
Glioma-Associated Microglia Characterization in the Glioblastoma Microenvironment through a ‘Seed-and Soil’ Approach: A Systematic Review

 , ,
, , 
Abstract
1. Introduction
- (a)
- developing new treatments targeting GAMs, thus rendering the tumor once again targetable by host immunity, and slowing its progression and aggressiveness by inhibiting the microglia and tumor cell crosstalk
- (b)
- defining the hystotypes of the tumor and degree of response to treatment based on the GAMs composition and infiltration pattern.
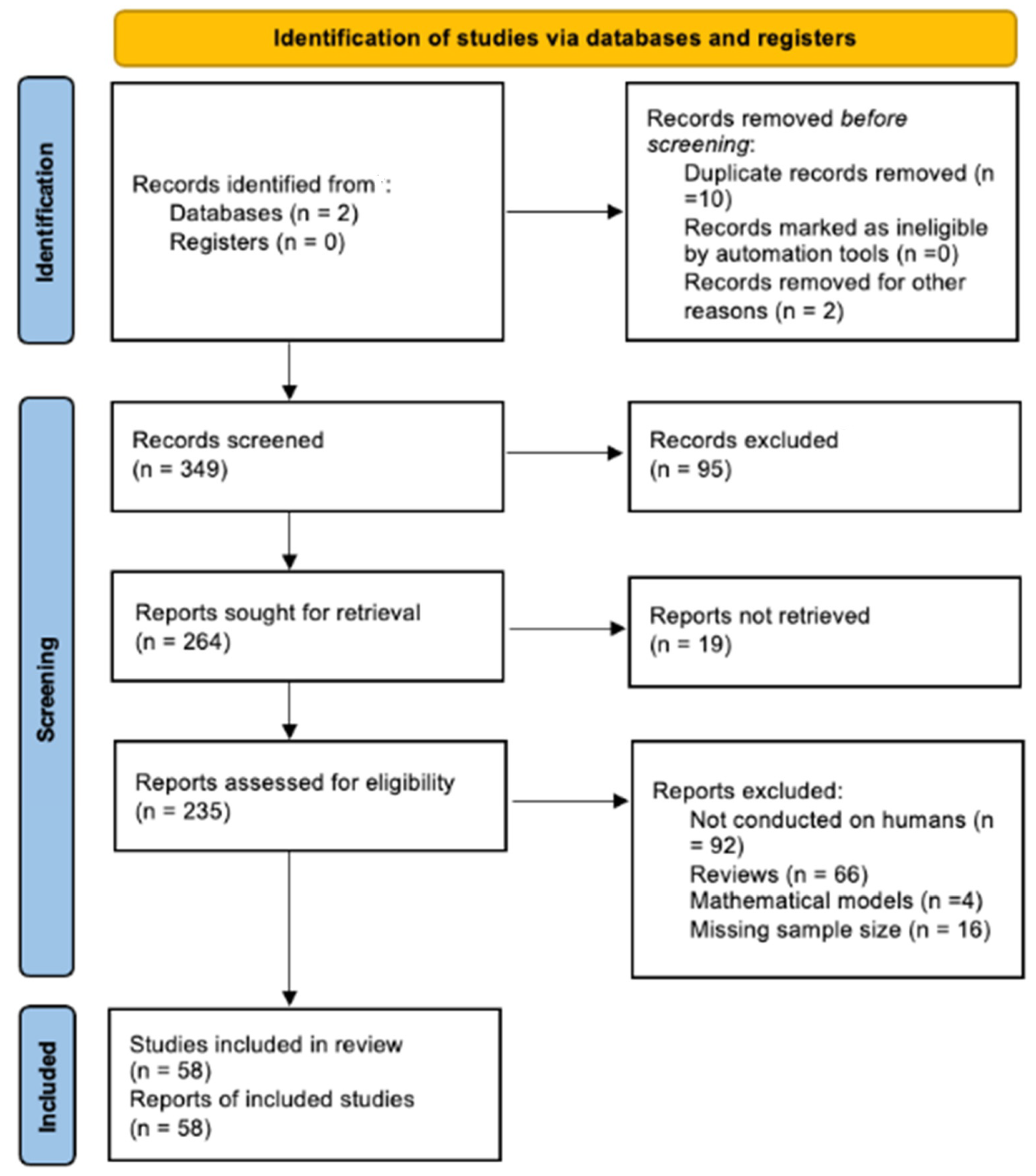
2. Methods
3. Results
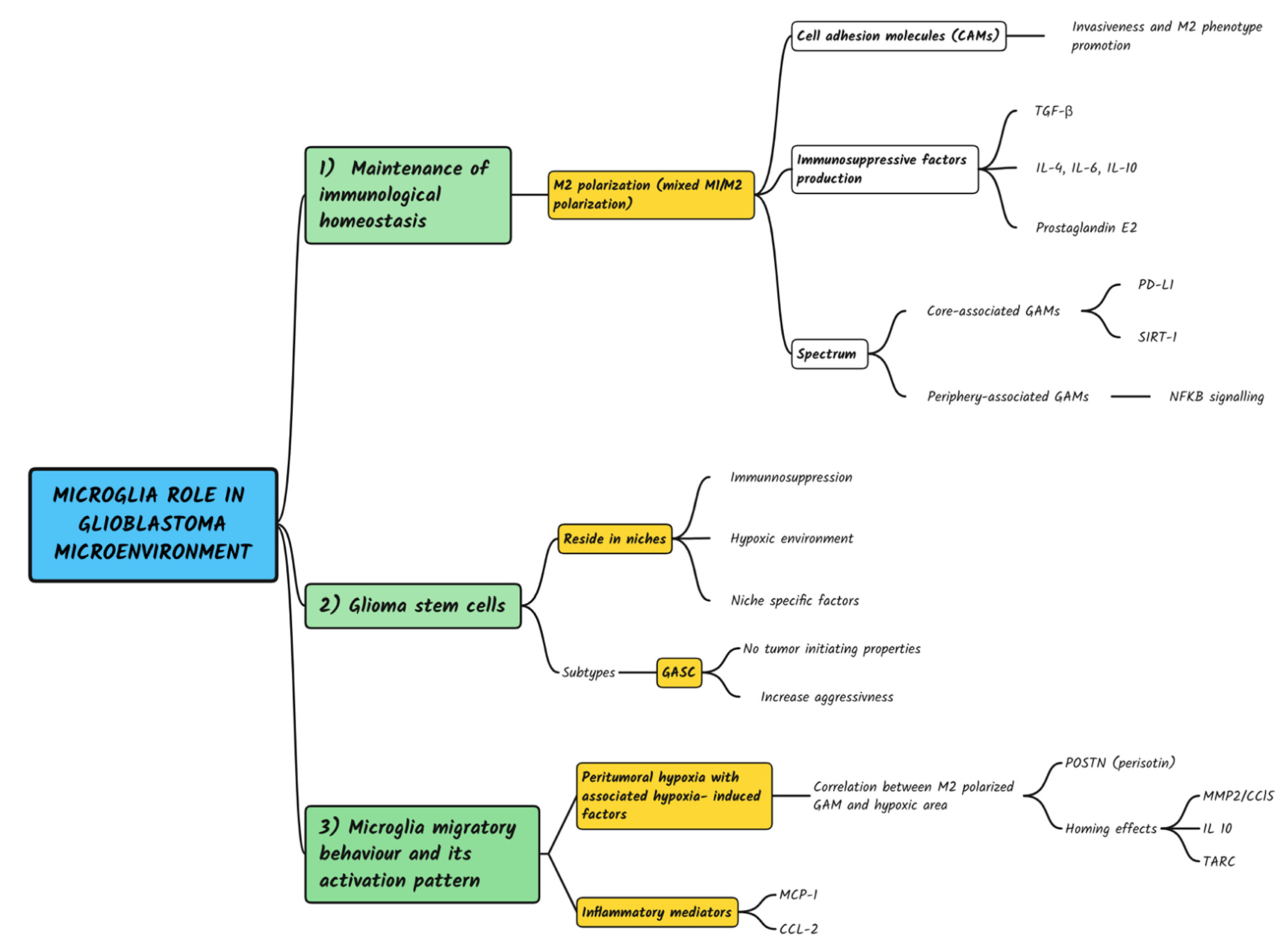
4. Discussion
4.1. Microglia Maintenance of Immunological Homeostasis and Protection against Autoimmunity
4.2. Microglia Crosstalk with Dedifferentiated and Stem-like Glioblastoma Cells
- -
- Host immune system: since immunosuppression is a cardinal feature of malignant tumors.
- -
- Metabolism: GBM is characterized by a hypoxic environment, which in turn increases the need for glycolysis, active in malignant cells, even under aerobic conditions. Hypoxic niches and perivascular niches have been reported both inside and outside of the tumor mass.
- -
- Niche-specific factors: perivascular niches develop along capillaries and arterioles where GASCs are in direct contact with the endothelium. Invasive niches are characterized by perivascular growth of single invasive neoplastic cells along the capillaries, between the endothelium and reactive astrocytes.
4.3. Microglia Migratory Behavior and Its Activation Pattern
4.4. Targeting the Tumor Microenvironment: The Disillusionment with Current Immunotherapeutic Treatment
- -
- Radiotherapy improves the blood-brain barrier (BBB) permeability to chemotherapy; triggers TME remodeling via increased GAM infiltrates and improved GSC radiation resistance by activating DNA damage checkpoints to repair DNA damage.
- -
- Temozolomide (TMZ) triggers a proinvasive TME phenotype by altering proteoglycans and glycosaminoglycans (GAGs) content.
- (1)
- PD-L1 quantitative expression has an impact on survival, independently of gender and age [98].
- (2)
- PD-L1 overexpression is significantly associated with poor OS for patients from Asia and America, while no significant association for the survival of patients from Europe. This “ethnic bias” of PD-L1 has been observed in several clinical studies for patients with other solid tumors, such as KEYNOTE-161 in esophageal squamous cell carcinoma and KEYNOTE-063 in advanced gastric or gastro-esophageal junction cancer [98].
- (3)
- IDH1-wildtype status in glioblastoma was PD-L1 expression positive, suggesting PD-L1/IDH1-wildtype association. From a molecular point of view, it could be that IDH1 mutation results in PD-L1 promoter hypermethylation, thus downregulating the expression of PD-L1. Therefore, PD-L1 immune checkpoint inhibitors analysis might not be advisable because of the globally low PD-L1 expression in patients with IDH1-mutant glioblastomas.
5. Conclusions
- (1)
- Microglia play a paramount role in the maintenance of immunological homeostasis and protection against autoimmunity and its activation pattern at the TME level, polarized toward an M2 phenotype as selected by environmental pressure. This suggests that further investigation of microglia phenotypic characterization at the microenvironment level (M1 vs. M2 phenotype) is needed.
- (2)
- Microglia crosstalk with dedifferentiated and stem-like glioblastoma cells in perivascular and perinecrotic hypoxic niches, where they start crosstalk with the staminal compartment ultimately promoting disease progression and relapse after treatments.
- (3)
- Microglia demonstrate migratory behavior with respect to infiltrative margins of tumor cells. However, there are still many issues to be investigated. While the classification of macrophages or microglial cells into the M1 or M2 polarized state is a well-established approach in most preclinical models, the same is not true in the clinical research setting, because of the high degree of diversity and plasticity shown by these cell types. Therefore, dichotomizing GAMS into M1 and M2 activation status might be over simplistic as, indeed, a clear distinction between these phenotypes cannot be clearly distinguished. The resulting definitions of transcriptomic-based functional phenotypes of GAMs from human and experimental rodent gliomas are conflicting and indicate a mixture of M1 and M2 phenotypes. Cells within the tumor often display a complex pattern of phenotypes, upregulating both M1 and M2 molecular markers, and the prevalence of one phenotype on the other might also depend on the stage of disease [99].
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| APCs | Antigen presenting cells |
| BBB | Blood brain barrier |
| CAMs | cell adhesion molecules |
| CNS | Central nervous system |
| ECM | Extracellular matrix |
| EMT | Epithelial–mesenchymal transition |
| EGFR | epidermal growth factors receptor |
| FDA | Food and Drug Administration |
| GAG | Glycosaminoglycans |
| GAM | Glioma-associated macrophage and microglia |
| GASC | Glioma Associated Stem cells |
| GBM | Glioblastoma |
| GCS | Glioma stem cells |
| MET | mesenchymal–epithelial transition |
| PD1 | Programmed cell Death-1 |
| PRISMA-P | Preferred Reporting Items for Systematic Review and Meta-Analysis Protocols |
| SIRT1 | Sirtuin 1 |
| TCR | T cell receptors |
| TGF-β | Transforming growth factor-β |
| TIL | Tumor-infiltrating lymphocytes |
| TME | Tumor Microenvironment |
| TMZ | Temozolomide |
References
- Prosniak, M.; Harshyne, L.A.; Andrews, D.W.; Kenyon, L.; Bedelbaeva, K.; Apanasovich, T.V.; Heber-Katz, E.; Curtis, M.T.; Cotzia, P.; Hooper, D. Glioma Grade Is Associated with the Accumulation and Activity of Cells Bearing M2 Monocyte Markers. Clin. Cancer Res. 2013, 19, 3776–3786. [Google Scholar] [CrossRef] [PubMed]
- Hattermann, K.; Sebens, S.; Helm, O.; Schmitt, A.D.; Mentlein, R.; Mehdorn, H.M.; Held-Feindt, J. Chemokine expression profile of freshly isolated human glioblastoma-associated macrophages/microglia. Oncol. Rep. 2014, 32, 270–276. [Google Scholar] [CrossRef] [PubMed]
- Zeiner, P.S.; Preusse, C.; Blank, A.-E.; Zachskorn, C.; Baumgarten, P.; Caspary, L.; Braczynski, A.K.; Weissenberger, J.; Bratzke, H.; Reiß, S.; et al. MIF Receptor CD74 is Restricted to Microglia/Macrophages, Associated with a M1-Polarized Immune Milieu and Prolonged Patient Survival in Gliomas. Brain Pathol. 2014, 25, 491–504. [Google Scholar] [CrossRef] [PubMed]
- Sørensen, M.D.; Dahlrot, R.H.; Boldt, H.B.; Hansen, S.; Kristensen, B.W. Tumour-associated microglia/macrophages predict poor prognosis in high-grade gliomas and correlate with an aggressive tumour subtype. Neuropathol. Appl. Neurobiol. 2018, 44, 185–206. [Google Scholar] [CrossRef] [PubMed]
- Zeiner, P.S.; Preusse, C.; Golebiewska, A.; Zinke, J.; Iriondo, A.; Muller, A.; Kaoma, T.; Filipski, K.; Müller-Eschner, M.; Bernatz, S.; et al. Distribution and prognostic impact of microglia/macrophage subpopulations in gliomas. Brain Pathol. 2018, 29, 513–529. [Google Scholar] [CrossRef]
- Lin, G.L.; Nagaraja, S.; Filbin, M.G.; Suvà, M.L.; Vogel, H.; Monje, M. Non-inflammatory tumor microenvironment of diffuse intrinsic pontine glioma. Acta Neuropathol. Commun. 2018, 6, 51. [Google Scholar] [CrossRef]
- Waters, M.R.; Gupta, A.S.; Mockenhaupt, K.; Brown, L.N.; Biswas, D.D.; Kordula, T. RelB acts as a molecular switch driving chronic inflammation in glioblastoma multiforme. Oncogenesis 2019, 8, 37. [Google Scholar] [CrossRef]
- Martinez-Lage, M.; Lynch, T.M.; Bi, Y.; Cocito, C.; Way, G.P.; Pal, S.; Haller, J.; Yan, R.E.; Ziober, A.; Nguyen, A.; et al. Immune landscapes associated with different glioblastoma molecular subtypes. Acta Neuropathol. Commun. 2019, 7, 203. [Google Scholar] [CrossRef]
- González-Tablas Pimenta, M.; Otero, Á.; Arandia Guzman, D.A.; Pascual-Argente, D.; Ruíz Martín, L.; Sousa-Casasnovas, P.; García-Martin, A.; Roa Montes de Oca, J.C.; Villaseñor-Ledezma, J.; Torres Carretero, L.; et al. Tumor cell and immune cell profiles in primary human glioblastoma: Impact on patient outcome. Brain Pathol. 2021, 31, 365–380. [Google Scholar] [CrossRef]
- Fu, W.; Wang, W.; Li, H.; Jiao, Y.; Weng, J.; Huo, R.; Yan, Z.; Wang, J.; Xu, H.; Wang, S.; et al. High Dimensional Mass Cytometry Analysis Reveals Characteristics of the Immunosuppressive Microenvironment in Diffuse Astrocytomas. Front. Oncol. 2020, 10, 78. [Google Scholar] [CrossRef]
- Klemm, F.; Maas, R.R.; Bowman, R.L.; Kornete, M.; Soukup, K.; Nassiri, S.; Brouland, J.-P.; Iacobuzio-Donahue, C.A.; Brennan, C.; Tabar, V.; et al. Interrogation of the Microenvironmental Landscape in Brain Tumors Reveals Disease-Specific Alterations of Immune Cells. Cell 2020, 181, 1643–1660.e17. [Google Scholar] [CrossRef] [PubMed]
- Zinnhardt, B.; Müther, M.; Roll, W.; Backhaus, P.; Jeibmann, A.; Foray, C.; Barca, C.; Döring, C.; Tavitian, B.; Dollé, F.; et al. TSPO imaging-guided characterization of the immunosuppressive myeloid tumor microenvironment in patients with malignant glioma. Neuro-Oncology 2020, 22, 1030–1043. [Google Scholar] [CrossRef] [PubMed]
- Fu, W.; Wang, W.; Li, H.; Jiao, Y.; Weng, J.; Huo, R.; Yan, Z.; Wang, J.; Xu, H.; Wang, S.; et al. CyTOF Analysis Reveals a Distinct Immunosuppressive Microenvironment in IDH Mutant Anaplastic Gliomas. Front. Oncol. 2021, 10, 560211. [Google Scholar] [CrossRef] [PubMed]
- Cai, X.; Yuan, F.; Zhu, J.; Yang, J.; Tang, C.; Cong, Z.; Ma, C. Glioma-Associated Stromal Cells Stimulate Glioma Malignancy by Regulating the Tumor Immune Microenvironment. Front. Oncol. 2021, 11, 672928. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Ma, W.; Fan, W.; Ren, C.; Xu, J.; Zeng, F.; Bao, Z.; Jiang, T.; Zhao, Z. Comprehensive transcriptomic characterization reveals core genes and module associated with immunological changes via 1619 samples of brain glioma. Cell Death Dis. 2021, 12, 1140. [Google Scholar] [CrossRef]
- Cui, X.; Wang, Q.; Zhou, J.; Wang, Y.; Xu, C.; Tong, F.; Wang, H.; Kang, C. Single-Cell Transcriptomics of Glioblastoma Reveals a Unique Tumor Microenvironment and Potential Immunotherapeutic Target Against Tumor-Associated Macrophage. Front. Oncol. 2021, 11, 710695. [Google Scholar] [CrossRef]
- Shen, C.-K.; Huang, B.-R.; Yeh, W.-L.; Chen, C.-W.; Liu, Y.-S.; Lai, S.-W.; Tseng, W.-P.; Lu, D.-Y.; Tsai, C.-F. Regulatory effects of IL-1β in the interaction of GBM and tumor-associated monocyte through VCAM-1 and ICAM-1. Eur. J. Pharmacol. 2021, 905, 174216. [Google Scholar] [CrossRef]
- Fries, G.; Perneczky, A.; Kempski, O. Glioblastoma-associated circulating monocytes and the release of epidermal growth factor. J. Neurosurg. 1996, 85, 642–647. [Google Scholar] [CrossRef]
- Dziurzynski, K.; Wei, J.; Qiao, W.; Hatiboglu, M.A.; Kong, L.-Y.; Wu, A.; Wang, Y.; Cahill, D.; Levine, N.; Prabhu, S.; et al. Glioma-Associated Cytomegalovirus Mediates Subversion of the Monocyte Lineage to a Tumor Propagating Phenotype. Clin. Cancer Res. 2011, 17, 4642–4649. [Google Scholar] [CrossRef]
- Kmiecik, J.; Poli, A.; Brons, N.H.C.; Waha, A.; Eide, G.E.; Enger, P.Ø.; Zimmer, J.; Chekenya, M. Elevated CD3+ and CD8+ tumor-infiltrating immune cells correlate with prolonged survival in glioblastoma patients despite integrated immunosuppressive mechanisms in the tumor microenvironment and at the systemic level. J. Neuroimmunol. 2013, 264, 71–83. [Google Scholar] [CrossRef]
- Shimato, S.; Anderson, L.M.; Asslaber, M.; Bruce, J.N.; Canoll, P.; Anderson, D.E.; Anderson, R.C.E. Inhibition of Caveolin-1 Restores Myeloid Cell Function in Human Glioblastoma. PLoS ONE 2013, 8, e77397. [Google Scholar] [CrossRef] [PubMed]
- Silver, D.J.; Siebzehnrubl, F.; Schildts, M.J.; Yachnis, A.T.; Smith, G.M.; Smith, A.A.; Scheffler, B.; Reynolds, B.A.; Silver, J.; Steindler, D.A. Chondroitin Sulfate Proteoglycans Potently Inhibit Invasion and Serve as a Central Organizer of the Brain Tumor Microenvironment. J. Neurosci. 2013, 33, 15603–15617. [Google Scholar] [CrossRef] [PubMed]
- Su, Z.; Roncaroli, F.; Durrenberger, P.F.; Coope, D.J.; Karabatsou, K.; Hinz, R.; Thompson, G.; Turkheimer, F.E.; Janczar, K.; Du Plessis, D.; et al. The 18-kDa Mitochondrial Translocator Protein in Human Gliomas: An 11C-(R)PK11195 PET Imaging and Neuropathology Study. J. Nucl. Med. 2015, 56, 512–517. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.; Stradmann-Bellinghausen, B.; Savaskan, N.; Regnier-Vigouroux, A. Human Monocyte-Derived Macrophages Exposed to Glioblastoma Cells and Tumor-Associated Microglia/Macrophages Differ in Glutamatergic Gene Expressions. In Glia; Wiley-Blackwell: Hoboken, NJ, USA, 2015; Volume 63, p. E466. [Google Scholar]
- Dijksterhuis, J.P.; Arthofer, E.; Marinescu, V.D.; Nelander, S.; Uhlén, M.; Pontén, F.; Mulder, J.; Schulte, G. High levels of WNT-5A in human glioma correlate with increased presence of tumor-associated microglia/monocytes. Exp. Cell Res. 2015, 339, 280–288. [Google Scholar] [CrossRef]
- Chen, J.E.; Lumibao, J.; Leary, S.; Sarkaria, J.N.; Steelman, A.J.; Gaskins, H.R.; Harley, B.A.C. Crosstalk between microglia and patient-derived glioblastoma cells inhibit invasion in a three-dimensional gelatin hydrogel model. J. Neuroinflamm. 2020, 17, 346. [Google Scholar] [CrossRef]
- Zhu, C.; Mustafa, D.; Zheng, P.-P.; Van Der Weiden, M.; Sacchetti, A.; Brandt, M.; Chrifi, I.; Tempel, D.; Leenen, P.J.M.; Duncker, D.J.; et al. Activation of CECR1 in M2-like TAMs promotes paracrine stimulation-mediated glial tumor progression. Neuro-Oncology 2017, 19, 648–659. [Google Scholar] [CrossRef]
- Wang, Z.; Zhang, C.; Liu, X.; Wang, Z.; Sun, L.; Li, G.; Liang, J.; Hu, H.; Liu, Y.; Zhang, W.; et al. Molecular and clinical characterization of PD-L1 expression at transcriptional level via 976 samples of brain glioma. OncoImmunology 2016, 5, e1196310. [Google Scholar] [CrossRef]
- Leite, D.M.; Zvar Baskovic, B.; Civita, P.; Neto, C.; Gumbleton, M.; Pilkington, G.J. A human co-culture cell model incorporating microglia supports glioblastoma growth and migration, and confers resistance to cytotoxics. FASEB J. 2020, 34, 1710–1727. [Google Scholar] [CrossRef]
- Caponegro, M.D.; Oh, K.; Madeira, M.M.; Radin, D.; Sterge, N.; Tayyab, M.; Moffitt, R.A.; Tsirka, S.E. A distinct microglial subset at the tumor–stroma interface of glioma. Glia 2021, 69, 1767–1781. [Google Scholar] [CrossRef]
- Sadahiro, H.; Kang, K.-D.; Gibson, J.T.; Minata, M.; Yu, H.; Shi, J.; Chhipa, R.R.; Chen, Z.; Lu, S.; Simoni, Y.; et al. Activation of the Receptor Tyrosine Kinase AXL Regulates the Immune Microenvironment in Glioblastoma. Cancer Res. 2018, 78, 3002–3013. [Google Scholar] [CrossRef]
- Couto, M.; Coelho-Santos, V.; Santos, L.; Fontes-Ribeiro, C.; Silva, A.P.; Gomes, C.M.F. The interplay between glioblastoma and microglia cells leads to endothelial cell monolayer dysfunction via the interleukin-6-induced JAK2/STAT3 pathway. J. Cell Physiol. 2019, 234, 19750–19760. [Google Scholar] [CrossRef] [PubMed]
- Gjorgjevski, M.; Hannen, R.; Carl, B.; Li, Y.; Landmann, E.; Buchholz, M.; Bartsch, J.W.; Nimsky, C. Molecular profiling of the tumor microenvironment in glioblastoma patients: Correlation of microglia/macrophage polarization state with metalloprotease expression profiles and survival. Biosci. Rep. 2019, 39, BSR20182361. [Google Scholar] [CrossRef] [PubMed]
- Lisi, L.; Ciotti, G.M.P.; Chiavari, M.; Pizzoferrato, M.; Mangiola, A.; Kalinin, S.; Feinstein, D.L.; Navarra, P. Phospho-mTOR expression in human glioblastoma microglia-macrophage cells. Neurochem. Int. 2019, 129, 104485. [Google Scholar] [CrossRef] [PubMed]
- Chiavari, M.; Ciotti, G.; Canonico, F.; Altieri, F.; Lacal, P.; Graziani, G.; Navarra, P.; Lisi, L. PDIA3 Expression in Glioblastoma Modulates Macrophage/Microglia Pro-Tumor Activation. Int. J. Mol. Sci. 2020, 21, 8214. [Google Scholar] [CrossRef] [PubMed]
- Fu, W.; Wang, W.; Li, H.; Jiao, Y.; Huo, R.; Yan, Z.; Wang, J.; Wang, S.; Wang, J.; Chen, D.; et al. Single-Cell Atlas Reveals Complexity of the Immunosuppressive Microenvironment of Initial and Recurrent Glioblastoma. Front. Immunol. 2020, 11, 835. [Google Scholar] [CrossRef]
- Tan, Y.Q.; Li, Y.T.; Yan, T.F.; Xu, Y.; Liu, B.H.; Yang, J.A.; Yang, X.; Chen, Q.X.; Zhang, H.B. Six Immune Associated Genes Construct Prognostic Model Evaluate Low-Grade Glioma. Front. Immunol. 2020, 11, 3326. [Google Scholar] [CrossRef]
- Wei, J.; Gabrusiewicz, K.; Heimberger, A. The Controversial Role of Microglia in Malignant Gliomas. Clin. Dev. Immunol. 2013, 2013, 285246. [Google Scholar] [CrossRef]
- Senjor, E.; Perišić Nanut, M.; Breznik, B.; Mitrović, A.; Mlakar, J.; Rotter, A.; Porčnik, A.; Lah Turnšek, T.; Kos, J. Cystatin F acts as a mediator of immune suppression in glioblastoma. Cell. Oncol. 2021, 44, 1051–1063. [Google Scholar] [CrossRef]
- Tanaka, S.; Ohgidani, M.; Hata, N.; Inamine, S.; Sagata, N.; Shirouzu, N.; Mukae, N.; Suzuki, S.O.; Hamasaki, H.; Hatae, R.; et al. CD206 Expression in Induced Microglia-Like Cells from Peripheral Blood as a Surrogate Biomarker for the Specific Immune Microenvironment of Neurosurgical Diseases Including Glioma. Front. Immunol. 2021, 12, 670131. [Google Scholar] [CrossRef]
- Nuñez, R.E.; del Valle, M.M.; Ortiz, K.; Almodovar, L.; Kucheryavykh, L. Microglial Cytokines Induce Invasiveness and Proliferation of Human Glioblastoma through Pyk2 and FAK Activation. Cancers 2021, 13, 6160. [Google Scholar] [CrossRef]
- Li, Y.; Dong, X.; Cai, J.; Yin, S.; Sun, Y.; Yang, D.; Jiang, C. SERPINA3 induced by astroglia/microglia co-culture facilitates glioblastoma stem-like cell invasion. Oncol. Lett. 2017, 15, 285–291. [Google Scholar] [CrossRef] [PubMed]
- Urbantat, R.M.; Jelgersma, C.; Brandenburg, S.; Nieminen-Kelhä, M.; Kremenetskaia, I.; Zollfrank, J.; Mueller, S.; Rubarth, K.; Koch, A.; Vajkoczy, P.; et al. Tumor-Associated Microglia/Macrophages as a Predictor for Survival in Glioblastoma and Temozolomide-Induced Changes in CXCR2 Signaling with New Resistance Overcoming Strategy by Combination Therapy. Int. J. Mol. Sci. 2021, 22, 11180. [Google Scholar] [CrossRef] [PubMed]
- Parney, I.F.; Waldron, J.S.; Parsa, A.T. Flow cytometry and in vitro analysis of human glioma-associated macrophages: Laboratory investigation. J. Neurosurg. 2009, 110, 572–582. [Google Scholar] [CrossRef] [PubMed]
- Della Pepa, G.M.; Menna, G. In Reply to the Letter to the Editor Regarding “5-Aminolevulinic Acid False Positives in Cerebral Neuro-Oncology: Not All That Is Fluorescent Is Tumor. A Case-Based Update and Literature Review”. World Neurosurg. 2022, 161, 218–219. [Google Scholar] [CrossRef]
- Guo, X.; Xue, H.; Shao, Q.; Wang, J.; Guo, X.; Chen, X.; Zhang, J.; Xu, S.; Li, T.; Zhang, P.; et al. Hypoxia promotes glioma-associated macrophage infiltration via periostin and subsequent M2 polarization by upregulating TGF-βeta and M-CSFR. Oncotarget 2016, 7, 80521–80542. [Google Scholar] [CrossRef]
- Mignogna, C.; Signorelli, F.; Vismara, M.F.M.; Zeppa, P.; Camastra, C.; Barni, T.; Donato, G.; Di Vito, A. A reappraisal of macrophage polarization in glioblastoma: Histopathological and immunohistochemical findings and review of the literature. Pathol.-Res. Pract. 2016, 212, 491–499. [Google Scholar] [CrossRef]
- Lisi, L.; Ciotti, G.; Braun, D.; Kalinin, S.; Currò, D.; Russo, C.D.; Coli, A.; Mangiola, A.; Anile, C.; Feinstein, D.; et al. Expression of iNOS, CD163 and ARG-1 taken as M1 and M2 markers of microglial polarization in human glioblastoma and the surrounding normal parenchyma. Neurosci. Lett. 2017, 645, 106–112. [Google Scholar] [CrossRef]
- Annovazzi, L.; Mellai, M.; Bovio, E.; Mazzetti, S.; Pollo, B.; Schiffer, D. Microglia immunophenotyping in gliomas. Oncol. Lett. 2017, 15, 998–1006. [Google Scholar] [CrossRef]
- Hide, T.; Shibahara, I.; Kumabe, T. Novel concept of the border niche: Glioblastoma cells use oligodendrocytes progenitor cells (GAOs) and microglia to acquire stem cell-like features. Brain Tumor Pathol. 2019, 36, 63–73. [Google Scholar] [CrossRef]
- Yu-Ju Wu, C.; Chen, C.H.; Lin, C.Y.; Feng, L.Y.; Lin, Y.C.; Wei, K.C.; Huang, C.Y.; Fang, J.Y.; Chen, P.Y. CCL5 of glioma-associated microglia/macrophages regulates glioma migration and invasion via calcium-dependent matrix metalloproteinase 2. Neuro-Oncol. 2020, 22, 253–266. [Google Scholar] [CrossRef]
- Landry, A.P.; Balas, M.; Alli, S.; Spears, J.; Zador, Z. Distinct regional ontogeny and activation of tumor associated macrophages in human glioblastoma. Sci. Rep. 2020, 10, 19542. [Google Scholar] [CrossRef] [PubMed]
- Tang, H.; Liu, W.; Xu, Z.; Zhao, J.; Wang, W.; Yu, Z.; Wei, M. Integrated microenvironment-associated genomic profiles identify LRRC15 mediating recurrent glioblastoma-associated macrophages infiltration. J. Cell. Mol. Med. 2021, 25, 5534–5546. [Google Scholar] [CrossRef] [PubMed]
- Kai, K.; Komohara, Y.; Esumi, S.; Fujiwara, Y.; Yamamoto, T.; Uekawa, K.; Ohta, K.; Takezaki, T.; Kuroda, J.; Shinojima, N.; et al. Macrophage/microglia-derived IL-1β induces glioblastoma growth via the STAT3/NF-κB pathway. Hum. Cell 2022, 35, 226–237. [Google Scholar] [CrossRef] [PubMed]
- Sielska, M.; Przanowski, P.; Wylot, B.; Gabrusiewicz, K.; Maleszewska, M.; Kijewska, M.; Zawadzka, M.; Kucharska, J.; Vinnakota, K.; Kettenmann, H.; et al. Distinct roles of CSF family cytokines in macrophage infiltration and activation in glioma progression and injury response. J. Pathol. 2013, 230, 310–321. [Google Scholar] [CrossRef] [PubMed]
- Butts, C.; Socinski, M.A.; Mitchell, P.L.; Thatcher, N.; Havel, L.; Krzakowski, M.; Nawrocki, S.; Ciuleanu, T.E.; Bosquée, L.; Trigo, J.M.; et al. Tecemotide (L-BLP25) versus placebo after chemoradiotherapy for stage III non-small-cell lung cancer (START): A randomised, double-blind, phase 3 trial. Lancet Oncol. 2014, 15, 59–68. [Google Scholar] [CrossRef]
- Shen, X.; Wang, X.; Shen, H.; Feng, M.; Wu, D.; Yang, Y.; Li, Y.; Yang, M.; Ji, W.; Wang, W.; et al. Transcriptomic Analysis Identified Two Subtypes of Brain Tumor Characterized by Distinct Immune Infiltration and Prognosis. Front. Oncol. 2021, 11, 734407. [Google Scholar] [CrossRef]
- Di Ianni, N.; Maffezzini, M.; Eoli, M.; Pellegatta, S. Revisiting the Immunological Aspects of Temozolomide Considering the Genetic Landscape and the Immune Microenvironment Composition of Glioblastoma. Front. Oncol. 2021, 11, 1–7. [Google Scholar] [CrossRef]
- Anghileri, E.; Patanè, M.; Di Ianni, N.; Sambruni, I.; Maffezzini, M.; Milani, M.; Maddaloni, L.; Pollo, B.; Eoli, M.; Pellegatta, S. Deciphering the Labyrinthine System of the Immune Microenvironment in Recurrent Glioblastoma: Recent Original Advances and Lessons from Clinical Immunotherapeutic Approaches. Cancers 2021, 13, 6156. [Google Scholar] [CrossRef]
- Pyonteck, S.M.; Akkari, L.; Schuhmacher, A.J.; Bowman, R.L.; Sevenich, L.; Quail, D.F.; Olson, O.C.; Quick, M.L.; Huse, J.T.; Teijeiro, V.; et al. CSF-1R inhibition alters macrophage polarization and blocks glioma progression. Nat. Med. 2013, 19, 1264–1272. [Google Scholar] [CrossRef]
- Butovsky, O.; Jedrychowski, M.P.; Moore, C.S.; Cialic, R.; Lanser, A.J.; Gabriely, G.; Koeglsperger, T.; Dake, B.; Wu, P.M.; Doykan, C.E. Erratum: Identification of a unique TGF-β-dependent molecular and functional signature in microglia. Nat. Neurosci. 2014, 17, 1286. [Google Scholar] [CrossRef]
- Pan, Y.; Smithson, L.J.; Ma, Y.; Hambardzumyan, D.; Gutmann, D.H. Ccl5 establishes an autocrine high-grade glioma growth regulatory circuit critical for mesenchymal glioblastoma survival. Oncotarget 2017, 8, 32977–32989. [Google Scholar] [CrossRef]
- Qian, J.; Luo, F.; Yang, J.; Liu, J.; Liu, R.; Wang, L.; Wang, C.; Deng, Y.; Lu, Z.; Wang, Y.; et al. TLR2 Promotes Glioma Immune Evasion by Downregulating MHC Class II Molecules in Microglia. Cancer Immunol. Res. 2018, 6, 1220–1233. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Hu, B.; Hu, X.; Kim, H.; Squatrito, M.; Scarpace, L.; DeCarvalho, A.C.; Lyu, S.; Li, P.; Li, Y.; et al. Tumor Evolution of Glioma-Intrinsic Gene Expression Subtypes Associates with Immunological Changes in the Microenvironment. Cancer Cell 2017, 32, 42–56.e6. [Google Scholar] [CrossRef]
- Feng, X.; Szulzewsky, F.; Yerevanian, A.; Chen, Z.; Heinzmann, D.; Rasmussen, R.D.; Alvarez-Garcia, V.; Kim, Y.; Wang, B.; Tamagno, I.; et al. Loss of CX3CR1 increases accumulation of inflammatory monocytes and promotes gliomagenesis. Oncotarget 2015, 6, 15077–15094. [Google Scholar] [CrossRef] [PubMed]
- Tomaszewski, W.; Sanchez-Perez, L.; Gajewski, T.F.; Sampson, J.H. Brain Tumor Microenvironment and Host State: Implications for Immunotherapy. Clin. Cancer Res. 2019, 25, 4202–4210. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Dube, C.; Gibert, M.; Cruickshanks, N.; Wang, B.; Coughlan, M.; Yang, Y.; Setiady, I.; Deveau, C.; Saoud, K.; et al. The p53 Pathway in Glioblastoma. Cancers 2018, 10, 297. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Sun, Y.; Zhang, Q.; Jin, W.; Gordon, R.E.; Zhang, Y.; Wang, J.; Sun, C.; Wang, Z.J.; Qi, X.; et al. Pro-inflammatory and proliferative microglia drive progression of glioblastoma. Cell Rep. 2021, 36, 109718. [Google Scholar] [CrossRef]
- Silver, D.J.; Lathia, J.D. Therapeutic Injury and Tumor Regrowth: Tumor Resection and Radiation Establish the Recurrent Glioblastoma Microenvironment. EBioMedicine 2018, 31, 13–14. [Google Scholar] [CrossRef]
- Zhang, H.; Du, J.; Wang, H.; Wang, H.; Jiang, J.; Zhao, J.; Lu, H. Comparison of diagnostic values of ultrasound micro-flow imaging and contrast-enhanced ultrasound for neovascularization in carotid plaques. Exp. Ther. Med. 2017, 14, 680–688. [Google Scholar] [CrossRef]
- Caponegro, M.D.; Moffitt, R.A.; Tsirka, S.E. Expression of neuropilin-1 is linked to glioma associated microglia and macrophages and correlates with unfavorable prognosis in high grade gliomas. Oncotarget 2018, 9, 35655–35665. [Google Scholar] [CrossRef]
- Yang, C.; Lee, D.-H.; Mangraviti, A.; Su, L.; Zhang, K.; Zhang, Y.; Zhang, B.; Li, W.; Tyler, B.; Wong, J.; et al. Quantitative correlational study of microbubble-enhanced ultrasound imaging and magnetic resonance imaging of glioma and early response to radiotherapy in a rat model. Med. Phys. 2015, 42, 4762–4772. [Google Scholar] [CrossRef]
- Wei, Q.; Singh, O.; Ekinci, C.; Gill, J.; Li, M.; Mamatjan, Y.; Karimi, S.; Bunda, S.; Mansouri, S.; Aldape, K.; et al. TNFα secreted by glioma associated macrophages promotes endothelial activation and resistance against anti-angiogenic therapy. Acta Neuropathol. Commun. 2021, 9, 67. [Google Scholar] [CrossRef] [PubMed]
- Hsu, J.-M.; Xia, W.; Hsu, Y.-H.; Chan, L.-C.; Yu, W.-H.; Cha, J.-H.; Chen, C.-T.; Liao, H.-W.; Kuo, C.-W.; Khoo, K.-H.; et al. STT3-dependent PD-L1 accumulation on cancer stem cells promotes immune evasion. Nat. Commun. 2018, 9, 1908. [Google Scholar] [CrossRef] [PubMed]
- Manini, I.; Caponnetto, F.; Dalla, E.; Ius, T.; Pepa, G.; Pegolo, E.; Bartolini, A.; Rocca, G.; Menna, G.; Loreto, C.; et al. Heterogeneity Matters: Different Regions of Glioblastoma Are Characterized by Distinctive Tumor-Supporting Pathways. Cancers 2020, 12, 2960. [Google Scholar] [CrossRef] [PubMed]
- Menna, G.; Manini, I.; Cesselli, D.; Skrap, M.; Olivi, A.; Ius, T.; Della Pepa, G.M. Immunoregulatory effects of glioma-associated stem cells on the glioblastoma peritumoral microenvironment: A differential PD-L1 expression from core to periphery? Neurosurg. Focus 2022, 52, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Altieri, R.; Barbagallo, D.; Certo, F.; Broggi, G.; Ragusa, M.; Di Pietro, C.; Caltabiano, R.; Magro, G.; Peschillo, S.; Purrello, M.; et al. Peritumoral Microenvironment in High-Grade Gliomas: From FLAIRectomy to Microglia–Glioma Cross-Talk. Brain Sci. 2021, 11, 200. [Google Scholar] [CrossRef]
- Yuan, J.X.; Bafakih, F.F.; Mandell, J.W.; Horton, B.J.; Munson, J.M. Quantitative Analysis of the Cellular Microenvironment of Glioblastoma to Develop Predictive Statistical Models of Overall Survival. J. Neuropathol. Exp. Neurol. 2016, 75, 1110–1123. [Google Scholar] [CrossRef][Green Version]
- Zhang, Q.; Wang, J.; Yao, X.; Wu, S.; Tian, W.; Gan, C.; Wan, X.; You, C.; Hu, F.; Zhang, S.; et al. Programmed Cell Death 10 Mediated CXCL2-CXCR2 Signaling in Regulating Tumor-Associated Microglia/Macrophages Recruitment in Glioblastoma. Front. Immunol. 2021, 12, 1933. [Google Scholar] [CrossRef]
- Schiffer, D.; Mellai, M.; Bovio, E.; Bisogno, I.; Casalone, C.; Annovazzi, L. Glioblastoma niches: From the concept to the phenotypical reality. Neurol. Sci. 2018, 39, 1161–1168. [Google Scholar] [CrossRef]
- Schiffer, D.; Annovazzi, L.; Casalone, C.; Corona, C.; Mellai, M. Glioblastoma: Microenvironment and Niche Concept. Cancers 2018, 11, 5. [Google Scholar] [CrossRef]
- Hide, T.; Komohara, Y.; Miyasato, Y.; Nakamura, H.; Makino, K.; Takeya, M.; Kuratsu, J.-I.; Mukasa, A.; Yano, S. Oligodendrocyte Progenitor Cells and Macrophages/Microglia Produce Glioma Stem Cell Niches at the Tumor Border. EBioMedicine 2018, 30, 94–104. [Google Scholar] [CrossRef] [PubMed]
- Westphal, M.; Maire, C.L.; Lamszus, K. EGFR as a Target for Glioblastoma Treatment: An Unfulfilled Promise. CNS Drugs 2017, 31, 723–735. [Google Scholar] [CrossRef]
- Wenger, K.J.; Wagner, M.; You, S.-J.; Franz, K.; Harter, P.N.; Burger, M.C.; Voss, M.; Ronellenfitsch, M.W.; Fokas, E.; Steinbach, J.P.; et al. Bevacizumab as a last-line treatment for glioblastoma following failure of radiotherapy, temozolomide and lomustine. Oncol. Lett. 2017, 14, 1141–1146. [Google Scholar] [CrossRef] [PubMed]
- Rath, B.H.; Wahba, A.; Camphausen, K.; Tofilon, P.J. Coculture with astrocytes reduces the radiosensitivity yof glioblastoma stem-like cells and identifies additional targets for radiosensitization. Cancer Med. 2015, 4, 1705–1716. [Google Scholar] [CrossRef] [PubMed]
- Litak, J.; Mazurek, M.; Grochowski, C.; Kamieniak, P.; Roliński, J. PD-L1/PD-1 Axis in Glioblastoma Multiforme. Int. J. Mol. Sci. 2019, 20, 5347. [Google Scholar] [CrossRef] [PubMed]
- He, X.; Xu, C. Immune checkpoint signaling and cancer immunotherapy. Cell Res. 2020, 30, 660–669. [Google Scholar] [CrossRef]
- Scheffel, T.B.; Grave, N.; Vargas, P.; Diz, F.M.; Rockenbach, L.; Morrone, F.B. Immunosuppression in Gliomas via PD-1/PD-L1 Axis and Adenosine Pathway. Front. Oncol. 2021, 10, 617385. [Google Scholar] [CrossRef]
- Yao, Y.; Tao, R.; Wang, X.; Wang, Y.; Mao, Y.; Zhou, L.F. B7-H1 is correlated with malignancy-grade gliomas but is not expressed exclusively on tumor stem-like cells. Neuro-Oncology 2009, 11, 757–766. [Google Scholar] [CrossRef]
- Wilmotte, R.; Burkhardt, K.; Kindler, V.; Belkouch, M.-C.; Dussex, G.; De Tribolet, N.; Walker, P.R.; Dietrich, P.-Y. B7-homolog 1 expression by human glioma: A new mechanism of immune evasion. Neuro Report 2005, 16, 1081–1085. [Google Scholar] [CrossRef]
- Xue, S.; Hu, M.; Iyer, V.; Yu, J. Blocking the PD-1/PD-L1 pathway in glioma: A potential new treatment strategy. J. Hematol. Oncol. 2017, 10, 81. [Google Scholar] [CrossRef]
- Li, X.; Yan, S.; Yang, J.; Wang, Y.; Lv, C.; Li, S.; Zhao, J.; Yang, Y.; Zhuo, M.; Wu, N. Efficacy and Safety of PD-1/PD-L1 Inhibitors Plus Chemotherapy Versus PD-1/PD-L1 Inhibitors in Advanced Non-Small Cell Lung Cancer: A Network Analysis of Randomized Controlled Trials. Front. Oncol. 2021, 10, 574752. [Google Scholar] [CrossRef] [PubMed]
- Hao, C.; Chen, G.; Zhao, H.; Li, Y.; Chen, J.; Zhang, H.; Li, S.; Zhao, Y.; Chen, F.; Li, W.; et al. PD-L1 Expression in Glioblastoma, the Clinical and Prognostic Significance: A Systematic Literature Review and Meta-Analysis. Front. Oncol. 2020, 10, 1015. [Google Scholar] [CrossRef] [PubMed]
- Nduom, E.K.; Wei, J.; Yaghi, N.K.; Huang, N.; Kong, L.-Y.; Gabrusiewicz, K.; Ling, X.; Zhou, S.; Ivan, C.; Chen, J.Q.; et al. PD-L1 expression and prognostic impact in glioblastoma. Neuro-Oncology 2015, 18, 195–205. [Google Scholar] [CrossRef] [PubMed]
- Kelly, W.J.; Giles, A.J.; Gilbert, M. T lymphocyte-targeted immune checkpoint modulation in glioma. J. Immunother. Cancer 2019, 8, e000379. [Google Scholar] [CrossRef] [PubMed]
- Litak, J.; Grajkowska, W.; Szumiło, J.; Krukow, P.; Maciejewski, R.; Roliński, J.; Grochowski, C. PD-L1 Expression Correlated with p53 Expression in Pediatric Glioblastoma Multiforme. Brain Sci. 2021, 11, 262. [Google Scholar] [CrossRef]
- Alexander, B.M.; Ba, S.; Berger, M.S.; Berry, D.A.; Cavenee, W.K.; Chang, S.M.; Cloughesy, T.F.; Jiang, T.; Khasraw, M.; Li, W. GBMAGILE Network. Adaptive global innovative learning environment for glioblastoma: GBMAGILE. Clin. Cancer Res. 2018, 24, 737–743. [Google Scholar] [CrossRef]
- Alexander, B.M.; Trippa, L.; Gaffey, S.C.; Arrillaga, I.; Lee, E.Q.; Tanguturi, S.K.; Ahluwalia, M.S.; Colman, H.; Galanis, E.; De Groot, J.F. Individualized screening trial of innovative glioblastoma therapy (INSIGhT). J. Clin. Oncol. 2017, 35, TPS2079. [Google Scholar] [CrossRef]
- Walentynowicz, K.A.; Ochocka, N.; Pasierbińska, M.; Wojnicki, K.; Stepniak, K.; Mieczkowski, J.; Ciechomska, I.; Kaminska, B. In Search for Reliable Markers of Glioma-Induced Polarization of Microglia. Front. Immunol. 2018, 9, 1329. [Google Scholar] [CrossRef]
- Szulzewsky, F.; Pelz, A.; Feng, X.; Synowitz, M.; Markovic, D.; Langmann, T.; Holtman, I.R.; Wang, X.; Eggen, B.J.L.; Boddeke, H.W.G.M.; et al. Glioma-Associated Microglia/Macrophages Display an Expression Profile Different from M1 and M2 Polarization and Highly Express Gpnmb and Spp1. PLoS ONE 2015, 10, e0116644. [Google Scholar] [CrossRef]
- La Rocca, G.; Sabatino, G.; Menna, G.; Altieri, R.; Ius, T.; Marchese, E.; Olivi, A.; Barresi, V.; Della Pepa, G.M. 5-Aminolevulinic Acid False Positives in Cerebral Neuro-Oncology: Not All That Is Fluorescent Is Tumor. A Case-Based Update and Literature Review. World Neurosurg. 2020, 137, 187–193. [Google Scholar] [CrossRef]
- Mieczkowski, J.; Kocyk, M.; Nauman, P.; Gabrusiewicz, K.; Sielska, M.; Przanowski, P.; Maleszewska, M.; Rajan, W.D.; Pszczolkowska, D.; Tykocki, T.; et al. Down-regulation of IKKβ expression in glioma-infiltrating microglia/macrophages is associated with defective inflammatory/immune gene responses in glioblastoma. Oncotarget 2015, 6, 33077–33090. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Author | Year (dd-mm-yyyy) | Type of Study | Sample Size | Microglia Population | Sample | Outcome | Observations |
|---|---|---|---|---|---|---|---|
| Prosniak et al. [1] | 5 June 2013 | Genomics and immunohistochemistry | 143 | Monocytes | Primary tumor or epilepsy samples as control | Grade II and IV astrocytomas can be clearly differentiated based on the expression of certain M2 markers (CD14, CD68, CD163, and CD204, which appear to be elevated, MDSC and CD15, which instead are reduced) in tumor tissues, whereas grade III astrocytomas exhibit a range of expression between the lower and higher grade specimens. The content of CD163þ cells distinguishes grade III astrocytoma subsets with different prognoses. | Levels of mRNAs specific for IL-10, TGF-β1, and monocyte and M2 phenotypic markers were found to concordantly increase with WHO glioma grade. |
| Hatterman et al. [2] | 23 May 2014 | Proteomics/ immunofluorescence | 20 humans | Macrophages | Tumor only | The mean expression of certain chemokines in GAMs (This population is characterized by CD11b+/CD45dim (microglia) and CD11b+/CD45high (macro- phages) phenotypes) is higher than in in vitro polarized M1 and M2 macrophages (M2-macrophages show elevated expression of the mannose (CD204/206) and scavenger receptor (CD163) and TGF-β1 and IL-10. M1-macrophages show L-1β, tumor necrosis factor (TNF)-α, interleukin (IL)-6 or IL-12 along with the ability to induce Th1-mediated immune responses) and displayed a very heterogeneous expression pattern with greater differences between single TAM-enriched fractions. | Comparison between TAMs and in vitro polarized M1- and M2-macrophages based on the expression of specific markers and chemokines. |
| Zeiner et al. [3] | 29 August 2014 | Cellular culture and histopathology | 303 | Macrophages | Tumor core, infiltration zone, and normal-appearing brain | This study provides evidence that CD74 expression is restricted to GAMs in gliomas contrasting findings of several other peripheral tumor entities. | As CD74 protein expression was absent from glioma cells but strongly present in glioma whole tissue, it was hypothesized that CD74 expression derives from non-neoplastic cells of the glioma microenvironment. |
| Meiczkowski et al. [4] | 29 September 2015 | Genomics | 49 | Macrophages | Not specified | The study uncovers new mechanisms by which defective ikkβ/nfκb signaling in myeloid cells affects antitumor immunity and facilitates glioblastoma progression. | Computational analysis of the immune response and TLR signaling genes led to the main finding of the reduced expression of IKBKB (a gene coding for ikkβ) in high-grade gliomas. |
| Zeiner et al. [5] | 3 December 2018 | Cellular culture and histopathology | 344 | Macrophages | Microarrays | This study provides evidence that the immune polarization phenotype of GAMs (Microglia/macrophage (M/M) are identified through CD68, CD163, CD206, and Iba-1. GAMs were isolated using CD11b and later characterized using Iba1, MCHII and CD45) might be distinct from TAMs in non-CNS cancers. | This study identified a rather mixed polarization phenotype with parallel expression of presumptive M1 and M2 markers and without evidence for an unequivocal M1 (CCL2, CCL4, CCL3, IL1B, TLR2, CD86, CCL5 and PTGS2) or M2 (MRC1, LGMN, IL10, MSR1, CD14, CD163) polarization state of GAMs according to the M1 and M2 model. |
| Lin et al. [6] | 28 June 2018 | Genomics | Not specified | Macrophages | Tumor | In contrast to adult GBM, the immune microenvironment of DIPG is noninflammatory and does not contain a significant adaptive immune component. | The data presented here indicate that DIPG-associated macrophages are strikingly less inflammatory compared to adult GBM-associated macrophages. |
| Waters et al. [7] | 29 May 2019 | Cellular culture | Not specified | Macrophages and astrocytes | Not specified | Although relb coordinates anti-inflammatory feedback in astrocytes, this mechanism does not function in GBM cells due to both the limited activity of SIRT1 and the presence of YY1 in the nuclei. | These data strongly support the idea that p52-independent relb signaling is critical in GBM development or progression. |
| Martinez-Lage et al. [8] | 29 November 2019 | Immunohistochemistry | 98 | Monocytes and T cells | Tumor | GBMs demonstrate high levels of intratumor heterogeneity in immune infiltrate, GBMs subtypes vary significantly in the percentage of immune cells in their microenvironment, and mesenchymal GBMs have the highest percentage of microglia, macrophage, and lymphocyte infiltration. | Found trends across the GBM subtypes for most cell types, there was also significant variation within the subtypes. The results demonstrate that the mesenchymal subtype has the highest degree of immune infiltration for all the populations evaluated. Thus, mesenchymal GBM may be the most immunogenic. |
| González-tablas et al. [9] | 3 December 2020 | Flow cytometry | 55 | All immune cells | Tumor | This study shows that immune cell infiltrates, which consist of distinct immune profiles, are systematically present at highly variable levels in GBM. | Combined staining for GFAP and CD45, allowed clear-cut discrimination between GFAP+CD45- GBM tumor and normal astrocytic cells and GFAP-CD45+immune cells. |
| Fu et al. [10] | 4 February 2020 | Mass cytometry | 14 | Monocytes and T cells | Tumor and peripheral blood | The immune cell composition of DA and OG was similar, and T cells in both diseases showed similar exhaustion characteristics. However, GAMs in DAs expressed higher levels of VEGF and TGF-β and exhibited more adverse immune-inhibitory characteristics than OGs. | The enrichment of exhausted T cell subpopulations, recruitment of Tregs, and the strong protumor phenotype of GAMs together contribute to the immunosuppressive microenvironment in DAs. GAMs of das exhibit more inhibitory characteristics than those of OGs. |
| Klemm et al. [11] | 28 May 2020 | Cellular culture, genomics, proteinomics, and more | Not specified | Macrophages | Not specified | By exploring the broad immune landscape, several pronounced differences between gliomas and BrMs were uncovered when directly compared side by side. | Gliomas contain an abundance of TAMs, whereas T cells were much fewer, particularly in IDH mut tumors. The clinical BrM samples showed a pronounced accumulation of lymphocytes and neutrophils. |
| Zinnhardt et al. [12] | 12 February 2020 | Imaging and histopathology | 9 | Myeloid cells | Tumor | A novel preoperative imaging protocol, including CE MRI, [18F]FET-, and [18F]DPA-714-PET-MRI, may allow comprehensive characterization of glioma extent and the heterogeneity of the immune tumor microenvironment in LGG and HGG. | The combination with the commonly used tracer [18F]FET provided complementary information about the glioma microenvironment, the degree of GAM infiltration and differential spatial extent, and the degree of tracer uptake. |
| Fu et al. [13] | 4 February 2021 | Mass cytometry | 15 | Macrophages | Tumor and peripheral blood | In this study, using cytof analyses, infiltrating immunocytes from surgically resected initial AG tissues, including aamut and AOD samples were analyzed. | It was verified that mononuclear phagocytes and T cells were the most abundant groups in the immune microenvironment of ags. Compared to that in the PBMCs, the ratios of immune checkpoint-positive-exhausted CD4+ T cells and CD8+ T cells were distinctly higher at the AG tumor sites |
| Cai et al. [14] | 29 April 2021 | Genomics | 1395 | Macrophages | Databases from TCGA and CCGA | The GASC is an important cell type in the microenvironment and might influence immunotherapy responses. | A strong correlation between the GASC score and tumorigenic cytokine score was found, indicating the tumor-supporting function of GASCs. |
| Y. Zhang et al. [15] | 8 December 2021 | Transcriptomics | 1619 cases from three datasets | Macrophages | Not specified | An immune gene signature was constructed and showed a favorable efficiency in predicting the prognosis of patients with glioma. A high-risk score of this signature predicted enrichment of macrophages and less response to ICB therapy in glioma. | The higher complexity of immune cells was shown in high-risk patients, and the risk score was significantly positively correlated with the abundance of macrophages. Genes in our signature are highly expressed in macrophages by an open single-cell sequencing dataset of GBM. Patients with a high-risk score showed enrichment of macrophage in a tumor-supportive state, but not an antitumor state. |
| Cui et al. [16] | 9 August 2021 | Genomics | 9 humans | TAMs | Not specified | The comprehensive characterization of immune cells from a total of nine GBM tissues revealed a unique immune landscape in GBM at the single-cell level and identified SPI1 as a potential immunotherapeutic target against TAMs in GBM. | The study’s results confirm that TAMs may occupy over one-third of the GBM tumor, with the ratio of the bone marrow-derived macrophages and the brain-resident microglia being 2:1, but infiltrating T lymphocytes comprising less than 2% of the tumor mass. SPI1 was a vital regulon in all states of TAMs |
| Shen et al. [17] | 15 October 2021 | Transcriptomics | 3810 | Monocytes and T cells | Datasets from NCBI, ICGC, CGGA | This reveals two molecular subtypes (i.e., C1/2) of brain tumors featured by distinct immune infiltration signatures and prognosis. | The C1/2 subtypes can distinguish glioma patients with different prognoses stratified by histology, tumor grade, and genomic alteration. In addition, the C1/2 subtypes can also reflect differences in microvascular infiltration, distant metastasis, and the radio-chemotherapy response of patients. |
| Author | Year (dd-mm-yyyy) | Type of Study | Sample Size | Microglia Population | Sample | Outcome | Observations |
|---|---|---|---|---|---|---|---|
| Fries et al. [18] | 10 January 1996 | Cellular culture | 26 | Monocytes | Peripheral blood | Demonstrated that glioblastoma-associated peripheral blood monocytes release substantially larger amounts of EGF even over a long period (100 days) than monocytes from healthy individuals or patients with nontumorous diseases. | First study that demonstrated that glioblastoma-associated circulating monocytes release EGF. This study confirms findings that glioblastomas retain large numbers of EGFR-positive cells and large numbers of tumor-infiltrating monocytes as well. |
| Dziurzynski et al. [19] | 13 April 2011 | Cellular culture | Not specified | Macrophages | Not specified | The data argue against the idea that CMV merely plays a bystander role in glioblastoma pathology but is contributing to oncogenesis. | A tropism for CMV antigen expression was found, specifically pp65, in the GCSCs and MS microglia. Finding of production of CMV IL-10 by the GCSCs and its subsequent effect on the MS microglia precursor, the monocyte. CMV IL-10 activates IE1 in monocytes and there is conversion to the immunosuppressive phenotype M2. |
| J. Kmiecik et al. [20] | 22 August 2013 | Histopathological | 65 | T cells | Not specified | This study showed the beneficial role of immune cell infiltration into the tumor in GBM patients, despite multiple mechanisms of tumor immune escape. | Significant positive correlation of increased cd3+ and cd8+ cellular infiltration into the tumor with improved patient survival. |
| Shimato et al. [21] | 9 October 2013 | Cellular culture | 13 | Monocytes | Tumors and brains from healthy donors | These data indicate that GBM-mediated suppression of tumor-associated myeloid cell function is mediated at least in part by CAV1, and importantly, that activity can be restored by suppressing CAV1. | Upregulation of both AXL and Caveolin-1 (CAV1) has been demonstrated to reduce inflammation via inhibition of TNF- alpha production and was therefore investigated further. There was significant upregulation of CAV1 and AXL when monocytes were stimulated with LPS in the presence of GBMs compared to stimulated monocytes alone as measured by quantitative PCR. |
| Silver et al. [22] | 25 September 2013 | Cellular culture | 4 | Not specified | Tumor | The presence of heavily glycosylated, microenvironmental cspgs inversely correlates with the invasive character of human glioma. | Invasion of high-grade glioma occurs in the absence of a CS-GAG-rich inhibitory matrix. The absence of glycosylated cspgs provides favorable conditions for diffuse infiltration that typifies high-grade glioma. |
| Su et al. [23] | 26 February 2015 | Imaging and histopathology | 22 | GAMs | Tumor only | This study proved that TSPO imaging has the potential to detect early anaplastic transformation of lggs, and it can be effective to stratify patients with glioma who are suitable for TSPO-targeted treatment. | TSPO is predominantly expressed in neoplastic cells, with GAMs only partially contributing to PET signal and no expression in reactive astrocytes. |
| Choi et al. [24] | 5 June 2015 | Genomics | 10 | Macrophages | Not specified | The data suggest a putative neuroprotective role of TAMs by taking up excess extracellular glutamate by increasing expressions of glutamate transporters and glutamine synthetase. | This is an immunosuppressive, anti-inflammatory milieu that led to alterations in gene expression of glutamate receptors and transporters as well as GS (glutamine synthase) that were found to differ between TAMs and mdms. |
| J.P. Dijksterhuis et al. [25] | 25 October 2015 | Immunohistochemistry | 48 | Macrophages | Tumor core and eight cores of normal tissue | The results show that WNT-5A is the only re- presentative of the WNT family upregulated in human glioma compared to nonmalignant control brain tissue. | There is an approximately 4-fold increase in WNT-5A expression between nonmalignant control brain tissue and GBM patients’ brain tissue. The highest levels of WNT-5A were found in mesenchymal GBM. |
| M. D. Sørensen et al. [26] | 2 August 2017 | Histopathological | 314 from different cohorts | Not specified | Tumor, tumor microarrays | Results demonstrate that the prognostic impact of TAMs in gliomas does not depend on the total amount of TAMs but on their acquired functional phenotype. | High levels of the M2-related marker CD204 correlated with increasing malignancy grade and poor patient survival in grades III and IV independently of clinical-pathological parameters. CD204+ TAMs were associated with a more aggressive tumor subtype and expressed proteins that could enable tumor progression. |
| Zhu et al. [27] | 4 January 2017 | Cellular culture | Not specified | Macrophages | From biobank | Demonstration of CECR1-mediated crosstalk mechanism between macrophages and glioma cells. | The influence of CECR1 on immune cells, and on macrophage polarization especially, seems to predominantly impact the systemic and cerebral vasculature and could affect the tumor vasculature in GBM. Expression analysis of typical M1 and m2a, b, and c subtype markers validated the presence of these TAM subtypes in human gliomas. |
| Wang et al. [28] | 10 July 2017 | Transcriptomics | 53 | Monocytes and T cells | Tumor | This study defines a strategy to determine transcriptional subtypes and associates expression subtypes to the tumor-associated immuno-environment. | The transcriptional glioma subtypes, defined through clustering based on tumor-intrinsic genes, strongly over lapped with the PN, CL, and MES subtypes, but identified the NE subtype as normal NE lineage contamination. |
| Li et al. [29] | 26 October 2017 | Cellular culture and histopathology | 80 | Microglia | Tumor and three healthy controls | Results indicate that SERPINA3 could play a critical role in glioma initiation and progression process. | Examination of the expression pattern of SERPINA3 at the various stages of glioma progression, including normal brain, pilocytic astrocytomas, diffuse astrocytomas, oligodendrocytes astrocytoma, anaplastic astrocytomas, and glioblastomas multiforme. |
| Caponegro et al. [30] | 2 November 2018 | Genomics | 667 from TCGA | Macrophages | RNA-seq data | NRP1 expression is correlated with poor prognosis and glioma grade, and associates with the mesenchymal GB subtype. In human GB, NRP1 expression is highly correlated with markers of monocytes and macrophages, as well as genes that contribute to the protumorigenic phenotype of these cells. | Functional gene analysis suggests that NRP1 is associated with markers of monocytic infiltration and protumorigenic GAMs in human GB. The results demonstrate that both LGG and GB patients with high NRP1 expression have enriched monocytic, macrophage, and M2 macrophage populations. AIF1 and ITGAM (Iba1 and CD11b, respectively) are pan markers of monocytes, macrophages, and microglia, and are highly upregulated across human GB subtypes. Significantly correlated with genes that characterize the M2 pro-tumorigenic GAM signature, such as Adm and Mrc1. Also, significantly correlated with TMEM119 and TMEM173. |
| Sadahiro et al. [31]. | 12 March 2018 | Cellular culture and xenograft | Not specified | Not specified | Patient-derived GBM (neuro)sphere cultures | This study identified that tumor-associated MG/Mø cells produce PROS1 that binds and activates AXL in GSCS. The PROS1/AXL signaling axis in GSCS subsequently activates the tumor-intrinsic nfkb pathway to promote the MES phenotype of GBM tumors. | Negative correlation between the expression of PDGFRA and PAXL in GSCS and GBM tumors. High AXL and PROS1 expression is associated with a poor prognosis in patients with GBM. The study demonstrates a role for PROS1-mediated AXL signaling in cancer, which is negatively regulated by pdgfra. |
| Couto et al. [32] | 6 March 2019 | Cellular culture | Not specified | GAMs | Cell lines from biobank | The data unravel a new oncogenic role for IL-6 in GBM through direct effects in microvascular ECs and regulation of endothelial permeability with a negative impact on brain barrier functions. | Data demonstrated significant alterations in the barrier properties of human ECs (endothelial cells) when exposed to cm-cc (conditioned medium of coculture), namely an increase in the permeability to smaller (4 kda) and larger (70 kda) molecular size dextrans and a decrease in the teer (transendothelial electric resistance) values. |
| Gjorgjevski et al. [33] | 20 June 2019 | Qpcr | 20 | Macrophages | Tumor | This study establishes M1- and M2-like markers CXCL10 and CCL13 for informative and reliable detection of GBM-associated microglia and macrophage polarization in conjunction with a defined protease profile as molecular determinants for GBM progression. | It was found that MMP9 and MMP14 are negatively correlated with GBM patient survival and associated with the markers to define a more M2-like microglia and macrophage phenotype. |
| L. Lisi et al. [34] | 10 June 2019 | Histopathological | 42 | Macrophages | Tumor core and tumor periphery | The mTOR pathway is activated in 39% of microglia–macrophage within GBM tumors, compared to 21% in peripheral tissues. | The mTOR pathway is fully activated in microglia cells under conditions mimicking the human GBM pathology. Microglia express an M2 protumor phenotype in the presence of glioma cells, and this M2 phenotype is downregulated in the presence of an mTOR inhibitor. |
| Leitte et al. [29] | 13 November 2019 | Cellular culture | One human and two human GBM cell lines from European cellular banks | Human MG cell line, CHME3 | Not specified | Results confirm the role of MG in GBM behavior using human cell lines cultured in human serum-based conditions and validate the use of in vitro controllable platforms that recapitulate the microenvironment of GBM as powerful tools for cancer studies. | CHME3 cells were shown to be responsive to classical proinflammatory signals through the Toll-like 4 receptor (TLR4) stimulation by LPS and IFN-γ, leading to activation of STAT1 and the nuclear factor-kb (NF-kb). When challenged with cytotoxic agents, GBM cells showed an increase in proliferation when in contact with MG. Even a low amount of MG (10%–20%) is able to confer resistance of GBM to cytotoxics. |
| Chiavari et al. [35] | 3 November 2020 | Cellular culture and histopathology | 18 | GAMs | Tumor and normal-appearing brain | The data showed the expression of PDIA3 not only in tumor cells but also in GAMs, supporting its potential role in cellular and molecular processes related to GB. | Data showed the expression of PDIA3 not only in tumor cells but also in GAMs, supporting its potential role in cellular and molecular processes related to GB. |
| Fu et al. [36] | 7 May 2020 | Mass cytometry | 16 | GAMs, T cells, NK cells | Tumor and peripheral blood | Recurrent and initial GBMs shared similar immune signatures (the cell types were identified based on the following parameters: T cells, CD45+ CD3+; natural killer (NK) cells, CD45+ CD3-CD16+ CD56+ (10, 19); B cells, CD45+ CD19+; monocytes, CD45+ CD14+ CD16+ (20); macrophages or microglial cells, CD45+ CD11b+ CD3-CD19- CD66b- (15); Tregs, CD45+ CD4+ CD25+ CD127- (21), and granulocytes, CD45+ CD66b+); however, the proportion of GAMs in the recurrent GBMs was decreased compared with that in initial GBMs. | This study confirmed that GAMs, as the dominant infiltrating immune cell population, exhibit substantial inter- and intratumoral heterogeneity in the GBM immune microenvironment, and increased proportions of exhausted T cell subpopulations and Tregs substantially contribute to local immune suppressive characteristics. |
| Chen et al. [26] | 18 November 2020 | Cellular culture | 2 (GBM12 and GBM39) | Not specified | Not specified | In the study, the combination of increased proliferation but decreased invasion aligns with the go-or-grow hypothesis but more importantly demonstrates that crosstalk between MG and GBM cells in the tumor microenvironment may have powerful effects on GBM activities tied directly to tumor progression and patient survival. | GO analyses revealed GBM-MG co-culture upregulated genes in a patient-derived GBM specimen associated with cell cycle, RNA and DNA division, and metabolic activity. However, genes involved in cell adhesion and migration showed significant downregulation as a result of GBM-MG co-culture. |
| Tan et al. [37] | 21 December 2020 | Genomics | 9 | Macrophages | Not specified | A prognostic model made of six genes was constructed to predict the outcomes of LGG and these are correlated with immune checkpoints which provide a valuable role in diagnosis, prognosis, and immunotherapy of glioma. | The phenotype of macrophages was related to whether the cell is neoplastic or not. The M2 macrophages are mainly gathered in neoplastic cells, while the M1 macrophages are located in non-neoplastic cells. CD163, FPR3, LPAR5, P2ry12, PLAUR, SIGLEC1. CD163, FPR3, SIGLEC1 are correlated to the M2 phenotype |
| Wei et al. [38] | 14 April 2021 | Cellular culture and histopathology | 63 | Macrophages | Cell lines from biobank | This study reports a new mechanism of endothelial cell activation in GBM, which is mediated by TNFα secreting GAMs. | This study shows that GBM cells secrete two important cytokines, IL-8 and CCL2, which stimulate GAMs to produce TNFα. Secreted TNFα then activates ECS to express a gene signature indicative of EC activation, including increased expression of VCAM1, ICAM1, CXCL5, and CXCL10. |
| C.-k. Shen et al. [17] | 28 May 2021 | Cellular culture | Not specified | Macrophages | U251 human GBM cells, u87 human GBM cells, thp-1 human monocyte cells | Propose the regulatory mechanism of IL-1β for interaction between GBM and TAM | Il-1β induces the expression of ICAM-1 and VCAM-1 in GBM by binding to the IL-1 receptor, which may play key roles in interacting with GBM and tumor-associated immune cells. |
| Senjor et al. [39] | 29 June 2021 | RT-qpcr | Not specified | Macrophages | Not specified | The data show that cystatin F mRNA and protein levels are increased in GBM tissues compared to those in nonmalignant brain tissues. We found that the levels of the transcription factor C/ebpα also increased with disease progression. Cystatin F is expressed by immune cells, differentiated GBM cells, and undifferentiated GSCS | Elevated mRNA levels of Cystatin F and its transcription factor C/ebpα were observed in advanced gliomas compared to nonmalignant brain tissues and in GSCS compared to differentiated GBM cells. Cystatin F was expressed in cells positive for CD68, Iba-1, GFAP, CD44, and SOX-2. |
| Tanaka et al. [40] | 29 June 2021 | Qrt-PCR and immunohistochemistry | 22 humans | GAMs | 15 tumors, seven epileptic surgeries | Peripheral IMG cells obtained by the authors’ previously developed technique can be used to gauge the properties of PMG from the tumor lesion microenvironment in the CNS. | The specific immune status of glioma might be monitored using peripheral IMG cells. There is specific upregulation of CD206 in IMG cells isolated from the peripheral blood of patients with glioma. Synchronous upregulation of CD206 expression levels was observed in most patients with glioma (6/9, 66.7%) and almost all patients with glioblastoma (4/5, 80%). |
| E. Nuñez et al. [41] | 7 December 2021 | Cellular culture | 20 | Macrophages | Not specified | Microglia-derived EGF, PDGFβ, SDF-1α, and IL-6 were identified as the primary activators driving PYK2 and FAK activation in glioma. | Microglia treated with glioma-conditioned medium (GCM) from primary cell lines increased the expression of genes encoding PDGFβ, SDF-1α, IL-6, IL-8, and EGF. |
| Liu et al. [42] | 14 September 2021 | Cellular culture and srna sequencing | Five individuals diagnosed with glioma + glioma specimens from 165 chemo- and radiotherapy-naive patients undergoing craniotomy from August 2016 to February 2020 (among these 21 fresh tissues were collected for the following cell culture) | Macrophages and monocyte-derived TAMs | Tumor | Targeting the HGG-AM may represent a promising therapeutic approach for agbm. | HGG-AM was demonstrated to be predominantly enriched in IDH-WT GBM and activated by SETD2-mut tumor cells via TGF-β1 secretion. P2RY12 expression was dramatically compromised in HGG-AM, suggesting a specific microglia activation driven by IDH-WT cancer cells. HGG-AM expressed PDGFRα, suggesting their proliferative capacity. |
| Urbantat et al. [43] | 16 October 2021 | Histopathological | 76 samples from 38 patients | TAMs | Paired initial and recurrent GBM | A significantly decreased infiltration of tumor-associated microglia and macrophages (TAM, identified through Iba-1) was observed in recurrent tumors, while a high TAM infiltration in primary tumors was associated with a reduced OS. | The analysis of 76 matched primary and recurrent GBM samples underlined important morphological differences between primary and recurrent GBM, with a higher infiltration of TAMs in the primary tumors. Infiltration of TAMs in primary tumors served as a negative predictor for patients’ OS. |
| Author | Year (dd-mm-yyyy) | Type of Study | Sample Size | Microglia Population | Sample | Outcomes | Observations |
|---|---|---|---|---|---|---|---|
| Parney et al. [44] | Mar 2009 | Cellular culture | Nine humans | Macrophages | Tumor only | It has been reported that primary glioma cultures have immunological characteristics that are lost in successive passages. | Monocyte-lineage cells formed the most prominent group of inflammatory cells infiltrating human gliomas. Unexpectedly, it was found that these cells were predominantly CD45bright/CD11b+. This phenotype corresponds to infiltrating systemic macrophages. |
| X. Yuan et al. [45] | 1 December 2016 | Histopathological | 33 | Macrophages | Tumor | From the patient profiles, specific combinations of expression were found, both throughout the tumor sample, and when specifically comparing the adjacent and bulk tissue regions it was possible to predict poorer or better survival. | Higher coverage of astrocytes in adjacent tissue regions increased the hazard of death, whereas positive staining within the tumor had no effect on survival. Increased coverage of microglia adjacent to the tumor decreased the hazard of death while microglia in the tumor increased the hazard of death. |
| Guo et al. [46] | 2 September 2016 | Cellular culture | 47 | Macrophages and monocyte-derived TAMs | Tumor and five normal brain tissues | Demonstrated that hypoxia enhanced the recruitment of TAMs by upregulating POSTN expression in glioma cells. The hypoxic glioma microenvironment polarized TAMs toward the M2 subtype by increasing the expression of M-CSFR in macrophages and TGF-β in glioma cells. | The enhanced directional migration of macrophages toward hypoxic areas has been attributed to the hypoxia-inducible expression of POSTN in glioma cells which may partially explain the mechanism by which macrophages become trapped in hypoxic regions after they were initially attracted to them. |
| C. Mignogna et al. [47] | 23 February 2016 | Histopathology and immunohistochemistry | 37 | Macrophages | Tumor only | Define the distribution and polarization of TAMs in GBM | Immunohistochemical evaluation of macrophage phenotype showed an interesting scenario; the number of cd163+ macrophages was significantly higher (p < 0.001) than those of cd68+ macrophages in the perinecrotic area, parenchyma, and, in particular, in perivascular areas. In confocal analysis, the cd163+ population consisted of both cd163+/cd68+ macrophages and single stained cd163+ cells. Interestingly, this finding was more evident in perivascular areas. |
| L. Lisi et al. [48] | 2 March 2017 | Hisopathology and immunohistochemistry | 41 | Macrophages | Tumor core and healthy brain | Results suggest that cd163 expression is higher within the tumor than in the surrounding periphery in both male and female patients. | In the present work, we show that cd163 expression is higher within GBM specimens than in the surrounding periphery in both male and female patients. We report that both inos (m1 marker) and arg-1 (m2 markers) are present both within the tumor and in peripheral parenchyma, albeit unevenly distributed. |
| Annovazzi et al. [49] | 12 September 2017 | Histopathological | 108 | Macrophages | Tumor and 10 healthy controls | Characterization of different patterns of microglia composition based on the degree of infiltration. | The transition from HIA (High infiltration area) to ST (solid tumor) was marked by an almost disappearance of RM (reactive microglia) forms, which could be reduced, destroyed, or switched to serve a different function. |
| T. Hide et al. [50] | 1 March 2018 | Genomics | 89 | Macrophages | Tumor core, border, and periphery | During tumor growth, opcs (oligodendrocyte precursor cells) and macrophages and microglia migrate and proliferate rapidly in the border region, where they secrete growth factors and cytokines, causing GBM cells to acquire stem cell profiles and chemo-radio resistance. | The rapid reaction potential of opcs and macrophages and microglia provides advantages for GBM cells in the formation of the GSC niche in the border because GBM cells possess higher proliferation and migration potential, thereby necessitating rapid adaptation of supportive cells. |
| Yu-Ju Wu et al. [51] | 8 October 2019 | Cellular culture | 47 | Macrophages | Tumor only | Demonstration that CCL5-regulated glioma migration and invasion are associated with the expression levels of p-PYK2 and MMP2. | Newly diagnosed cases with a high CCL5 level were associated with increased tumor volume. GAMs of GBM were involved in the production of CCL. It was observed that high-grade glioma contained a higher level of GAM infiltration. Glioma cells elicited a more robust homing response toward GM-CSF–activated GAM-CM. |
| Landry et al. [52] | 11 November 2020 | Genomics | 14 tumors | Macrophages | Tumor core and tumor periphery | Analyze geographical differences in macrophage recruitment and activation through sequential activation states. Find distinct activation and maturation processes between tumor core and periphery. | It was found that TAMs in the tumor core mostly originate from the bone marrow-derived pool whereas those in the tumor periphery are largely derived from microglial cells, supporting prior research. Cells in the tumor core evolve from a “pre-activation” state toward a proinflammatory state. Tumor periphery, by contrast, it was found that cells transition from a preactivation state towards pro-oncogenic activation (M2 or “alternative” activation state). |
| Tang et al. [53] | 8 April 2021 | Genomics | 133 | Macrophages | Databases from CCGA | The study comprehensively analyzed the rGBM microenvironment gene signatures, and integrated rGBM microenvironment-associated genes and up-degs in rGBM to identify a novel prognostic immune-related gene, LRRC15. | Confirmed the negative survival role of LRRC15 as an independent prognostic factor, which indicated that LRRC15 could be used as a novel biomarker for predicting the rGBM patients’ outcomes. |
| K. Kai et al. [54] | 30 September 2021 | Cellular culture | Not specified | TAMs | Human GBM cell lines t98g and u251 obtained from atcc | Tam-derived il-1β increases growth rate of gbm, promotes TAMs infiltration in the TME and shows higher expression patterns in perinecrotic areas | Involvement of il-1β in GBM (cell lines t98g and u251, with emphasis on u251) growth by means of in vitro studies. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Menna, G.; Mattogno, P.P.; Donzelli, C.M.; Lisi, L.; Olivi, A.; Della Pepa, G.M. Glioma-Associated Microglia Characterization in the Glioblastoma Microenvironment through a ‘Seed-and Soil’ Approach: A Systematic Review. Brain Sci. 2022, 12, 718. https://doi.org/10.3390/brainsci12060718
Menna G, Mattogno PP, Donzelli CM, Lisi L, Olivi A, Della Pepa GM. Glioma-Associated Microglia Characterization in the Glioblastoma Microenvironment through a ‘Seed-and Soil’ Approach: A Systematic Review. Brain Sciences. 2022; 12(6):718. https://doi.org/10.3390/brainsci12060718
Chicago/Turabian StyleMenna, Grazia, Pier Paolo Mattogno, Carlo Maria Donzelli, Lucia Lisi, Alessandro Olivi, and Giuseppe Maria Della Pepa. 2022. "Glioma-Associated Microglia Characterization in the Glioblastoma Microenvironment through a ‘Seed-and Soil’ Approach: A Systematic Review" Brain Sciences 12, no. 6: 718. https://doi.org/10.3390/brainsci12060718
APA StyleMenna, G., Mattogno, P. P., Donzelli, C. M., Lisi, L., Olivi, A., & Della Pepa, G. M. (2022). Glioma-Associated Microglia Characterization in the Glioblastoma Microenvironment through a ‘Seed-and Soil’ Approach: A Systematic Review. Brain Sciences, 12(6), 718. https://doi.org/10.3390/brainsci12060718