

A Systematic Review on Attenuation of PCSK9 in Relation to Atherogenesis Biomarkers Associated with Natural Products or Plant Bioactive Compounds in In Vitro Studies: A Critique on the Quality and Imprecision of Studies


 , , and
, , and
Abstract
1. Introduction
2. Methods
2.1. Definitions
2.2. Search Criteria
2.3. Inclusion and Exclusion Criteria
2.4. Study Identification and Selection
2.5. Data Synthesis
2.6. Quality Assessment
3. Results
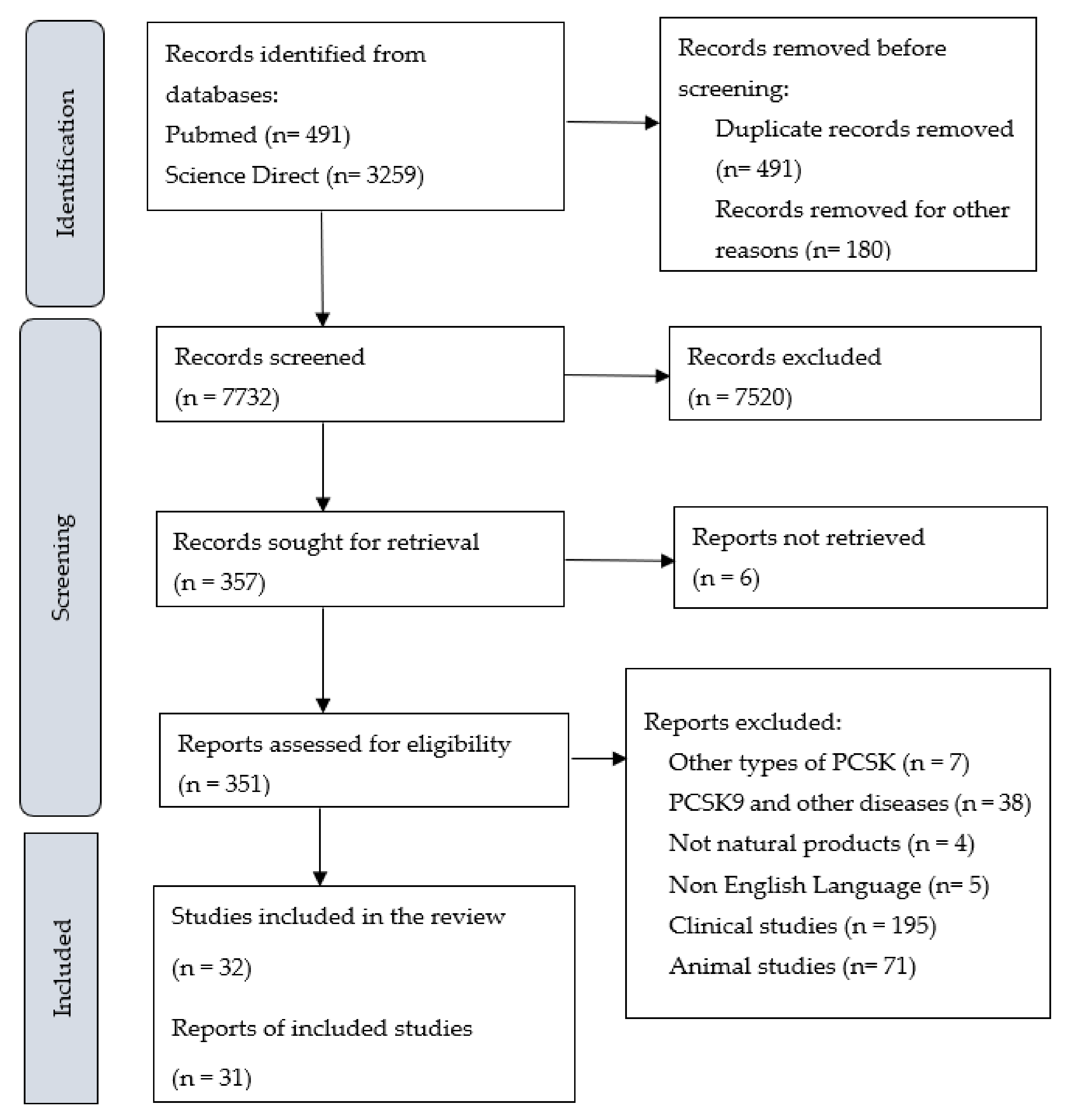
3.1. Literature Searches and Inclusion Assessment
3.2. PCSK9 in Relation to Atherogenesis Biomarkers
3.2.1. In Vitro Models
3.2.2. Protein and Gene Expression of PCSK9 In Vitro Models
PCSK9 in Relation to LDLR, SREBP, and HNF1α Biomarkers
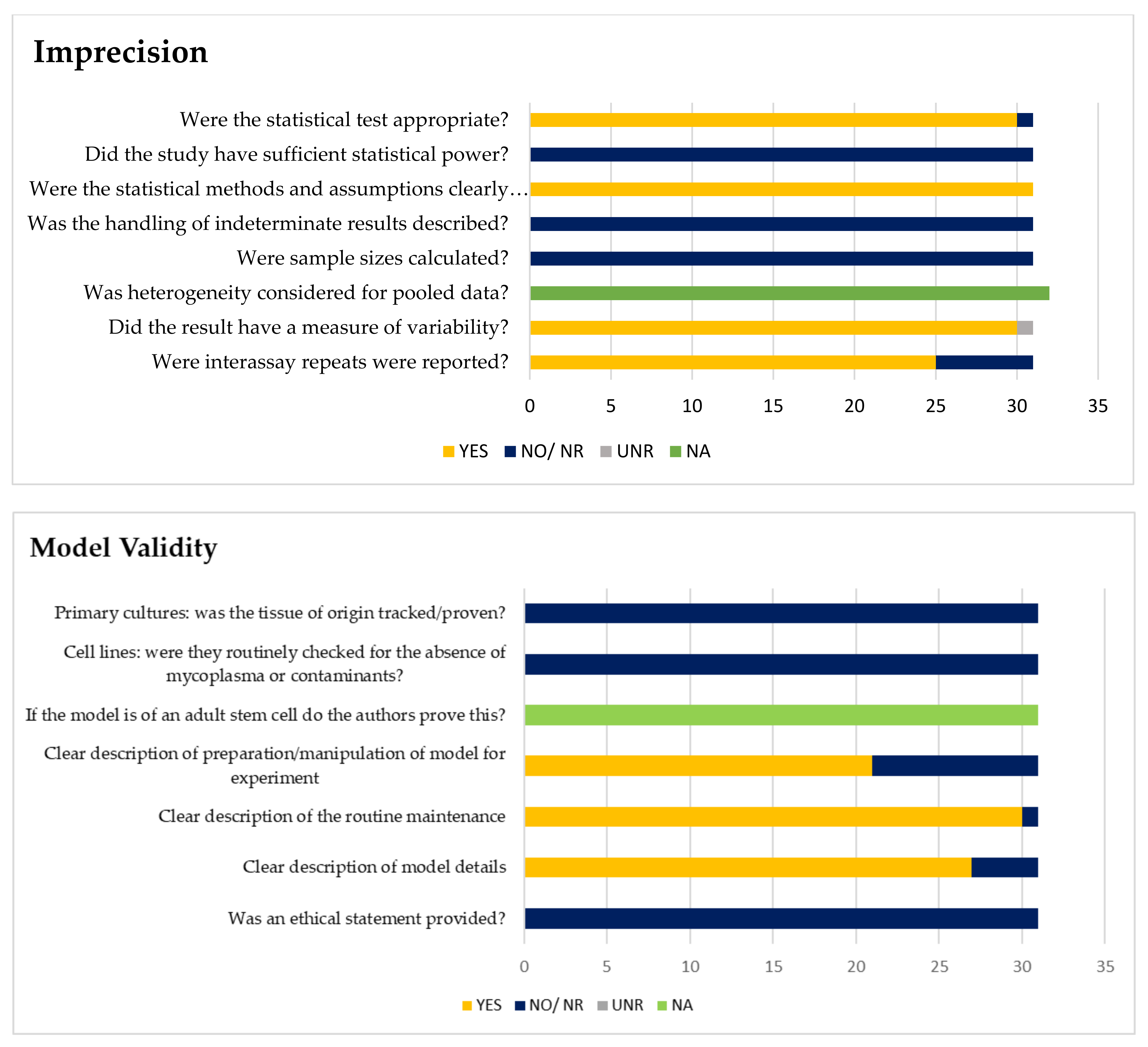
3.2.3. Imprecision and Validity Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Appendix A

Appendix B

{kind=link}
{kind=link}
{kind=link}
| Search | Query |
|---|---|
| PubMed (k = 491) | |
| #4 | (#1 AND #2 AND #3 AND #4) |
| #3 | Cell OR Cells OR Endothelial cell OR Endothelial cells [tiab] |
| #2 | PCSK9 Inhibition [tiab] |
| #1 | Proprotein Convertase Subtilisin Kexin 9 Inhibitor*[tiab] OR PCSK9 Inhibitor*[tiab] |
| Science Direct (k = 3259) | |
| #3 | (#1 AND #2 AND #3) |
| #2 | Topic: Cell* OR Endothelial Cell* |
| #1 | Topic: PCSK9 Inhibitor* OR PCSK9 Inhibition* |
| Scopus (k = 4653) | |
| #3 | (#1 AND #2 AND #3 AND #4) |
| #2 | Topic: PCSK9 Inhibitor* AND (Endothelial Cell*) |
| #1 | Topic: (PCSK9 Inhibitor* AND Cell* |
Appendix C
| Signalling Question | Notes | Answer | |
|---|---|---|---|
| TECHNICAL REPORTING | 1. How many technical repeats were performed per experiment? | Intra-assay variability | Free text |
| 2. Observer variability: Did the experiment give the same result when repeated? | Inter-assay variability | Free text | |
| 3. Is it clear whether the technical repeat is true or a combination of technical and observer variability? | Yes/no/not applicable/unclear or not reported | ||
| 4. Did the result include a measure of variability? Or was the data presented as a scatter plot? | EB = error bars (unclear error), SE = standard error, SEM = standard error of the mean, SD = standard deviation CI = confidence intervals | Free text | |
| 5. Did the authors pool data between experiments? If so, was heterogeneity measured to test that pooling was appropriate? | (Important when using multiple patient/animal samples) | Yes/no/not applicable/unclear or not reported | |
| Overall reporting rating | Low = no concern for bias. Unclear = insufficient data to make a judgment. High risk = there is a concern of high risk. If 1,2 and 4 are fulfilled this can be given a low rating for this review. | Low/Unclear/High | |
| SAMPLE SIZE | 6. Were sample sizes calculated? | For the given experiment/effect did the authors calculate the number of repeats that would be required for significance? | Yes/no/not applicable/unclear or not reported |
| 7. How were indeterminate results, missing results, and outliers handled? | Free text | ||
| 8. Did the study have sufficient statistical power? | Yes: clearly meets the sample size. Likely: >10 repeats with inter-assay repeats. Unclear: >10 repeats, no inter-assay repeats. No: ≤10 technical repeats. | Yes/no/unclear/likely | |
| Did the study have sufficient statistical power? Justification | Based on questions 6-8 | Free text | |
| Overall sample size rating | Low = no concern (or likely statistical power or Unclear statistical power plus variability reported). Unclear: not enough information to make judgement and no high risk for 6-8. High risk: there is a concern of high risk for 6-8. | Low/Unclear/High | |
| STATISTICAL TEST | 9. Description of statistical methods and assumptions. | P S TT = Paired student t-test; US TT = unpaired student t test; x2-test = XT; Fishers exact test = FET; others possible | Free text |
| 10. Were the statistical tests appropriate? | In this review t-tests were the predominant test (other statistical tests are possible). A paired tt-testis the most appropriate test for comparisons between the same cell lines or non-human models, because these are assumed to be homogeneous populations. An unpaired t test should be used for comparisons between primary cultures, human tissues, or different mutants or strains, because these will be heterogeneous populations. | Yes/no/not applicable/unclear or not reported | |
| Were the statistical tests appropriate? Justification | Free text | ||
| 11. Evidence of data dredging | https://en.wikipedia.org/wiki/Data_dredging (accessed on 31 August 2022). | Yes/no/not applicable/unclear or not reported | |
| Statistical test rating | Low = no concern. Unclear = not enough information to make judgement. High risk = there is a concern of high risk | Low/Unclear/High | |
| Other Concerns | Free text |
| First Author Surname and Year | Experiment | 1. | 2. | 3. | 4. | 5. | Reporting Rating | 6. | 7. | 8. | Sample Size Rating | 9. | 10. | 11. | Statistical Test Rating | Overall Rating | Justification |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| HUVECs (Human Umbilical Vein Endothelial Cells) | |||||||||||||||||
| Wang et al. (2019) | Ginkgolide B | 3 | NR | NR | SEM | NA | Low | NA | NR | NO | UNR | S TT | YES | UNR | Low | Low | The observer variability was not reported. The sample size rating is unclear. It is not applicable because it is not a common practice to calculate sample size in cell culture studies. |
| Huh7 (Human Hepatocytes) | |||||||||||||||||
| Wang et al. (2020) | Ascorbic acid | 5 | NR | NR | SE | NA | Low | NA | NR | NO | UNR | P TT | YES | UNR | Low | Low | The technical repeats were high. The observer variability was not reported. Sample size rating is unclear. It is not applicable because it is not a common practice to calculate sample size in cell culture studies. |
| Mbikay et al. (2014) | Quercetin-3-glucoside | 3 | NR | NR | SEM | NA | Low | NA | NR | NO | UNR | S TT | YES | UNR | Low | Low | The observer variability was not reported. Sample size rating is unclear. It is not applicable because it is not a common practice to calculate sample size in cell culture studies. |
| LO2 (hepatocytes) | |||||||||||||||||
| Jing et al. (2019) | Resveratrol | 3 | NR | NR | SEM | NA | Low | NA | NR | NO | UNR | ANOVA | YES | UNR | Low | Low | The observer variability was not reported. The sample size rating is unclear. It is not applicable because it is not a common practice to calculate sample size in cell culture studies. |
| HepG2 (Human Hepatoma) | |||||||||||||||||
| Fan et al. (2021) | Berberine derivative (9k) | 3 | NR | NR | SEM | NA | Low | NA | NR | NO | UNR | S TT | YES | UNR | Low | Low | The observer variability was not reported. One domain was reported unclear. Sample size rating is unclear. It is not applicable because it is not a common practice to calculate sample size in cell culture studies. The observer variability was not reported. One domain was reported unclear. Sample size rating is unclear. It is not applicable because it is not a common practice to calculate sample size in cell culture studies. |
| Masagalli et al. (2021) | Moracin C and Its Derivatives with a 2-arylbenzofuran Motif (Compound 7) | 3 | NR | NR | SEM | NA | Low | NA | NR | NO | UNR | Dunnet TT | YES | UNR | Low | Low | |
| Wang et al. (2020) | Gypenoside LVI | 3 | NR | NR | SD | NA | Low | NA | NR | NO | UNR | S TT | YES | UNR | Low | Low | |
| Fu et al. (2020) | 17β-estradiol (βE2) | 3 | NR | NR | SEM | NA | Low | NA | NR | NO | UNR | Duncan T | YES | UNR | Low | Low | |
| Hwang et al. (2020) | Butein | 3 | NR | NR | SD | NA | Low | NA | NR | NO | UNR | S TT | YES | UNR | Low | Low | |
| Kim et al. (2020) | Piceatannol | 3 | NR | NR | SD | NA | Low | NA | NR | NO | UNR | S TT | YES | UNR | Low | Low | |
| Li et al. (2020) | 23,24- Dihydrocucurbitacin B | 3 | NR | NR | SD | NA | Low | NA | NR | NO | UNR | S TT | YES | UNR | Low | Low | |
| Ahn et al. (2019) | Sophora tonkinensis (erybraedin D-compound 16) | 3 | NR | NR | SEM | NA | Low | NA | NR | NO | UNR | D TT | YES | UNR | Low | Low | |
| Dong et al. (2019) | Siblinin A | 3 | NR | NR | SD | NA | Low | NA | NR | NO | UNR | Post Hoc T | YES | UNR | Low | Low | |
| Lammi et al. (2019) | Lupin peptide T9 (GQEQSHQDEGVIVR) | 4 | NR | NR | SD | NA | Low | NA | NR | NO | UNR | Dunnet T | YES | UNR | Low | Low | |
| Lupo et al. (2019) | red yeast rice RYR (monacolin K), | 3 | NR | NR | SD | NA | Low | NA | NR | NO | UNR | S TT | YES | UNR | Low | Low | |
| Berberis aristata cortex BCE (Berberine) | 3 | NR | NR | SD | NA | Low | NA | NR | NO | UNR | S TT | YES | UNR | Low | Low | ||
| Morus alba leaves extract MLE (1-deoxynojirimycin) | 3 | NR | NR | SD | NA | Low | NA | NR | NO | UNR | S TT | YES | UNR | Low | Low | ||
| Wu et al. (2019) | tetrahydroprotoberberine derivatives (THPBs) (Compound 22) | 3 | NR | NR | SEM | NA | Low | NA | NR | NO | UNR | S TT | YES | UNR | Low | Low | The observer variability was not reported. One domain was reported unclear. The sample size rating is unclear. It is not applicable because it is not a common practice to calculate sample size in cell culture studies. |
| Chae et al. (2018) | Saucinone | 3 | NR | NR | SEM | NA | Low | NA | NR | NO | UNR | D TT | YES | UNR | Low | Low | |
| Yang et al. (2018) | Liraglutide | 3 | NR | NR | SE | NA | Low | NA | NR | NO | UNR | S TT | YES | UNR | Low | Low | |
| Yang et al. (2018) | Chitosan oligosaccharides | 3 | NR | NR | SD | NA | Low | NA | NR | NO | UNR | S TT | YES | UNR | Low | Low | |
| Gu et al. (2017) | Lunasin | 3 | NR | NR | SEM | NA | Low | NA | NR | NO | UNR | ANOVA | YES | UNR | Low | Low | |
| Chen et al. (2016) | Salvia miltiorrhiza Bunge (Tanshinone IIA) | 3 | NR | NR | SD | NA | Low | NA | NR | NO | UNR | D TT | YES | UNR | Low | Low | |
| Cameron et al. (2008) | Berberine | 3 | NR | NR | SEM | NA | Low | NA | NR | NO | UNR | P TT | YES | UNR | Low | Low | |
| Gao et al. (2018) | Pinostrobin | 3 | YES | NR | U | NA | Low | NA | NR | NO | UNR | Post Hoc T (Dunnet) | YES | UNR | Low | Low | The only article that reported on observer variability was reported. The measurement of variability is not clear. One domain was reported unclear. The sample size rating is unclear. It is not applicable because it is not a common practice to calculate sample size in cell culture studies. |
| Hwang et al. (2021) | Capsella Bursa-Pastoris | NR | NR | NR | SD | NA | UNR | NA | NR | NO | UNR | S TT | YES | UNR | Low | UNR | Two domains were reported as unclear. The number of technical repeats was not mentioned in the article. The observer variability was not reported. Sample size rating is unclear. It is not applicable because it is not a common practice to calculate sample size in cell culture studies. |
| Weng et al. (2021) | Gynostemma pentaphyllum [dammarane-type glycosides (2, 3, 15)] | NR | NR | NR | SD | NA | UNR | NA | NR | NO | UNR | ANOVA | YES | UNR | Low | UNR | |
| Pel et al. (2020) | Chromolaena odorata – involve many extraction & many compounds (Compound 6) | NR | NR | NR | SEM | NA | UNR | NA | NR | NO | UNR | Dunnet TT | YES | UNR | Low | UNR | |
| Choi et al. (2017) | Welsh onion (Allium fistulosum L. [family Amaryllidaceae]) | NR | NR | NR | SD | NA | UNR | NA | NR | NO | UNR | S TT | YES | UNR | Low | UNR | |
| Choi et al. (2017) | Welsh onion (Allium fistulosum L. [family Amaryllidaceae]) | NR | NR | NR | SD | NA | UNR | NA | NR | NO | UNR | S TT | YES | UNR | Low | UNR | |
| Pel et al. (2017) | Schisandra chinensis (Turcz.) (Compound 10) | NR | NR | NR | SEM | NA | UNR | NA | NR | NO | UNR | Dunnet TT | YES | UNR | Low | UNR | |
| Aggrey et al. (2019) | Nauclea latifolia (Compound 5) | 3 | NR | NR | SEM | NA | Low | NA | NR | NO | UNR | NR | NO | UNR | UNR | UNR | The observer variability was not reported. Two domains were reported unclear. The sample size rating is unclear. It is not applicable because it is not a common practice to calculate sample size in cell culture studies. The statistical test rating was reported unclear because no analysis was reported on the comparison. |
| JLM3 (hepatocellular carcinoma cells) | |||||||||||||||||
| He et al. (2017) | Actinidia chinensis Planch root extract | 3 | NR | NR | SD | NA | Low | NA | NR | NO | UNR | S TT | YES | UNR | Low | Low | The observer variability was not reported. The sample size rating is unclear. It is not applicable because it is not a common practice to calculate sample size in cell culture studies. |
Appendix D
| Signalling Question | Notes | Answer |
|---|---|---|
| 1. Ethical statement | Was an ethical statement provided for animal handling? | Yes/NR—add details to justification |
| 2. Clear description of model details | Brief description of basic model followed by source, species, strain sex, developmental stage, age, passage number, etc.). | Free text |
| 3. Is the model transgenic? | Whether purchased or created. | Yes/no/unclear |
| 4. Clear description of the routine maintenance of the model | Free text | |
| 5. Further preparation of the model for experimentation | Description of how model was manipulated to obtain result: to include preparation for imaging, how daughter or mother organelle were induced to differentiate. This should be used to make it clear how result was derived. | Free text |
| 6. If the model is of an adult stem cell do the authors prove this? | Cells must be capable of dividing and renewing for long periods; undifferentiated; multipotent. | NA/partial/NR/yes/no—add details to justification. |
| 7. Cell lines: were they routinely checked for the absence of mycoplasma or contaminants? | Yes/no/NR | |
| 8. Primary cultures: was the tissue of origin tracked/proven? | Yes/no/NR | |
| 9. Additional comments/concerns | NA/partial/No/yes—add details to justification. | |
| Overall rating/reporting of model. | Low = all domains clearly reported, and there were no additional concerns. Unclear = Any domain was unclear, but not high risk. High risk = there is a concern of high risk. Note that for this review routine maintenance was not essential for low order organisms. | High/Low/Unclear or not reported |
| Justification | Text to justify why model was given unclear or high rating. Additional text for details regarding questions 1, 6-9. | Free text. |
| Study ID | 1. | 2. | 3. | 4. | 5. | 6. | 7. | 8. | 9. | Overall Rating/ Reporting of Model | Justification |
|---|---|---|---|---|---|---|---|---|---|---|---|
| HUVECs (Human Umbilical Vein Endothelial Cells) | |||||||||||
| Wang et al. (2019) | NR | Human Umbilical Vein Endothelial Cells (from American Type Culture Collection; ATCC) | YES | Cells were cultured under standard culture conditions in DMEM containing 10% heat Inactivated FBS, 2 Mm glutamine, and antibiotics (100 U/mL). | To study the impact of oxidatively modified-LDL on various biochemical and molecular parameters, HUVECs were incubated with Ox-LDL (25–100 µg/mL). | NA | NR | NR | NA | Low | No description of its routine maintenance nor check for contaminants |
| Huh7 (Human Hepatocytes) | |||||||||||
| Wang et al. (2020) | NR | Huh7 was obtained from ATCC. | YES | Huh7 cells, the human hepatic cell lines, were cultured in high glucose DMEM containing 10% FBS, 50 mg/mL penicillin and streptomycin, and 2 Mm glutamine. | After reaching, 80% confluence, cells received treatment in medium containing 2% FBS. | NA | NR | NR | NA | Low | No clear description of model nor its routine maintenance. No inducer was given to stimulate the cells. |
| Mbikay et al. (2014) | NR | Huh7 human liver cells were obtained from the Japanese Collection of Research Bioresources | YES | Huh7 cell incubations were carried out at 37 °C in a humidified 5% CO2–95% air atmosphere in DMEM containing 10% FBS for maintenance or LPDS for experiments, and 50 g/mL gentamycin | LPDS was used for experiments, and 50 µg/mL gentamycin; they were incubated overnight and then treated or not with Q3G, or simvastatin, or both, at defined concentrations, and for defined lengths of time. | NA | NR | NR | NR | Low | |
| LO2 (hepatocytes) | |||||||||||
| Jing et al. (2019) | NR | Human L02 hepatocytes were obtained from the Cell Bank of the Chinese Academy of Sciences (Shanghai, China) | YES | LO2 hepatocytes were cultured overnight in DMEM, supplemented with 10% FBS 100 U/mL of penicillin, and 100 μg/mL of streptomycin at 37 °C in a humidified atmosphere of 5% CO2 and 95% air | To induce cellular steatosis, the cells were exposed to a mixture of FFA (oleate: palmitate = 2:1) at a final concentration of 1 mM for 24 h | NA | NR | NR | NR | Low | No description of its routine maintenance nor check for contaminants |
| HepG2 (Human Hepatoma) | |||||||||||
| Fan et al. (2021) | NR | NR | NO | Cells were cultured in Eagle EMEM, supplemented with 10% FBS 1% nonessential amino acids, and 1% sodium pyruvate | The stable pGL4-PCSK9-P transfected HepG2 cells, named as PCSK9p-Luc HepG2 cells and used as PCSK9 transcriptional inhibitor HTS assay, were cultured in MEM supplemented with 10% FBS, 1% nonessential amino acids, 1% sodium pyruvate and additional G418 (700 mg/mL, Invitrogen). Cells were maintained at 37 °C in the presence of 5% CO2. | NA | NR | NR | NR | Low | No description of the origin or source of cell lines. |
| Masagalli et al. (2021) | NR | HepG2 cells were obtained from the Chinese Academy of Cell Resource Center (Xiangf bio, Shanghai, China) | YES | Cells were maintained in low glucose DMEM containing 10% FBS at 37 °C under 5% CO2 atmosphere | During experiment, cells were seeded in corresponding culture vessels, after reaching 50- 60% confluence, culture media were changed to DMEM supplemented with 5% LPDS while the control group changed to fresh 5% FBS. | NA | NR | NR | NR | Low | No description of its routine maintenance nor check for contaminants |
| Wang et al. (2021) | NR | HepG2 cells were obtained from the Chinese Academy of Cell Resource Center (Xiangf bio, Shanghai, China) | YES | Cells were maintained in low glucose DMEM containing 10% FBS at 37 °C under 5% CO2 atmosphere | During the experiment, cells were seeded in corresponding culture vessels, after reaching 50–60% confluence, culture media were changed to DMEM supplemented with 5% LPDS, while the control group changed to fresh 5% FBS. | NA | NR | NR | NR | Low | |
| Fu et al. (2020) | NR | HepG2 cells (ATCC, USA) | YES | Cells were maintained at 37 °C in phenol red-free DMEM supplemented with 10% FBS, 100 IU/mL penicillin, and 100 µg/mL streptomycin | For all assays, the cells were pre-treated with 1µM G15 for 15 min prior to the addition of βE2 to block GPER action. After a series of wash steps with PBS, internalized AF−PCSK9 was directly observed under an inverted fluorescence microscope, and the fluorescence intensity of AF−PCSK9 in isopropyl alcohol was detected by a SpectraMax M5 reader and reported in RFUs. | NA | NR | NR | NR | UNR | |
| Hwang et al. (2020) | NR | HepG2 cells (HB-8065) were purchased from ATCC (Manassas, VA, USA). | YES | The cells were cultured with DMEM high glucose; supplemented with 10% FBS and 1% antibiotic and antimycotic solution in an incubator (37 °C and 5% CO2) | After 24 h, the media were changed to either DMEM supplemented with FBS or delipidated serum (DLPS)17 (day 1). The media were then changed to media supplemented with either FBS or DLPS + butein | NA | NR | NR | NR | Low | No description of the model nor its routine maintenance |
| Kim et al. (2020) | NR | HepG2 cells (HB-8065) were purchased from the American Type Culture Collection (Manassas, VA, USA). | YES | The cells were cultured with high glucose DMEM supplemented with 10% FBS and 1% antibiotics in a humidified atmosphere of 5% CO2 at 37 °C | After reaching ≈50% confluence (day 0), the medium was changed to either DMEM supplemented with FBS or DLPS, andthe next day, the medium was changed to either FBS or DLPS supplemented DMEM with piceatannol alone or in combination with rosuvastatin or simvastatin | NA | NR | NR | NR | Low | |
| Li et al. (2020) | NR | HepG2 cells (catalogue number: HB-8065, ATCC, Manassas, VA, USA) | YES | Cells were maintained in DMEM with 10% FBS and incubated under a humidified atmosphere of 95% O2 and 5% CO2 at 37 °C. The cells were subcultured once every 2 days. | LDL and LPDS were separated from the pooled plasma of healthy volunteers by ultracentrifugation and were then dialyzed in dialysis buffer and phosphate-buffered saline (PBS). After specific treatments, the culture medium was changed to DiI-LDL DMEM (20 μg/mL) or changed to 2% LPDS. | NA | NR | NR | NR | Low | Human plasma was obtained from Shanghai Xuhui Central Hospital, China, after informed consent was obtained and approval was granted by the Ethics Committee. The procedures conformed to the principles outlined in the Declaration of Helsinki Cells within 4–11 passages were used for experiments. |
| Ahn et al. (2019) | NR | HepG2 cell line was obtained from the Korea Research Institute of Bioscience and Biotechnology (South Korea) | YES | Cells were grown in EMEM, supplemented with 10% FBS and 100 U/mL penicillin/streptomycin sulfate. Cells were incubated in a humidified incubator at 37 °C in a 5% CO2 atmosphere | NR | NA | NR | NR | NR | UNR | No description of the model nor its routine maintenance |
| Dong et al. (2019) | NR | HepG2 cells were obtained from the ATCC | YES | Cells were cultured in DMEM supplemented with 10% FBS and 1% penicillin/streptomycin solution. All cells were incubated in a cell culture chamber at 37 °C under a humidified atmosphere with 5% CO2. | NR | NA | NR | NR | NR | UNR | |
| Lammi et al. (2019) | NR | The HepG2 cell line was bought from ATCC (HB-8065, ATCC from LGC Standards, Milan, Italy) | YES | The HepG2 cell line was cultured in DMEM high-glucose with stable L-glutamine supplemented with 10% FBS, 100 U/mL penicillin, and 100 μg/mL streptomycin (complete growth medium) and incubated at 37 °C under 5% CO2 atmosphere | Cells at a 70–90% confluence were transfected with the mixture containing 1.0 µg pcDNA3+PCSK9D374Y-FLAG plasmid and 2.0 µL TurboFect Transfection Reagent in 100 µL of serum-free DMEM for 48 h. After 24 h, transfected HepG2 cells were treated with peptide T9 (100 µM) and incubated for 24 h at 37 °C under 5% CO2 atmosphere | NA | NR | NR | NR | Low | HepG2 cells were used for no more than 20 passages after thawing |
| Lupo et al. (2019) | NR | NR | NO | HepG2 was cultured in MEM supplemented with 10% FCS, L-glutamine, sodium-pyruvate and non-essential amino acids, penicillin/streptomycin at 37 °C in a humidified atmosphere of 5% CO2 and 95% air. | NR | NA | NR | NR | NR | UNR | No description of the model nor its routine maintenance |
| Wu et al. (2019) | NR | NR | NO | The cell line HepG2 was maintained in DMEM, supplemented with 10% FBS, 100 units/mL penicillin, and 100 mg/mL streptomycin and cultured in a 37 °C 2incubator with 5% CO2 in the air | NR | NA | NR | NR | NR | UNR | |
| Chae et al. (2018) | NR | HepG2 human hepatocellular liver cell line was obtained from the Korea Research Institute of Bioscience and Biotechnology (South Korea) | YES | Cells were grown in EMEM) containing 10% FBS and 100U/Mlpenicillin/streptomycin sulfate. Cells were incubated in a humidified 5% CO2 atmosphere at 37 °C. | NR | NA | NR | NR | NR | UNR | No description of the model nor its routine maintenance |
| Yang et al. (2018) | NR | The human hepatoma cell line, HepG2, was obtained from Cell Resource Center, IBMS, CAMS/PUMC (Beijing, China) | YES | Cells were cultured in DMEM containing 10% FBS 1% non-essential amino acids (NEAA) and 1% penicillin–streptomycin at 37 °C, 5% (v/v) CO2. | HepG2 cells were serum-starved for 18 h and then treated with liraglutide at various concentrations for 24 h | NA | NR | NR | NR | Low | No description of its routine maintenance nor check for contaminants |
| Yang et al. (2018) | NR | The HepG2 cell line was obtained from the American Type Culture Collection (ATCC; Manassas, VA | YES | The cells were cultured in DMEM containing 10% FBS at 37 °C and 5% CO2 atmosphere. After reaching 70–80% confluence, the HepG2 cells were pre-treated with vehicle or COS (50–200 μg/mL) in DMEM with 4% FBS for 24 h. | After reaching 70–80% confluence, the HepG2 cells were pre-treated with vehicle or COS (50–200 μg/mL) in DMEM with 4% FBS for 24 h | NA | NR | NR | NR | Low | |
| Gu et al. (2017) | NR | Human hepatic HepG2 cells were obtained from China Infrastructure of Cell Line Resources (Beijing, China) | YES | Cells were cultured in a complete medium consisting of MEM supplemented with penicillin (100 U/mL), streptomycin (100 μg/mL) and 10% FBS in a humidified 5% CO2 atmosphere at 37 °C. | OptiMEM media was used in the Lunasin dose-response and time-course experiments to measure the amount of PCSK9 secreted into the culture media and LDLR expression | NA | NR | NR | NR | Low | |
| Chen et al. (2016) | NR | HepG2 cells were obtained from the Bioresource Collection and Research Center (Hsinchu, Taiwan) | YES | Cells were maintained in a DMEM medium containing 10% FBS. | The cells were seeded and cultured in normal serum medium overnight; then, the medium was changed to DMEM supplemented with 5% LPDS and was cultured for 24 h. | NA | NR | NR | NR | Low | |
| Cameron et al. (2008) | NR | HepG2 cells (European collection of cell cultures, Wiltshire, UK) | YES | Cells were maintained in MEM, containing penicillin (50 U/mL), streptomycin (50 (g/mL), l-glutamine (2 mM) and 10% fetal calf serum (FCS) in a humidified atmosphere (37 °C, 5% CO2) | OptiMEM (Gibco) media was used instead of media containing 10% LPDS in the dose-response and time-course experiments | NA | NR | NR | NR | Low | No description of its routine maintenance nor check for contaminants |
| Gao et al. (2018) | NR | The HepG2 cell line was obtained from the Bioresource Collection and Research Center 124 (Hsinchu, Taiwan). | YES | The cells were cultured in DMEM containing 10% FBS and 1x non-essential amino acid (NEAA) solution. | For compound treatment, the cells were seeded in a culture medium for 24 h. The medium was replaced with 127 DMEM supplemented with 5% LPDS) for 24 h incubation | NA | NR | NR | NR | Low | |
| Hwang et al. (2021) | NR | HepG2 cells (HB-8065; ATCC, Manassas, VA, USA) | YES | Cells were cultured in high-glucose DMEM supplemented with 10% FBS and 1% antibiotic and anti-mycotic solution DLPS was prepared. | After reaching 70–80% confluence, the cells were seeded in well plates (day 0), and the medium was changed to either DMEM supplemented with FBS or DMEM supplemented with DLPS (day 1). After 24 h incubation, the medium was changed to media supplemented with either FBS or DLPS, and simultaneously treated with either samples (CBE or chemical compounds) or DMSO (day 2). After an additional hour of incubation (day 3), the cells were either washed with cold DPBS or collected for subsequent experiments. | NA | NR | NR | NR | Low | |
| Weng et al. (2021) | NR | Human hepatoma HepG2 cells were purchased from the Chinese Academy of Sciences (Shanghai, China). | YES | HepG2 cells were cultured in DMEM (low glucose), and media were supplemented with 10% FBS and 1% penicillin-streptomycin at 37 °C in a humid atmosphere with 5% CO2. | The cells were inoculated in 12-well plates at 1 × 105 /well, which cultured in DMEM (low glucose) containing 10% FBS at 37 °C with 5% CO2. After cell adherence, the media were replaced with DMEM containing 5% LPDS and incubated for 23 h in the incubator | NA | NR | NR | NR | Low | |
| Pel et al. (2020) | NR | The HepG2 human hepatocellular liver cell line was provided by the Korea Research Institute of Bioscience and Biotechnology, Republic of Korea | YES | Cells were grown in EMEM containing 10% FBS and 100 U/Ml penicillin/streptomycin sulfate. Cells were incubated in a humidified 5% CO2 atmosphere at 37 °C. | NR | NA | NR | NR | NR | UNR | No description of the model nor its routine maintenance |
| Choi et al. (2017) | NR | HepG2 cells (HB-8065; ATCC, Manassas, VA, USA) | YES | Cells were cultured with high glucose DMEM supplemented with 10% FBS and 1% antibiotic and antimycotic solution | After reaching 70–80% confluence, the cells were seeded in 96-well plates (day 0), and the medium was changed to either DMEM supplemented with FBS or DMEM supplemented DLPS; day 1. DLPS was prepared as previously described.26 After 24 hours of incubation, the medium was changed to media supplemented with either FBS or DLPS. | NA | NR | NR | NR | Low | No description of its routine maintenance nor check for contaminants |
| Pel et al. (2017) | NR | NR | NO | NR | NR | NA | NR | NR | NR | High | All domains were not reported/not applicable. |
| Aggrey et al. (2019) | NR | HepG2 cells (ATCC HB-8065) | YES | Cells were maintained in DMEM supplemented with 10% FBS. Cells were incubated under a humidified atmosphere of 95% O2 and 5% CO2 at 37 °C | NR | NA | NR | NR | NR | UNR | No description of the model nor its routine maintenance |
| JLM3 (hepatocellular carcinoma cells) | |||||||||||
| He et al. (2017) | NR | RAW264.7 murine macrophages were obtained from the Korean Research Institute of Bioscience and Biotechnology (Daejeon, Korea) | YES | Cells were grown in RPMI 1640 medium supplemented with 10% FBS and 100 U/ML penicillin/streptomycin sulfate. | NR | NA | NR | NR | NR | UNR | No description of the model nor its routine maintenance |
Appendix E
| Domain | Signalling Question | Notes | Answer |
|---|---|---|---|
| Validation of Marker | 1. Functional validation according to report aims or methods. | Free text | |
| 2. Cellular localisation according to Genecard confidence 5 or cellular components according to Flybase | http://www.genecards.org/ (accessed on 31 August 2022). http://flybase.org/ (accessed on 31 August 2022). | Free text or NA/NR | |
| 3. gene ontology—cellular component terms according to Genecards | http://www.genecards.org/ (accessed on 31 August 2022). | Free text or NA/NR | |
| 4. Do the authors present data for functional validation in results? | This includes: Is the marker in the correct location? Any functional experiments? | Yes/NR/Referenced If yes add free text to justification. | |
| 5. Were co-localisation experiments performed with a second marker/was the result confirmed with a second marker? | |||
| Validation rating | Low= no concerns. Unclear/not reported = insufficient data to make a judgement or not reported High risk = there are concerns | Low/UNR/High/ Referenced If UNR/High add free text to justification | |
| Controls | 6. Is there an appropriate positive control? | Molecular: Result in the presence of another tagged protein/gene that marks the organelle of interest. IHC: Result in another model that expresses the marker | Yes/NR/NA If yes add free text to justification. |
| 7. Is there an appropriate negative control? | Molecular: Result in the presence of a tagged protein that does not mark the organelle of interest OR in the absence of a tagged protein (e.g. empty vector, tag only). IHC: Result in absence of marker, AND result in another model than does not express the marker | Yes/NR/NA If yes add free text to justification. | |
| Control rating | Low= no concerns. Unclear/not reported = insufficient data to make a judgement or not reported. High risk = there are concerns | Low/UNR/High If UNR/High add free text to justification | |
| Experimental performance, reporting flaws | 8. Were there sufficient details to judge the performance of molecular experiments? | Detailed =, allowing repetition of the experiment. Partial = some details, but could not repeat the experiment easily. NR = not reported | D/P/NR/NA |
| 9. Did the authors provide evidence that the genetic manipulation did not influence the observed effect? | Yes/NR/NA If yes add free text to justification. | ||
| 10. Molecular techniques: Additional comments/concerns | Yes/NR/NA If yes add free text to justification. | ||
| 11. Were there sufficient details to judge the performance of immunochemistry? | Detailed = allowing easy repetition of the experiment. Partial = some details, but could not repeat the experiment easily. NR = not reported | D/P/NR/NA | |
| 12. Immunotechniques: Additional comments/concerns | Yes/NR/NA If yes add free text to justification. | ||
| 13. Type of image analysis. | Confocal fluorescent/fluorescent NR/light | ||
| 14. Were there sufficient details to repeat the image analysis? | Detailed = allowing easy repetition of the experiment. Partial = some details, but could not repeat the experiment easily. NR = not reported | D/P/NR/NA | |
| 15. Was the optical plane considered? | Yes/NR/NA | ||
| 16. Additional comments/concerns regarding imaging | Yes/NR/NA If yes add free text to justification. | ||
| Experimental performance rating | Low = no concerns. Unclear/not reported = insufficient data to make a judgement or not reported. High risk = there are concerns | Low/UNR/High If UNR/High add free text to justification | |
| Applicability/generalisability | Model variability (Did the experiment give the same result in a different model?) | Yes = low NR = UNR | Low/UNR If low add free text to justification |
| Additional Biases | 17. Any experimental assumptions? | Yes/NR/NA If yes add free text to justification. | |
| 18. Other concerns/ How was asymmetry measured? | Was subjective assessment used, if yes the results should be verified independently | Yes/NR/NA If yes add free text to justification. | |
| 19. Was the marker stated a priori? | The marker should be stated a priori in the introduction or methods. Or the authors should assess a range of markers clearly stated in the aims. If the authors list the marker in the methods or results only (with no further details or intention) this is unclear/NR. | Yes/NR/No | |
| additional rating | Low= no concerns. Unclear/not reported = insufficient data to make a judgement or not reported. High risk = there are concerns | Low/UNR/High If UNR/High add free text to justification | |
| OVERALL RATING | Low = all domains clearly reported. Unclear = Any domains are unclear, but not high risk. High risk = there is a concern of high risk | Low/UNR/High | |
| JUSTIFICATION | Free text to explain UNR or High ratings, plus additional free text from signalling questions | ||
| Marker | 1. | 2. | 3. | 4. | 5. | Marker Validation Rating | 6. Positive | 7. Negative | Control Rating | |
|---|---|---|---|---|---|---|---|---|---|---|
| HUVEC (Human Umbilical Vein Endothelial Cells) | ||||||||||
| Wang 2019 | PCSK9 | PCSK9 released from HUVEC | Cytoplasm, Endosome, Lysosome, Cell surface, Endoplasmic reticulum, Golgi apparatus. | Extracellular region, extracellular space, cytoplasm, lysosome, and lysosomal membrane. | NR | NR | Low | NR | Yes | UNR |
| LDLR | LDLR released from HUVEC | Cell membrane, Single-pass type I membrane protein, Membrane, clathrin-coated pit, Golgi apparatus. Endosome, Lysosome. | Lysosome, endosome, and golgi Apparatus. | NR | NR | Low | NR | NR | High | |
| ICAM-1 | ICAM-1 released from HUVEC | Membrane. Single pass type I membrane protein. | Immunological synapse, extracellular space, plasma membrane, integral component of plasma membrane, and focal adhesion | NR | NR | Low | NR | Yes | UNR | |
| VCAM | VCAM-1 released from HUVEC | Membrane. Single pass type I membrane protein. | Podosome, extracellular space, early endosome, endoplasmic reticulum, and golgi apparatus. | NR | NR | Low | NR | Yes | UNR | |
| SREBP2 | SREBP2 released from HUVEC | Endoplasmic reticulum membrane. Multi-pass membrane protein. Golgi apparatus membrane. Multi-pass membrane protein. Cytoplasmic vesicle, COPII-coated vesicle membrane. Multi-pass membrane protein. | Golgi membrane, chromarin, nucleus, nucleoplasm, and cytoplasm. | NR | NR | Low | NR | Yes | UNR | |
| IL-1α | IL-1α released from HUVEC | Cytoplasm. | Extracellular region, extracellular space, cytoplasm, cytosol, and plasma membrane. | NR | NR | Low | NR | Yes | UNR | |
| IL-1β | IL-1β released from HUVEC | Cytoplasm, cytosol. Lysosome. Secreted, extracellular exosome. | Extracellular region, extracellular space, cytoplasm, lysosome, and cytosol. | NR | NR | Low | NR | Yes | UNR | |
| IL-6 | IL-6 released from HUVEC | Endoplasmic reticulum, Extracellular exosome, cytosol, nucleus. | Extracellular region, extracellular space, endoplasmic reticulum lumen and interleukin 6 receptor complex. | NR | NR | Low | NR | Yes | UNR | |
| MCP-1 | MCP-1 released from HUVEC | - | - | NR | NR | UNR | NR | Yes | UNR | |
| CXCL-1 | CXCL-1 released from HUVEC | Extracellular exosome. | Extracellular region, extracellular space, and granule lumen. | NR | NR | Low | NR | Yes | UNR | |
| CXCL-2 | CXCL-2 released from HUVEC | - | - | NR | NR | UNR | NR | Yes | UNR | |
| NOX-4 | NOX-4 released from HUVEC | Endoplasmic reticulum membrane. | Nucleus, nucleolus, mitochondria, and endoplasmic reticulum. | NR | NR | Low | NR | Yes | UNR | |
| LOX-1 | LOX-1 released from HUVEC | - | - | NR | NR | UNR | NR | Yes | UNR | |
| Huh7 (Human Hepatocytes) | ||||||||||
| Mbikay 2014 | PCSK9 | PCSK9 released from Huh7 | Cytoplasm, Endosome, Lysosome, Cell surface, Endoplasmic reticulum, Golgi apparatus. | Extracellular region, extracellular space, cytoplasm, lysosome, and lysosomal membrane. | NR | NR | Low | Yes | Yes | Low |
| LDLR | LDLR released from Huh7 | Cell membrane, Single-pass type I membrane protein, Membrane, clathrin-coated pit, Golgi apparatus. Endosome, Lysosome. | Lysosome, endosome, and golgi apparatus. | NR | NR | Low | Yes | Yes | Low | |
| SREBP2 | SREBP2 released from Huh7 | Endoplasmic reticulum membrane. Multi-pass membrane protein. Golgi apparatus membrane. Multi-pass membrane protein. Cytoplasmic vesicle, COPII-coated vesicle membrane. Multi-pass membrane protein. | Golgi membrane, chromarin, nucleus, nucleoplasm, and cytoplasm. | NR | NR | Low | NR | Yes | UNR | |
| Wang 2020 | PCSK9 | PCSK9 released from Huh7 | Cytoplasm, Endosome, Lysosome, Cell surface, Endoplasmic reticulum, Golgi apparatus. | Extracellular region, extracellular space, cytoplasm, lysosome, and lysosomal membrane. | NR | NR | Low | Yes | Yes | Low |
| LDLR | LDLR released from Huh7 | Cell membrane, Single-pass type I membrane protein, Membrane, clathrin-coated pit, Golgi apparatus. Endosome, Lysosome. | Lysosome, endosome, and golgi apparatus. | NR | NR | Low | Yes | Yes | Low | |
| PPARg | PPARg released from Huh7 | Nucleus | Chromatin, nucleus, nucleoplasm, cytoplasm, and cytosol. | NR | NR | Low | NR | Yes | UNR | |
| FoxO3a | FoxO3a released from Huh7 | Cytoplasm, cytosol, nucleus, mitochondrion matrix, mitochondrion outer membrane, peripheral membrane protein, and cytoplasmic side. | Chromatin, nucleus, nucleoplasm, cytoplasm, and mitochondria. | NR | NR | Low | Yes | Yes | Low | |
| LO2 (hepatocytes) | ||||||||||
| Jing 2019 | PCSK9 | PCSK9 released from LO2 | Cytoplasm, Endosome, Lysosome, Cell surface, Endoplasmic reticulum, Golgi apparatus. | Extracellular region, extracellular space, cytoplasm, lysosome, and lysosomal membrane. | NR | NR | Low | NR | Yes | UNR |
| LDLR | LDLR released from LO2 | Cell membrane, Single-pass type I membrane protein, Membrane, clathrin-coated pit, Golgi apparatus. Endosome, Lysosome. | Lysosome, endosome, and golgi apparatus. | NR | NR | Low | NR | Yes | UNR | |
| SREBP 1c | SREBP 1c released from LO2 | Nucleoplasm, cytosol, and golgi apparatus. | Golgi membrane, chromarin, nucleus, nuclear envelope, and nucleoplasm. | NR | NR | Low | NR | Yes | UNR | |
| HepG2 (Human Hepatoma) | ||||||||||
| Aggrey 2019 | PCSK9 | PCSK9 released from HepG2 | Cytoplasm, Endosome, Lysosome, Cell surface, Endoplasmic reticulum, Golgi apparatus. | Extracellular region, extracellular space, cytoplasm, lysosome, and lysosomal membrane. | NR | NR | Low | NR | Yes | UNR |
| LDLR | LDLR released from HepG2 | Cell membrane, Single-pass type I membrane protein, Membrane, clathrin-coated pit, Golgi apparatus. Endosome, Lysosome. | Lysosome, endosome, and golgi apparatus. | NR | NR | Low | NR | Yes | UNR | |
| Ahn 2019 | PCSK9 | PCSK9 released from HepG2 | Cytoplasm, Endosome, Lysosome, Cell surface, Endoplasmic reticulum, Golgi apparatus. | Extracellular region, extracellular space, cytoplasm, lysosome, and lysosomal membrane. | NR | NR | Low | NR | Yes | UNR |
| Cameron 2008 | PCSK9 | PCSK9 released from HepG2 | Cytoplasm, Endosome, Lysosome, Cell surface, Endoplasmic reticulum, Golgi apparatus. | Extracellular region, extracellular space, cytoplasm, lysosome, and lysosomal membrane. | NR | NR | Low | Yes | Yes | Low |
| LDLR | LDLR released from HepG2 | Cell membrane, Single-pass type I membrane protein, Membrane, clathrin-coated pit, Golgi apparatus. Endosome, Lysosome. | Lysosome, endosome, and golgi apparatus. | NR | NR | Low | Yes | Yes | Low | |
| Chae 2018 | PCSK9 | PCSK9 released from HepG2 | Cytoplasm, Endosome, Lysosome, Cell surface, Endoplasmic reticulum, Golgi apparatus. | Extracellular region, extracellular space, cytoplasm, lysosome, and lysosomal membrane. | NR | NR | Low | Yes | Yes | Low |
| LDLR | LDLR released from HepG2 | Cell membrane, Single-pass type I membrane protein, Membrane, clathrin-coated pit, Golgi apparatus. Endosome, Lysosome. | Lysosome, endosome, and golgi apparatus. | NR | NR | Low | Yes | Yes | Low | |
| Chen 2016 | PCSK9 | PCSK9 released from HepG2 | Cytoplasm, Endosome, Lysosome, Cell surface, Endoplasmic reticulum, Golgi apparatus. | Extracellular region, extracellular space, cytoplasm, lysosome, and lysosomal membrane. | NR | NR | Low | NR | Yes | UNR |
| LDLR | LDLR released from HepG2 | Cell membrane, Single-pass type I membrane protein, Membrane, clathrin-coated pit, Golgi apparatus. Endosome, Lysosome. | Lysosome, endosome, and golgi apparatus. | NR | NR | Low | NR | Yes | UNR | |
| Choi 2017 | PCSK9 | PCSK9 released from HepG2 | Cytoplasm, Endosome, Lysosome, Cell surface, Endoplasmic reticulum, Golgi apparatus. | Extracellular region, extracellular space, cytoplasm, lysosome, and lysosomal membrane. | NR | NR | Low | Yes | Yes | Low |
| LDLR | LDLR released from HepG2 | Cell membrane, Single-pass type I membrane protein, Membrane, clathrin-coated pit, Golgi apparatus. Endosome, Lysosome. | Lysosome, endosome, and golgi apparatus. | NR | NR | Low | NR | Yes | UNR | |
| SREBP2 | SREBP2 released from Huh7 | Endoplasmic reticulum membrane. Multi-pass membrane protein. Golgi apparatus membrane. Multi-pass membrane protein. Cytoplasmic vesicle, COPII-coated vesicle membrane. Multi-pass membrane protein. | Golgi membrane, chromarin, nucleus, nucleoplasm, and cytoplasm. | NR | NR | Low | NR | Yes | UNR | |
| HNF1α | HNF1α released from HepG2 | Nucleus | Chromatin, nucleus, transcription regulator complex, and cytoplasm. | NR | NR | Low | NR | Yes | UNR | |
| Dong 2019 | PCSK9 | PCSK9 released from HepG2 | Cytoplasm, Endosome, Lysosome, Cell surface, Endoplasmic reticulum, Golgi apparatus. | Extracellular region, extracellular space, cytoplasm, lysosome, and lysosomal membrane. | NR | NR | Low | NR | Yes | UNR |
| Fan 2021 | PCSK9 | PCSK9 released from HepG2 | Cytoplasm, Endosome, Lysosome, Cell surface, Endoplasmic reticulum, Golgi apparatus. | Extracellular region, extracellular space, cytoplasm, lysosome, and lysosomal membrane. | NR | NR | Low | NR | Yes | UNR |
| LDLR | LDLR released from HepG2 | Cell membrane, Single-pass type I membrane protein, Membrane, clathrin-coated pit, Golgi apparatus. Endosome, Lysosome. | Lysosome, endosome, and golgi apparatus. | NR | NR | Low | NR | Yes | UNR | |
| Gao 2018 | PCSK9 | PCSK9 released from HepG2 | Cytoplasm, Endosome, Lysosome, Cell surface, Endoplasmic reticulum, Golgi apparatus. | Extracellular region, extracellular space, cytoplasm, lysosome, and lysosomal membrane. | NR | NR | Low | Yes | Yes | Low |
| LDLR | LDLR released from HepG2 | Cell membrane, Single-pass type I membrane protein, Membrane, clathrin-coated pit, Golgi apparatus. Endosome, Lysosome. | Lysosome, endosome, and golgi apparatus. | NR | NR | Low | Yes | Yes | Low | |
| SREBP2 | SREBP2 released from HepG2 | Endoplasmic reticulum membrane. Multi-pass membrane protein. Golgi apparatus membrane. Multi-pass membrane protein. Cytoplasmic vesicle, COPII-coated vesicle membrane. Multi-pass membrane protein. | Golgi membrane, chromarin, nucleus, nucleoplasm, and cytoplasm. | NR | NR | Low | NR | Yes | UNR | |
| HNF1α | HNF1α released from HepG2 | Nucleus | Chromatin, nucleus, transcription regulator complex, and cytoplasm. | NR | NR | Low | NR | Yes | UNR | |
| FoxO3a | FoxO3a released from Huh7 | Cytoplasm, cytosol, nucleus, mitochondrion matrix, mitochondrion outer membrane, peripheral membrane protein, and cytoplasmic side. | Chromatin, nucleus, nucleoplasm, cytoplasm, and mitochondria. | NR | NR | Low | NR | Yes | UNR | |
| Fu 2020 | PCSK9 | PCSK9 released from HepG2 | Cytoplasm, Endosome, Lysosome, Cell surface, Endoplasmic reticulum, Golgi apparatus. | Extracellular region, extracellular space, cytoplasm, lysosome, and lysosomal membrane. | NR | NR | Low | NR | Yes | UNR |
| LDLR | LDLR released from HepG2 | Cell membrane, Single-pass type I membrane protein, Membrane, clathrin-coated pit, Golgi apparatus. Endosome, Lysosome. | Lysosome, endosome, and golgi apparatus. | NR | NR | Low | NR | Yes | UNR | |
| Gu 2017 | PCSK9 | PCSK9 released from HepG2 | Cytoplasm, Endosome, Lysosome, Cell surface, Endoplasmic reticulum, Golgi apparatus. | Extracellular region, extracellular space, cytoplasm, lysosome, and lysosomal membrane. | NR | NR | Low | NR | Yes | UNR |
| LDLR | LDLR released from HepG2 | Cell membrane, Single-pass type I membrane protein, Membrane, clathrin-coated pit, Golgi apparatus. Endosome, Lysosome. | Lysosome, endosome, and golgi apparatus. | NR | NR | Low | NR | Yes | UNR | |
| HNF1α | HNF1α released from HepG2 | Nucleus | Chromatin, nucleus, transcription regulator complex, and cytoplasm. | NR | NR | Low | NR | Yes | UNR | |
| SREBP2 | SREBP2 released from HepG2 | Endoplasmic reticulum membrane. Multi-pass membrane protein. Golgi apparatus membrane. Multi-pass membrane protein. Cytoplasmic vesicle, COPII-coated vesicle membrane. Multi-pass membrane protein. | Golgi membrane, chromarin, nucleus, nucleoplasm, and cytoplasm. | NR | NR | Low | NR | Yes | UNR | |
| Hwang 2020 | PCSK9 | PCSK9 released from HepG2 | Cytoplasm, Endosome, Lysosome, Cell surface, Endoplasmic reticulum, Golgi apparatus. | Extracellular region, extracellular space, cytoplasm, lysosome, and lysosomal membrane. | NR | NR | Low | NR | Yes | UNR |
| LDLR | LDLR released from HepG2 | Cell membrane, Single-pass type I membrane protein, Membrane, clathrin-coated pit, Golgi apparatus. Endosome, Lysosome. | Lysosome, endosome, and golgi apparatus. | NR | NR | Low | NR | Yes | UNR | |
| HNF1α | HNF1α released from HepG2 | Nucleus | Chromatin, nucleus, transcription regulator complex, and cytoplasm. | NR | NR | Low | NR | Yes | UNR | |
| SREBP2 | SREBP2 released from HepG2 | Endoplasmic reticulum membrane. Multi-pass membrane protein. Golgi apparatus membrane. Multi-pass membrane protein. Cytoplasmic vesicle, COPII-coated vesicle membrane. Multi-pass membrane protein. | Golgi membrane, chromarin, nucleus, nucleoplasm, and cytoplasm. | NR | NR | Low | NR | Yes | UNR | |
| HMGCR | S HMGCR released from HepG2 | Endoplasmic reticulum membrane. Multi-pass membrane protein. Peroxisome membrane. Multi-pass membrane protein. | Peroxisome, peroxisomal membrane, endoplasmic reticulum, endoplasmic reticulum membrane, and membrane. | NR | NR | Low | NR | Yes | UNR | |
| Hwang 2021 | PCSK9 | PCSK9 released from HepG2 | Cytoplasm, Endosome, Lysosome, Cell surface, Endoplasmic reticulum, Golgi apparatus. | Extracellular region, extracellular space, cytoplasm, lysosome, and lysosomal membrane. | NR | NR | Low | Yes | Yes | Low |
| LDLR | LDLR released from HepG2 | Cell membrane, Single-pass type I membrane protein, Membrane, clathrin-coated pit, Golgi apparatus. Endosome, Lysosome. | Lysosome, endosome, and golgi apparatus. | NR | NR | Low | Yes | Yes | Low | |
| HNF1α | HNF1α released from HepG2 | Nucleus | Chromatin, nucleus, transcription regulator complex, and cytoplasm. | NR | NR | Low | Yes | Yes | Low | |
| SREBP2 | SREBP2 released from HepG2 | Endoplasmic reticulum membrane. Multi-pass membrane protein. Golgi apparatus membrane. Multi-pass membrane protein. Cytoplasmic vesicle, COPII-coated vesicle membrane. Multi-pass membrane protein. | Golgi membrane, chromarin, nucleus, nucleoplasm, and cytoplasm. | NR | NR | Low | Yes | Yes | Low | |
| Kim 2020 | PCSK9 | PCSK9 released from HepG2 | Cytoplasm, Endosome, Lysosome, Cell surface, Endoplasmic reticulum, Golgi apparatus. | Extracellular region, extracellular space, cytoplasm, lysosome, and lysosomal membrane. | NR | NR | Low | Yes | Yes | Low |
| LDLR | LDLR released from HepG2 | Cell membrane, Single-pass type I membrane protein, Membrane, clathrin-coated pit, Golgi apparatus. Endosome, Lysosome. | Lysosome, endosome, and golgi apparatus. | NR | NR | Low | Yes | Yes | Low | |
| HNF1α | HNF1α released from HepG2 | Nucleus | Chromatin, nucleus, transcription regulator complex, and cytoplasm. | NR | NR | Low | NR | Yes | UNR | |
| SREBP2 | SREBP2 released from HepG2 | Endoplasmic reticulum membrane. Multi-pass membrane protein. Golgi apparatus membrane. Multi-pass membrane protein. Cytoplasmic vesicle, COPII-coated vesicle membrane. Multi-pass membrane protein. | Golgi membrane, chromarin, nucleus, nucleoplasm, and cytoplasm. | NR | NR | Low | NR | Yes | UNR | |
| Lammi 2019 | PCSK9 | PCSK9 released from HepG2 | Cytoplasm, Endosome, Lysosome, Cell surface, Endoplasmic reticulum, Golgi apparatus. | Extracellular region, extracellular space, cytoplasm, lysosome, and lysosomal membrane. | NR | NR | Low | NR | Yes | UNR |
| LDLR | LDLR released from HepG2 | Cell membrane, Single-pass type I membrane protein, Membrane, clathrin-coated pit, Golgi apparatus. Endosome, Lysosome. | Lysosome, endosome, and golgi apparatus. | NR | NR | Low | NR | Yes | UNR | |
| HNF1α | HNF1α released from HepG2 | Nucleus | Chromatin, nucleus, transcription regulator complex, and cytoplasm. | NR | NR | Low | NR | Yes | UNR | |
| Li 2020 | PCSK9 | PCSK9 released from HepG2 | Cytoplasm, Endosome, Lysosome, Cell surface, Endoplasmic reticulum, Golgi apparatus. | Extracellular region, extracellular space, cytoplasm, lysosome, and lysosomal membrane. | NR | NR | Low | NR | Yes | UNR |
| LDLR | LDLR released from HepG2 | Cell membrane, Single-pass type I membrane protein, Membrane, clathrin-coated pit, Golgi apparatus. Endosome, Lysosome. | Lysosome, endosome, and golgi apparatus. | NR | NR | Low | Yes | Yes | Low | |
| SREBP2 | SREBP2 released from HepG2 | Endoplasmic reticulum membrane. Multi-pass membrane protein. Golgi apparatus membrane. Multi-pass membrane protein. Cytoplasmic vesicle, COPII-coated vesicle membrane. Multi-pass membrane protein. | Golgi membrane, chromarin, nucleus, nucleoplasm, and cytoplasm. | NR | NR | Low | NR | Yes | UNR | |
| HNF1α | HNF1α released from HepG2 | Nucleus | Chromatin, nucleus, transcription regulator complex, and cytoplasm. | NR | NR | Low | Yes | Yes | Low | |
| Masagalli 2021 | PCSK9 | PCSK9 released from HepG2 | Cytoplasm, Endosome, Lysosome, Cell surface, Endoplasmic reticulum, Golgi apparatus. | Extracellular region, extracellular space, cytoplasm, lysosome, and lysosomal membrane. | NR | NR | Low | NR | Yes | UNR |
| Pel 2020 | PCSK9 | PCSK9 released from HepG2 | Cytoplasm, Endosome, Lysosome, Cell surface, Endoplasmic reticulum, Golgi apparatus. | Extracellular region, extracellular space, cytoplasm, lysosome, and lysosomal membrane. | NR | NR | Low | NR | Yes | UNR |
| LDLR | LDLR released from HepG2 | Cell membrane, Single-pass type I membrane protein, Membrane, clathrin-coated pit, Golgi apparatus. Endosome, Lysosome. | Lysosome, endosome, and golgi apparatus. | NR | NR | Low | NR | Yes | UNR | |
| HNF1α | HNF1α released from HepG2 | Nucleus | Chromatin, nucleus, transcription regulator complex, and cytoplasm. | NR | NR | Low | NR | Yes | UNR | |
| Pel 2017 | PCSK9 | PCSK9 released from HepG2 | Cytoplasm, Endosome, Lysosome, Cell surface, Endoplasmic reticulum, Golgi apparatus. | Extracellular region, extracellular space, cytoplasm, lysosome, and lysosomal membrane. | NR | NR | Low | Yes | Yes | Low |
| Weng 2021 | PCSK9 | PCSK9 released from HepG2 | Cytoplasm, Endosome, Lysosome, Cell surface, Endoplasmic reticulum, Golgi apparatus. | Extracellular region, extracellular space, cytoplasm, lysosome, and lysosomal membrane. | NR | NR | Low | Yes | Yes | Low |
| Wang 2021 | PCSK9 | PCSK9 released from HepG2 | Cytoplasm, Endosome, Lysosome, Cell surface, Endoplasmic reticulum, Golgi apparatus. | Extracellular region, extracellular space, cytoplasm, lysosome, and lysosomal membrane. | NR | NR | Low | NR | Yes | UNR |
| LDLR | LDLR released from HepG2 | Cell membrane, Single-pass type I membrane protein, Membrane, clathrin-coated pit, Golgi apparatus. Endosome, Lysosome. | Lysosome, endosome, and golgi apparatus. | NR | NR | Low | NR | Yes | UNR | |
| SREBP2 | SREBP2 released from HepG2 | Endoplasmic reticulum membrane. Multi-pass membrane protein. Golgi apparatus membrane. Multi-pass membrane protein. Cytoplasmic vesicle, COPII-coated vesicle membrane. Multi-pass membrane protein. | Golgi membrane, chromarin, nucleus, nucleoplasm, and cytoplasm. | NR | NR | Low | NR | Yes | UNR | |
| Wu 2019 | PCSK9 | PCSK9 released from HepG2 | Cytoplasm, Endosome, Lysosome, Cell surface, Endoplasmic reticulum, Golgi apparatus. | Extracellular region, extracellular space, cytoplasm, lysosome, and lysosomal membrane. | NR | NR | Low | Yes | Yes | Low |
| LDLR | LDLR released from HepG2 | Cell membrane, Single-pass type I membrane protein, Membrane, clathrin-coated pit, Golgi apparatus. Endosome, Lysosome. | Lysosome, endosome, and golgi apparatus. | NR | NR | Low | Yes | Yes | Low | |
| Wu 2021 | PCSK9 | PCSK9 released from HepG2 | Cytoplasm, Endosome, Lysosome, Cell surface, Endoplasmic reticulum, Golgi apparatus. | Extracellular region, extracellular space, cytoplasm, lysosome, and lysosomal membrane. | NR | NR | Low | NR | Yes | UNR |
| LDLR | LDLR released from HepG2 | Cell membrane, Single-pass type I membrane protein, Membrane, clathrin-coated pit, Golgi apparatus. Endosome, Lysosome. | Lysosome, endosome, and golgi apparatus. | NR | NR | Low | NR | Yes | UNR | |
| SREBP2 | SREBP2 released from HepG2 | Endoplasmic reticulum membrane. Multi-pass membrane protein. Golgi apparatus membrane. Multi-pass membrane protein. Cytoplasmic vesicle, COPII-coated vesicle membrane. Multi-pass membrane protein. | Golgi membrane, chromarin, nucleus, nucleoplasm, and cytoplasm. | NR | NR | Low | NR | Yes | UNR | |
| HMGCR | S HMGCR released from HepG2 | Endoplasmic reticulum membrane. Multi-pass membrane protein. Peroxisome membrane. Multi-pass membrane protein. | Peroxisome, peroxisomal membrane, endoplasmic reticulum, endoplasmic reticulum membrane, and membrane. | NR | NR | Low | NR | Yes | UNR | |
| HNF1α | HNF1α released from HepG2 | Nucleus | Chromatin, nucleus, transcription regulator complex, and cytoplasm. | NR | NR | Low | NR | Yes | UNR | |
| Yang 2018 | PCSK9 | PCSK9 released from HepG2 | Cytoplasm, Endosome, Lysosome, Cell surface, Endoplasmic reticulum, Golgi apparatus. | Extracellular region, extracellular space, cytoplasm, lysosome, and lysosomal membrane. | NR | NR | Low | NR | Yes | UNR |
| HNF1α | HNF1α released from HepG2 | Nucleus | Chromatin, nucleus, transcription regulator complex, and cytoplasm. | NR | NR | Low | NR | Yes | UNR | |
| Yang 2018 | PCSK9 | PCSK9 released from HepG2 | Cytoplasm, Endosome, Lysosome, Cell surface, Endoplasmic reticulum, Golgi apparatus. | Extracellular region, extracellular space, cytoplasm, lysosome, and lysosomal membrane. | NR | NR | Low | NR | Yes | UNR |
| SREBP2 | SREBP2 released from HepG2 | Endoplasmic reticulum membrane. Multi-pass membrane protein. Golgi apparatus membrane. Multi-pass membrane protein. Cytoplasmic vesicle, COPII-coated vesicle membrane. Multi-pass membrane protein. | Golgi membrane, chromarin, nucleus, nucleoplasm, and cytoplasm. | NR | NR | Low | NR | Yes | UNR | |
| HNF1α | HNF1α released from HepG2 | Nucleus | Chromatin, nucleus, transcription regulator complex, and cytoplasm. | NR | NR | Low | NR | Yes | UNR | |
| FoxO3a | FoxO3a released from Huh7 | Cytoplasm, cytosol, nucleus, mitochondrion matrix, mitochondrion outer membrane, peripheral membrane protein, and cytoplasmic side. | Chromatin, nucleus, nucleoplasm, cytoplasm, and mitochondria. | NR | NR | Low | NR | Yes | UNR | |
| Lupo 2019 | PCSK9 | PCSK9 released from HepG2 | Cytoplasm, Endosome, Lysosome, Cell surface, Endoplasmic reticulum, Golgi apparatus. | Extracellular region, extracellular space, cytoplasm, lysosome, and lysosomal membrane. | NR | NR | Low | Yes | Yes | Low |
| LDLR | LDLR released from HepG2 | Cell membrane, Single-pass type I membrane protein, Membrane, clathrin-coated pit, Golgi apparatus. Endosome, Lysosome. | Lysosome, endosome, and golgi apparatus. | NR | NR | Low | Yes | Yes | Low | |
| HMGCR | HMGCR released from HepG2 | Endoplasmic reticulum membrane. Multi-pass membrane protein. Peroxisome membrane. Multi-pass membrane protein. | Peroxisome, peroxisomal membrane, endoplasmic reticulum, endoplasmic reticulum membrane, and membrane. | NR | NR | Low | Yes | Yes | Low | |
| FAS | FAS released from HepG2 | Cell membrane. Single-pass type I membrane protein. Membrane raft. | Extracellular region, cytosol, plasma membrane, and cell surface, | NR | NR | Low | NR | Yes | UNR | |
| Wang 2020 | PCSK9 | PCSK9 released from HepG2 | Cytoplasm, Endosome, Lysosome, Cell surface, Endoplasmic reticulum, Golgi apparatus. | Extracellular region, extracellular space, cytoplasm, lysosome, and lysosomal membrane. | NR | NR | Low | Yes | Yes | Low |
| LDLR | LDLR released from HepG2 | Cell membrane, Single-pass type I membrane protein, Membrane, clathrin-coated pit, Golgi apparatus. Endosome, Lysosome. | Lysosome, endosome, and golgi apparatus. | NR | NR | Low | Yes | Yes | Low | |
| PPARg | PPARg released from Huh7 | Nucleus | Chromatin, nucleus, nucleoplasm, cytoplasm, and cytosol. | NR | NR | Low | NR | Yes | UNR | |
| FoxO3a | FoxO3a released from Huh7 | Cytoplasm, cytosol, nucleus, mitochondrion matrix, mitochondrion outer membrane, peripheral membrane protein, and cytoplasmic side. | Chromatin, nucleus, nucleoplasm, cytoplasm, and mitochondria. | NR | NR | Low | Yes | Yes | Low | |
| JLM3 (hepatocellular carcinoma cells) | ||||||||||
| He 2017 | PCSK9 | PCSK9 released from HepG2 | Cytoplasm, Endosome, Lysosome, Cell surface, Endoplasmic reticulum, Golgi apparatus. | Extracellular region, extracellular space, cytoplasm, lysosome, and lysosomal membrane. | NR | NR | Low | Yes | Yes | Low |
| LDLR | LDLR released from HepG2 | Cell membrane, Single-pass type I membrane protein, Membrane, clathrin-coated pit, Golgi apparatus. Endosome, Lysosome. | Lysosome, endosome, and golgi apparatus. | NR | NR | Low | Yes | Yes | Low | |
References
- Collis, A.; Ross, J.; Lang, S.H. A systematic review of the asymmetric inheritance of cellular organelles in eukaryotes: A critique of basic science validity and imprecision. PLoS ONE 2017, 12, e0178645. [Google Scholar] [CrossRef]
- Batista, N.T.; Rebersek, M.; Vernier, P.T.; Mali, B.; Miklavcic, D. Effects of high voltage nanosecond electric pulses on eucaryotic cells (in vitro): A systematic review. Bioelectrochemistry 2016, 110, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Pavan, L.M.; Rego, D.F.; Elias, S.T.; De Luca Canto, G.; Guerra, E.N. In vitro Anti-Tumor Effects of Statins on Head and Neck Squamous Cell Carcinoma: A Systematic Review. PLoS ONE 2015, 10, e0130476. [Google Scholar] [CrossRef] [PubMed]
- Bus, P.; Siersema, P.D.; Van Baal, J.W.P.M. Cell culture models for studying the development of Barrett’s esophagus: A systematic review. Cell. Oncol. 2012, 35, 149–161. [Google Scholar] [CrossRef]
- De Vries, R.B.; Wever, K.E.; Avey, M.T.; Stephens, M.L.; Sena, E.S.; Leenaars, M. The usefulness of systematic reviews of animal experiments for the design of preclinical and clinical studies. ILAR J. 2014, 55, 427–437. [Google Scholar] [CrossRef]
- Sabatine, M.S.; Giugliano, R.P.; Keech, A.C.; Honarpour, N.; Wiviott, S.D.; Murphy, S.A. Evolocumab and clinical outcomes in patients with cardiovascular disease. N. Engl. J. Med. 2017, 376, 1713–1722. [Google Scholar] [CrossRef]
- Ballantyne, C.M.; Neutel, J.; Cropp, A.; Duggan, W.; Wang, E.Q.; Plowchalk, D.; Sweeney, K.; Kaila, N.; Vincent, J.; Bays, H. Results of bococizumab, a monoclonal antibody against proprotein convertase subtilisin/Kexin type 9, from a randomized, placebo-controlled, dose-ranging study in statin-treated subjects with hypercholesterolemia. Am. J. Cardiol. 2015, 115, 1212–1221. [Google Scholar] [CrossRef]
- Kereiakes, D.J.; Robinson, J.G.; Cannon, C.P.; Lorezento, C.; Pordy, R.; Chaudari, U.; Colhoun, H.M. Efficacy and safety of the PCSK9 inhibitor alirocumab among high cardiovascular risk patients on maximally tolerated statin therapy: The ODYSSEY COMBO I study. Am. Heart J. 2015, 169, 906–915. [Google Scholar] [CrossRef]
- Seidah, N.G.; Benjannet, S.; Wickham, L.; Marcinkiewicz, J.; Jasmin, S.B.; Stifani, S. The secretory proprotein convertase neural apoptosis-regulated convertase 1 (NARC-1): Liver regeneration and neuronal differentiation. Proc. Natl. Acad. Sci. USA 2003, 100, 928–933. [Google Scholar] [CrossRef]
- Soutar, A.K.; Naoumova, R.P. Mechanisms of disease: Genetic causes of familial hypercholesterolemia. Nat. Clin. Pract. Cardiovasc. Med. 2007, 4, 214–225. [Google Scholar] [CrossRef]
- Seidah, N.G.; Awan, Z.; Chretien, M.; Mbikay, M. PCSK9: A key modulator of cardiovascular health. Circ. Res. 2014, 114, 1022–1036. [Google Scholar] [CrossRef]
- Rashid, S.; Curtis, D.E.; Garuti, R.; Anderson, N.N.; Bashmakov, Y.; Ho, Y.K.; Hammer, R.E.; Moon, Y.A.; Horton, J.D. Decreased plasma cholesterol and hypersensitivity to statins in mice lacking Pcsk9. Proc. Natl. Acad. Sci. USA 2005, 102, 5374–5379. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.; Tuakli-Wosornu, Y.; Lagace, T.A.; Kinch, L.; Grishin, N.V.; Horton, J.D.; Cohen, J.C.; Hobbs, H.H. Molecular characterization of loss-of-function mutations in PCSK9 and identification of a compound heterozygote. Am. J. Hum. Genet. 2006, 79, 514–523. [Google Scholar] [CrossRef]
- Robinson, J.G.; Farnier, M.; Krempf, M.; Bergeron, J.; Luc, G.; Averna, M.; Stroes, E.S.; Langslet, G.; Raal, F.J.; Shahawy, M.; et al. Efficacy and safety of alirocumab in reducing lipids and cardiovascular events. N. Engl. J. Med. 2015, 372, 1489–1499. [Google Scholar] [CrossRef]
- Raal, F.J.; Honarpour, N.; Blom, D.J. Inhibition of PCSK9 with evolocumab in homozygous familial hypercholesterolaemia (TESLA Part B): A randomised, double-blind, placebo-controlled trial. Lancet 2015, 385, 341–350. [Google Scholar] [CrossRef]
- Shapiro, M.D.; Fazio, S. PCSK9 and atherosclerosis-lipids and beyond. J. Atheroscler. Thromb. 2017, 24, 462–472. [Google Scholar] [CrossRef]
- Barale, C.; Melchionda, E.; Morotti, A.; Russo, I. PCSK9 Biology and Its Role in Atherothrombosis. Int. J. Mol. Sci. 2021, 22, 5880. [Google Scholar] [CrossRef]
- Nicholls, S.J.; Puri, R.; Anderson, T.; Ballantyne, C.M.; Cho, L.; Kastelein, J.J. Effect of evolocumab on progression of coronary disease in statin-treated patients: The GLAGOV randomised clinical trial. JAMA 2016, 316, 2373–2384. [Google Scholar] [CrossRef]
- Adorni, M.P.; Zimetti, F.; Lupo, M.G.; Ruscica, M.; Ferri, N. Naturally Occurring PCSK9 Inhibitors. Nutrients 2020, 12, 1440. [Google Scholar] [CrossRef]
- Mohd Ariff, A.; Abu Bakar, N.A.; Abd Muid, S.; Omar, E.; Ismail, N.H.; Ali, A.M.; Mohd Kasim, N.A.; Mohd Nawawi, H. Ficus deltoidea suppresses endothelial activation, inflammation, monocytes adhesion and oxidative stress via NF-κB and eNOS pathways in stimulated human coronary artery endothelial cells. BMC Complement. Med Ther. 2020, 20, 56. [Google Scholar] [CrossRef]
- Chumachenko, P.; Leusova, Y.Y.; Kheimets, G.; Chasova, E. Adhesion molecules and subpopulation of mononuclear cells in pulmonary arteries. Kardiologiia 2016, 56, 35–39. [Google Scholar] [CrossRef] [PubMed]
- Khodabandehlou, K.; Masehi-Lano, J.J.; Poon, C.; Wang, J.; Chung, E.J. Targeting cell adhesion molecules with nanoparticles using in vivo and flow-based in vitro models of atherosclerosis. Exp. Biol. Med. 2017, 242, 799–812. [Google Scholar] [CrossRef] [PubMed]
- Higgins, J.P.T.; Green, S. Cochrane Handbook for Systematic Reviews of Interventions; Version 5.1.0; The Cochrane Collaboration: London, UK, 2011. [Google Scholar]
- Page, M.J.; McKenzie, J.E.; Bossuyt, P.M.; Boutron, I.; Hoffmann, T.C.; Mulrow, C.D.; Shamseer, L.; Tetzlaff, J.M.; Akl, E.A.; Brennan, S.E.; et al. The PRISMA 2020 statement: An updated guideline for reporting systematic reviews. BMJ 2021, 372, n71. [Google Scholar] [CrossRef]
- Abifadel, M.; Varret, M.; Rabes, J.P.; Allard, D.; Ouguerram, K.; Devillers, M.; Cruaud, C.; Benjannet, S.; Wickham, L.; Erlich, D.; et al. Mutations in PCSK9 cause autosomal dominant hypercholesterolemia. Nat. Genet. 2003, 34, 154–156. [Google Scholar] [CrossRef]
- Hooijmans, C.R.; Rovers, M.M.; de Vries, R.B.; Leenaars, M.; Ritskes-Hoitinga, M.; Langendam, M.W. SYRCLE’s risk of bias tool for animal studies. BMC Med. Res. Methodol. 2014, 14, 43. [Google Scholar] [CrossRef]
- Wang, G.; Liu, Z.; Li, M.; Li, Y.; Alvi, S.S.; Ansari, I.A.; Khan, M.S. Ginkgolide B Mediated Alleviation of Inflammatory Cascades and Altered Lipid Metabolism in HUVECs via Targeting PCSK-9 Expression and Functionality. BioMed Res. Int. 2019, 2019, 7284767. [Google Scholar] [CrossRef]
- Mbikay, M.; Sirois, F.; Simoes, S.; Mayne, J.; Chretien, M. Quercetin-3-glucoside increases low-density lipoprotein receptor (LDLR) expression, attenuates proprotein convertase subtilisin/kexin 9 (PCSK9) secretion and stimulates LDL uptake by Huh7 human hepatocytes in culture. FEBS Open Bio. 2014, 24, 755–762. [Google Scholar] [CrossRef]
- Wang, D.; Yang, X.; Chen, Y.; Gong, K.; Yu, M.; Gao, Y.; Wu, X.; Hu, H.; Liao, C.; Han, J.; et al. Ascorbic acid enhances low-density lipoprotein receptor expression by suppressing proprotein convertase subtilisin/kexin 9 expression. J. Biol. Chem. 2020, 295, 15870–15882. [Google Scholar] [CrossRef]
- Jing, Y.; Hu, T.; Lin, C.; Xiong, Q.; Liu, F.; Yuan, J.; Zhao, X.; Wang, R. Resveratrol downregulates PCSK9 expression and attenuates steatosis through estrogen receptor α-mediated pathway in L02 cells. Eur. J. Pharmacol. 2019, 855, 216–226. [Google Scholar] [CrossRef]
- Aggrey, M.O.; Li, H.H.; Wang, W.Q.; Wang, Y.; Xuan, L.J. Indole alkaloid from Nauclea latifolia promotes LDL uptake in HepG2 cells by inhibiting PCSK9. Phytomedicine 2019, 55, 264–268. [Google Scholar] [CrossRef]
- Ahn, J.; Kim, Y.M.; Chae, H.S.; Choi, Y.H.; Ahn, H.C.; Yoo, H.; Kang, M.; Kim, J.; Chin, Y.W. Prenylated Flavonoids from the Roots and Rhizomes of Sophora tonkinensis and Their Effects on the Expression of Inflammatory Mediators and Proprotein Convertase Subtilisin/Kexin Type 9. J. Nat. Prod. 2019, 82, 309–317. [Google Scholar] [CrossRef] [PubMed]
- Cameron, J.; Ranheim, T.; Kulseth, M.A.; Leren, T.P.; Berge, K.E. Berberine decreases PCSK9 expression in HepG2 cells. Atherosclerosis 2008, 201, 266–273. [Google Scholar] [CrossRef] [PubMed]
- Chae, H.S.; You, B.H.; Kim, D.Y.; Lee, H.; Ko, H.W.; Ko, H.J.; Choi, Y.H.; Choi, S.S.; Chin, Y.W. Sauchinone controls hepatic cholesterol homeostasis by the negative regulation of PCSK9 transcriptional network. Sci. Rep. 2018, 8, 6737. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.C.; Chen, P.Y.; Wu, M.J.; Tai, M.H.; Yen, J.H. Tanshinone IIA Modulates Low-Density Lipoprotein Uptake via Down-Regulation of PCSK9 Gene Expression in HepG2 Cells. PLoS ONE 2016, 11, e0162414. [Google Scholar] [CrossRef]
- Choi, H.K.; Hwang, J.T.; Nam, T.G.; Kim, S.H.; Min, D.K.; Park, S.W.; Chung, M.Y. Welsh onion extract inhibits PCSK9 expression contributing to the maintenance of the LDLR level under lipid depletion conditions of HepG2 cells. Food Funct. 2017, 8, 4582–4591. [Google Scholar] [CrossRef] [PubMed]
- Dong, Z.; Zhang, W.; Chen, S.; Liu, C. Silibinin A decreases statin-induced PCSK9 expression in human hepatoblastoma HepG2 cells. Mol. Med. Rep. 2019, 20, 1383–1392. [Google Scholar] [CrossRef] [PubMed]
- Fan, T.Y.; Yang, Y.X.; Zeng, Q.X.; Wang, X.L.; Wei, W.; Guo, X.X.; Zhao, L.P.; Song, D.Q.; Wang, Y.X.; Wang, L.; et al. Structure-activity relationship and biological evaluation of berberine derivatives as PCSK9 down-regulating agents. Bioorg. Chem. 2021, 113, 104994. [Google Scholar] [CrossRef] [PubMed]
- Gao, W.Y.; Chen, P.Y.; Chen, S.F.; Wu, M.J.; Chang, H.Y.; Yen, J.H. Pinostrobin Inhibits Proprotein Convertase Subtilisin/Kexin-type 9 (PCSK9) Gene Expression through the Modulation of FoxO3a Protein in HepG2 Cells. J. Agric. Food Chem. 2018, 66, 6083–6093. [Google Scholar] [CrossRef]
- Fu, W.; Gao, X.P.; Zhang, S.; Dai, Y.P.; Zou, W.J.; Yue, L. M17β-Estradiol Inhibits PCSK9-Mediated LDLR Degradation Through GPER/PLC Activation in HepG2 Cells. Front. Endocrinol. 2020, 10, 930. [Google Scholar] [CrossRef]
- Gu, L.; Wang, Y.; Xu, Y.; Tian, Q.; Lei, G.; Zhao, C.; Gao, Z.; Pan, Q.; Zhao, W.; Nong, L.; et al. Lunasin functionally enhances LDL uptake via inhibiting PCSK9 and enhancing LDLR expression in vitro and in vivo. Oncotarget 2017, 8, 80826–80840. [Google Scholar] [CrossRef]
- Hwang, J.T.; Kim, H.J.; Choi, H.K.; Park, J.H.; Chung, S.; Chung, M.Y. Butein Synergizes with Statin to Upregulate Low-Density Lipoprotein Receptor Through HNF1α-Mediated PCSK9 Inhibition in HepG2 Cells. J. Med. Food. 2020, 23, 1102–1108. [Google Scholar] [CrossRef]
- Hwang, J.T.; Choi, E.; Choi, H.K.; Park, J.H.; Chung, M.Y. The Cholesterol-Lowering Effect of Capsella Bursa-Pastoris Is Mediated via SREBP2 and HNF-1α-Regulated PCSK9 Inhibition in Obese Mice and HepG2 Cells. Foods 2021, 10, 408. [Google Scholar] [CrossRef]
- Kim, H.J.; Lee, J.; Chung, M.Y.; Hong, S.; Park, J.H.; Lee, S.H.; Park, S.W.; Choi, H.K.; Hwang, J.T. Piceatannol reduces resistance to statins in hypercholesterolemia by reducing PCSK9 expression through p300 acetyltransferase inhibition. Pharmacol. Res. 2020, 161, 105205. [Google Scholar] [CrossRef]
- Lammi, C.; Bollati, C.; Lecca, D.; Abbracchio, M.P.; Arnoldi, A. Lupin Peptide T9 (GQEQSHQDEGVIVR) Modulates the Mutant PCSK9D374Y Pathway: In vitro Characterization of its Dual Hypocholesterolemic Behavior. Nutrients 2019, 11, 1665. [Google Scholar] [CrossRef]
- Li, H.H.; Li, J.; Zhang, X.J.; Li, J.M.; Xi, C.; Wang, W.Q.; Lu, Y.L.; Xuan, L.J. 23,24-Dihydrocucurbitacin B promotes lipid clearance by dual transcriptional regulation of LDLR and PCSK9. Acta Pharmacol. Sin. 2020, 41, 327–335. [Google Scholar] [CrossRef]
- Masagalli, J.N.; BasavanaGowda, M.K.; Chae, H.S.; Choi, W.J. Synthesis of Moracin C and Its Derivatives with a 2-arylbenzofuran Motif and Evaluation of Their PCSK9 Inhibitory Effects in HepG2 Cells. Molecules 2021, 26, 1327. [Google Scholar] [CrossRef]
- Pel, P.; Chae, H.S.; Nhoek, P.; Kim, Y.M.; Khiev, P.; Kim, G.J.; Nam, J.W.; Choi, H.; Choi, Y.H.; Chin, Y.W. A stilbene dimer and flavonoids from the aerial parts of Chromolaena odorata with proprotein convertase subtilisin/kexin type 9 expression inhibitory activity. Bioorg. Chem. 2020, 99, 103869. [Google Scholar] [CrossRef]
- Pel, P.; Chae, H.S.; Nhoek, P.; Kim, Y.M.; Chin, Y.W. Chemical Constituents with Proprotein Convertase Subtilisin/Kexin Type 9 mRNA Expression Inhibitory Activity from Dried Immature Morus alba Fruits. J. Agric. Food Chem. 2017, 65, 5316–5321. [Google Scholar] [CrossRef]
- Weng, X.; Lou, Y.Y.; Wang, Y.S.; Huang, Y.P.; Zhang, J.; Yin, Z.Q.; Pan, K. New dammarane-type glycosides from Gynostemma pentaphyllum and their lipid-lowering activity. Bioorg. Chem. 2021, 111, 104843. [Google Scholar] [CrossRef]
- Wang, J.; Wang, Y.S.; Huang, Y.P.; Jiang, C.H.; Gao, M.; Zheng, X.; Yin, Z.Q.; Zhang, J. Gypenoside LVI improves hepatic LDL uptake by decreasing PCSK9 and upregulating LDLR expression. Phytomedicine 2021, 91, 153688. [Google Scholar] [CrossRef]
- Wu, C.; Xi, C.; Tong, J.; Zhao, J.; Jiang, H.; Wang, J.; Wang, Y.; Liu, H. Design, synthesis, and biological evaluation of novel tetrahydroprotoberberine derivatives (THPBs) as proprotein convertase subtilisin/kexin type 9 (PCSK9) modulators for the treatment of hyperlipidemia. Acta Pharm. Sin. B 2019, 9, 1216–1230. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.-R.; Li, L.; Sun, X.-C.; Wang, J.; Ma, C.-Y.; Zhang, Y.; Qu, H.-L.; Xu, R.-X.; Li, J.-J. Diallyl disulfide improves lipid metabolism by inhibiting PCSK9 expression and increasing LDL uptake via PI3K/Akt-SREBP2 pathway in HepG2 cells. Nutr. Metab. Cardiovasc. Dis. 2020, 31, 322–332. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.H.; Xu, R.X.; Cui, C.J.; Wang, Y.; Du, Y.; Chen, Z.G.; Yao, Y.H.; Ma, C.Y.; Zhu, C.G.; Guo, Y.L.; et al. Liraglutide downregulates hepatic LDL receptor and PCSK9 expression in HepG2 cells and db/db mice through a HNF-1a dependent mechanism. Cardiovasc. Diabetol. 2018, 17, 48. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Zhang, J.; Chen, L.; Wu, Q.; Yu, C. Chitosan oligosaccharides enhance lipid droplets via down-regulation of PCSK9 gene expression in HepG2 cells. Exp. Cell Res. 2018, 366, 152–160. [Google Scholar] [CrossRef]
- Lupo, M.G.; Macchi, C.; Marchiano, S.; Cristofani, R.; Greco, M.F.; Dall’Acqua, S.; Chen, H.; Sirtori, C.R.; Corsini, A.; Ruscica, M.; et al. Differential effects of red yeast rice, Berberis aristata and Morus alba extracts on PCSK9 and LDL uptake. Nutr. Metab. Cardiovasc. Dis. 2019, 29, 1245–1253. [Google Scholar] [CrossRef]
- He, M.; Hou, J.; Wang, L.; Zheng, M.; Fang, T.; Wang, X.; Xia, J. Actinidia chinensis Planch root extract inhibits cholesterol metabolism in hepatocellular carcinoma through upregulation of PCSK9. Oncotarget 2017, 8, 42136–42148. [Google Scholar] [CrossRef]
- Liu, Y.; Beyer, A.; Aebersold, R. On the dependency of cellular protein levels on mRNA abundance. Cell 2016, 165, 535–550. [Google Scholar] [CrossRef]
- Csardi, G.; Franks, A.; Choi, D.S.; Airoldi, E.M.; Drummond, D.A. Accounting for experimental noise reveals that mRNA levels, amplified by post-transcriptional processes, largely determine steady-state protein levels in yeast. PLoS Genet. 2015, 11, e1005206. [Google Scholar] [CrossRef]
- Silva, G.M.; Vogel, C. Quantifying gene expression: The importance of being subtle. Mol. Syst. Biol. 2016, 12, 885. [Google Scholar] [CrossRef]
- Cheng, Z.; Teo, G.; Krueger, S.; Rock, T.M.; Koh, H.W.; Choi, H.; Vogel, C. Differential dynamics of the mammalian mRNA and protein expression response to misfolding stress. Mol. Syst. Biol. 2016, 12, 855. [Google Scholar] [CrossRef]
- Garg, A.; Simha, V. Update on dyslipidemia. J. Clin. Endocrinol. Metab. 2007, 92, 1581–1589. [Google Scholar] [CrossRef]
- Zhang, D.W.; Lagace, T.A.; Garuti, R.; Zhao, Z.; McDonald, M.; Horton, J.D.; Cohen, J.C.; Hobbs, H.H. Binding of proprotein convertase subtilisin/Kexin type 9 to epidermal growth factor-like repeat a of low-density lipoprotein receptor decreases receptor recycling and increases degradation. J. Biol. Chem. 2007, 282, 18602–18612. [Google Scholar] [CrossRef]
- Jeong, H.J.; Lee, H.S.; Kim, K.S.; Kim, Y.K.; Yoon, D.; Park, S.W. Sterol-dependent regulation of proprotein convertase subtilisin/kexin type 9 expression by sterol-regulatory element binding protein-2. J. Lipid Res. 2008, 49, 399–409. [Google Scholar] [CrossRef]
- Horton, J.D.; Cohen, J.C.; Hobbs, H.H. Molecular biology of PCSK9: Its role in LDL metabolism. Trends Biochem. Sci. 2007, 32, 71–77. [Google Scholar] [CrossRef]
- Costet, P.; Cariou, B.; Lambert, G.; Lalanne, F.; Lardeux, B.; Jarnoux, A.L.; Grefhorst, A.; Staels, B.; Krempf, M. Hepatic PCSK9 expression is regulated by nutritional status via insulin and sterol regulatory element-binding protein 1c. J. Biol. Chem. 2006, 281, 6211–6218. [Google Scholar] [CrossRef]
- Li, H.; Dong, B.; Park, S.W.; Lee, H.S.; Chen, W.; Liu, J. Hepatocyte nuclear factor 1alpha plays a critical role in PCSK9 gene transcription and regulation by the natural hypocholesterolemic compound berberine. J. Biol. Chem. 2009, 284, 28885–28895. [Google Scholar] [CrossRef]


| Cell Lines | Study ID | Natural Product/ Plant Bioactive Compound | Biomarkers | Expression at Effective Concentration | |
|---|---|---|---|---|---|
| Proteins | Genes | ||||
| HUVEC | Wang 2019 [27] | Ginkgolide B ** | PCSK9 | Downregulated | Downregulated |
| LDLR | Upregulated | Upregulated | |||
| ICAM-1 | Downregulated | Downregulated | |||
| VCAM-1 | Downregulated | Downregulated | |||
| SREBP2 | Downregulated | Downregulated | |||
| IL-1α | Downregulated | Downregulated | |||
| IL-1β | Downregulated | Downregulated | |||
| IL-6 | Downregulated | Downregulated | |||
| MCP-1 | Downregulated | Downregulated | |||
| CXCL-1 | Downregulated | Downregulated | |||
| CXCL-2 | Downregulated | Downregulated | |||
| NOX-4 | Downregulated | Downregulated | |||
| LOX-1 | Downregulated | Downregulated | |||
| Huh7 | Mbikay 2014 [28] | Quercetin-3-glucoside ** | PCSK9 | Downregulated | Downregulated |
| LDLR | Upregulated | Upregulated | |||
| SREBP2 | Not reported | Not affected | |||
| Wang 2020 [29] | Ascorbic acid ** | PCSK9 | Downregulated | Downregulated | |
| LDLR | Upregulated | Upregulated | |||
| PPARg | Not affected | Not affected | |||
| FoxO3a | Upregulated | Upregulated | |||
| LO2 | Jing 2019 [30] | Resveratrol ** | PCSK9 | Downregulated | Downregulated |
| LDLR | Upregulated | Upregulated | |||
| SREBP 1c | Downregulated | Downregulated | |||
| HepG2 | Aggrey 2019 [31] | 3R3,14-dihydroangustoline ** | PCSK9 | Downregulated | Not reported |
| LDLR | Upregulated | Not reported | |||
| Ahn 2019 [32] | Erybraedin D ** | PCSK9 | Downregulated | Downregulated | |
| Cameron 2008 [33] | Berberine ** | PCSK9 | Downregulated | Downregulated | |
| Chae 2018 [34] | Saucinone ** | PCSK9 | Not reported | Downregulated | |
| LDLR | Not reported | Upregulated | |||
| Chen 2016 [35] | Tanshinone IIA ** | PCSK9 | Downregulated | Downregulated | |
| LDLR | Upregulated | NSC | |||
| Choi 2017 [36] | Allium fistulosum L. * | PCSK9 | Downregulated | Downregulated | |
| LDLR | Downregulated | Downregulated | |||
| SREBP2 | Downregulated | Downregulated | |||
| HNF1α | Not affected | Downregulated | |||
| Dong 2019 [37] | Siblinin A ** | PCSK9 | Downregulated | Downregulated | |
| HepG2 | Fan 2021 [38] | Berberine derivative (9k) ** | PCSK9 | Downregulated | Not reported |
| LDLR | Upregulated | Not reported | |||
| Gao 2018 [39] | Pinostrobin ** | PCSK9 | Downregulated | Downregulated | |
| LDLR | Upregulated | NSC | |||
| SREBP2 | NSC | Not reported | |||
| HNF1α | NSC | Not reported | |||
| FoxO3a | Upregulated | Not reported | |||
| Fu 2020 [40] | 17β-estradiol (βE2) ** | PCSK9 | Downregulated | Not reported | |
| LDLR | Upregulated | No changed | |||
| Gu 2017 [41] | Lunasin ** | PCSK9 | Downregulated | Downregulated | |
| LDLR | Upregulated | Upregulated | |||
| HNF1α | Not reported | Downregulated | |||
| SREBP2 | Upregulated | Upregulated | |||
| Hwang 2020 [42] | Butein ** | PCSK9 | Downregulated | Downregulated | |
| LDLR | Upregulated | Upregulated | |||
| HNF1α | Downregulated | Downregulated | |||
| SREBP2 | NSC | Downregulated | |||
| HMGCR | Not reported | Downregulated | |||
| Hwang 2021 [43] | Capsellabursa-pastoris * | PCSK9 | Downregulated | Downregulated | |
| LDLR | Not affected | Downregulated | |||
| HNF1α | Downregulated | Downregulated | |||
| SREBP2 | Downregulated | Downregulated | |||
| Kim 2020 [44] | Piceatannol ** | PCSK9 | Downregulated | Downregulated | |
| LDLR | Upregulated | Not affected | |||
| HNF1α | Not reported | Downregulated | |||
| SREBP2 | Not reported | Downregulated | |||
| Lammi 2019 [45] | Lupin peptide T9 ** | PCSK9 | Downregulated | Not reported | |
| LDLR | Upregulated | Not reported | |||
| HNF1α | Downregulated | Not reported | |||
| Li 2020 [46] | 23,24-Dihydrocucurbitacin B ** | PCSK9 | Downregulated | Downregulated | |
| LDLR | Upregulated | Upregulated | |||
| SREBP2 | Upregulated | Not reported | |||
| HNF1α | Downregulated | Not reported | |||
| Masagalli 2021 [47] | Moracin C ** | PCSK9 | Downregulated | Downregulated | |
| Pel 2020 [48] | 5,6,7,4’-tetramethoxyflavanone ** | PCSK9 | Downregulated | Downregulated | |
| LDLR | Upregulated | NSC | |||
| HNF1α | Not reported | Downregulated | |||
| Pel 2017 [49] | (+)-pinoresinol ** | PCSK9 | Downregulated | Downregulated | |
| Weng 2021 [50] | Gynostemma pentaphyllum * | PCSK9 | Downregulated | Downregulated | |
| HepG2 | Wang 2021 [51] | Gypenoside LVI ** | PCSK9 | Downregulated | Downregulated |
| LDLR | Not reported | Not affected | |||
| SREBP2 | Not affected | Not affected | |||
| Wu 2019 [52] | Tetrahydroprotoberberi-ne derivatives ** | PCSK9 | Downregulated | Downregulated | |
| LDLR | Upregulated | Not reported | |||
| Wu 2021 [53] | Diallyl disulfide ** | PCSK9 | Downregulated | Downregulated | |
| LDLR | Upregulated | Upregulated | |||
| SREBP2 | Downregulated | Downregulated | |||
| HMGCR | Downregulated | Downregulated | |||
| HNF1α | Not affected | Not affected | |||
| Yang 2018 [54] | Liraglutide ** | PCSK9 | Downregulated | Downregulated | |
| HNF1α | Downregulated | Downregulated | |||
| Yang 2018 [55] | Chitosan oligosaccharides ** | PCSK9 | Downregulated | Downregulated | |
| SREBP2 | Upregulated | Upregulated | |||
| HNF1α | Upregulated | Upregulated | |||
| FoxO3a | Upregulated | Upregulated | |||
| Lupo 2019 [56] | Monacolin K ** | PCSK9 | Upregulated | Upregulated | |
| LDLR | Upregulated | Not reported | |||
| HMGCR | Not reported | Upregulated | |||
| FAS | Not reported | Upregulated | |||
| Berberine ** | PCSK9 | Downregulated | Downregulated | ||
| LDLR | Upregulated | Not reported | |||
| HMGCR | Not reported | Downregulated | |||
| FAS | Not reported | Downregulated | |||
| 1-deoxynojirimycin ** | PCSK9 | Downregulated | Downregulated | ||
| LDLR | Upregulated | Not reported | |||
| HMGCR | Not reported | Downregulated | |||
| FAS | Not reported | Downregulated | |||
| Wang 2020 [29] | Ascorbic acid ** | PCSK9 | Downregulated | Downregulated | |
| LDLR | Upregulated | Upregulated | |||
| PPARg | Not affected | Not affected | |||
| FoxO3a | Upregulated | Upregulated | |||
| JLM3 | He 2017 [57] | Actinidia chinensis * | PCSK9 | Not reported | Upregulated |
| LDLR | Not reported | Upregulated | |||
| Biomarkers | Cell Lines | Study ID | Model Validity | Imprecision | Biomarker Validity |
|---|---|---|---|---|---|
| PCSK9 | HUVEC | Wang 2019 [27] | Low | Low | Unclear |
| Huh7 | Mbikay 2014 [28] | Low | Low | Low | |
| Wang 2020 [29] | Low | Low | Low | ||
| LO2 | Jing 2019 [30] | Low | Low | Unclear | |
| JLM3 | He 2017 [57] | Unclear | Low | Low | |
| HepG2 | Aggrey 2019 [31] | Unclear | Unclear | Unclear | |
| Ahn 2019 [32] | Unclear | Low | Unclear | ||
| Cameron 2008 [33] | Low | Low | Low | ||
| Chae 2018 [34] | Unclear | Low | Low | ||
| Chen 2016 [35] | Low | Low | Unclear | ||
| Choi et 2017 [36] | Low | Unclear | Low | ||
| Dong 2019 [37] | Unclear | Low | Unclear | ||
| Fan 2021 [38] | Low | Low | Unclear | ||
| Gao 2018 [39] | Low | Low | Low | ||
| Fu 2020 [40] | Unclear | Low | Unclear | ||
| Gu 2017 [41] | Low | Low | Unclear | ||
| Hwang 2020 [42] | Low | Low | Unclear | ||
| Hwang 2021 [43] | Low | Unclear | Low | ||
| Kim 2020 [44] | Low | Low | Low | ||
| Lammi 2019 [45] | Low | Low | Unclear | ||
| Li 2020 [46] | Low | Low | Unclear | ||
| Masagalli 2021 [47] | Low | Low | Unclear | ||
| Pel 2020 [48] | Unclear | Unclear | Unclear | ||
| Pel 2017 [49] | High | Unclear | Low | ||
| Weng 2021 [50] | Low | Unclear | Low | ||
| Wang 2020 [51] | Low | Low | Unclear | ||
| Wu 2019 [52] | Unclear | Low | Low | ||
| Wu 2021 [53] | Low | Low | Unclear | ||
| Yang 2018 [54] | Low | Low | Unclear | ||
| Yang 2018 [55] | Low | Low | Unclear | ||
| Lupo 2019 [56] | Unclear | Low | Low | ||
| Wang 2020 [29] | Low | Low | Low | ||
| LDLR | HUVEC | Wang 2019 [27] | Low | Low | High |
| Huh7 | Mbikay 2014 [28] | Low | Low | Low | |
| Wang 2020 [29] | Low | Low | Low | ||
| LO2 | Jing 2019 [30] | Low | Unclear | Unclear | |
| JLM3 | He 2017 [57] | Unclear | Low | Low | |
| HepG2 | Aggrey 2019 [31] | Unclear | Unclear | Unclear | |
| Cameron 2008 [33] | Low | Low | Low | ||
| Chae 2018 [34] | Unclear | Low | Low | ||
| Chen 2016 [35] | Low | Low | Unclear | ||
| Choi 2017 [36] | Low | Unclear | Unclear | ||
| Fan 2021 [38] | Low | Low | Unclear | ||
| Gao 2018 [39] | Low | Low | Low | ||
| Fu 2020 [40] | Unclear | Low | Unclear | ||
| Gu 2017 [41] | Low | Low | Unclear | ||
| Hwang 2020 [42] | Low | Low | Unclear | ||
| Hwang 2021 [43] | Low | Unclear | Low | ||
| Kim 2020 [44] | Low | Low | Low | ||
| Lammi 2019 [45] | Low | Low | Unclear | ||
| Li 2020 [46] | Low | Low | Low | ||
| Pel 2017 [49] | High | Unclear | Unclear | ||
| Wang 2021 [51] | Low | Low | Unclear | ||
| Wu 2019 [52] | Unclear | Low | Low | ||
| Wu 2021 [53] | Low | Low | Unclear | ||
| Lupo 2019 [56] | Unclear | Low | Low | ||
| Wang 2020 [29] | Low | Low | Low | ||
| SREBP | HUVEC | Wang 2019 [27] | Low | Low | Unclear |
| Huh7 | Mbikay 2014 [28] | Low | Low | Unclear | |
| LO2 | Jing 2019 [30] | Low | Unclear | Unclear | |
| HepG2 | Choi et 2017 [36] | Low | Unclear | Unclear | |
| Gao 2018 [39] | Low | Low | Unclear | ||
| Gu 2017 [41] | Low | Low | Unclear | ||
| Hwang 2020 [42] | Low | Low | Unclear | ||
| Hwang 2021 [43] | Low | Unclear | Low | ||
| Kim 2020 [44] | Low | Low | Unclear | ||
| Li 2020 [46] | Low | Low | Unclear | ||
| Wang 2021 [51] | Low | Low | Unclear | ||
| Wu 2021 [53] | Low | Low | Unclear | ||
| Yang 2018 [55] | Low | Low | Unclear | ||
| HNF1α | HepG2 | Choi 2017 [36] | Low | Unclear | Unclear |
| Gao 2018 [39] | Low | Low | Unclear | ||
| Gu 2017 [41] | Low | Low | Unclear | ||
| Hwang 2020 [42] | Low | Low | Unclear | ||
| Hwang 2021 [43] | Low | Unclear | Low | ||
| Kim 2020 [44] | Low | Low | Unclear | ||
| Li 2020 [46] | Low | Low | Low | ||
| Pel 2020 [48] | Unclear | Unclear | Unclear | ||
| Wu 2021 [53] | Low | Low | Unclear | ||
| Yang 2018 [55] | Low | Low | Unclear | ||
| FoxO3a | Huh7 | Wang 2020 [29] | Low | Low | Low |
| HepG2 | Gao 2018 [39] | Low | Low | Unclear | |
| Yang 2018 [55] | Low | Low | Unclear | ||
| Wang 2020 [29] | Low | Low | Low | ||
| HMGCR | HepG2 | Hwang 2020 [42] | Low | Low | Unclear |
| Wu 2021 [53] | Low | Low | Unclear | ||
| Lupo 2019 [56] | Unclear | Low | Low | ||
| PPARg | Huh7 | Wang 2020 [29] | Low | Low | Unclear |
| HepG2 | Wang 2020 [29] | Low | Low | Unclear | |
| FAS | HepG2 | Lupo 2019 [56] | Unclear | Low | Unclear |
| LOX-1 | HUVEC | Wang 2019 [27] | Low | Low | Unclear |
| NOX-4 | HUVEC | Wang 2019 [27] | Low | Low | Unclear |
| ICAM | HUVEC | Wang 2019 [27] | Low | Low | Unclear |
| VCAM | HUVEC | Wang 2019 [27] | Low | Low | Unclear |
| (IL)-1α, | HUVEC | Wang 2019 [27] | Low | Low | Unclear |
| IL-1β | HUVEC | Wang 2019 [27] | Low | Low | Unclear |
| IL-6 | HUVEC | Wang 2019 [27] | Low | Low | Unclear |
| MCP-1 | HUVEC | Wang 2019 [27] | Low | Low | Unclear |
| CXCL-1 | HUVEC | Wang 2019 [27] | Low | Low | Unclear |
| CXCL-2 | HUVEC | Wang 2019 [27] | Low | Low | Unclear |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zulkapli, R.; Yusof, M.Y.P.M.; Abd Muid, S.; Wang, S.M.; Firus Khan, A.Y.; Nawawi, H. A Systematic Review on Attenuation of PCSK9 in Relation to Atherogenesis Biomarkers Associated with Natural Products or Plant Bioactive Compounds in In Vitro Studies: A Critique on the Quality and Imprecision of Studies. Int. J. Environ. Res. Public Health 2022, 19, 12878. https://doi.org/10.3390/ijerph191912878
Zulkapli R, Yusof MYPM, Abd Muid S, Wang SM, Firus Khan AY, Nawawi H. A Systematic Review on Attenuation of PCSK9 in Relation to Atherogenesis Biomarkers Associated with Natural Products or Plant Bioactive Compounds in In Vitro Studies: A Critique on the Quality and Imprecision of Studies. International Journal of Environmental Research and Public Health. 2022; 19(19):12878. https://doi.org/10.3390/ijerph191912878
Chicago/Turabian StyleZulkapli, Rahayu, Mohd Yusmiaidil Putera Mohd Yusof, Suhaila Abd Muid, Seok Mui Wang, Al’Aina Yuhainis Firus Khan, and Hapizah Nawawi. 2022. "A Systematic Review on Attenuation of PCSK9 in Relation to Atherogenesis Biomarkers Associated with Natural Products or Plant Bioactive Compounds in In Vitro Studies: A Critique on the Quality and Imprecision of Studies" International Journal of Environmental Research and Public Health 19, no. 19: 12878. https://doi.org/10.3390/ijerph191912878
APA StyleZulkapli, R., Yusof, M. Y. P. M., Abd Muid, S., Wang, S. M., Firus Khan, A. Y., & Nawawi, H. (2022). A Systematic Review on Attenuation of PCSK9 in Relation to Atherogenesis Biomarkers Associated with Natural Products or Plant Bioactive Compounds in In Vitro Studies: A Critique on the Quality and Imprecision of Studies. International Journal of Environmental Research and Public Health, 19(19), 12878. https://doi.org/10.3390/ijerph191912878