC4 and MUC5B transcripts are the prevalent mucin messenger ribonucleic acids of the human endocervix. Biol. Reprod. 1999, 60, 58–64. [Google Scholar] [CrossRef] [PubMed]
- Garcea, N.; Moneta, E.; Caruso, A. Cervical mucus. Ann. Ostet. Ginecol. 1970, 92, 701–720. [Google Scholar]
- Kamran, S.; Moghissi, M.D. Cervical Mucus Changes and Ovulation Prediction and Detection. J. Reprod. Med. 1986, 31, 748–753. [Google Scholar]
- Moghissi, K.S.; Syner, F.N.; Borin, B. Cyclic changes of cervical mucus enzymes related to the time of ovulation. I. Alkaline phosphatase. Am. J. Obstet. Gynecol. 1976, 125, 1044–1048. [Google Scholar] [CrossRef]
- Billings, E.L.; Brown, J.B.; Billings, J.J.; Burger, H.G. Symptoms and hormonal changes accompanying ovulation. Lancet 1972, 1, 282–284. [Google Scholar] [CrossRef]
- Thijssen, A.; Meier, A.; Panis, K.; Ombelet, W. Fertility Awareness-Based Methods and subfertility: A systematic review. Facts Views Vis. ObGyn 2014, 6, 113–123. [Google Scholar]
- Curlin, M.; Bursac, D. Cervical mucus: From biochemical structure to clinical implications. Front. Biosci.-Sch. 2013, 5, 507–515. [Google Scholar] [CrossRef]
- Fehring, R.J. Accuracy of the peak day of cervical mucus as a biological marker of fertility. Contraception 2002, 66, 231–235. [Google Scholar] [CrossRef]
- Scarpa, B.; Dunson, D.B. Bayesian selection of predictors of conception probabilities across the menstrual cycle. Paediatr. Périnat. Epidemiol. 2006, 20, 30–37. [Google Scholar] [CrossRef] [PubMed]
- Ryder, B.; Campbell, H. Natural family planning in the 1990s. Lancet 1995, 346, 233–234. [Google Scholar] [CrossRef]
- Billings, E.L.; Westmore, A. The Billings Method, 9th ed.; O’Donovan, A., Ed.; Melbourne Publishing: Melbourne, Australia, 2011. [Google Scholar]
- Parrilla, J.J.; Delgado, J.L. Métodos naturales de regulacioón de la fertilidad: Perspectivas actuales. Ventajas e inconvenientes. Prog. Obstet. Ginecol. 1997, 40, 82–102. [Google Scholar]
- Milardi, D.; Grande, G.; Vincenzoni, F.; Castagnola, M.; Marana, R. Proteomics of human seminal plasma: Identification of biomarker candidates for fertility and infertility and the evolution of technology. Mol. Reprod. Dev. 2013, 80, 350–357. [Google Scholar] [CrossRef]
- Gipson, I.K.; Moccia, R.; Spurrmichaud, S.; Argueso, P.; Gargiulo, A.R.; Hill, J.A.; Offner, G.D.; Keutmann, H.T. The amount of MUC5B mucin in cervical mucus peaks at midcycle. J. Clin. Endocrinol. Metab. 2001, 86, 594–600. [Google Scholar] [CrossRef]
- Thelen, J.J.; Miernyk, J.A. The proteomic future: Where mass spectrometry should be taking us. Biochem. J. 2012, 444, 169–181. [Google Scholar] [CrossRef]
- Issaq, H.J.; Veenstra, T.D.; Conrads, T.P.; Felschow, D. The SELDI-TOF MS approach to proteomics: Protein profiling and biomarker identification. Biochem. Biophys. Res. Commun. 2002, 292, 587–592. [Google Scholar] [CrossRef]
- Panicker, G.; Lee, D.R.; Unger, E.R. Optimization of SELDI-TOF protein profiling for analysis of cervical mucous. J. Proteom. 2009, 71, 637–646. [Google Scholar] [CrossRef]
- Andersch-Björkman, Y.; Thomsson, K.A.; Holmén Larsson, J.M.; Ekerhovd, E.; Hansson, G.C. Large scale identification of proteins, mucins, and their O-glycosylation in the endocervical mucus during the menstrual cycle. Mol. Cell. Proteom. 2007, 6, 708–716. [Google Scholar] [CrossRef]
- Gipson, I.K. Mucins of the human endocervix. Front. Biosci. 2001, 6, D1245–D1255. [Google Scholar] [CrossRef]
- Van Kooij, R.J.; Roelofs, H.J.; Kathmann, G.A.; Kramer, M.F. Human cervical mucus and its mucous glycoprotein during the menstrual cycle. Fertil. Steril. 1980, 34, 226–233. [Google Scholar] [CrossRef]
- Gilks, C.B.; Reid, P.E.; Clement, P.B.; Owen, D.A. Histochemical changes in cervical mucus-secreting epithelium during the normal menstrual cycle. Fertil. Steril. 1989, 51, 286–291. [Google Scholar] [CrossRef]
- Argueso, P.; Spurr-Michaud, S.; Tisdale, A.; Gipson, I.K. Variation in the amount of T antigen and N-acetyllactosamine oligosaccharides in human cervical mucus secretions with the menstrual cycle. J. Clin. Endocrinol. Metab. 2002, 87, 5641–5648. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Iacobelli, S.; Garcea, N.; Angeloni, C. Biochemistry of mucus: A comparative analysis of the secretion from preovulatory, postovulatory, and pregnancy periods. Fertil. Steril. 1971, 22, 727–734. [Google Scholar] [CrossRef]
- Chantler, E.; Debruyne, E. Factors regulating the changes in cervical mucus in different hormonal states. In Mucus in Health and Disease; Springer: Boston, MA, USA, 1977; Volume 89, pp. 131–141. [Google Scholar]
- Shaw, J.L.; Smith, C.R.; Diamandis, E.P. Proteomic analysis of human cervico-vaginal fluid. J. Proteome Res. 2007, 6, 2859–2865. [Google Scholar] [CrossRef]
- Tang, L.J.; De Seta, F.; Odreman, F.; Venge, P.; Piva, C.; Guaschino, S.; Garcia, R.C. Proteomic analysis of human cervical-vaginal fluids. J. Proteome Res. 2007, 6, 2874–2883. [Google Scholar] [CrossRef]
- Panicker, G.; Ye, Y.; Wang, D.; Unger, E.R. Characterization of the Human Cervical Mucous Proteome. Clin. Proteom. 2010, 6, 18–28. [Google Scholar] [CrossRef]
- Casado-Vela, J.; Rodriguez-Suarez, E.; Iloro, I.; Ametzazurra, A.; Alkorta, N.; García- Velasco, J.A.; Matorras, R.; Prieto, B.; González, S.; Nagore, D.; et al. Comprehensive proteomic analysis of human endometrial fluid aspirate. J. Proteome Res. 2009, 8, 4622–4632. [Google Scholar] [CrossRef]
- Lee, D.C.; Hassan, S.S.; Romero, R.; Tarca, A.L.; Bhatti, G.; Gervasi, M.T.; Caruso, J.A.; Stemmer, P.M.; Kim, C.J.; Hansen, L.K.; et al. Protein profiling underscores immunological functions of uterine cervical mucus plug in human pregnancy. J. Proteom. 2011, 74, 817–828. [Google Scholar] [CrossRef]
- Grande, G.; Milardi, D.; Vincenzoni, F.; Pompa, G.; Biscione, A.; Astorri, A.L. Proteomic characterization of the qualitative and quantitative differences in cervical mucus composition during the menstrual cycle. Mol. BioSyst. 2015, 11, 1717–1725. [Google Scholar] [CrossRef]
- Odeblad, E. The discovery of different types of cervical mucus and the Billings Ovulation Method. Bull. Nat. Fam. Plan. Counc. Vic. 1994, 21, 1–35. [Google Scholar]
- Grande, G.; Vincenzoni, F.; Mancini, F.; Baroni, S.; Luca, G.; Calafiore, R.; Marana, R.; Castagnola, M.; Pontecorvi, A.; Milardi, D. Semen Proteomics Reveals the Impact of Enterococcus faecalis on male Fertility. Protein Pept. Lett. 2018, 25, 472–477. [Google Scholar] [CrossRef] [PubMed]
- Odeblad, E. Physical properties of cervical mucus. In Mucus in Health and Disease; Springer: Boston, MA, USA, 1977; Volume 89, pp. 217–225. [Google Scholar]
- King, A.E.; Fleming, D.C.; Critchley, H.O.; Kelly, R.W. Regulation of natural antibiotic expression by inflammatory mediators and mimics of infection in human endometrial epithelial cells. Mol. Hum. Reprod. 2002, 8, 341–349. [Google Scholar] [CrossRef]
- Moriyama, A.; Shimoya, K.; Ogata, I.; Kimura, T.; Nakamura, T.; Wada, H.; Ohashi, K.; Azuma, C.; Saji, F.; Murata, Y. Secretory leukocyte protease inhibitor (SLPI) concentrations in cervical mucus of women with normal menstrual cycle. Mol. Hum. Reprod. 1999, 5, 656–661. [Google Scholar] [CrossRef] [PubMed]
- King, A.E.; Critchley, H.O.; Kelly, R.W. Presence of secretory leukocyte protease inhibitor in human endometrium and first trimester decidua suggests an antibacterial protective role. Mol. Hum. Reprod. 2000, 6, 191–196. [Google Scholar] [CrossRef]
- Eggert-Kruse, W.; Botz, I.; Pohl, S.; Rohr, G.; Strowitzki, T. Antimicrobial activity of human cervical mucus. Hum. Reprod. 2000, 15, 778–784. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Hermida, Y.; Grande, G.; Menarguez, M.; Astorri, A.L.; Azagra, R. Proteomic Markers in Cervical Mucus. Protein Pept. Lett. 2018, 25, 463–471. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Age | ID Number of the Sample (Day. Volunteer ID Number) | Percentage of the Different CM Types in Each Sample | Phase of the Menstrual Cycle |
|---|---|---|---|
| 28 | 1.1 | 100% G | Early Estrogenic |
| 2.1 | 100% G | Early Estrogenic | |
| 3.1 | 100% G+ | Gestagenic | |
| 25 | 1.2 | 100% G | Early Estrogenic |
| 2.2 | 50% G 50% L | Late estrogenic | |
| 3.2 | 85% G 10% L. 5% P | Late Ovulatory | |
| 27 | 1.3 | 100% G | Early Estrogenic |
| 2.3 | 100% G | Early Estrogenic | |
| 3.3 | 100% G | Early Estrogenic | |
| 30 | 1.4 | 100% G | Early Estrogenic |
| 2.4 | 100% G | Early Estrogenic | |
| 3.4 | 80% L 16% S 4% P | Early Ovulatory | |
| 29 | 1.5 | 100% G | Early Estrogenic |
| 2.5 | 90% G 10% L | Early Estrogenic | |
| 3.5 | 23% G 50% L 15% S 2% P | Early Ovulatory | |
| 28 | 1.6 | 100% G | Early Estrogenic |
| 2.6 | 100% G | Early Estrogenic | |
| 3.6 | 80% L 17% S 3% P | Early Ovulatory |
| UniPROT | Gene Name | Molecular Weigth [kDa] | Isoelectric Point | Description |
|---|---|---|---|---|
| P04217 | A1BG | 13.2 | 6.13 | Alpha-1B-glycoprotein |
| P01859 | IGHG2 | 69.3 | 6.28 | Immunoglobulin heavy constant gamma 2 |
| P01860 | IGHG3 | 18.1 | 8.57 | Immunoglobulin heavy constant gamma 3 |
| P01857 | IGHG1 | 14.3 | 8.75 | Immunoglobulin heavy constant gamma 1 |
| P08311 | CTSG | 37.6 | 6.51 | Cathepsin G |
| P02533 | KRT14 | 11.0 | 5.50 | Keratin, type I cytoskeletal 14 |
| P05109 | S100A8 | 16.5 | 9.16 | Protein S100-A8 |
| P01834 | IGKC | 15.2 | 8.68 | Immunoglobulin kappa constant |
| P22528 | SPRR1B | 15.2 | 7.01 | Cornifin-B |
| P01591 | JCHAIN | 22.8 | 6.40 | Immunoglobulin J chain |
| P01876 | IGHA1 | 16.0 | 7.28 | Immunoglobulin heavy constant alpha 1 |
| P02787 | TF | 53.5 | 6.10 | Serotransferrin |
| B3EWG3 | FAM25A | 8.6 | 5.92 | Protein FAM25A |
| P00751 | CFB | 13.9 | 10.32 | Complement factor B |
| P61626 | LYZ | 11.1 | 7.56 | Lysozyme C |
| P02511 | CRYAB | 9.9 | 8.48 | Alpha-crystallin B chain |
| P35321 | SPRR1A | 9.9 | 8.48 | Cornifin-A |
| P04792 | HSPB1 | 11.8 | 6.52 | Heat shock protein beta-1 |
| Q96KK5 | HIST1H2AH | 10.2 | 6.99 | Histone H2A type 1-H |
| P13646 | KRT13 | 10.8 | 7.03 | Keratin, type I cytoskeletal 13 |
| P32926 | DSG3 | 11.3 | 7.87 | Desmoglein-3 |
| P04080 | CSTB | 78.1 | 8.12 | Cystatin-B |
| P01040 | CSTA | 13.0 | 4.84 | Cystatin-A |
| P60709 | ACTB | 9.3 | 6.15 | Actin, cytoplasmic 1 |
| P06702 | S100A9 | 12.3 | 8.82 | Protein S100-A9 |
| Q07654 | TFF3 | 77.0 | 7.12 | Trefoil factor 3 |
| P02765 | AHSG | 41.3 | 7.90 | Alpha-2-HS-glycoprotein |
| P04083 | ANXA1 | 66.0 | 8.12 | Annexin A1 |
| P02788 | LTF | 60.0 | 8.00 | Lactotransferrin |
| P05164 | MPO | 36.1 | 8.19 | Myeloperoxidase |
| P04264 | KRT1 | 41.7 | 5.48 | Keratin, type II cytoskeletal 1 |
| Q14508 | WFDC2 | 28.8 | 11.19 | WAP four-disulfide core domain protein 2 |
| P19957 | PI3 | 62.3 | 7.74 | Elafin |
| Q01469 | FABP5 | 35.9 | 7.59 | Fatty acid-binding protein, epidermal |
| P13647 | KRT5 | 20.1 | 7.33 | Keratin, type II cytoskeletal 5 |
| P01833 | PIGR | 83.8 | 8.97 | Polymeric immunoglobulin receptor |
| P19013 | KRT4 | 39.3 | 5.72 | Keratin, type II cytoskeletal 4 |
| P69905 | HBA1 | 51.5 | 5.16 | Hemoglobin subunit alpha |
| P0CG04 | IGLC1 | 18.1 | 5.24 | Immunoglobulin lambda constant 1 |
| Q9UBG3 | CRNN | 49.6 | 4.96 | Cornulin |
| Q9NQ38 | SPINK5 | 13.9 | 10.89 | Serine protease inhibitor Kazal-type 5 |
| P03973 | SLPI | 57.2 | 6.61 | Antileukoproteinase |
| P68871 | HBB | 54.2 | 5.86 | Hemoglobin subunit beta |
| P23527 | HIST1H2BO | 120.6 | 8.06 | Histone H2B type 1-O |
| P02768 | ALB | 83.2 | 5.74 | Serum albumin |
| P02538 | KRT6A | 38.7 | 7.02 | Keratin, type II cytoskeletal 6A |
| Q9UBC9 | SPRR3 | 85.5 | 7.06 | Small proline-rich protein 3 |
| P59665 | DEFA1 | 107.5 | 5.00 | Neutrophil defensin 1 |
| UniPROT | Gene Name | Description |
|---|---|---|
| P01834 | IGKC | Immunoglobulin kappa constant |
| P01857 | IGHG1 | Immunoglobulin heavy constant gamma 1 |
| P01859 | IGHG2 | Immunoglobulin heavy constant gamma 2 |
| P01860 | IGHG3 | Immunoglobulin heavy constant gamma 3 |
| P08311 | CTSG | Cathepsin G |
| P05109 | S100A8 | Protein S100-A8 |
| P04217 | A1BG | Alpha-1B-glycoprotein |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fernandez-Hermida, Y.; Vincenzoni, F.; Milardi, D.; Astorri, A.L.; Urbani, A.; Grande, G.; Azagra, R. Light Microscopy and Proteomic Patterns of Ovulation in Cervical Mucus. Life 2022, 12, 1815. https://doi.org/10.3390/life12111815
Fernandez-Hermida Y, Vincenzoni F, Milardi D, Astorri AL, Urbani A, Grande G, Azagra R. Light Microscopy and Proteomic Patterns of Ovulation in Cervical Mucus. Life. 2022; 12(11):1815. https://doi.org/10.3390/life12111815
Chicago/Turabian StyleFernandez-Hermida, Yolanda, Federica Vincenzoni, Domenico Milardi, Anna Laura Astorri, Andrea Urbani, Giuseppe Grande, and Rafael Azagra. 2022. "Light Microscopy and Proteomic Patterns of Ovulation in Cervical Mucus" Life 12, no. 11: 1815. https://doi.org/10.3390/life12111815
APA StyleFernandez-Hermida, Y., Vincenzoni, F., Milardi, D., Astorri, A. L., Urbani, A., Grande, G., & Azagra, R. (2022). Light Microscopy and Proteomic Patterns of Ovulation in Cervical Mucus. Life, 12(11), 1815. https://doi.org/10.3390/life12111815