Abstract
Radiotherapy is focused on the tumor but also reaches healthy tissues, causing toxicities that are possibly related to genomic factors. In this context, radiogenomics can help reduce the toxicity, increase the effectiveness of radiotherapy, and personalize treatment. It is important to consider the genomic profiles of populations not yet studied in radiogenomics, such as the indigenous Amazonian population. Thus, our objective was to analyze important genes for radiogenomics, such as ATM, TGFB1, RAD51, AREG, XRCC4, CDK1, MEG3, PRKCE, TANC1, and KDR, in indigenous people and draw a radiogenomic profile of this population. The NextSeq 500® platform was used for sequencing reactions; for differences in the allelic frequency between populations, Fisher’s Exact Test was used. We identified 39 variants, 2 of which were high impact: 1 in KDR (rs41452948) and another in XRCC4 (rs1805377). We found four modifying variants not yet described in the literature in PRKCE. We did not find any variants in TANC1—an important gene for personalized medicine in radiotherapy—that were associated with toxicities in previous cohorts, configuring a protective factor for indigenous people. We identified four SNVs (rs664143, rs1801516, rs1870377, rs1800470) that were associated with toxicity in previous studies. Knowing the radiogenomic profile of indigenous people can help personalize their radiotherapy.
1. Introduction
Radiotherapy (RT) is one of the most important treatments for cancer. It is used in more than half of cancer patients and can have a curative or palliative goal by reducing the rate of local recurrence and improving overall survival. In RT, the tumor tissue is irradiated with ionizing radiation, which can penetrate the adjacent healthy tissue and cause toxic effects there, as the radiation damages the DNA [1,2]. In this sense, the toxic effects, adverse effects caused by the intensity of the dose, lead to losses both for the individual and due to the poor quality of life. Particular attention must be paid to childhood cancers, as these can be long-lasting, with correspondingly high impacts of radiotherapy and on the healthcare system due to the increase in public expenditure [1,3].
Radiotoxicity (RTX) may vary depending on the tissue irradiated [1]. Mucositis, xerostomia, subcutaneous fibrosis, and dysphagia [4] occur with squamous cell carcinoma; esophagitis may occur with radiation to the lung [5]. RT to the chest can cause pain, fibrosis, upper limb edema, pericarditis, and ischemic and valvular heart disease [6]. RT in the prostate can lead to dermatitis, urinary incontinence, and bladder and erectile dysfunction [7].
Radiosensitivity is a tendency to develop adverse effects in tissues under the effects of cell death due to radiation; this generates resistance to treatment and is influenced by biological or genetic factors, for example, tumor heterogeneity [8,9]. It is understood that radiosensitivity driven by genetic variation is one of the fundamental factors for radiotoxicity [10]. Given this, radiogenomics searches for biomarkers of the response to radiotherapy have been used to personalize treatments among individuals undergoing radiotherapy according to their radiogenomic profile [3,10].
RT dosing follows the guidelines of the Radiation Therapy Oncology Group, which has established the same standards for the same tumor sites. However, due to individual differences, some doses may be excessive or insufficient [11]. This scenario may be even more complicated when considering the profile of genetically diverse populations such as indigenous populations, due to their process of geographic isolation and to genetic and evolutionary factors such as genetic drift, founder effects, natural selection, and inbreeding [12].
Therefore, it is pertinent to analyze important genes related to radiogenomics in the Amazonian indigenous population, to obtain information regarding the genomic profile of this population regarding the response to radiotherapy and, in the future, to obtain pertinent results through the control of mutations. Thus, the objective of the study was to analyze the exome of the ATM, TGFB1, RAD51, AREG, XRCC4, CDK1, MEG3, PRKCE, TANC1, and KDR genes in the indigenous population and draw a genomic profile of this population, which has been poorly studied with regard to radiogenomics.
We emphasize that the Brazilian population is one of the most heterogeneous in the world, marked by indigenous, African, and European ancestry; the indigenous population contributes significantly to the formation of Latin American groups, with a share of about 30, especially in the northern region of Brazil [13,14]. In addition, studies conducted with indigenous peoples from other countries have shown high rates of cancer deaths [15]. On the other hand, there have been few genetic studies aiming to understand oncologic mechanisms and therapies that have dealt with this group. We can also point out that there have been no studies evaluating the impact of variants important for radiogenomics on the indigenous population and the population mixed with it.
2. Materials and Methods
2.1. Population Analysis for the Study
The study participants and their ethnic leaders signed the Free and Informed Consent Form. This study was approved by the National Ethics and Research Commission (CONEP) and the Research Ethics Committee of the Center for Tropical Medicine of the Federal University of Pará (UFPA) under CAAE 20654313.6.0000.5172. The population is made up of 64 indigenous people from the Amazon region of Brazil, representing 12 ethnicities: Asurini do Xingu, Arara, Araweté, Asurini do To-cantins, Awagujá, Kayapó/Xikrin, Zo’é, Wajãpi, Karipuna, Phurere, Munduruku, and Ju-runa. The population was healthy and did not undergo radiotherapy. The genetic ancestry of the population group was obtained through a panel of 61 ancestry informative markers (AIMs), which were used to identify individual ancestry and a mixture of three continents (European, African, and Amerindian). The results obtained were associated with other world populations, according to information available in version 3 of the 1000 Genomes database (available online at http://www.1000genomes.org, accessed on 25 October 2023) and the Exome Aggregation Consortium (ExAC). These populations are composed of 661 Africans (AFR), 346 Americans (AMR), 504 East Asians (EAS), 503 Europeans (EUR), and 489 South Asians (SAS).
2.2. DNA Extraction and Exome Analysis
The method used for DNA extraction was phenol–chloroform extraction [16]. Verification of the integrity of the sample was carried out using a NanoDrop 8000 spectrophotometer, and tracing was performed using the Nextera Rapid Capture Exome (Illumina®, San Diego, CA, USA) and SureSelect Human All Exon V6 kits (Agilent, Santa Clara, CA, USA). The NextSeq 500® platform (Illumina®, San Diego, CA, USA) was used for sequencing reactions using the NextSeq 500 High-output v2 300 cycle kit (Illumina®, San Diego, CA, USA).
2.3. Selection of Genes and Variants
The first criterion for variant consideration was 10 minimum coverage values, and then the impact of each variant was considered according to the SNPeff v 4.3 classification (https://pcingola.github.io/SnpEff/; accessed on 20 February 2024). In addition, we considered only variants with a significant allelic frequency in at least three continental populations. As a result of the exome analysis, 143 variants were found. After selection based on the above criteria, 39 variants remained to be followed up in this study.
Ten genes were selected for this study based on their association with radiotherapy efficacy and/or toxicity; some have been included in previous studies and are indexed in databases such as PubMed. The genes are described in Table 1.

Table 1.
The functions of the AREG, ATM, KDR, MEG3, PRKCE, RAD51, TGFB1, TANC1, and XRCC4 genes.
2.4. Bioinformatics Analysis
FASTQ was used to analyze the quality of the reads (FastQCv.0.11—https://www.bioinformatics.babraham.ac.uk/projects/fastqc/; accessed on 20 February 2024), and filters were applied to the samples in order to disregard low-quality reads (fastx_tools v.0.13—http://hannonlab.cshl.edu/fastx_toolset/; accessed on 20 February 2024). The reference genome (GRCH38) was used to map and align the samples, using BWA v.0.7 (http://bio-bwa.sourceforge.net/; accessed on 20 February 2024). The file was indexed and classified according to the alignment generated by the reference genome (SAMtools v.1.2—http://sourceforge.net/projects/samtools/; accessed on 20 February 2024). From there, the alignment was processed to remove PCR duplication (Picard Tools v.1.129—http://broadinstitute.github.io/picard/; accessed on 20 February 2024) and to perform structuring quality readjustment and local realignment (GATK v. 3.2—https://www.broadinstitute.org/gatk/; accessed on 20 February 2024). Then, the results were processed to establish the reference genome variants (GATK v.3.2). ViVa1 (Viewer of Variants) software (v1)—developed by the bioinformatics team at the Federal University of Rio Grande do Norte (UFRN)—was used to analyze the variant annotations. The databases employed for variant annotations were SnpEff v.4.3., Ensembl Variant Effect Predictor (Ensembl version 99), and ClinVar (v.2018-10). For in silico predictions of pathogenicity, SIFT (v.6.2.1), PolyPhen-2 (v.2.2), LRT (November 2009), Mutation Evaluator (v.3.0), and Mutation Tester (v. 2.0) were used. FATHMM (v.2.3), PROVEAN (v.1.1.3), MetaSVM (v1.0), M-CAP (v1.4), and FATHMM-MKL (http://fathmm.biocompute.org.uk/about.html; accessed on 20 February 2024) were also used. More information about bioinformatics analysis is given in the works of Rodrigues et al. [37] and Ribeiro-dos-Santos et al. [38].
2.5. Statistical Analysis
Fisher’s Exact Test was used to determine statistically significant differences (p value ≤ 0.05) between the frequencies of the variants in the world populations (AFR, EUR, AMR, EAS, and SAS). The allele frequencies of the variants found in the study population were determined by allele enumeration. Comparisons between the allele frequencies of the variants found in the indigenous population and in the five world populations were performed using Fisher’s Exact Test. To avoid discrepancies due to different population sizes, we leveled all variant frequencies with the size of the indigenous population. The frequencies of the five populations were taken from Phase 3 of the 1000 Genomes Project (http://www.1000genomes.org; accessed on 20 February 2024) and the Exome Aggregation Consortium (ExAC). These allele frequencies were also used to calculate the dissimilarity matrix and to generate the multidimensional scaling (MDS). The analyses were performed with RStudio v. 3.5.1.
3. Results
From the analyses, we identified 39 variants distributed across the 9 genes: 1 belonging to the AREG gene, 6 to the ATM gene, 9 to the KDR gene, 4 to the MEG3 gene, 10 to the PRKCE gene, 3 to the RAD51 gene, 1 to the TGFB1 gene, 4 to the TANC1 gene, and 1 to the XRCC4 gene. Figure 1 presents the distribution of variants according to their high, modifier, or moderate impact; thus, 2 were classified as high-impact, 28 as modifiers, and 5 as moderate.
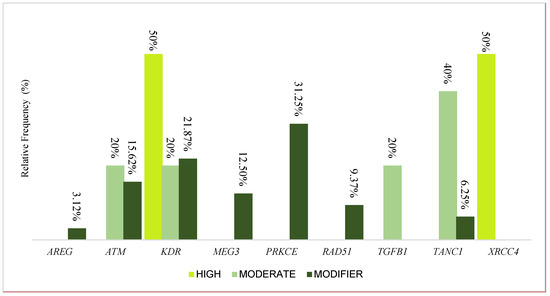
Figure 1.
The relative contributions of variants discriminated according to high, modifier, or moderate impact in the AREG, ATM, KDR, MEG3, PRKCE, RAD51, TGFB1, TANC1, and XRCC4 genes.
Two high-impact variants were identified, distributed in the KDR and XRCC4 genes; for the modifying variants, 3.12% (1) were in the AREG gene, 15.62% (5) in ATM, 21.87% (7) in KDR, 12.50% (4) in MEG3, 31.25% (10) in PRKCE, 9.37% (3) in RAD51, and 6.25% (2) in TANC1. Regarding the moderate variants, ATM, KDR, and TGFB1 each had one moderate variant, while two were in the TANC1 gene. The variants are described in Table 2 with information about the ID, the impact predicted by SNPeff software v 4.3, the region, the type, the nucleotide change, and the frequency presented in world populations. The low-impact variants are presented in Supplementary Table S1.
Table 2.
Descriptions of the variants in the AREG, ATM, KDR, MEG3, PRKCE, RAD51, TGFB1, TANC1, and XRCC4 genes according to their impact, region, variant type, and change in nucleotide.
As for the variants not yet described in the literature, four were found, all in the PRKCE gene on chromosome 2; these are listed in Table 3 and are characterized by a single nucleotide change and an insertion/deletion. The variants are sequentially located at 45652078, 45652096, 45652087, and 45652092. Of these variants, three had a frequency of 0.0833 in the indigenous population and one had a frequency of 0.01677. We intend to conduct further studies to further explore the effects of these variants on protein activity to obtain additional information on radiogenomics in a poorly studied population.
Table 3.
Description of new variants found in the indigenous population from the Brazilian Amazon in genes relevant to radiotherapy.
Of the variants described in Table 2, rs41452948 and rs1805377 are of high impact, the first occurring in the KDR gene and the second in the XRCC4 gene, the first on chromosome 4 and the second on chromosome 5. Both are represented by a change in nucleotide from G to A. These variants showed a significant allelic frequency in the indigenous population compared to the AFR, AMR, EUR, and SAS populations and a high, modifying, and moderate impact, as shown in Table 3 Comparisons with low-impact variants can be found in Supplementary Table S2.
In addition, 43 variants showed a nonzero allelic frequency in at least three global populations. However, the differences in frequency in the five continental populations compared to the indigenous population were significant for only 18 of these alleles, as described in Table 4.
Table 4.
Comparison between the allelic frequencies in Indigenous Amazonian (INDG) and continental populations (AFR, AMR, EAS, EUR, and SAS) described in the 1000 Genomes and the Exome Aggregation Consortium (ExAC).
Figure 2 shows the different genotypes in the global population and the indigenous population on a multidimensional scale (MDS) determined using Fisher’s Exact Test. Looking at the genes selected for this exome, our results show that the indigenous population (INDG) has a genetic profile that differs significantly from those of the five world populations, especially those of East Asians (EAS) and Africans (AFR).
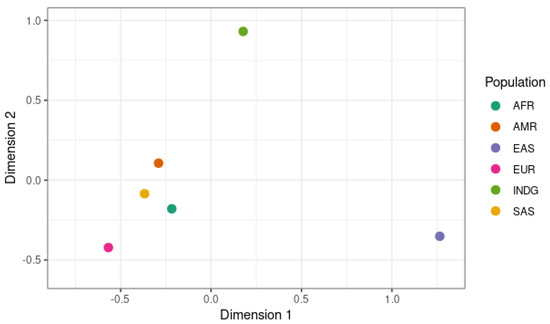
Figure 2.
Differences in allele frequencies of the variants studied in the continental populations and indigenous population, plotted on an MDS.
4. Discussion
The curative potential of radiotherapy is limited by the intrinsic radioresistance of tumor cells, which is related to the heterogeneity of the tumor and the surrounding microenvironment, as well as to various genetic alterations [39]. In addition to radioresistance, patients may experience clinically significant side effects, namely, acute reactions such as erythema, follicular reaction, pruritus, moist or dry desquamation, ulceration, and necrosis, or delayed reactions such as tissue fibrosis, atrophy, firm subcutaneous tissue, and subcutaneous swelling [40,41].
GWA studies are often used to identify genetic variants associated with complex and multifactorial diseases [42,43]. GWA studies linking radiogenomics to radiotherapy toxicity show that genetic variants, treatment variables, and other clinical factors are independent predictors of radiotoxicity. This contributes to the suggestion that common variants may improve traditional models of the likelihood of complications in normal tissues [44,45]. When DNA is exposed to radiation, DNA double-strand breaks occur, which favors a response to this damage, such as cellular radioresistance [46,47]. This was the first exome study to examine genes associated with the response to radiotherapy in indigenous populations, a group that is usually underrepresented in genetic studies, especially in precision medicine [48].
In our results, we found 39 variants with nonzero allele frequencies in the study population, 4 of which have never been found in other genetic databases and none of which are in the CDK1 gene. Therefore, of the 35 variants present in all five populations, we highlight two high-impact variants, rs41452948 in the KDR gene and rs1805377 in the XRCC4 gene, as presenting statically significant differences when comparing the indigenous population with African, American, European, and South Asian populations, as shown in Table 4.
We emphasize that many variants were identified in intronic regions, where they were believed not to encompass gene functions. However, intronic regions play an important role in genome regulation. For example, they influence the maturation of mRNA and transcriptional regulation through alternative splicing, which can directly influence a gene’s response to radiotoxicity [49]. The rs1805377 in the splice site acceptor + intron could have consequences such as disruption of the normal splicing process, leading to important changes in parts of the final mRNA messenger gene, which, in turn, could cause genetic disorders or disease, depending on the gene affected and the specific type of variant [50].
The variants found only in the indigenous population are all located in the PRKCE gene, which is described in the literature as a gene associated with the response to radiation [27]. Members of the PKC family phosphorylate a variety of protein targets and are involved in cell signaling pathways [26]. These variants have a modifying clinical effect; three of the variants had an allelic frequency of 0.0833 in indigenous people and one had a frequency of 0.1667. These mutations with a modifying effect could be potential markers for indigenous populations in the Amazon region. However, we emphasize the intention to conduct further studies to consolidate the knowledge of these potential new variants.
Polymorphisms in the RAD51 gene have been associated in previous studies with symptoms of heart failure and the appearance of a new primary tumor in patients with breast cancer treated with trastuzumab and RT [6], suggesting an important gene for RTX; however, such polymorphisms were not detected in the population studied. Other studies have also investigated variations in DNA repair genes—such as ATM, RAD51, and XRCC4—that cause differences in the response to radiation between individuals. These differences are characterized by increased toxicity, which normally emanates from irradiated tissue [51], but these variants were also not detected in the Indigenous population, suggesting protection from these toxicities.
We discovered four variants in the TANC1 gene, two with a moderate impact (rs34588551, rs4664277) and two with a modifying impact in the intronic region (rs34344829, rs146371641). In our analyses, we found a statistically significant difference regarding rs146371641 in all five world populations compared to the indigenous group (Table 4), which is related to the fact that TANC1 is an important gene for radiogenomics; this could show that the indigenous population has a different profile in the use of radiotherapy.
TANC1 plays a crucial role in the recruitment of fusion-capable myoblasts during myotube formation. In addition, expression of this gene has been found in various tissues such as adipose tissue and the adrenal cortex [3,31]. TANC1 was associated with late radiotherapy toxicity in a large genomic study cohort with a European population. The results suggested that this gene is involved in muscle damage processes induced by radiation. However, in our study, the variants presented in the European study were not found, which may be explained by the genomic difference between the population originating from Brazil and the European population [37]. The influencing variant rs664143, located in the ATM intron, showed significant differences in the five world populations. This variant showed a worse prognosis in patients with squamous cell carcinoma of the esophagus and non-small cell lung cancer after radiotherapy. In addition, the A allele was associated with shorter survival, while the G allele indicated an increased risk of disease progression [52].
The rs1801516 variant, which has a moderate impact on ATM, showed significant differences in the indigenous population compared to the American, European, and South Asian populations (see Table 4). This variant has been studied with regard to the effects after radiotherapy, with results suggesting that individuals with the minor allele have an increased risk of developing late fibrosis after radiotherapy [53]. In studies by McDuff et al. [54], Andreassen et al. [55], and Kerns et al. [45], the variant was associated with the risk of general toxicity, acute toxicity, late toxicity, acute skin toxicity, acute rectal toxicity, telangiectasia, and fibrosis.
The variant with a moderate impact on the KDR gene, rs1870377, showed a significant difference, especially when comparing the indigenous and European populations (Table 4). A study by Tinhofer et al. [56] showed that despite the poorer prognosis of patients with squamous cell carcinoma of the head and neck, the A > T allele of germline variant rs1870377 was associated with longer survival in risk groups of patients with stage IV squamous cell carcinoma of the oropharynx and hypopharynx who underwent chemotherapy. On the other hand, Butkiewicz et al. [57] analyzed 422 patients with squamous cell carcinoma of the head and neck undergoing radiotherapy and concluded that VEGFR2 rs1870377 TT is a significant borderline risk factor for lower local recurrence and an independent predictor of poor prognosis.
When analyzing the toxicity of the rs1870377 variant in patients undergoing 5-fluorouracil-based chemotherapy for locally advanced rectal cancer, the VEGFR2 H472Q Q/Q genotype was associated with a higher risk of grade 3 mucositis of the proximal upper gastrointestinal tract in arm 2 of the study (induction and concomitant prolonged intravenous infusion of 5-FU with radiotherapy). However, in arm 1 (bolus of 5-FU followed by prolonged intravenous infusion of 5-FU with radiotherapy), this genotype was associated with a lower risk of mucositis [22].
The variant with a moderate influence on the TGFB1 gene, rs1800470, showed statistically significant differences between the five global populations and the indigenous population (Table 4). According to Xiao Y et al. [58], the T allele of variant rs1800470 (CT/TT) is associated with the risk of developing radiation-induced pneumonia in patients with esophageal squamous cell carcinoma. In patients with oropharyngeal squamous cell carcinoma undergoing radiotherapy, the presence of the rs1800470 variant of TGFβ1 may reduce and modify the risk of death and recurrence [59].
Previous research also found mitigating differences between the genetic profile of indigenous peoples and those of the other five world populations examined here [60,61,62]. However, the physical and geographic isolation of these peoples contributes to a gap in epidemiologic and genetic information. Therefore, knowledge of their molecular profile may be of importance to indigenous populations, as well as those intermixed with them [63].
In the European world, the applications of radiogenomics concepts are more advanced, considering studies such as those by O’Sullivan, NJ; Kelly [64], which used models to predict metastases after neoadjuvant radiotherapy and showed how these concepts can be used to identify low-risk patients in order not to subject them to interventions such as tumor dissection. In addition, radiogenomics has also been used to predict genetic mutations in colorectal cancer to optimize the outcome of radiotherapy and enable targeted therapy [65].
5. Conclusions
This study investigated the presence of genetic variants involved in the process of radiotherapy toxicity in an indigenous Amazonian population. No variants already associated with late RTX were found. We found two high-impact variants and four variants that have not yet been described in the other five world populations examined and may be exclusive to this population. Future research on therapeutic approaches targeting indigenous and admixed populations is critical.
Personalized medicine research has gained much attention for its role in targeting therapies and making predictions. This can help doctors plan more appropriate treatment regimens, reduce radiation damage in high-risk patients, and optimize outcomes in low-risk patients.
Supplementary Materials
The following support information can be downloaded from: https://www.mdpi.com/article/10.3390/jpm14050484/s1. Table S1: Descriptions of low-impact AREG, ATM, KDR, MEG3, PRKCE, RAD51, TGFB1, TANC1, and XRCC4 gene variants, as well as allele frequencies in continental populations (African (AFR), American (AMR), East Asian (EAS), European (EUR), and South Asian (SAS)) described in the 1000 genome database. Table S2: Paired comparisons (p-values) with significant results of allele frequencies in the Amazonian indigenous population (INDG) and in the continental populations (African (AFR), American (AMR), East Asian (EAS), European (EUR), and South Asian (SAS)) described in the 1000 genome database.
Author Contributions
Conceptualization: M.C.d.L. and N.P.C.d.S. methodology: N.M., J.C.G.R. and N.P.C.d.S.; formal analysis: N.M. and J.C.G.R.; writing—original draft preparation: M.C.d.L., C.C.d.C., K.E.C.A., G.G.d.C.N. and A.C.A.d.C.; writing—review and editing: M.C.d.L. and N.P.C.d.S.; supervision: J.F.G., Â.R.-d.-S., P.P.d.A., R.M.R.B., M.R.F., S.E.B.d.S. and N.P.C.d.S.; project administration: N.P.C.d.S. and J.F.G.; resources: Â.R.-d.-S., P.P.d.A. and S.E.B.d.S.; validation: R.M.R.B. and M.R.F. All authors have read and agreed to the published version of the manuscript.
Funding
Pró-Reitoria de Pesquisa e Pós-Graduação (PROPESP) and the Federal University of Pará (UFPA).
Institutional Review Board Statement
This study was conducted according to the guidelines of the Declaration of Helsinki and approved by the Research Ethics Committee of the Health Sciences Institute of the Federal University of Pará and by the National Research Ethics Committee (CONEP), identification number 20654313.6.0000.5172, approved on 2 March 2015.
Informed Consent Statement
Informed consent was obtained from all subjects involved in this study.
Data Availability Statement
Data are contained within the article and Supplementary Materials.
Acknowledgments
We acknowledge Universidade Federal do Pará (UFPA), the Oncology Research Center (NPO/UFPA), and the Human and Medical Genetics Laboratory (LGHM/UFPA).
Conflicts of Interest
The authors declare no conflicts of interest.
References
- De Ruysscher, D.; Niedermann, G.; Burnet, N.G.; Siva, S.; Lee, A.W.; Hegi-Johnson, F. Radiotherapy toxicity. Nat. Rev. Dis. Primers 2019, 5, 13. [Google Scholar] [CrossRef] [PubMed]
- Early Breast Cancer Trialists’ Collaborative Group (EBCTCG); Darby, S.; McGale, P.; Correa, C.; Taylor, C.; Arriagada, R.; Clarke, M.; Cutter, D.; Davies, C.; Ewertz, M.; et al. Effect of radiotherapy after breast-conserving surgery on recurrence at 10 years and death from breast cancer at 15 years: Meta-analysis of individual patient data for 10,801 women in 17 randomized trials. Lancet 2011, 378, 1707–1716. [Google Scholar] [PubMed]
- Rosenstein, B.S. Radiogenomics: Identification of Genomic Predictors for Radiation Toxicity. Semin. Radiat. Oncol. 2017, 27, 300–309. [Google Scholar] [CrossRef]
- Thiagarajan, A.; Iyer, N.G. Genomics of radiation sensitivity in squamous cell carcinomas. Pharmacogenomics 2019, 20, 457–466. [Google Scholar] [CrossRef]
- Holgersson, G.; Bergström, S.; Liv, P.; Nilsson, J.; Edlund, P.; Blomberg, C.; Nyman, J.; Friesland, S.; Ekman, S.; Asklund, T.; et al. Effect of Increased Radiotoxicity on Survival of Patients with Non-small Cell Lung Cancer Treated with Curatively Intended Radiotherapy. Anticancer Res. 2015, 35, 5491–5497. [Google Scholar] [PubMed]
- Goričar, K.; Dugar, F.; Dolžan, V.; Marinko, T. NBN, RAD51, and XRCC3 Polymorphisms as Potential Predictive Biomarkers of Adjuvant Radiotherapy Toxicity in Early HER2-Positive Breast Cancer. Cancers 2022, 14, 4365. [Google Scholar] [CrossRef] [PubMed]
- Mališić, E.; Petrović, N.; Brengues, M.; Azria, D.; Matić, I.Z.; Srbljak Ćuk, I.; Kopčalić, K.; Stanojković, T.; Nikitović, M. Association of polymorphisms in TGFB1, XRCC1, XRCC3 genes and CD8 T-lymphocyte apoptosis with adverse effect of radiotherapy for prostate cancer. Sci. Rep. 2022, 12, 21306. [Google Scholar] [CrossRef] [PubMed]
- El-Nachef, L.; Al-Choboq, J.; Restier-Verlet, J.; Granzotto, A.; Berthel, E.; Sonzogni, L.; Ferlazzo, M.L.; Bouchet, A.; Leblond, P.; Combemale, P.; et al. Human radiosensitivity and radiosusceptibility: What are the differences? Int. J. Mol. Sci. 2021, 22, 7158. [Google Scholar] [CrossRef] [PubMed]
- Guhlich, M.; Hubert, L.; Mergler, C.P.N.; Rave-Fraenk, M.; Dröge, L.H.; Leu, M.; Schmidberger, H.; Rieken, S.; Hille, A.; Schirmer, M.A. Identification of Risk Loci for Radiotoxicity in Prostate Cancer by Comprehensive Genotyping of TGFB1 and TGFBR1. Cancers 2021, 13, 5585. [Google Scholar] [CrossRef]
- Jiang, M.; Yang, J.; Li, K.; Liu, J.; Jing, X.; Tang, M. Insights into the theranostic value of precision medicine on advanced radiotherapy to breast cancer. Int. J. Med. Sci. 2021, 18, 626–638. [Google Scholar] [CrossRef]
- Bai, X.; Wang, B.; Wang, S.; Wu, Z.; Gou, C.; Hou, Q. Radiotherapy dose distribution prediction for breast cancer using deformable image registration. BioMedical Engineering OnLine 2020, 19, 39. [Google Scholar] [CrossRef] [PubMed]
- Amorim, C.E.G.; Wang, S.; Marrero, A.R.; Salzano, F.M.; Ruiz-Linares, A.; Bortolini, M.C. X-Chromosomal Genetic Diversity and Linkage Disequilibrium Patterns in Amerindians and Non-Amerindian Populations. Am. J. Hum. Biol. 2011, 23, 299–304. [Google Scholar] [CrossRef] [PubMed]
- BRAZIL. Ministry of Indigenous Peoples. Data from the 2022 Census Reveals that Brazil has 1.7 Million Indigenous People. 2023. Available online: https://en.wikipedia.org/wiki/Ministry_of_Native_People_(Brazil) (accessed on 20 February 2024).
- Rodrigues, J.C.G.; Fernandes, M.R.; Guerreiro, J.F.; da Silva, A.L.D.C.; Ribeiro-Dos-Santos, Â.; Santos, S.; Santos, N.P.C.D. Polymorphisms of ADME-related genes and their implications for drug safety and efficacy in Amazonian Amerindians. Sci. Rep. 2019, 9, 7201. [Google Scholar] [CrossRef]
- Moore, S.P.; Forman, D.; Piñeros, M.; Fernández, S.M.; de Oliveira Santos, M.; Bray, F. Cancer in indigenous people in Latin America and the Caribbean: A review. Cancer Med. 2014, 3, 70–80. [Google Scholar] [CrossRef] [PubMed]
- Green, M.R.; Sambrook, J. Isolation of high-molecular-weight DNA using organic solvents. Cold Spring Harb. Protoc. 2017, 2017, pdb-prot093450. [Google Scholar] [CrossRef] [PubMed]
- Bouchet, A.; Sakakini, N.; Atifi, M.E.; Le Clec’h, C.; Bräuer-Krisch, E.; Rogalev, L.; Laissue, J.A.; Rihet, P.; Le Duc, G.; Pelletier, L. Identification of AREG and PLK1 pathway modulation as a potential key of the response of intracranial 9L tumor to microbeam radiation therapy. Int. J. Cancer 2015, 136, 2705–2716. [Google Scholar] [CrossRef] [PubMed]
- García, M.E.G.; Kirsch, D.G.; Reitman, Z.J. Targeting the ATM Kinase to Enhance the Efficacy of Radiotherapy and Outcomes for Cancer Patients. Semin. Radiat. Oncol. 2022, 32, 3–14. [Google Scholar] [CrossRef]
- Liu, J.; Tang, X.; Shi, F.; Li, C.; Zhang, K.; Liu, J.; Wang, G.; Yin, J.; Li, Z. Genetic polymorphism contributes to 131I radiotherapy-induced toxicities in patients with differentiated thyroid cancer. Pharmacogenomics 2018, 19, 1335–1344. [Google Scholar] [CrossRef]
- Massacci, G.; Perfetto, L.; Sacco, F. The Cyclin-dependent kinase 1: More than a cell cycle regulator. Br. J. Cancer 2023, 129, 1707–1716. [Google Scholar] [CrossRef]
- Huang, R.X.; Zhou, P.K. DNA damage response signaling pathways and targets for radiotherapy sensitization in cancer. Signal Transduct. Target. Ther. 2020, 5, 60. [Google Scholar] [CrossRef]
- Zaman, N.; Dass, S.S.; Du Parcq, P.; Macmahon, S.; Gallagher, L.; Thompson, L.; Khorashad, J.S.; LimbÄck-Stanic, C. The KDR (VEGFR-2) Genetic Polymorphism Q472H and c-KIT Polymorphism M541L Are Associated with More Aggressive Behaviour in Astrocytic Gliomas. Cancer Genom. Proteom. 2020, 17, 715–727. [Google Scholar] [CrossRef]
- Bohanes, P.; Rankin, C.J.; Blanke, C.D.; Winder, T.; Ulrich, C.M.; Smalley, S.R.; Rich, T.A.; Martensen, J.A.; Benson, A.B., III; Mayer, R.J.; et al. Pharmacogenetic Analysis of INT 0144 Trial: Association of Polymorphisms with Survival and Toxicity in Rectal Cancer Patients Treated with 5-FU and Radiation. Clin Cancer Res. 2015, 21, 1583–1590. [Google Scholar] [CrossRef][Green Version]
- Ashrafizadeh, M.; Gholami, M.H.; Mirzaei, S.; Zabolian, A.; Haddadi, A.; Farahani, M.V.; Kashani, S.H.; Hushmandi, K.; Najafi, M.; Zarrabi, A.; et al. Dual relationship between long non-coding RNAs and STAT3 signaling in different cancers: New insight to proliferation and metastasis. Life Sci. 2021, 270, 119006. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Guo, Z.; Zhao, Y.; Jin, Y.; An, L.; Wu, B.; Liu, Z.; Chen, X.; Chen, X.; Zhou, H.; et al. Genetic polymorphisms of genes in the lncRNA-p53 regulatory network are associated with concomitant toxicities and efficacy of chemoradiotherapy in patients with nasopharyngeal carcinoma. Sci. Rep. 2017, 7, 8320. [Google Scholar] [CrossRef] [PubMed]
- Gene. Bethesda (MD): National Library of Medicine (USA), National Center for Biotechnology Information; 2004. Available online: https://www.ncbi.nlm.nih.gov/gene/5581#summary (accessed on 27 February 2024).
- Pu, X.; Wang, L.; Chang, J.Y.; Hildebrandt, M.A.; Ye, Y.; Lu, C.; Skinner, H.D.; Niu, N.; Jenkins, G.D.; Komaki, R.; et al. Inflammation-related genetic variants predict toxicity following definitive radiotherapy for lung cancer. Clin. Pharmacol. Ther. 2014, 96, 609–615. [Google Scholar] [CrossRef]
- Greenhough, L.A.; Liang, C.C.; Belan, O.; Kunzelmann, S.; Maslen, S.; Rodrigo-Brenni, M.C.; Anand, R.; Skehel, M.; Boulton, S.J.; West, S.C. Structure and function of the RAD51B-RAD51C-RAD51D-XRCC2 tumor suppressor. Nature 2023, 619, 650–657. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Zhang, Y.; Shi, Y.; Liu, S. Comprehensive Analysis of Prognostic and Immune Infiltrates for RAD51 in Human Breast Cancer. Crit. Rev. Eukaryot. Gene Expr. 2021, 31, 71–79. [Google Scholar] [CrossRef]
- Xu, H.; Xiong, C.; Chen, Y.; Zhang, C.; Bai, D. Identification of RAD51 as a prognostic biomarker correlated with immune infiltration in hepatocellular carcinoma. Bioengineered 2021, 12, 2664–2675. [Google Scholar] [CrossRef]
- Data for This Paper Were Retrieved from the Alliance of Genome Resources. Version 6.0.0. Available online: https://www.alliancegenome.org (accessed on 20 February 2024).
- Fachal, L.; Gómez-Caamaño, A.; Barnett, G.C.; Peleteiro, P.; Carballo, A.M.; Calvo-Crespo, P.; Kerns, S.L.; Sánchez-García, M.; Lobato-Busto, R.; Dorling, L.; et al. A three-stage genome-wide association study identifies a susceptibility locus for late radiotherapy toxicity at 2q24.1. Nat. Genet. 2014, 46, 891–894. [Google Scholar] [CrossRef]
- Aula, H.; Skyttä, T.; Tuohinen, S.; Luukkaala, T.; Hämäläinen, M.; Virtanen, V.; Raatikainen, P.; Moilanen, E.; Kellokumpu-Lehtinen, P.L. Decreases in TGF-β1 and PDGF levels are associated with echocardiographic changes during adjuvant radiotherapy for breast cancer. Radiat. Oncol. 2018, 13, 201. [Google Scholar] [CrossRef]
- Delgado, B.D.; Enguix-Riego, M.V.; de Bobadilla, J.C.F.; Rivera, D.H.; Gómez, J.M.N.G.; Praena-Fernández, J.M.; Del Campo, E.R.; Gordillo, M.J.O.; Fernandez, M.D.C.F.; Guerra, J.L.L. Association of single nucleotide polymorphisms at HSPB1 rs7459185 and TGFB1 rs11466353 with radiation esophagitis in lung cancer. Radiother. Oncol. 2019, 135, 161–169. [Google Scholar] [CrossRef]
- Talibova, G.; Bilmez, Y.; Ozturk, S. Increased double-strand breaks in aged mouse male germ cells may result from altered expression of genes essential for homologous recombination or nonhomologous end-joining repair. Histochem. Cell Biol. 2023, 159, 127–147. [Google Scholar] [CrossRef]
- Wen, Y.; Dai, G.; Wang, L.; Fu, K.; Zuo, S. Silencing of XRCC4 increases the radiosensitivity of triple-negative breast cancer cells. Biosci. Rep. 2019, 39, BSR20180893. [Google Scholar] [CrossRef]
- Rodrigues, J.C.G.; de Souza, T.P.; Pastana, L.F.; dos Santos, A.M.R.; Fernandes, M.R.; Pinto, P.; Wanderley, A.V.; de Souza, S.J.; Kroll, J.E.; Pereira, A.L.; et al. Identification of NUDT15 gene variant in Amazonian Amerindians and admixed individuals from northern Brazil. PLoS ONE 2020, 15, e0231651. [Google Scholar] [CrossRef]
- Ribeiro-dos-Santos, A.M.; Vidal, A.F.; Vinasco-Sandoval, T.; Guerreiro, J.; Santos, S.; Ribeiro-dos-Santos, Â.; De Souza, S.J. Exome Sequencing of Native Populations from the Amazon Reveals Patterns on the Peopling of South America. Front. Genet. 2020, 11, 548507. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Song, Y.; Wang, R.; Wang, T. Molecular mechanisms of tumor resistance to radiotherapy. Mol. Cancer 2023, 22, 96. [Google Scholar] [CrossRef]
- Lee, E.; Eum, S.Y.; Slifer, S.H.; Martin, E.R.; Takita, C.; Wright, J.L.; Hines, R.B.; Hu, J.J. Association between polymorphisms in DNA damage repair genes and radiation therapy–induced early adverse skin reactions in a breast cancer population: A polygenic risk score approach. Int. J. Radiat. Oncol. Biol. Phys. 2020, 106, 948–957. [Google Scholar] [CrossRef] [PubMed]
- Jandu, H.K.; Veal, C.D.; Fachal, L.; Luccarini, C.; Aguado-Barrera, M.E.; Altabas, M.; Azria, D.; Baten, A.; Bourgier, C.; Bultijnck, R.; et al. Genome-wide association study of treatment-related toxicity two years following radiotherapy for breast cancer. Radiother. Oncol. 2023, 187, 109806. [Google Scholar] [CrossRef] [PubMed]
- Ko, D.C.; Urban, T.J. Understanding Human Variation in Susceptibility to Infectious Diseases through Clinical and Cellular GWAS. PLoS Pathog. 2013, 9, e1003424. [Google Scholar] [CrossRef]
- Nikpay, M.; Goel, A.; Won, H.H.; Hall, L.M.; Willenborg, C.; Kanoni, S.; Saleheen, D.; Kyriakou, T.; Nelson, C.P.; Hopewell, J.C.; et al. A comprehensive genome-wide association meta-analysis of 1000 coronary artery disease genomes. Nat. Genet. 2015, 47, 1121–1130. [Google Scholar]
- El Naqa, I.; Pandey, G.; Aerts, H.; Chien, J.T.; Andreassen, C.N.; Niemierko, A.; Ten Haken, R.K. Radiotherapy outcome models in the era of radiomics and radiogenomics: Uncertainties and validation. Int. J. Radiat. Oncol. Biol. Phys. 2018, 102, 1070–1073. [Google Scholar] [CrossRef]
- Kerns, S.L.; Fachal, L.; Dorling, L.; Barnett, G.C.; Baran, A.; Peterson, D.R.; Hollenberg, M.; Hao, K.; Narzo, A.D.; Ahsen, M.E.; et al. Radiogenomics Consortium Genome-Wide Association Study Meta-Analysis of Late Toxicity After Prostate Cancer Radiotherapy. J. Natl. Cancer Inst. 2020, 112, 179–190. [Google Scholar] [CrossRef]
- Ranjha, L.; Howard, S.M.; Cejka, P. Main steps in DNA double-strand break repair: An introduction to homologous recombination and related processes. Chromosoma 2018, 127, 187–214. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Wu, Q.; Zhu, L.; Xie, S.; Tu, L.; Yang, Y.; Wu, K.; Zhao, Y.; Wang, Y.; Xu, Y.; et al. SERPINE2/PN-1 regulates the DNA damage response and radioresistance by activating ATM in lung cancer. Cancer Lett. 2022, 524, 268–283. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, M.R.; Rodrigues, J.C.G.; Maroñas, O.; Latorre-Pellicer, A.; Cruz, R.; Guerreiro, J.F.; Burbano, R.M.R.; Assumpção, P.P.D.; Ribeiro-dos-Santos, A.; Santos, S.E.B.D.; et al. Genetic Diversity of Drug-Related Genes in Native Americans of the Brazilian Amazon. Pharmacogenom. Pers. Med. 2021, 14, 117–133. [Google Scholar] [CrossRef] [PubMed]
- Chorev, M.; Carmel, L. The function of introns. Front. Genet. 2012, 3, 55. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Chaldebas, M.; Ogishi, M.; Al Qureshah, F.; Ponsin, K.; Feng, Y.; Rinchai, D.; Milisavljevic, B.; Han, J.E.; Moncada-Vélez, M.; et al. Genome-wide detection of human intronic AG-gain variants located between splicing branch points and canonical splice acceptor sites. Proc. Natl. Acad. Sci. USA 2023, 120, e2314225120. [Google Scholar] [CrossRef]
- Mumbrekar, K.D.; Goutham, H.V.; Vadhiraja, B.M.; Bola Sadashiva, S.R. Polymorphisms in double-strand break repair related genes influence radiosensitivity phenotype in lymphocytes from healthy individuals. DNA Repair 2016, 40, 27–34. [Google Scholar] [CrossRef] [PubMed]
- Du, Z.; Zhang, W.; Zhou, Y.; Yu, D.; Chen, X.; Chang, J.; Qiao, Y.; Zhang, M.; Huang, Y.; Wu, C.; et al. Associations of ATM Polymorphisms with Survival in Advanced Esophageal Squamous Cell Carcinoma Patients Receiving Radiation Therapy. Int. J. Radiat. Oncol. Biol. Phys. 2015, 93, 181–189. [Google Scholar] [CrossRef]
- Zhang, Y.; Liu, Z.; Wang, M.; Tian, H.; Su, K.; Cui, J.; Dong, L.; Han, F. Single Nucleotide Polymorphism rs1801516 in Ataxia Telangiectasia-Mutated Gene Predicts Late Fibrosis in Cancer Patients after Radiotherapy: A PRISMA-Compliant Systematic Review and Meta-Analysis. Medicine 2016, 95, e3267. [Google Scholar] [CrossRef]
- McDuff, S.G.R.; Bellon, J.R.; Shannon, K.M.; Gadd, M.A.; Dunn, S.; Rosenstein, B.S.; Ho, A.Y. ATM Variants in Breast Cancer: Implications for Breast Radiation Therapy Treatment Recommendations. Int. J. Radiat. Oncol. Biol. Phys. 2021, 110, 1373–1382. [Google Scholar] [CrossRef] [PubMed]
- Andreassen, C.N.; Rosenstein, B.S.; Kerns, S.L.; Ostrer, H.; De Ruysscher, D.; Cesaretti, J.A.; Barnett, G.C.; Dunning, A.M.; Dorling, L.; West, C.M.; et al. International Radiogenomics Consortium. Individual patient data meta-analysis shows a significant association between the ATM rs1801516 SNP and toxicity after radiotherapy in 5456 breast and prostate cancer patients. Radiother. Oncol. 2016, 121, 431–439. [Google Scholar] [CrossRef] [PubMed]
- Tinhofer, I.; Stenzinger, A.; Eder, T.; Konschak, R.; Niehr, F.; Endris, V.; Distel, L.; Hautmann, M.G.; Mandic, R.; Stromberger, C.; et al. Targeted next-generation sequencing identifies molecular subgroups in squamous cell carcinoma of the head and neck with distinct outcomes after concurrent chemoradiation. Ann. Oncol. 2016, 27, 2262–2268. [Google Scholar] [CrossRef] [PubMed]
- Butkiewicz, D.; Gdowicz-Kłosok, A.; Krześniak, M.; Rutkowski, T.; Krzywon, A.; Cortez, A.J.; Domińczyk, I.; Składowski, K. Association of Genetic Variants in ANGPT/TEK and VEGF/VEGFR with Progression and Survival in Head and Neck Squamous Cell Carcinoma Treated with Radiotherapy or Radiochemotherapy. Cancers 2020, 12, 1506. [Google Scholar] [CrossRef]
- Xiao, Y.; Yuan, X.; Qiu, H.; Li, Q. Single-nucleotide polymorphisms of TGFβ1 and ATM associated with radiation-induced pneumonitis: A prospective cohort study of thoracic cancer patients in China. Int. J. Clin. Exp. Med. 2015, 8, 16403–16413. [Google Scholar] [PubMed]
- Niu, Z.; Sun, P.; Liu, H.; Wei, P.; Wu, J.; Huang, Z.; Gross, N.D.; Shete, S.; Wei, Q.; Zafereo, M.E.; et al. Functional Genetic Variants in TGFβ1 and TGFβR1 in miRNA-Binding Sites Predict Outcomes in Patients with HPV-positive Oropharyngeal Squamous Cell Carcinoma. Clin. Cancer Res. 2023, 29, 3081–3091. [Google Scholar] [CrossRef] [PubMed]
- Pastana, L.F.; Silva, T.A.; Gellen, L.P.A.; Vieira, G.M.; de Assunção, L.A.; Leitão, L.P.C.; da Silva, N.M.; Coelho, R.D.C.C.; de Alcântara, A.L.; Vinagre, L.W.M.S.; et al. The Genomic Profile Associated with Risk of Severe Forms of COVID-19 in Amazonian Native American Populations. J. Pers. Med. 2022, 12, 554. [Google Scholar] [CrossRef]
- Aguiar, K.E.C.; Oliveira, I.D.S.; Cohen-Paes, A.D.N.; Coelho, R.D.C.C.; Vinagre, L.W.M.S.; Rodrigues, J.C.G.; Ribeiro-Dos-Santos, A.M.; De Souza, S.J.; Ribeiro-Dos-Santos, Â.; Guerreiro, J.F.; et al. Molecular Profile of Variants in CDH1, TP53, PSCA, PRKAA1 and TTN Genes Related to Gastric Cancer Susceptibility in Amazonian Indigenous Population. J. Pers. Med. 2023, 13, 1364. [Google Scholar] [CrossRef]
- Coelho, R.d.C.C.; Martins, C.L.e.L.P.; Pastana, L.F.; Rodrigues, J.C.G.; Aguiar, K.E.C.; Cohen-Paes, A.d.N.; Gellen, L.P.A.; Moraes, F.C.A.d.; Calderaro, M.C.L.; de Assunção, L.A.; et al. Molecular Profile of Variants Potentially Associated with Severe Forms of COVID-19 in Amazonian Indigenous Populations. Viruses 2024, 16, 359. [Google Scholar] [CrossRef]
- Cohen-Paes, A.; de Alcântara, A.L.; de Souza Menezes, E.; Moreira, F.C.; Fernandes, M.R.; Guerreiro, J.F.; Ribeiro-Dos-Santos, Â.; Dos Santos, S.E.B.; Santos, N.P.C.D. Characterization of DNA Polymerase Genes in Amazonian Amerindian Populations. Genes 2022, 14, 53. [Google Scholar] [CrossRef]
- O’Sullivan, N.J.; Kelly, M.E. Radioomics and Radiogenomics in Pelvic Oncology: Current Applications and Future Directions. Curr. Oncol. 2023, 30, 4936–4945. [Google Scholar] [CrossRef] [PubMed]
- Badic, B.; Tixier, F.; Cheze Le Rest, C.; Hatt, M.; Visvikis, D. Radiogenomics in Colorectal Cancer. Cancers 2021, 13, 973. [Google Scholar] [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).