Amelioration of Scopolamine-Induced Amnesic, Anxiolytic and Antidepressant Effects of Ficus benghalensis in Behavioral Experimental Models


 , and
, and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Preparation of Plant Extract
2.3. Drugs and Chemicals
2.4. Identification of Phytocomponents
2.4.1. Gas Chromatography–Mass Spectrometry (GC–MS) Analysis
2.4.2. Derivatization
2.5. Behavioral Tests for Memory and Learning
2.5.1. Passive Avoidance Task (Step-Through)
2.5.2. Y-maze Task
2.5.3. Morris Water Maze Task
2.6. Behavioral Tests for Anxiety
2.6.1. Open Field Test
2.6.2. Elevated Plus-Maze Test
2.7. Behavioral Tests for Depression
Forced Swimming Test
2.8. Statistical Analysis
3. Results
3.1. Phytochemical Analysis
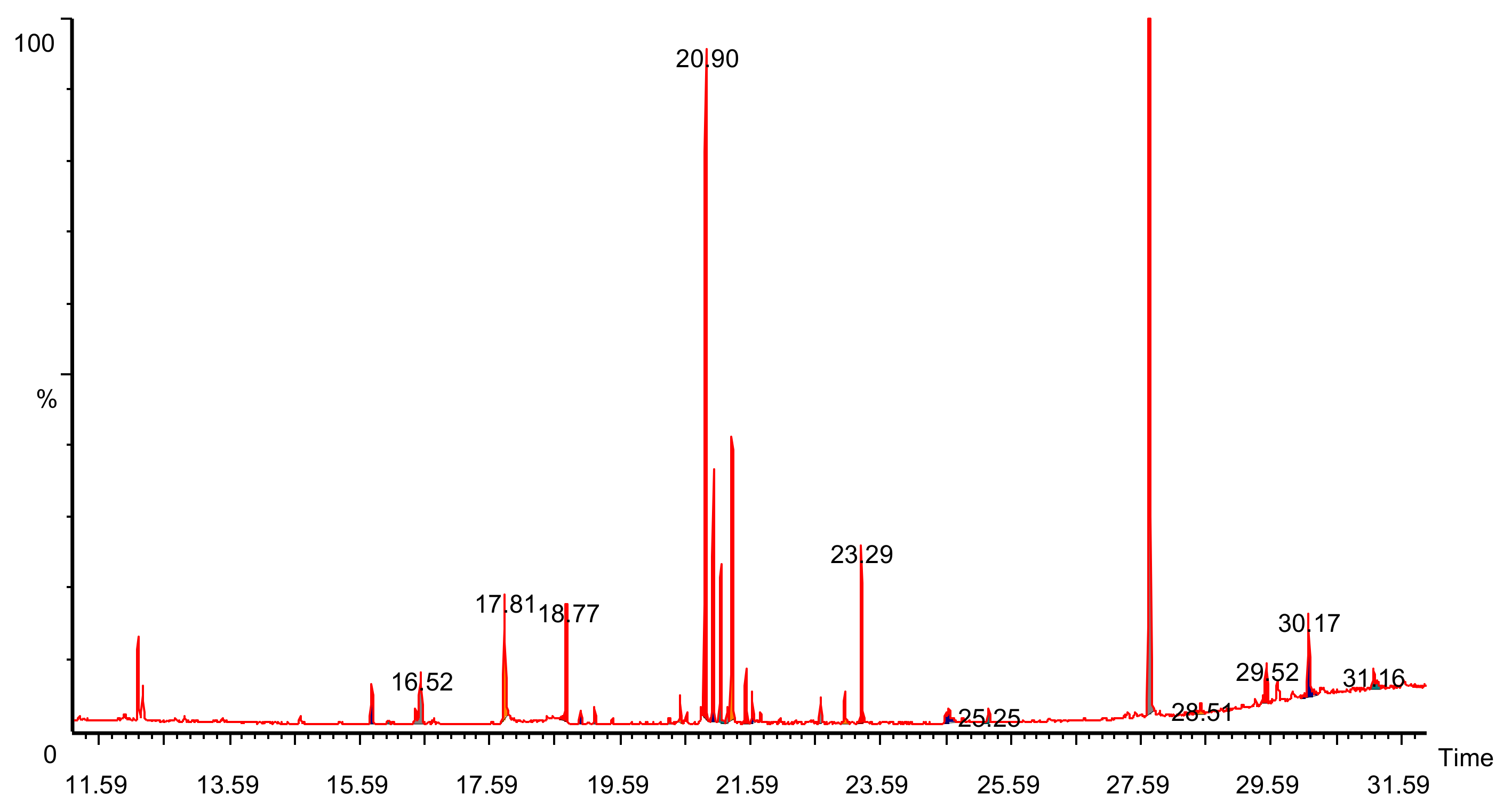
3.2. GC–MS Analysis for Phytocomponents
3.3. Effect on Spatial Memory and Learning
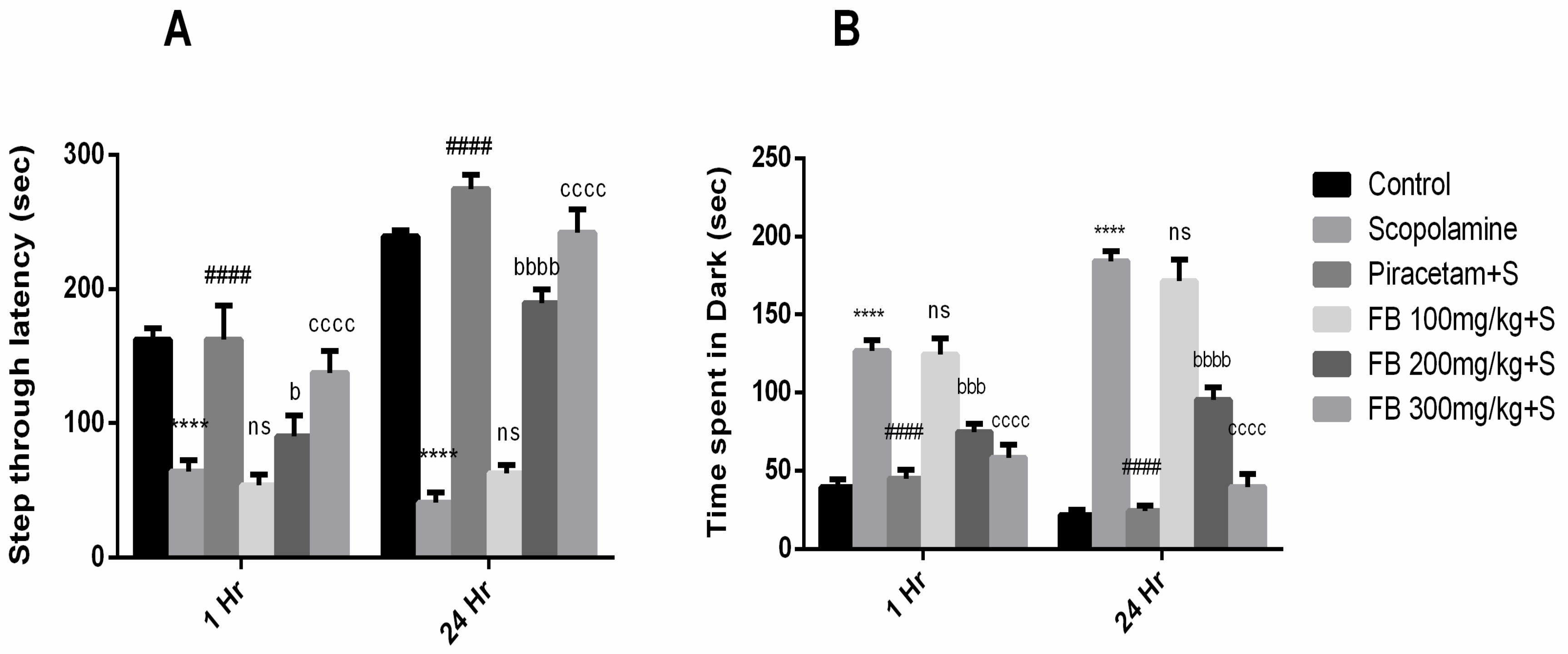
3.3.1. Passive Avoidance Test
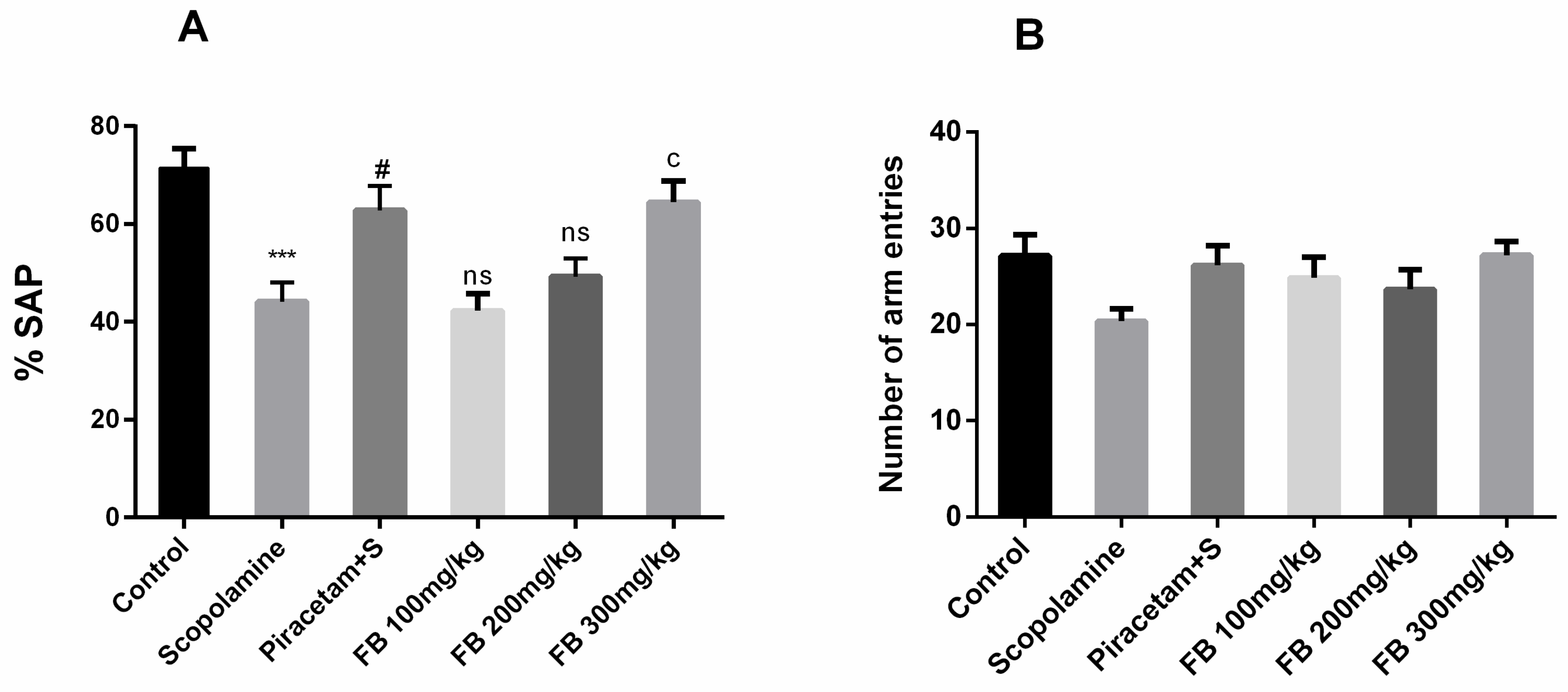
3.3.2. Y-maze Test
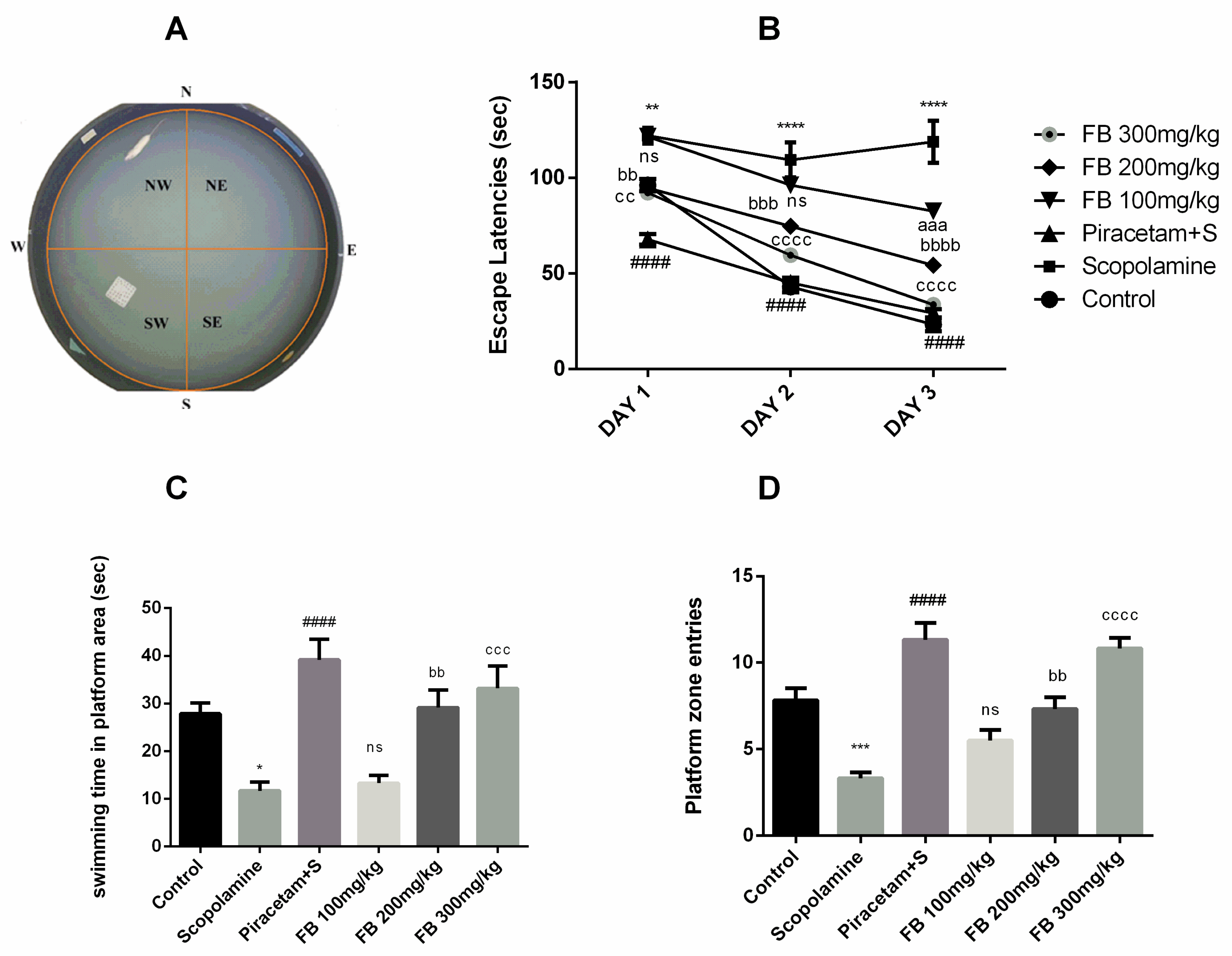
3.3.3. Morris Water Maze Test
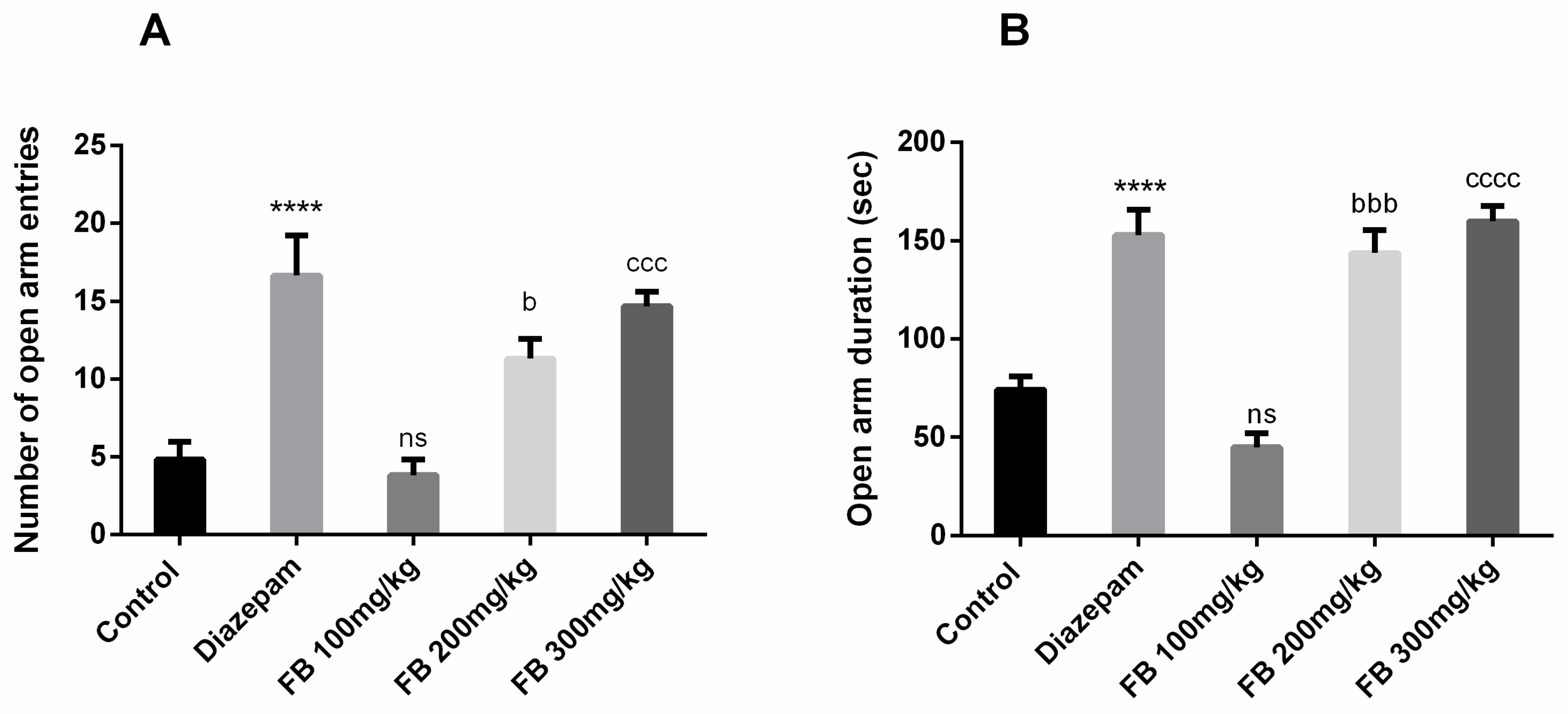
3.4. Anxiolytic Effect
3.4.1. Open Field Test
3.4.2. Elevated Plus Maze Test
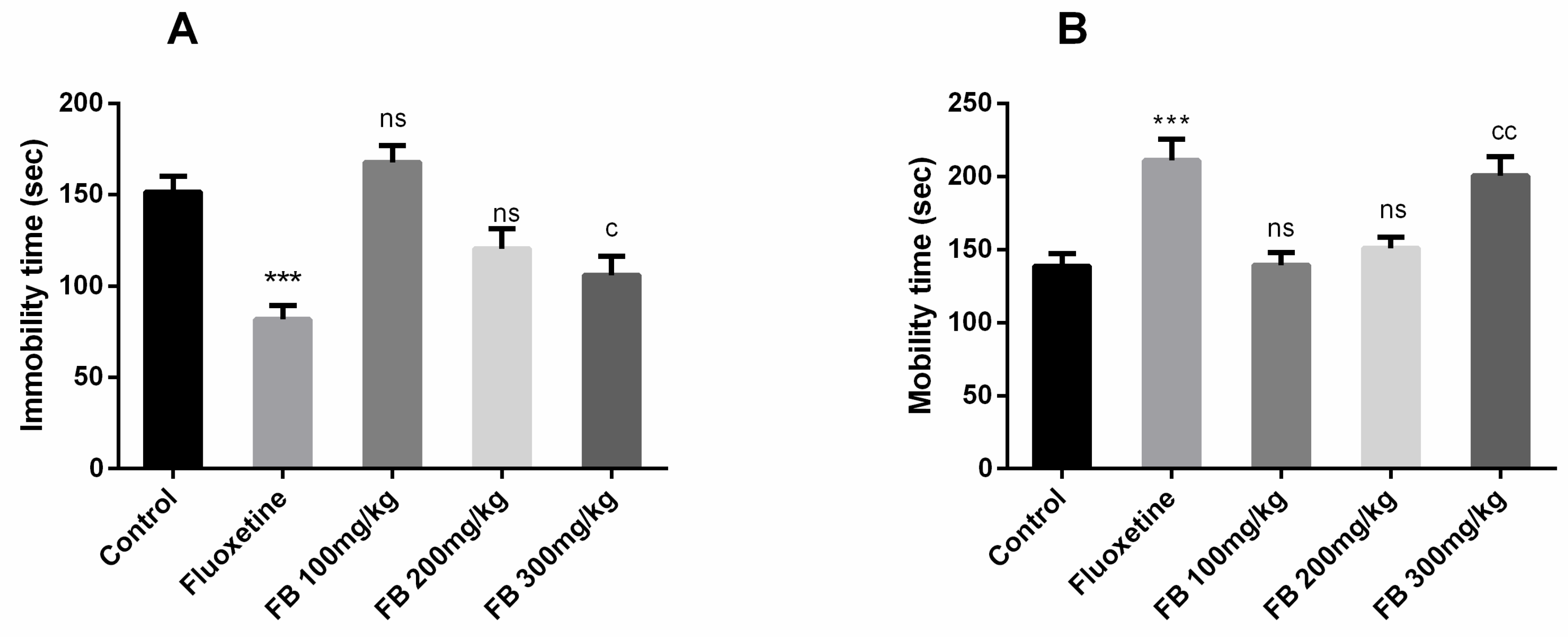
3.5. Antidepressant Effect
Forced swimming test
4. Discussion
5. Conclusion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- WHO. Mental Disorders Affect One in Four People. Available online: https://www.who.int/whr/2001/media_centre/press_release/en/ (accessed on 6 March 2020).
- Ferrara, M.; Langiano, E.; Di Brango, T.; De Vito, E.; Di Cioccio, L.; Bauco, C. Prevalence of stress, anxiety and depression in with Alzheimer caregivers. Health Qual. Life Outcomes 2008, 6, 93. [Google Scholar] [CrossRef]
- Nordberg, A. Pharmacological treatment of cognitive dysfunction in dementia disorders. Acta Neurol. Scand. 1996, 94, 87–92. [Google Scholar] [CrossRef]
- Jucker, M.; Walker, L.C. Pathogenic protein seeding in Alzheimer disease and other neurodegenerative disorders. Ann. Neurol. 2011, 70, 532–540. [Google Scholar] [CrossRef] [PubMed]
- Morrison, A.S.; Lyketsos, C. Review the Pathophysiology of Alzheimer’s Disease and Directions in Treatment. Adv. Stud. Nurs. 2005, 3, 256–270. [Google Scholar]
- Bores, G.M.; Huger, F.P.; Petko, W.; Mutlib, A.E.; Camacho, F.; Rush, D.K.; Selk, D.E.; Wolf, V.; Kosley, R.W.; Davis, L.; et al. Pharmacological evaluation of novel Alzheimer’s disease therapeutics: Acetylcholinesterase inhibitors related to galanthamine. J. Pharmacol. Exp. Ther. 1996, 277, 728–738. [Google Scholar] [PubMed]
- Stein, D.J.; Scott, K.M.; de Jonge, P.; Kessler, R.C. Epidemiology of anxiety disorders: From surveys to nosology and back. Dialogues Clin. Neurosci. 2017, 19, 127–136. [Google Scholar]
- Testa, A.; Giannuzzi, R.; Sollazzo, F.; Petrongolo, L.; Bernardini, L.; Daini, S. Psychiatric emergencies (part I): Psychiatric disorders causing organic symptoms. Eur. Rev. Med. Pharmacol. Sci. 2013, 17 (Suppl. 1), 55–64. [Google Scholar]
- Parsaik, A.K.; Mascarenhas, S.S.; Khosh-Chashm, D.; Hashmi, A.; John, V.; Okusaga, O.; Singh, B. Mortality associated with anxiolytic and hypnotic drugs—A systematic review and meta-analysis. Aust. N. Z. J. Psychiatry 2016, 50, 520–533. [Google Scholar] [CrossRef]
- Kraeuter, A.K.; Guest, P.C.; Sarnyai, Z. The Y-Maze for Assessment of Spatial Working and Reference Memory in Mice. In Methods in Molecular Biology; Humana Press Inc.: Totowa, NJ, USA, 2019; Volume 1916, pp. 105–111. [Google Scholar]
- Maj, M. Development and validation of the current concept of major depression. Psychopathology 2012, 45, 135–146. [Google Scholar] [CrossRef]
- Hirschfeld, R.M.A. The epidemiology of depression and the evolution of treatment. J. Clin. Psychiatry 2012, 73, 5–9. [Google Scholar] [CrossRef]
- Belzung, C.; Willner, P.; Philippot, P. Depression: From psychopathology to pathophysiology. Curr. Opin. Neurobiol. 2015, 30, 24–30. [Google Scholar] [CrossRef]
- Ressler, K.J.; Nemeroff, C.B. Role of serotonergic and noradrenergic systems in the pathophysiology of depression and anxiety disorders. Depress. Anxiety 2000, 12, 2–19. [Google Scholar] [CrossRef]
- Dunlop, B.W.; Nemeroff, C.B. The role of dopamine in the pathophysiology of depression. Arch. Gen. Psychiatry 2007, 64, 327–337. [Google Scholar] [CrossRef]
- Si, X.; Miguel-Hidalgo, J.J.; O’Dwyer, G.; Stockmeier, C.A.; Rajkowska, G. Age-dependent reductions in the level of glial fibrillary acidic protein in the prefrontal cortex in major depression. Neuropsychopharmacology 2004, 29, 2088–2096. [Google Scholar] [CrossRef]
- Gunnell, D.; Ashby, D. Antidepressants and suicide: What is the balance of benefit and harm. Br. Med. J. 2004, 329, 34–38. [Google Scholar] [CrossRef]
- Rush, A.J. STAR*D: What have we learned? Am. J. Psychiatry 2007, 164, 201–204. [Google Scholar] [CrossRef]
- Cassano, P.; Fava, M. Tolerability issues during long-term treatment with antidepressants. Ann. Clin. Psychiatry 2004, 16, 15–25. [Google Scholar] [CrossRef]
- Khawam, E.A.; Laurencic, G.; Malone, D.A. Side effects of antidepressants: An overview. Cleve. Clin. J. Med. 2006, 73, 351–361. [Google Scholar] [CrossRef]
- Ekor, M. The growing use of herbal medicines: Issues relating to adverse reactions and challenges in monitoring safety. Front. Neurol. 2014, 4, 177. [Google Scholar] [CrossRef]
- Gopukumar, S.T.; Praseetha, P.K. Ficus benghalensis Linn—The sacred Indian medicinal tree with potent pharmacological remedies. Int. J. Pharm. Sci. Rev. Res. 2015, 32, 223–227. [Google Scholar]
- Thite, A.T.; Patil, R.R.; Naik, S.R. Anti-arthritic activity profile of methanolic extract of Ficus bengalensis: Comparison with some clinically effective drugs. Biomed. Aging Pathol. 2014, 4, 207–217. [Google Scholar] [CrossRef]
- Panday, D.R.; Rauniar, G.P. Effect of root-extracts of Ficus benghalensis (Banyan) in memory, anxiety, muscle co-ordination and seizure in animal models. BMC Complement. Altern. Med. 2016, 16, 429. [Google Scholar] [CrossRef]
- Kunwar, R.M.; Bussmann, R.W. Ficus (Fig) species in Nepal: A review of diversity and indigenous uses. Lyonia 2006, 11, 85–97. [Google Scholar]
- Basu, N.K.; Singh, G.B. Investigations on Indian medicinal plants; Vitex-negundo Linn. Q. J. Pharm. Pharmacol. 1947, 20, 136. [Google Scholar]
- Naquvi, K.J.; Ali, M.; Ahamad, J. Two new phytosterols from the stem bark of Ficus bengalensis L. J. Saudi Chem. Soc. 2015, 19, 650–654. [Google Scholar] [CrossRef]
- Ahmed, H.; Khan, M.Z.I.; Waseem, D.; Nazli, A.; Baig, M.W. Phytochemical Analysis and Antioxidant Potential of Ficus Benghalensis L. J. Bioresour. Manag. 2017, 4, 3. [Google Scholar]
- Verma, V.K.; Sehgal, N.; Prakash, O. Characterization and screening of bioactive compounds in the extract prepared from aerial roots of Ficus benghalensis. Int. J. Pharm. Sci. Res 2015, 6, 5056–5069. [Google Scholar]
- Ghasemi, S.; Moradzadeh, M.; Hosseini, M.; Beheshti, F.; Sadeghnia, H.R. Beneficial effects of Urtica dioica on scopolamine-induced memory impairment in rats: Protection against acetylcholinesterase activity and neuronal oxidative damage. Drug Chem. Toxicol. 2019, 42, 167–175. [Google Scholar] [CrossRef]
- Miedel, C.J.; Patton, J.M.; Miedel, A.N.; Miedel, E.S.; Levenson, J.M. Assessment of spontaneous alternation, novel object recognition and limb clasping in transgenic mouse models of amyloid-β and tau neuropathology. J. Vis. Exp. 2017, 123, e55523. [Google Scholar] [CrossRef]
- McNamara, R.K.; Skelton, R.W. Diazepam impairs acquisition but not performance in the Morris water maze. Pharmacol. Biochem. Behav. 1991, 38, 651–658. [Google Scholar] [CrossRef]
- Vorhees, C.V.; Williams, M.T. Morris water maze: Procedures for assessing spatial and related forms of learning and memory. Nat. Protoc. 2006, 1, 848–858. [Google Scholar] [CrossRef]
- Angelucci, M.E.M.; Cesário, C.; Hiroi, R.H.; Rosalen, P.L.; Da Cunha, C. Effects of caffeine on learning and memory in rats tested in the Morris water maze. Brazilian J. Med. Biol. Res. 2002, 35, 1201–1208. [Google Scholar] [CrossRef]
- Wang, J.; Yuan, J.; Pang, J.; Ma, J.; Han, B.; Geng, Y.; Shen, L.; Wang, H.; Ma, Q.; Wang, Y.; et al. Effects of Chronic Stress on Cognition in Male SAMP8 Mice. Cell. Physiol. Biochem. 2016, 39, 1078–1086. [Google Scholar] [CrossRef]
- Moniruzzaman, M.; Sharoti Bhattacharjee, P.; Rahman Pretty, M.; Sarwar Hossain, M. Sedative and Anxiolytic-Like Actions of Ethanol Extract of Leaves of Glinus oppositifolius (Linn.) Aug. DC. Evid. Based Complement. Altern. Med. 2016, 2016, 8541017. [Google Scholar] [CrossRef]
- Walf, A.A.; Frye, C.A. The use of the elevated plus maze as an assay of anxiety-related behavior in rodents. Nat. Protoc. 2007, 2, 322–328. [Google Scholar] [CrossRef]
- Sawantdesai, N.; Kale, P.; Savai, J. Evaluation of anxiolytic effects of aripiprazole and hydroxyzine as a combination in mice. J. Basic Clin. Pharm. 2016, 7, 97. [Google Scholar]
- Campus, P.; Colelli, V.; Orsini, C.; Sarra, D.; Cabib, S. Evidence for the involvement of extinction-associated inhibitory learning in the forced swimming test. Behav. Brain Res. 2015, 278, 348–355. [Google Scholar] [CrossRef]
- Bayarsukh, N. Strengthening the conservation and sustainable use of plant genetic resources for food and agriculture in Mongolia. Mong. J. Agric. Sci. 2015, 15, 173–179. [Google Scholar] [CrossRef][Green Version]
- Altemimi, A.; Lakhssassi, N.; Baharlouei, A.; Watson, D.G.; Lightfoot, D.A. Phytochemicals: Extraction, isolation, and identification of bioactive compounds from plant extracts. Plants 2017, 6, 42. [Google Scholar] [CrossRef]
- Donovan, N.J.; Amariglio, R.E.; Zoller, A.S.; Rudel, R.K.; Gomez-Isla, T.; Blacker, D.; Hyman, B.T.; Locascio, J.J.; Johnson, K.A.; Sperling, R.A.; et al. Subjective cognitive concerns and neuropsychiatric predictors of progression to the early clinical stages of Alzheimer disease. Am. J. Geriatr. Psychiatry 2014, 22, 1642–1651. [Google Scholar] [CrossRef]
- Deiana, S.; Platt, B.; Riedel, G. The cholinergic system and spatial learning. Behav. Brain Res. 2011, 221, 389–411. [Google Scholar] [CrossRef] [PubMed]
- Krypotos, A.M.; Effting, M.; Kindt, M.; Beckers, T. Avoidance learning: A review of theoretical models and recent developments. Front. Behav. Neurosci. 2015, 9, 189. [Google Scholar] [CrossRef] [PubMed]
- Hejazian, S.; Karimi, S.; Hosseini, M.; Mousavi, S.; Soukhtanloo, M. Protection against brain tissues oxidative damage as a possible mechanism for improving effects of low doses of estradiol on scopolamine-induced learning and memory impairments in ovariectomized rats. Adv. Biomed. Res. 2016, 5, 123. [Google Scholar] [PubMed]
- Eilam, D.; Golani, I. Home base behavior of rats (Rattus norvegicus) exploring a novel environment. Behav. Brain Res. 1989, 34, 199–211. [Google Scholar] [CrossRef]
- Hughes, R.N. The value of spontaneous alternation behavior (SAB) as a test of retention in pharmacological investigations of memory. Neurosci. Biobehav. Rev. 2004, 28, 497–505. [Google Scholar] [CrossRef]
- Blokland, A.; Geraerts, E.; Been, M. A detailed analysis of rats’ spatial memory in a probe trial of a Morris task. Behav. Brain Res. 2004, 154, 71–75. [Google Scholar] [CrossRef]
- Spencer, J.P.E. Flavonoids and brain health: Multiple effects underpinned by common mechanisms. Genes Nutr. 2009, 4, 243–250. [Google Scholar] [CrossRef]
- Khan, H.; Amin, S.; Kamal, M.A.; Patel, S. Flavonoids as acetylcholinesterase inhibitors: Current therapeutic standing and future prospects. Biomed. Pharmacother. 2018, 101, 860–870. [Google Scholar] [CrossRef]
- Maurer, S.V.; Williams, C.L. The cholinergic system modulates memory and hippocampal plasticity via its interactions with non-neuronal cells. Front. Immunol. 2017, 8, 1489. [Google Scholar] [CrossRef]
- Liu, H.; Leak, R.K.; Hu, X. Neurotransmitter receptors on microglia. Stroke Vasc. Neurol. 2016, 1, 52–58. [Google Scholar] [CrossRef]
- Thameem Dheen, S.; Kaur, C.; Ling, E.-A. Microglial Activation and its Implications in the Brain Diseases. Curr. Med. Chem. 2007, 14, 1189–1197. [Google Scholar] [CrossRef]
- Spencer, J.P.E. The impact of fruit flavonoids on memory and cognition. Br. J. Nutr. 2010, 104 (Suppl. 3), S40–S47. [Google Scholar] [CrossRef]
- Hernandez, P.J.; Abel, T. A molecular basis for interactions between sleep and memory. Sleep Med. Clin. 2011, 6, 71–84. [Google Scholar] [CrossRef][Green Version]
- Burg, V.K.; Grimm, H.S.; Rothhaar, T.L.; Grösgen, S.; Hundsdörfer, B.; Haupenthal, V.J.; Zimmer, V.C.; Mett, J.; Weingärtner, O.; Laufs, U.; et al. Plant sterols the better cholesterol in Alzheimer’s disease? A mechanistical study. J. Neurosci. 2013, 33, 16072–16087. [Google Scholar] [CrossRef]
- (PDF) Therapeutic Efficacy of Phytochemicals as Anti-Anxiety-A Review. Available online: https://www.researchgate.net/publication/311913094_Therapeutic_efficacy_of_Phytochemicals_as_Anti-Anxiety-A_Review (accessed on 6 March 2020).
- Hanrahan, J.R.; Chebib, M.; Johnston, G.A.R. Flavonoid modulation of GABA A receptors. Br. J. Pharmacol. 2011, 163, 234–245. [Google Scholar] [CrossRef]
- Cryan, J.F.; Markou, A.; Lucki, I. Assessing antidepressant activity in rodents: Recent developments and future needs. Trends Pharmacol. Sci. 2002, 23, 238–245. [Google Scholar] [CrossRef]
- Lochmann, D.; Richardson, T. Selective Serotonin Reuptake Inhibitors. Handb. Exp. Pharmacol. 2019, 250, 135–144. [Google Scholar]
- Castrén, E.; Rantamäki, T. The role of BDNF and its receptors in depression and antidepressant drug action: Reactivation of developmental plasticity. Dev. Neurobiol. 2010, 70, 289–297. [Google Scholar] [CrossRef]
- Matias, I.; Buosi, A.S.; Gomes, F.C.A. Functions of flavonoids in the central nervous system: Astrocytes as targets for natural compounds. Neurochem. Int. 2016, 95, 85–91. [Google Scholar] [CrossRef]
- Salim, S. Oxidative Stress and Psychological Disorders. Curr. Neuropharmacol. 2014, 12, 140–147. [Google Scholar] [CrossRef]
- Harbottle, L.; Schonfelder, N. Nutrition and depression: A review of the evidence. J. Ment. Heal. 2008, 17, 576–587. [Google Scholar] [CrossRef]
- Spencer, J.P.; Schroeter, H.; Kuhnle, G.; Srai, S.K.; Tyrrell, R.M.; Hahn, U.; Rice-Evans, C. Epicatechin and its in vivo metabolite, 3’-O-methyl epicatechin, protect human fibroblasts from oxidative-stress-induced cell death involving caspase-3 activation. Biochem. J. 2001, 354, 493–500. [Google Scholar] [CrossRef]
- Schroeter, H.; Williams, R.J.; Matin, R.; Iversen, L.; Rice-Evans, C.A. Phenolic antioxidants attenuate neuronal cell death following uptake of oxidized low-density lipoprotein. Free Radic. Biol. Med. 2000, 29, 1222–1233. [Google Scholar] [CrossRef]
- Gould, E.; Tanapat, P. Stress and hippocampal neurogenesis. Biol. Psychiatry 1999, 46, 1472–1479. [Google Scholar] [CrossRef]
- The Role of Dietary Polyphenols on Adult Hippocampal Neurogenesis: Molecular Mechanisms and Behavioural Effects on Depression and Anxiety. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3395274/ (accessed on 6 March 2020).
- Oxidative Stress-Related Biomarkers in Essential Hypertension and Ischemia-Reperfusion Myocardial Damage. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3856219/ (accessed on 6 March 2020).
- Taslimi, P.; Akıncıoglu, H.; Gülçin, İ. Synephrine and phenylephrine act as α-amylase, α-glycosidase, acetylcholinesterase, butyrylcholinesterase, and carbonic anhydrase enzymes inhibitors. J. Biochem. Mol. Toxicol. 2017, 31, e21973. [Google Scholar] [CrossRef]
- Rahman, A.U.; Choudhary, M.I. Bioactive natural products as a potential source of new pharmacophores. A theory of memory. Pure Appl. Chem. 2001, 73, 555–560. [Google Scholar] [CrossRef]
- Darvesh, S.; Cash, M.K.; Reid, G.A.; Martin, E.; Mitnitski, A.; Geula, C. Butyrylcholinesterase is associated with β-amyloid plaques in the transgenic APPSWE/PSEN1dE9 mouse model of alzheimer disease. J. Neuropathol. Exp. Neurol. 2012, 71, 2–14. [Google Scholar] [CrossRef]
- Greig, N.H.; Utsuki, T.; Ingram, D.K.; Wang, Y.; Pepeu, G.; Scali, C.; Yu, Q.S.; Mamczarz, J.; Holloway, H.W.; Giordano, T.; et al. Selective butyrylcholinesterase inhibition elevates brain acetylcholine, augments learning and lowers Alzheimer β-amyloid peptide in rodent. Proc. Natl. Acad. Sci. USA 2005, 102, 17213–17218. [Google Scholar] [CrossRef]
- Song, D.K.; Suh, H.W.; Jung, J.S.; Wie, M.B.; Son, K.H.; Kim, Y.H. Antidepressant-like effects of p-synephrine in mouse models of immobility tests. Neurosci. Lett. 1996, 214, 107–110. [Google Scholar] [CrossRef]
- Lin, Y.; De Vaca, S.C.; Carr, K.D.; Stone, E.A. Role of α1-adrenoceptors of the locus coeruleus in self-stimulation of the medial forebrain bundle. Neuropsychopharmacology 2007, 32, 835–841. [Google Scholar] [CrossRef]
- Kim, K.W.; Kim, H.D.; Jung, J.S.; Woo, R.S.; Kim, H.S.; Suh, H.W.; Kim, Y.H.; Song, D.K. Characterization of antidepressant-like effects of p-synephrine stereoisomers. Naunyn. Schmiedebergs. Arch. Pharmacol. 2001, 364, 21–26. [Google Scholar] [CrossRef] [PubMed]
- Alpha-1A Adrenergic Receptor Modulates Anxiety and Depression in Mice|The FASEB Journal. Available online: https://www.fasebj.org/doi/abs/10.1096/fasebj.23.1_supplement.962.3 (accessed on 6 March 2020).








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| S.No | Phytocomponents | RT | Area% | Area |
|---|---|---|---|---|
| 1 | Butedioic acid | 15.77 | 2.230 | 507,182 |
| 2 | Ribitol | 16.3 | 0.350 | 79,119 |
| 3 | Butanoic acid | 16.52 | 1.710 | 387,201 |
| 4 | L-asparagine | 18.77 | 3.150 | 714,501 |
| 5 | Synephrine | 18.97 | 0.590 | 133,192 |
| 6 | Adonitol | 19.19 | 0.610 | 138,566 |
| 7 | Azelaic acid | 20.34 | 0.220 | 50,773 |
| 8 | Citric acid | 20.51 | 0.960 | 218,233 |
| 9 | Cyanuric acid | 20.90 | 24.53 | 5,570,956 |
| 10 | D-Fructose | 21.02 | 7.33 | 1,665,137 |
| 11 | Galactose | 21.24 | 0.520 | 117,565 |
| 12 | Glucitol | 21.31 | 9.21 | 2,090,264 |
| 13 | Glucose | 21.52 | 1.65 | 37,4594 |
| 14 | Adonitol | 21.75 | 0.49 | 11,0464 |
| 15 | Inositol | 22.67 | 0.850 | 19,2694 |
| 16 | Hexadecanoic acid | 23.03 | 0.610 | 13,7463 |
| 17 | Myo-inositol | 23.29 | 5.13 | 117,5429 |
| 18 | Trans-9-octadecanoic acid | 24.64 | 0.38 | 86877 |
| 19 | 2-O-Glycerol alpha -D-Glucopyranoside | 25.25 | 0.510 | 116,099 |
| 20 | Alpha-D glucopyranoside | 27.72 | 31.75 | 7,210,211 |
| 21 | Turanose | 28.51 | 0.25 | 56,761 |
| 22 | Benzoic acid | 29.52 | 2.59 | 588,105 |
| 23 | Melibiose | 30.17 | 3.46 | 784,751 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Malik, H.; Javaid, S.; Fawad Rasool, M.; Samad, N.; Rizwan Ahamad, S.; Alqahtani, F.; Imran, I. Amelioration of Scopolamine-Induced Amnesic, Anxiolytic and Antidepressant Effects of Ficus benghalensis in Behavioral Experimental Models. Medicina 2020, 56, 144. https://doi.org/10.3390/medicina56030144
Malik H, Javaid S, Fawad Rasool M, Samad N, Rizwan Ahamad S, Alqahtani F, Imran I. Amelioration of Scopolamine-Induced Amnesic, Anxiolytic and Antidepressant Effects of Ficus benghalensis in Behavioral Experimental Models. Medicina. 2020; 56(3):144. https://doi.org/10.3390/medicina56030144
Chicago/Turabian StyleMalik, Humna, Sana Javaid, Muhammad Fawad Rasool, Noreen Samad, Syed Rizwan Ahamad, Faleh Alqahtani, and Imran Imran. 2020. "Amelioration of Scopolamine-Induced Amnesic, Anxiolytic and Antidepressant Effects of Ficus benghalensis in Behavioral Experimental Models" Medicina 56, no. 3: 144. https://doi.org/10.3390/medicina56030144
APA StyleMalik, H., Javaid, S., Fawad Rasool, M., Samad, N., Rizwan Ahamad, S., Alqahtani, F., & Imran, I. (2020). Amelioration of Scopolamine-Induced Amnesic, Anxiolytic and Antidepressant Effects of Ficus benghalensis in Behavioral Experimental Models. Medicina, 56(3), 144. https://doi.org/10.3390/medicina56030144