Abstract
During the Late Miocene, Bramatherium was the main representative of the giraffid subfamily Sivatheriinae in the Indian Subcontinent, with sparse and uncertain records from adjacent regions. In the present paper, we describe and compare two ossicones of the same individual, unearthed from the Upper Miocene site of Fourka in Chalkidiki Peninsula, Northern Greece, and attribute them with certain confidence to the species Bramatherium perimense. The definite record of Bramatherium so far away from the Indian Subcontinent and in close proximity to the well-known Pikermian sivatheriine Helladotherium duvernoyi enabled us to re-discuss both the intrageneric diversity of Bramatherium and the debatable Bramatherium–Helladotherium taxonomy. Our review allows us (i) to recognize only two Bramatherium species in Asia: the large-sized B. grande and the smaller B. perimense and (ii) to confirm and further support with morphological and biogeographic evidence the synonymy of Helladotherium with Bramatherium.
1. Introduction
Giraffidae Gray, 1821 (Mammalia, Ruminantia), is a family of pecoran ruminants, represented by only two genera in modern African fauna. However, they were much more diverse and widespread across the Old World from the Late Miocene to the Early to Middle Pleistocene. One of the main extinct subfamilies of Giraffidae is the Sivatheriinae Zittel, 1893. They constitute a monophyletic clade, characterized by secondary shortening of the metapodials and the neck, p3, with a continuous lingual wall in the talonid, the existence of two pairs of ossicones, and gigantism [1,2,3,4,5,6,7,8]. The subfamily likely arose during the late Middle Miocene, with the species Libytherium proton Ríos, Abbas, Khan & Solounias, 2022, and Vishnutherium priscillum Matthew, 1929, both from the Chinji Formation of the Siwaliks, being its possible earliest representatives [9,10]. Sivatheriinae thrived during the Late Miocene in Eurasia. Their most occidental members were found in the Iberian Peninsula: the primitive Decennatherium Crusafont, 1952, and the more derived Birgerbohlinia Crusafont, 1952 (e.g., [3,6,7,11,12]). Palaeogiraffa Bonis & Bouvrain, 2003, and Helladotherium Gaudry, 1860, were, during the same period, the main representatives of the subfamily in the so-called Greco–Irano–Afghan Province [13]. The mostly primitive Palaeogiraffa constitutes three species from the Vallesian of Northern Greece, Eastern Thrace, and Anatolia, Türkiye [8,14,15], whereas the single known species Helladotherium duvernoyi (Gaudry & Lartet, 1856) is widely recognized among several later Vallesian to Turolian mammal assemblages in today’s Greece, Bulgaria, North Macedonia, Türkiye, and Iran [7,16,17,18,19,20,21,22,23,24].
Further east, Bramatherium Falconer, 1845 (known by the species names Bramatherium perimense Falconer, 1845; Bramatherium megacephalum (Lydekker, 1876); and Bramatherium grande (Pilgrim, 1911)), predominated in the Late Miocene of today’s India and Pakistan and was mostly found in the Middle Siwaliks [25,26,27,28,29,30]. That taxon reached as far east as Thailand and Myanmar [31] and as far north as northern China [32], whereas its record in the United Arab Emirates [33] remains dubious.
This clear-cut biogeographic picture is, however, significantly disturbed and perplexed by several additional findings. The recently described Decennatherium asiaticum Ríos, Danowitz & Solounias, 2019, from the Late Miocene of the Siwaliks [34], the postcranial remains of Helladotherium from the peri-Black Sea areas and France [35,36,37,38], the recognition of B. perimense among the Turolian fauna of Kavakdere, Türkiye [7,39], and the alleged occurrence of Bramatherium sp. in Samos [40] suggest wider though strongly discontinuous and highly overlapping distributions. Complicating the situation even further, the two most widespread and large-sized sivatheres of the Late Miocene of Eurasia, Helladotherium and Bramatherium, appear very similar in terms of morphology and biometry, raising doubts about their taxonomic distinction and allowing several scholars to propose the former as representing females of the latter [17,26,27,39,41].
From the Early Pliocene onwards, sivatheres became extinct in Europe but persisted in Africa and Asia up to the Early/Middle Pleistocene with genera such as Sivatherium Falconer & Cautley, 1836, and Libytherium Pomel, 1892 [42,43,44,45,46]. The Early Pliocene Sivatherium cf. S. hendeyi Harris, 1976, from Spain and the rather Pliocene Sivatherium garevskii Geraads, 2009, from North Macedonia and likely Turkish Thrace [17,47] may mark the last occurrence of the subfamily on the subcontinent. Although the Spanish Pliocene record suggests some Mio-Pliocene Afro-Iberian dispersal [48], the southern Balkan record is supposed to be more closely involved with the Helladotherium-Bramatherium controversy [17].
In this study, we describe two sivatheriine ossicones, belonging to the same individual, from the Late Miocene of Chalkidiki Peninsula, Greece. They come from the Fourka sandpits (FRK) and were originally referred by Lazaridis [22] to Sivatheriinae indet. due to morphological similarities with Bramatherium. This attribution, made in the context of the ongoing Helladotherium–Bramatherium debate, is re-examined here in light of new evidence. They represent the fourth evidence of sivatheriine ossicones in the Balkans and the second in the Late Miocene of the Greco-Irano-Afghan Province along with the ossicone-bearing cranium of B. perimense from Kavakdere, Türkiye [39]. The systematic study of the FRK ossicones provides valuable new insights regarding the taxonomy, biogeography, and phylogenetic relationships of Late Miocene west Eurasian sivatheriines.
2. Materials and Methods
2.1. Geological and Chronological Setting
The Fourka sandpits (FRK) are distributed around the homonymous village of Fourka, in the Kassandra Peninsula of Chalkidiki (Figure 1). In this area, vertebrate fossils are usually found isolated or in small spots of a few specimens in several natural or technical outcrops. The studied ossicones (FRK13a and b) were found by the local Xanthos Makris in a natural outcrop.
Figure 1.
The locality of Fourka (FRK), Chalkidiki Peninsula, Greece, indicated with the red star.
The gray sands and gravels of Fourka sandpits belong to the upper part of the Antonios Formation (Fm) consisting of gray-white conglomerates, gravels, and sands. Basal sediments of the Antonios Fm are dated biochronologically to the Lower/Middle Miocene boundary [49,50,51], whereas top sands, largely exposed on Kassandra Peninsula, are considered of Upper Miocene age. Antonios Fm is overlain by the red beds of Triglia Fm, within which several Turolian mammal assemblages have been recovered [22,52,53,54,55]. The presence of the Vallesian suid Propotamochoerus cf. palaeochoerus [22] in FRK, along with remains of Deinotherium giganteum Kaup, 1829, found in sands similar to those of FRK at Aghia Paraskevi to the southeast of Kassandra Peninsula [56] led Lazaridis et al. [57] to propose a Vallesian age for the FRK fauna. Other taxa known from FRK are Tetralophodon longirostris (Kaup, 1832), ?Hippotherium sp., Urmiatherium kassandriensis Lazaridis, Kostopoulos, Lyras & Roussiakis, 2017, Chalicotheriidae indet., and Rhinocerotidae indet. [22,56].
2.2. Studied Material and Phylogenetic Analysis and Comparison
The studied ossicones (LGPUT FRK-13a and LGPUT FRK-13b) are housed at the Museum of Geology-Palaeontology-Palaeoanthropology, School of Geology, Aristotle University of Thessaloniki (LGPUT). A giraffid M3 (AMPG(V)-110), a proximal radius (AMPG(V)-107), and two distal metapodials (AMPG(V)-105, AMPG(V)-106) were also found in FRK and housed in the Museum of Paleontology and Geology of the National Kapodistrian University of Athens (AMPG). Although Lazaridis [22] ascribed them all to Sivatheriinae indet., the DAP and DT of AMPG(V)-105 are 42.2 and 73.5, respectively, whereas the DAP and DT of AMPG(V)-106 are 48.9 and 71.0. Those dimensions fit better biometrically with a smaller giraffid, of the size of Samotherium boissieri Forsyth Major, 1888, which shows a size range of 38.7–49.0 for the DAP of the metapodials and a size range of 69.0–90.2 for the DT of the metapodials [20]. Therefore, we excluded the metapodials from our current sample. Their presence suggests, however, that at least two giraffids occurred in the FRK fauna.
Part of the comparative material is also kept in LGPUT, whereas [6,17,20,58] were used for further ossiconal comparisons. The ossicones of Decennatherium rex Ríos, Sánchez & Morales, 2017, were studied in person (KL) at the Regional Archaeological Museum of Madrid (MARPA), Alcalá de Henares, Spain. Comparative measurements of Helladotherium and Bramatherium postcranials were taken from several works, and they are given along with their references in Supp. File 1.
The phylogenetic analysis was conducted based on the character matrix by Ríos et al. [6], in which we included the FRK-13 specimens (Supp. File 2); the analysis contains 32 taxa and 111 characters in total. All the characters were treated as unweighted and non-additive. We performed two modifications in the original dataset changing the state of character 13 (Anterior ossciones section) from 2 (Expanded-Amorphous) to 1 (Circular) for S. hendeyi and S. giganteum. We used the Maximum Parsimony criterium to plot FRK-13 to the giraffid phylogenetic tree. The character matrix was built on Mesquite 3.61 software [59], and the cladistic analysis was performed using TNT 1.5 [60]. The Traditional Search method with Tree Bisection Reconnection (TBR) and 1000 replicates (holding the ten most parsimonious trees for each replicate) was used. We used Bremer statistics (BREMER.RUN) to assess branch support.
2.3. Museum Abbreviations
AM: American Museum of Natural History, New York;
FM: Fossil Mammal Collection, Natural History Museum of Sofia;
NHMA: Natural History Museum of Aegean, Samos;
NHMB: Natural History Museum of Basel;
NHML: Natural History Museum of London;
PIM: Geologische—Palaeontologische Sammlungen, Universitaet Muenster;
YPM: Yale Peabody Museum, New Haven, CT.
3. Results
3.1. Systematic Paleontology
Class: Mammalia Linnaeus, 1758.
Order: Artiodactyla Owen, 1848.
Suborder: Ruminantia Scopoli, 1777.
Family: Giraffidae Gray, 1821.
Subfamily: Sivatheriinae Zittel, 1893.
Genus: Bramatherium Falconer, 1845.
Synonyms: Helladotherium Gaudry, 1860;
Hydaspidotherium Lydekker, 1876;
Hydaspitherium Lydekker, 1878.
Species: Bramatherium perimense Falconer, 1845.
Synonyms: Helladotherium duvernoyi (Gaudry & Lartet, 1856);
Hydaspidotherium megacephalum Lydekker, 1876;
Bramatherium megacephalum (Lydekker, 1876);
Hydaspitherium megacephalum Lydekker, 1878.
Emended Diagnosis: Large-sized giraffid. Males with two pairs of ossicones. The anterior ossicones are straight and porous and rise vertically from a common frontoparietal base. The posterior ossicones are curved, hollow, and laterally oriented. The surfaces of the ossicones of both pairs demonstrate deep ridges, grooves, and bumps. Blunt apices in both ossicones. Hornless females. Fully molarized p4. Sivatheriine-like p3. Metapodials moderately elongated and robust. Metatarsal with strongly separated articular surfaces for the naviculocuboideum and the os cuneiforme intermediolaterale.
Description
The two ossicones (FRK-13a and FRK-13b, Figure 2) were found in close proximity and show similar taphonomic and fossilization characteristics in a way that suggests that they belong to the same individual [22]. However, they demonstrate clear morphological differences which indicate that they belong to different pairs of ossicones, with one being supraorbital and the other located more distally towards the parietal or occipital [22]. Both ossicones come from the right side of the cranium.

Figure 2.
Bramatherium perimense Falconer, 1845, ossicones from Fourka, Chalkidiki Peninsula, Greece. (A–D) Anterior ossicone FRK-13a in cranial (A), lateral (B), caudal (C), and medial (D) views. (E–H) Posterior ossicone FRK-13b in caudal (E), ventral (F), cranial (G), and dorsal (H) views. Scale bars equal 20 cm.
The anterior ossicone (FRK-13a, Figure 2A–D) is attached to its base by a ridge medially. That ridge is probably a part of a frontoparietal base from which the two anterior ossicones are arising. The preserved shape of the base and the lack of the paired ossicone do not allow us to fully understand their positioning in respect to the cranium, i.e., completely vertically or inclined laterally. In the cross section, the anterior ossicone is porous, like in the extant Giraffa Brisson, 1762. Its cross section is semi-triangular towards its base; its lateral and anterior borders are straight and perpendicular to each other, whereas the postero-medial border is slightly curved. Moving towards the apex, the lateral and posterior surfaces are straight and perpendicular to each other, whereas the antero-medial border becomes curved. The surface of the ossicone is heavily textured with multiple elongated grooves and ridges. Some of the grooves are very deep and wide, whereas others are narrower and shallower. A very pronounced ridge lies exactly on the border between the anterior and the lateral surface. There are two other pronounced ridges: one in the middle of the lateral surface and the other one on the posterior surface. Numerous bumps occur on the surface of the ridge, indicating secondary bone growth. The size of the bumps varies, and several of them seem to be extended in neighboring ridges (interrupted by the intermediate grooves). Both the ridges and the grooves are clearly extended to the lateral part of the frontoparietal base. They seem to faint more medially as the grooves, ridges, and bumps are much less developed in this part. The apex of the ossicone is partially damaged but the ossicone probably ends in a blunt apex, which is no more expanded than the ossicone shaft. The surface of the apex is finely textured, with small bumps and pits, indicating that secondary bone growth was also present in this region.
The posterior ossicone (FRK-13b; Figure 2E–H) is broken towards its base; however, it seems that it was pneumatized at that area, with several sinuses being observed. It is rather unlikely that it was arising from a common base with the other posterior ossicone, as is the case in the anterior part. Moreover, it is more curved than the anterior ossicone, its interior is hollowed, and its cross section is circular to oval. It seems to be shorter and slenderer than the anterior ossicone. Unfortunately, its orientation cannot be safely traced. Its surface is also heavily textured (less than the anterior though), with numerous grooves, ridges, bumps, and pits of varying degrees of development. Its apex seems similar to that of the anterior ossicone, being finely textured and not more expanded than the shaft of the ossicone.
Comparisons
The giraffid individual FRK-13 probably bears two pairs of ossicones, whereas the palaeotragines (Palaeotragus Gaudry, 1861; Samotherium Forsyth-Major, 1888; and Schansitherium Bohlin, 1926), the giraffines (Giraffa; Bohlinia Matthew, 1929; Honanotherium Bohlin, 1926) and Okapia Lankester, 1901, bear only a single pair of ossicones (apart from Giraffa, which have a third nasal ossicone). Morphologically, FRK-13a and b are totally different from the palaeotragine ossicones, which have smooth surfaces and pointy apices [20,61,62]. They are also different from the ossicones of the Giraffinae (Bohlinia and Honanotherium), which end in a blunt tip, more expanded than the ossicone’s shaft [63,64]. The extant Giraffa’s main ossicones are also much shorter, their surfaces do not have deep grooves and ridges, and they bear very enlarged apical bumps [65,66]. Okapia’s ossicones are also different in that the shaft is smooth and only the apex is textured [61,63].
The presence of two pairs of ossicones has been widely considered as a sivatheriine apomorphy [3,6,24,32]. D. rex from the Vallesian of Los Batallones, Spain is considered a stem-sivathere by Ríos et al. [6], united to the more advanced sivatheres on the basis of a second pair of ossicones. Its ossicones are profoundly different from those of FRK-13: the anterior ossicones of D. rex are very small and not arising from a common base, in contrast to the massive FRK-13a. The posterior ossicones of D. rex, as well as of D. asiaticum, are closer in length to FRK-13b; however, they are slenderer and more curved, and their bumps are much larger. Birgerbohlinia schaubi Crusafont, 1952, is a sivathere, native to the Iberian Peninsula. It resembles D. rex in that the anterior ossicones are significantly smaller than the posterior ones [3], although the distinction is much more extreme in D. rex. In that respect, Bi. schaubi differs from FRK-13 in which both ossicones appear very large, having only minor differences in overall size. The posterior ossicones CR-513 and CR-514 of Bi. schaubi [3] bear surficial, irregular, longitudinal grooves like FRK-13. However, they are slenderer and more curved. FRK-13 ossicones are also completely different from those of Sivatherium, in which the anterior pair of ossicones are very small compared to the posterior ones. Moreover, in S. giganteum Falconer & Cautley, 1836, and S. garevskii, the posterior pair consists of extremely flattened, large, branching ossicones [6,17], very unlike the posterior ossicone FRK-13b. The ossicones of S. hendeyi are very similar in shape to the ossicones of the Iberian sivatheres D. rex and Bi. schaubi, although larger [44]; hence, they are different from FRK-13. The posterior ossicone is also more curved than FRK-13b. Libytherium is a taxon closely related to Sivatherium, known by the species Libytherium maurisium (Pomel, 1893) and L. proton. It is uncertain whether L. maurisium and L. proton have one or two pairs of ossicones. L. maurisium bears very curved ossicones with massive bumps [9,17,67], whereas L. proton’s ossicones have a general shape similar to those of D. rex, Bi. Schaubi, and S. hendeyi; hence, they are more curved than that of FRK-13.
Actually, the ossicones of FRK-13 seem identical to those of Bramatherium in that there is a pair of frontoparietal ossicones arising from a common base and a pair of laterally oriented parietal ossicones (Figure 3). Two Bramatherium species are known by their ossicones, B. perimense and B. megacephalum. The main difference between the two species is that the ossicones of B. perimense are reported as stouter and shorter. FRK-13 ossicones are dimensionally closer to B. megacephalum; the anterior ossicone FRK-13a has a length of about 335–340 mm from fork to tip, similar to the anterior ossicones of B. megacephalum, which have a length of 350 mm [27]. It was not possible to precisely measure the length of the posterior ossicone FRK-13b due to the way it is broken. Bramiscus micros Ríos, Abbas, Khan & Solounias, 2024, from the Middle Miocene of Pakistan possesses similar ossicones to B. megacephalum, though much smaller.
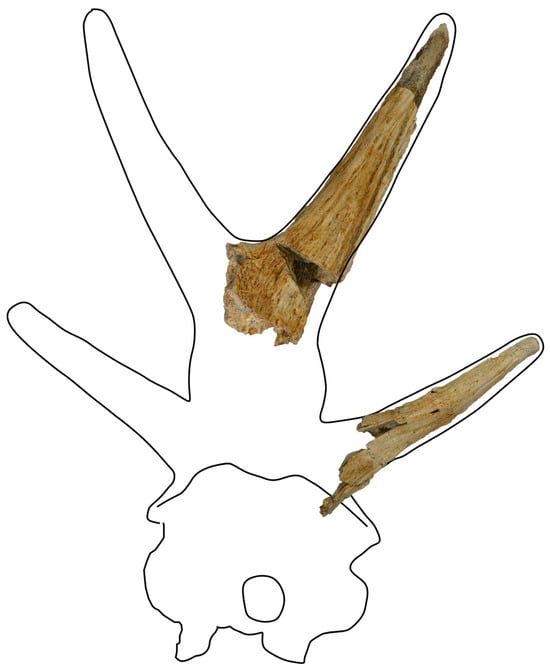
Figure 3.
The ossicones FRK-13 plotted upon the Bramatherium megacephalum cranium. The sketch of the cranium is based on YPM 13881 (Lewis, 1939: Pl. 2). Images are not to scale.
3.2. Phylogenetic Analysis
The phylogenetic analysis produced two most parsimonious trees (MPT; Figure 4) of 276 steps each, in which FRK-13 groups along with either B. megacephalum or B. perimense in a monophyletic clade. The node containing the FRK13, B. perimense and B. megacephalum, demonstrates a relatively high Bremer Support of 3 and relatively high Jacknife and Bootstrap values (79 and 75, respectively). The FRK-13 shares four synapomorphies with B. megacephalum and B. perimense: they have no Decennatherium-like ossicone core, the anterior ossicones are fused at the base, the ossicones are slightly curved, and the ossicones of the anterior pair are larger than the posterior ones. These results confirm the taxonomic assignment of FRK-13 as Bramatherium although without distinguishing between B. perimense and B. megacephalum.
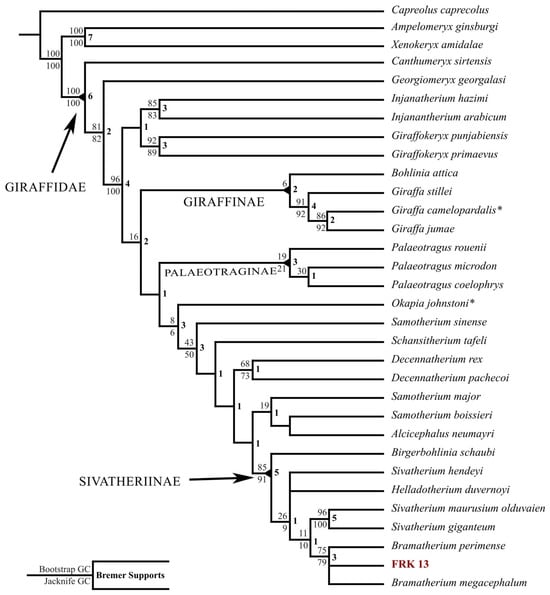
Figure 4.
Consensus tree of the two most parsimonious trees (276 steps). Extant taxa are indicated with *.
4. Discussion
4.1. Systematics of Bramatherium and the Assignment of FRK-13
The taxonomy of the large Eurasian Late Miocene sivatheres has been quite perplexing. That is especially true for the taxa known from Southwest Asia. Falconer [25] erected Bramatherium, with its type species being B. perimense, from the Piram Island of India based on a cranium. Later on, Lydekker [68,69] and then Pilgrim [70,71] raised several new genera and species (e.g., Hydaspidotherium megacephalum, Hydaspitherium megacephalum, Hydaspitherium magnum, Hydaspitherium grande, and Helladotherium grande) based on material from the Middle Siwaliks. Matthew [41] and Colbert [26] acknowledged that differences among the aforementioned genera are rather subtle but kept them distinct. Lewis [27] described a well-preserved cranium of B. megacephalum (YPM 13881; Lewis, 1939: Pl. 2) and included all previous species in the genus Bramatherium. The viewpoint of Lewis [27] that Bramatherium is actually the only large-sized sivathere genus from the Late Miocene of Siwaliks (and Southern–Southwestern Asia in general, with the exception of H. duvernoyi from Maragheh [23]) has been widely accepted to this day [1,29,30,39,72]. The intrageneric diversity of Bramatherium has been a subject of discussion ever since Lewis [27] assigned all taxa to just two species: B. perimense and B. megacephalum. Recently, Khan et al. [29,30] synonymized Helladotherium grande and Hydaspitherium magnum, under Bramatherium grande, suggesting that their subtle differences might be due to sexual dimorphism or ontogenetic variation. They kept B. megacephalum as a distinct species based on its smaller size [29], although they admitted that more material is needed for more solid claims. In fact, the distinction between B. perimense and B. megacephalum is also dubious. Their main reported difference is that the ossicones of the latter are slenderer and longer, but as Lewis [27] already stated, the cranial differences between these species “are of degree rather than of kind”, and it should not be excluded that there might be differences due to sexual dimorphism as well. Geraads & Güleç [39] described a juvenile skull and some postcranial bones from Kavakdere, Türkiye, which constitute the westernmost occurrence of Bramatherium to date. Based on the ossicone morphology, they attributed it to B. perimense but also stressed that the cranium YPM 13881 of B. megacephalum [27] belongs to a very old individual, and thus, B. megacephalum and B. perimense could be included in the same species, and their differences may be attributed to different ontogenetic stages. Harris [73], Ríos & Morales [74], and Ríos et al. [6,58] have already shown that the ossicones of sivatheres demonstrate extreme variability, and hence, observed small differences may be inadequate to segregate the two taxa. Dental and postcranial remains of B. perimense and B. megacephalum are extremely scarce and, for the most part, are insufficient for detailed comparisons. Metatarsal comparisons show that B. perimense and B. megacephalum have, to some extent, overlapping length distributions, whereas their robusticity index (R.I.; breadth in the middle of the shaft/total length of the shaft*100) is indistinguishable (Supp. File 1). Hamilton [1] and Khan et al. [30] also considered the possibility that B. megacephalum and B. perimense could be conspecific, but they kept them apart for the time being, until a more thorough analysis is possible.
The phylogenetic analysis showed that FRK-13 is indeed more closely related to the Bramatherium clade, but as we lack postcranial, dental, or cranial features, a higher resolution is prohibited. Bramatherium grande is not included in our phylogenetic analysis. However, an assignment of the FRK large giraffid to B. grande is excluded since the dimensions of the M3 (AMPG(V)-110) are much smaller than those of B. grande and within the size range of both B. perimense and B. megacephalum (Figure 5). Based on the ossicone slenderness and elongation, the FRK-13 specimens are similar to B. megacephalum.
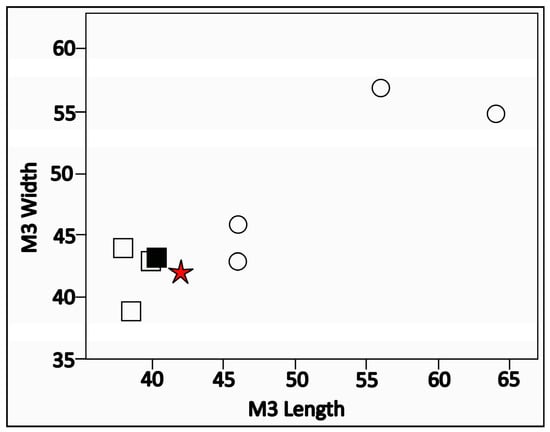
Figure 5.
Scatter plot with the dimensions of the M3 of various Bramatherium species. White square: Bramatherium megacephalum; black square: Bramatherium perimense; white circle: Bramatherium grande; red star: AMPG(V)-110.
4.2. Paleobiogeography of Bramatherium
For the time being, the easternmost occurrences of Bramatherium are from the early Late Miocene of Thailand and Myanmar where dental material, two metatarsals, and a naviculocuboid are referred to as Bramatherium sp. [31]. Danowitz et al. [32] also cite Bramatherium sp. from North China, whereas there are dubious findings assigned to Bramatherium indet. from the United Arab Emirates [33].
Solounias and Jukar [40]: Figure 12.6 recently suggested reassigning the juvenile cranium PIM-298 from Samos to Bramatherium based on the presence of a median boss located slightly caudally to the posterior orbital rims, the short and wide shape of the auditory bullae, the presence of occipital fossae, and the morphology of the deciduous teeth. Apart from the fact that the specimen illustrated in their Figure 12.6A1 and A3 (A and C, respectively, in their legend) is PIM-299, while the specimen illustrated in Figure 12.6A2 (B in their legend) is neither PIM-298 nor PIM-299, none of the authors’ arguments confirm their taxonomic conclusion. A bump with a smooth surface medially located just behind the coronal suture is a common character in Samotherium, as evidenced by several crania (e.g., NHML M4215, NHMA MTLA-540, and NHMB Sa-29); in young individuals such as PIM-298, PIM-299 likely appears less localized than in adult ones. Both Bramatherium and Samotherium show laterally compressed auditory bullae; however, they are more curved in the former taxon. The morphology and proportion of the auditory bulla of PIM-298 perfectly match the illustrated PIM-299, as well as that of other Samotherium crania (e.g., NHMA MTLA-540). The morphology of the occipital face is difficult to fully assess in PIM-298 as the ventral half is missing but, in any case, is fully compatible with that of PIM-299 and NHMB Sa-29 (ossicone-less, adult); it sharply differs from the strongly curved lateral outline seen in Bramatherium and Helladotherium crania. Finally, although the morphology of DP2 and DP3 in PIM-298 and in PIM-299 deviates slightly from the usual morphology seen in Samotherium, it is completely different from that of Bramatherium and Helladotherium in the less-well-defined hypocone and stronger entostyle associated with a mesially oriented basal flange/pillar on DP2, the presence of a more molariform (crescentic) anterior lobe on DP3, and the presence of basal pillars on both of these deciduous premolars. Based on this information, we consider that the presence of Bramatherium in Samos has not been proven to date.
The type species B. perimense is known from the type locality of Piram Island, India [25], and Kavakdere, Türkiye [7,39]. Welcomme et al. [28] suggested that B. perimense could be present in Dera Bugti, Pakistan, and Colbert [26], Mahmood et al. [75], Draz et al. [76], and Raza et al. [10] cited B. perimense from the Middle Siwaliks. We should, however, note that B. perimense from Dera Bugti and the Siwaliks are not properly described to our knowledge, except for the partial skull AM 19771 attributed to B. perimense by Colbert [26]. However, that skull only preserves the bases of the anterior and posterior ossicones. By what is illustrated, it cannot be excluded that it belongs to B. megacephalum. Hence, we consider that further confirmation is needed on the presence of B. perimense in Dera Bugti and the Siwalkis. Excluding doubtful occurrences, a huge geographical gap appears between the only two solid records of B. perimense; the Piram Island and the Kavakdere in Anatolia. The area in between is occupied by either B. grande or B. megacephalum, which are definitely known to originate from Middle Siwaliks [27,29,30]. Currently, there are no known occurrences of B. grande or B. megacephalum outside the Middle Siwaliks. The existence of FRK-13 in the Late Miocene of Fourka constitutes the westernmost occurrence of a B. megacephalum-like giraffid to date, as well as of Bramatherium in general, significantly expanding the geographical range of the genus (Figure 6). Although several Late Miocene mammalian taxa expanded from East Asia (e.g., China) to the Balkans, Bramatherium is one of the few ruminants (e.g., along with Tragoportax Pilgrim, 1937, and Nisidorcas Bouvrain, 1979) that connect the Indian subcontinent to the Aegean region [17].
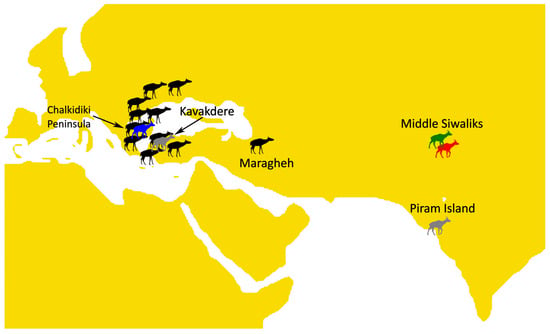
Figure 6.
Late Miocene map showing the distribution of the large sivatheres. Black sketch: Helladotherium duvernoyi; gray sketch: Bramatherium perimense; red sketch: Bramatherium megacephalum; green sketch: Bramatherium grande; blue sketch: FRK Bramatherium. Map adapted from Ríos et al. [34]. Bramatherium silhouette by Steven Traver, adapted and licensed under CC0 1.0 “https://creativecommons.org/publicdomain/zero/1.0/ (accessed on 8 July 2025)”. Helladotherium silhouette by Zimices (Julián Bayona), adapted and licensed under CC BY-SA 3.0 “https://creativecommons.org/licenses/by-sa/3.0/ (accessed on 8 July 2025)”.
The fact that the distributions of B. megacephalum and B. perimense show such large but suspicious gaps might be another indication that the two taxa are indeed conspecific, but their classification into different species was biased by the rareness of the fossil record and was since retained for historical reasons. Namely, all the material from Kavakdere and Piram Island was classified as B. perimense because crania with stout ossicones were found in those sites, whereas smaller Bramatherium from the Middle Siwaliks with long, slenderer ossicones was identified as B. megacephalum [27] (AM 13881). If the differences on the ossicones are indeed due to ontogenetic reasons, as suggested by Geraads & Güleç [39], then all the material from the Piram Island to Fourka could be associated with just a single species.
4.3. Is Helladotherium a Female Bramatherium?
Matthew [41] proposed for the first time that Helladotherium and Bramatherium could be congeneric, and Colbert [26] pointed out that the lack of ossicones in Helladotherium suggests that it could represent a female Bramatherium. Geraads & Güleç [39] observed that the crania of those taxa are very similar to each other and Geraads [17] tentatively proposed that these two genera might indeed represent two sexes of the same species. Apart from Kavakdere, these two genera have never been found in the same locality, or at least in neighboring sites [7]. Chalkidiki Peninsula would be the second area in which Bramatherium and Helladotherium are reported in proximity, as dental and postcranial material of H. duvernoyi has also been found in the latest Vallesian locality of Nikiti-1 [21]. At the same time, the local overlap of their records again calls into question their taxonomic distinctiveness.
Co-existence of two large herbivores, very close phylogenetically and very similar morphometrically, might be expected when they occupy different niches. Instead, in a mesowear analysis, Danowitz et al. [32] showed that both Helladotherium and Bramatherium were predominantly browsers. We should note that Bramatherium showed patterns of mixed feeding (also supported by a dental microwear study by Patnaik [77]), whereas the Helladotherium sample appeared more homogenous (also supported by a dental microwear study by Merceron et al. [78]). Danowitz et al. [32] studied Bramatherium from North China and Helladotherium from Pikermi and Samos, whereas Patnaik [77] studied Bramatherium from the Siwaliks, and Merceron et al. [78] studied Helladotherium from Axios Valley, Greece. The studied giraffid material therefore represents populations that lived in remote sites and at different time slices, and alternative methods were applied in each study. Hence, the suggested differences in the paleodietary profile of Bramatherium and Helladotherium might just reflect flexibility in food resources and be fully justified.
The hypothesis of the very morphologically similar Helladotherium and Bramatherium occupying different ecological niches in a single locality they are reported to coexist (Kavakdere, Türkiye) cannot unfortunately be tested due to the insufficiency of dental material. Xafis et al. [7] assigned very scarce postcranial material from Kavakdere to B. perimense, namely two metatarsals and a proximal phalanx. According to these authors, Bramatherium metatarsals are distinguished from those of H. duvernoyi from the same site by the presence of a pygmaios and a remnant of the fifth metatarsal, features which, according to Ríos et al. [5], are missing from the latter taxon. However, we were able to observe clear pygmaios in H. duvernoyi metatarsals from the Turolian locality of Perivolaki [19], from Samos [20], and from Pikermi. Additionally, Yohe & Solounias [79] proposed that a remnant of the fifth metatarsal exists in every giraffid. Xafis et al. [7] also used the irregularly shaped articular surface for the medial cuneiforme to assign those metatarsals to Bramatherium. These surfaces are, in our opinion, not so different from the circular ones observed in H. duvernoyi. The phalanx from Kavakdere (PV-226) is larger than any known phalanx of H. duvernoyi (as well as of any phalanx of Bramatherium), and for that reason, Xafis et al. [7] attributed it to B. perimense. However, no other phalanx has ever been assigned to B. perimense, so the assignment of PV-226 in that taxon is rather arbitrary. In fact, PV-226 is very close dimensionally to PIM-412 from Samos and FM-2323 from Strumyani, Bulgaria, both assigned to H. duvernoyi by Kostopoulos [20] and Geraads et al. [80], respectively (Figure 7A). In conclusion, in Kavakdere, we either have two very similar taxa, possibly occupying very similar niches, with the first one only definitely known from cranial material (B. perimense) and the other one only known from postcranials (H. duvernoyi), or all of the referred Kavakdere sivatheriine material should be attributed to a single taxon. Evidence is in favor of the latter scenario.
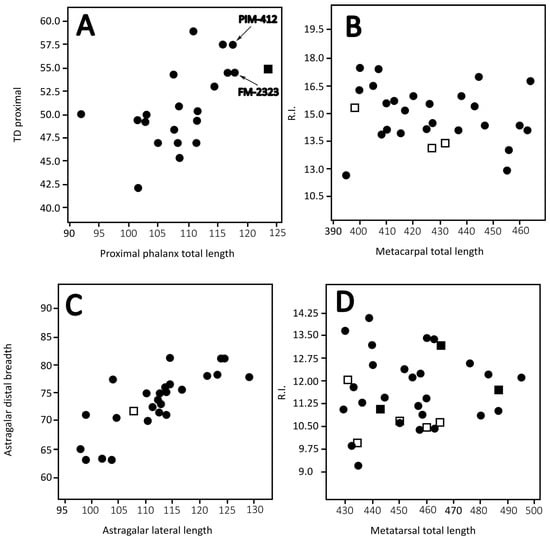
Figure 7.
Scatter plots of the dimensions of several postcranials of Helladotherium and Bramatherium. Black circle: Helladotherium duvernoyi; black square: Bramatherium perimense; white square: Bramatherium megacephalum. Proximal Phalanx total length—proximal TD (A); Metacarpal total length—R.I. (B); Astragalus lateral length—distal breadth; (C) and Metatarsal total length—R.I. (D).
Solounias & Danowitz [81] mentioned slight differences between the astragali of Bramatherium and Helladotherium. As they stated, the lateral notch between the head and trochlea is very prominent in Bramatherium, whereas it is more subtle in Helladotherium. On the contrary, the lateral ridge of the trochlea is notably thicker in Helladotherium. Ríos et al. [5,82] added that the epicondyles in Bramatherium’s metacarpals splay significantly more than in Helladotherium, and that the medial ridge in Bramatherium metatarsals is much thicker than the lateral one.
The splaying of the epicondyles of the metacarpals is in our opinion statistically insignificant in these two taxa. The thicker medial ridge of Bramatherium metatarsals [82] is another dubious character. Xafis et al. [7] ascribed a metatarsal to H. duvernoyi from Kavakdere (PV-152), which lacks a pygmaios and demonstrates an extremely thick medial ridge compared to the lateral [7] (Figure 2H). However, in the H. duvernoyi metatarsals we studied from Pikermi and Perivolaki, the medial ridge was always thicker than the lateral, although the differences were not so great sometimes, as those demonstrated by Ríos et al. [82] and Xafis et al. [7]. As for the astragali, we detected a pronounced lateral notch in a cast of a H. duvernoyi astragalus from Samos (LGPUT MGPP-C 66), very similar to the one sketched by Solounias & Danowitz [81] for B. megacephalum. Moreover, Solounias & Danowitz [81] proposed that the lateral ridge of the trochlea of the astragalus is notably thicker in H. duvernoyi than in B. megacephalum; however, there are no biometrical data for solid comparisons. Solounias & Danowitz [81] based their differences on rather limited material (four astragali for Helladotherium and two for Bramatherium), and as a result, the intrageneric or intraspecific diversity could not be addressed. Given that postcranial variability tends to increase with body mass in ruminants (e.g., [83]), we either found no significant morphological postcranial differences between Helladotherium and Bramatherium, or, in some cases, the available material was inadequate and failed to account for possible intrageneric or intraspecific disparity (e.g., the astragali).
Although we did not have the opportunity to perform cranial comparisons between Bramatherium and Helladotherium in this work, previous scholars clearly showed that they are very similar as well [6,27,39]. The dental morphology also supports this conclusion. The p3s of H. duvernoyi demonstrate the sivatheriine synapomorphy of a continuous talonid wall that isolates a reduced entocristid (e.g., [6,21]). The only published p3 of Bramatherium belongs to B. megacephalum [26] (AM 19669) and retains a primitive condition without a talonid wall and an isolated reduced entocristid. However, Ríos et al. [6] classify both B. megacephalum’s and B. perimense’s p3s as apomorphic in their dataset, and they also describe that synapomorphy as “Bramatherium-like” [12] (S1; character 80). Nevertheless, there is at least one H. duvernoyi p3 from the type locality of Pikermi (AS II 645) with primitive features, identical to AM 19669. Hence, not only do both taxa have similar p3s, but they also seem to demonstrate the same patterns of intraspecific variation. The p4 is also diagnostic for Giraffidae. In both Helladotherium and Bramatherium, that tooth is completely molarized, and its distal lobe is relatively enlarged compared to other lineages of giraffids [6].
The biometrical comparisons between the two taxa are also very limited, as very scarce material is published for Bramatherium. Except for a single radius from Kavakdere [39], which is distinct by any known H. duvernoyi, all of the other postcranials of Bramatherium and Helladotherium are rather indistinguishable (Figure 7). We should also note that since Xafis et al. [7] also cited smaller giraffids than Helladotherium and Bramatherium from Kavakdere, the radius studied by Geraads & Güleç [39] could belong to one of these smaller taxa.
In the most inclusive phylogenetic analysis of Giraffidae to date [6], Bramatherium was not found to be the sister taxon of Helladotherium, but it was recovered in a monophyletic clade with S. maurisium/olduvaiense and S. giganteum, as in our analysis (Figure 4). A large percentage of that dataset comprises ossiconal characters, whereas the postcranial characters are rather limited. Hence, the fact that Bramatherium and Helladotherium were not recovered in a monophyletic clade is attributed to the latter’s lack of ossicones.
5. Conclusions
In conclusion, Helladotherium and Bramatherium are two taxa with highly overlapping distributions for the most part (and supposedly coexisting in the Late Miocene faunas of the Chalkidiki Peninsula and Kavakdere), with very similar biometry, indistinguishable dental morphology, similar cranial morphology, and minor morphological postcranial differences. They also seem to occupy a similar niche. It is therefore highly probable that they are congeneric, and the westernmost remains are attributed to Helladotherium, whereas the easternmost remains are attributed to Bramatherium for historical reasons. The absence of Helladotherium ossicones in the Balkans contributed traditionally to that distinction but the Fourka record challenges this condition. The fact that they were not recovered as sister taxa in relevant phylogenetic analyses is obviously due to the lack of ossiconal features for Helladotherium.
Considering these findings, we suggest synonymizing the two genera under the name Bramatherium, which has priority. The available material and studies are inadequate to segregate the two taxa, and we believe that the question of their synonymy could be addressed with higher confidence through (a) the enrichment of the Bramatherium fossil record, especially eastwards of the Greco-Irano-Afghan Province; (b) more thorough studies of the Bramatherium morphology, both cranial and postcranial; and (c) a phylogenetic analysis that is more character-inclusive than the existing ones.
Bramatherium grande from the Middle Siwaliks is distinguished from the other Bramatherium species due to its larger size. The distinction of B. perimense from B. megacephalum is based solely on the differences in the ossicones, whereas the material previously attributed to H. duvernoyi could be classified as either B. perimense or B. megacephalum, as we did not detect any serious biometrical or morphological differences among the three species. We consider the size differences in the ossicones as inadequate to segregate the taxa, especially since Ríos et al. [6,58] and Ríos & Morales [74] have shown that in D. rex (which belongs to the same subfamily with Bramatherium), there is extreme intrageneric variation in the ossiconal size. Hence, all the material of the small Bramatherium could belong to just one species, with B. perimense having priority over the two other names. The synonymization of the three species under B. perimense also resolves the biogeographic issues emerging from the previously recorded mosaic distribution for B. megacephalum and B. perimense.
The large ossicones unearthed in the Late Miocene locality of Fourka, Chalkidiki, are classified with certain confidence to Bramatherium. The size of the associated M3 (AMPG(V)-110) excludes an assignment to B. grande. Dimensionally the FRK-13 ossicones are very close to those belonging to the skull YPM 13881, previously assigned to B. megacephalum. After synonymizing B. perimense and B. megacephalum, we assign FRK-13 in B. perimense.
In summary, the findings of Bramatherium ossicones in Fourka, so close to Helladotherium bearing localities, enabled us to debate the synonymization of Helladotherium with Bramatherium on a new basis. The proposed synonymization, along with the attribution of the smaller species in B. perimense, is the most parsimonious taxonomic solution given the currently available material. This approach resolves the biogeographic issues created by historical nomenclatural conventions and provides a more coherent taxonomic framework for interpreting the distribution of the genus. Nevertheless, a more definitive assessment of the genus’ taxonomy will depend on the recovery of additional and, principally, more diverse material, particularly from regions east of the Greco-Irano-Afghan Province and especially from the Indian Subcontinent.
Supplementary Materials
The following supporting information can be downloaded at https://www.mdpi.com/article/10.3390/fossils3040017/s1, S1: Measurements; S1: Phylogenetic Matrix.
Author Contributions
Conceptualization, K.L. and D.S.K.; methodology, K.L., E.V., and D.S.K.; software, K.L. and E.V.; validation, G.L., E.T., and D.S.K.; formal analysis, K.L.; data curation, G.L. and E.T.; writing—original draft preparation, K.L.; writing—review and editing, G.L., E.T., and D.S.K.; visualization, K.L.; supervision D.S.K. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Data Availability Statement
The research data are shared in the Supplementary Files.
Acknowledgments
We thank all the colleagues who worked during the past years to unearth and prepare the fossil material from Fourka and to the local Xanthos Makris who found the FRK13 ossicones and provided them to the excavating team. The first author was benefited by Erasmus+ Short-Term Doctoral Program to visit Spain and study Iberian giraffids. Special thanks to Soledad Domingo at the Complutense University of Madrid for being the hostess of the first author in Madrid and Elena Carrión, curator at the Regional Archaeological Museum of Madrid (MARPA), in Alcalá de Henares, and all her colleagues for providing access to the rich ossiconal material of D. rex. We also thank Socrates Roussiakis from the National and Kapodistrian University of Athens for allowing us to access the Pikermian giraffid material. Furthermore, we thank George D. Koufos and all the workers in the Natural History Museum of Aegean in Samos (NHMA) for allowing us to investigate the rich giraffid material there. Last but not least, we thank our two anonymous reviewers as their suggestions improved the quality of our manuscript. The Willi Hennig Society sponsors the use of TNT 1.5 software.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Hamilton, W.R. Fossils giraffes from the Miocene of Africa and a revision of the phylogeny of the Giraffoidea. Phil. Trans. R. Soc. Lond. 1978, B283, 165–229. [Google Scholar]
- Geraads, D. Remarques sur la systématique et la phylogénie des Giraffidae (Artiodactyla, Mammalia). Geobios 1986, 19, 465–477. [Google Scholar] [CrossRef]
- Montoya, P.; Morales, J. Birgerbohlinia schaubi Crusafont, 1952 (Giraffidae, Mammalia) del Turoliense inferior de Crevillente-2 (Alicante, España). Filogenia e historia biogeográfica de la subfamilia Sivatheriinae. Bull. Mus. Natl. Hist. Nat. Paris 1991, 13, 177–200. [Google Scholar]
- Danowitz, M.; Vasilyev, A.; Kortlandt, V.; Solounias, N. Fossil evidence and stages of elongation of the Giraffa camelopardalis neck. R. Soc. Open Sci. 2015, 2, 150393. [Google Scholar] [CrossRef]
- Ríos, M.; Danowitz, M.; Solounias, N. First comprehensive morphological analysis on the metapodials of Giraffidae. Palaeontol. Electron. 2016, 19.3.50A, 1–39. [Google Scholar] [CrossRef]
- Ríos, M.; Sánchez, I.M.; Morales, J. A new giraffid (Mammalia, Ruminantia, Pecora) from the Late Miocene of Spain, and the evolution of the sivathere-samothere lineage. PLoS ONE 2017, 12, e0185378. [Google Scholar] [CrossRef]
- Xafis, A.; Mayda, S.; Grímsson, F.; Nagel, D.; Kaya, T. Fossil Giraffidae (Mammalia, Artiodactyla) from the early Turolian of Kavakdere (Central Anatolia, Türkiye). Comptes Rendus Palevol 2019, 18, 619–642. [Google Scholar] [CrossRef]
- Laskos, K.; Kostopoulos, D.S. A review of Palaeogiraffa (Giraffidae, Mammalia) from the Vallesian of the Eastern Mediterranean. Geobios 2024, 84, 25–43. [Google Scholar] [CrossRef]
- Ríos, M.; Abbas, S.G.; Khan, M.A.; Solounias, N. Distinction of Sivatherium from Libytherium and a new species of Libytherium (Giraffidae, Ruminantia, Mammalia) from the Siwaliks of Pakistan (Miocene). Geobios 2022, 74, 67–76. [Google Scholar] [CrossRef]
- Raza, T.; Yasin, R.; López-Torres, S.; Warburton, N.M.; Samiullah, K.; Ghaffar, A.; Khan, M.N.; Ara, C.; Muzaffar, E. New sivatheriine giraffid (Ruminantia, Mammalia) craniodental material from the Siwaliks of Pakistan. J. Vertebr. Paleontol. 2024, 44, e2376241. [Google Scholar] [CrossRef]
- Crusafont, M. Los jiráfidos fósiles de España. In Memorias y Comunicaciones del Instituto Geológico VIII: Premio Extraordinario de Doctorado; Diputación Provincial de Barcelona: Barcelona, Spain, 1952. [Google Scholar]
- Ríos, M.; Sánchez, I.M.; Morales, J. Comparative anatomy, phylogeny, and systematics of the Miocene giraffid Decennatherium pachecoi Crusafont, 1952 (Mammalia, Ruminantia, Pecora): State of the art. J. Vertebr. Paleontol. 2016, 36, e1187624. [Google Scholar] [CrossRef]
- de Bonis, L.; Brunet, M.; Heintz, E.; Sen, S. La province Gréco-Irano-Afghane et la répartition des faunes mammaliennes au Miocène supérieur. Paleontol. Evol. 1992, 24–25, 103–112. [Google Scholar]
- de Bonis, L.; Bouvrain, G. Nouveaux Giraffidae du Miocène supérieur de Macédoine (Grèce). Adv. Vertebr. Paleontol. Hen Panta 2003, 5–16. [Google Scholar]
- Gentry, A.W. Ruminantia (Artiodactyla). In Geology and Paleontology of the Miocene Sinap Formation, Türkiye; Fortelius, M., Kappelman, J., Sen, S., Bernor, R., Eds.; Columbia University Press: New York, NY, USA, 2003; pp. 332–379. [Google Scholar]
- Geraads, D. Les Giraffidés du Miocène Supérieur de la Région de Thessalonique (Grèce). Unpublished. Ph.D. Thesis, University of Paris, Paris, France, 1974. [Google Scholar]
- Geraads, D. Giraffidae (Mammalia) de la fin du Néogène de la République de Macédoine (ARYM). Geodiversitas 2009, 31, 893–908. [Google Scholar] [CrossRef]
- Iliopoulos, G. The Giraffidae (Mammalia, Artiodactyla) and the Study of the Histology and Chemistry of Fossil Mammal Bone from the Late Miocene of Kerassia (Euboea Island, Greece). Unpublished. Ph.D. Thesis, University of Leicester, Leicester, UK, 2003. [Google Scholar]
- Kostopoulos, D.S.; Koufos, G.D. The Late Miocene vertebrate locality of Perivolaki, Thessaly, Greece. 8. Giraffidae. Palaeontogr. Abt. 2006, A 276, 135–149. [Google Scholar] [CrossRef]
- Kostopoulos, D.S. The Late Miocene mammal faunas of the Mytilinii Basin, Samos Island, Greece: New collection: 13. Giraffidae. In The Late Miocene Mammal Faunas of Samos; Koufos, G.D., Nagel, D., Eds.; Beiträge zur Paläontologie: Vienna, Austria, 2009; pp. 299–343. [Google Scholar]
- Kostopoulos, D.S. Palaeontology of the upper Miocene vertebrate localities of Nikiti (Chalkidiki Peninsula, Macedonia, Greece): Artiodactyla. Geobios 2016, 49, 119–134. [Google Scholar] [CrossRef]
- Lazaridis, G. Study of the Late Miocene vertebrate locality of Kryopigi and other localities of Kassandra Peninsula, Chalkidiki (Greece). Systematics, Taphonomy, Paleoecology, Biochronology. Unpublished. Ph.D. Thesis, Scientific Annals of the School of Geology, Aristotle University, Thessaloniki, Greece, 2015. (In Greek). [Google Scholar]
- Solounias, N.; Danowitz, M. The Giraffidae of Maragheh and the identification of a new species of Honanotherium. Palaeobiol. Palaeoenviron. 2016, 96, 489–506. [Google Scholar] [CrossRef]
- Xafis, A.; Tsoukala, E.; Solounias, N.; Mandic, O.; Harzhauser, M.; Grímsson, F.; Nagel, D. Fossil Giraffidae (Mammalia, Artiodactyla) from the late Miocene of Thermopigi (Macedonia, Greece). Palaeontol. Electron. 2019, 22.3.67, 38. [Google Scholar] [CrossRef]
- Falconer, H. Description of some fossil remains of Dinotherium, Giraffe and other Mammalia, from the Gulf of Cambay, Western Coast of India, Chiefly from the collection presented by Captain Fulljames, of the Bombay Engineers, to the Museum of the Geological Society. Q. J. Geol. Soc. 1845, 1, 356–372. [Google Scholar] [CrossRef]
- Colbert, E.H. Siwalik Mammals in the American Museum of Natural History. Trans. Am. Philos. Soc. 1935, XXVI, 294–377. [Google Scholar]
- Lewis, G.E. A new Bramatherium Skull. Am. J. Sci. 1939, 237, 275–280. [Google Scholar] [CrossRef]
- Welcomme, J.L.; Antoine, P.O.; Duranthon, F.; Mein, P.; Gingsburg, L. Nouvelles découvertes de Vertébrés miocènes dans le synclinal de Dera Bugti (Balouchistan, Pakistan). C. R. Acad. Sci. Paris. 1997, 325, 531–536. [Google Scholar] [CrossRef]
- Khan, M.A.; Akhtar, M.; Irum, A. Bramatherium (Artiodactyla, Ruminantia, Giraffidae) from the Middle Siwaliks of Hasnot, Pakistan: Biostratigraphy and palaeoecology. Turk. J. Earth Sci. 2014, 23, 308–320. [Google Scholar] [CrossRef]
- Khan, M.A.; Babar, M.A.; Ríos, M. New material of Bramatherium grande from the Siwaliks of Pakistan sheds light on dental intraclade morphological variability of Late Miocene sivatheres. J. Vertebr. Paleontol. 2021, 41, e1898976. [Google Scholar] [CrossRef]
- Nishioka, Y.; Hanta, R.; Jintasakul, P. Note on giraffe remains from the Miocene of continental Southeast Asia. J. Sci. Technol. MSL 2014, 33, 365–377. [Google Scholar]
- Danowitz, M.; Hou, S.; Mihlbachler, M.C.; Hastings, V.; Solounias, N. A combined-mesowear analysis of Late Miocene giraffids from North Chinese and Greek localities of the Pikermian Biome. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2016, 449, 194–204. [Google Scholar] [CrossRef]
- Whybrow, P.J.; Andrew, H.P. Fossil Vertebrates of Arabia: With Emphasis on the Late Miocene faunas, Geology, and Palaeoenvironments of the Emirate of Abu Dhabi, United Arab Emirates; Yale University Press: New Haven, CT, USA, 1999. [Google Scholar]
- Ríos, M.; Danowitz, M.; Solounias, N. First identification of Decennatherium Crusafont, 1952 (Mammalia, Ruminantia, Pecora) in the Siwaliks of Pakistan. Geobios 2019, 57, 97–110. [Google Scholar] [CrossRef]
- Gaudry, A. Animaux fossiles du Mont Luberon (Vaucluse). Étude sur les vertébrés. F. Savy Editions; Biodiversity Heritage Library: Paris, France, 1873. [Google Scholar]
- Godina, A.Y. Istoricheskoe razvitie zhiraf. Rod Palaeotragus [Historical Development of Girrafids. Genus Palaeotragus]. Trudy 177; Akademia Nauk USSR, Paleontologicheskii Institut: Moscow, Russia, 1979. (In Russian) [Google Scholar]
- Kaakinen, A. NOW_20200330_all_Basins_combined_AK.xls. NOW Locality Basin Data Import 2020. Available online: https://nowdatabase.luomus.fi/ (accessed on 27 October 2025).
- Ţibuleac, P.; Laskos, K.; Rӑţoi, B.-G.; Haiduc, B.S.; Merlan, V.; Ursachi, L. A link of the Late Miocene giraffid migration pathway from the peri-Aegean lands to the northeastern Eurasian areas. Geobios 2025, 88–89, 251–263. [Google Scholar] [CrossRef]
- Geraads, D.; Güleç, E. A Bramatherium skull (Giraffidae, Mammalia) from the Late Miocene of Kavakdere (Central Türkiye) Biogeographic and phylogenetic implications. Bull. Miner. Res. Explor. 1999, 121, 51–56. [Google Scholar]
- Solounias, N.; Jukar, A.M. A Reassessment of Some Giraffidae Specimens from the Late Miocene Faunas of Eurasia. In Evolution of Cenozoic Land Mammal Faunas and Ecosystems; Casanovas-Vilar, I., van den Hoek Ostende, L., Janis, C.M., Saarinen, J., Eds.; Springer Nature: Cham, Switzerland, 2023; pp. 189–200. [Google Scholar]
- Matthew, W.D. Critical observations upon Siwaliks mammals. Bull. Am. Mus. Nat. Hist. 1929, LVI, 437–560. [Google Scholar]
- Arambourg, C. Precisions nouvelles sur Libytherium maurusium Pomel, giraffide du Villafranchien d’Afrique. Bull. De La Société Géologique De Fr. 1960, S7-II, 888–894. [Google Scholar] [CrossRef]
- Hendey, Q.B. Quaternary Vertebrate Fossil Sites in the South-Western Cape Province. S. Afr. Archaeol. Bull. 1969, 24, 96–105. [Google Scholar] [CrossRef]
- Harris, J.M. Pliocene Giraffoidea (Mammalia, Artiodactyla) from the Cape Province. Ann. S. Afr. Mus. 1976, 69, 325–353. [Google Scholar]
- Geraads, D. Le Sivatherium (Giraffidae, Mammalia) du Pliocène final d’Ahl al Oughlam (Casablanca, Maroc) et l’evolution du genre en Afrique. Paläontologische Z. 1996, 70, 623–629. [Google Scholar] [CrossRef]
- Khan, A.A.; Khan, M.A.; Iqbal, M.; Akhtar, M.; Sarwar, M. Sivatherium (Artiodactyla, Ruminantia, Giraffidae) from the Upper Siwaliks, Pakistan. J. Anim. Plant Sci. 2011, 21, 202–206. [Google Scholar]
- Abel, O. Über einen Fund von Sivatherium giganteum bei Adrianopel. Sitzungsberichte Der Math. Naturwissenschaftlichen Kl. Der Kais. Akad. Der Wiss. 1904, 113, 1–22. [Google Scholar]
- Ríos, M.; Montoya, P.; Morales, J.; Romero, G. First occurrence of Sivatherium Falconer and Cautley, 1836 (Mammalia, Ruminantia, Giraffidae) in the Iberian Peninsula. J. Vertebr. Paleontol. 2021, 41, e1985507. [Google Scholar] [CrossRef]
- Koufos, G.D.; Syrirdes, G.E. A new Early/Middle Miocene mammal locality from Macedonia, Greece. Comptes Rendus De Académie Des Sci.-Ser. IIA-Earth Planet. Sci. 1997, 325, 511–516. [Google Scholar] [CrossRef]
- Koufos, G.D. Palaeoecology and chronology of the Vallesian (late Miocene) in the Eastern Mediterranean region. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2006, 234, 127–145. [Google Scholar] [CrossRef]
- Koufos, G.D. Carnivores from the early/middle Miocene locality of Antonios (Chalkidiki, Macedonia, Greece). Geobios 2008, 41, 365–380. [Google Scholar] [CrossRef]
- Syrides, G.E. Lithostratigraphic, Biostratigraphic and Palaeogeographic Study of the Neogene-Quaternary Sedimentary Deposits of Chalkidiki Peninsula, Macedonia, Greece. Ph.D. Thesis, Aristotle University of Thessaloniki, Scientific Annals of the School of Geology, Thessaloniki, Greece, 1990. Volume 11. pp. 1–243. [Google Scholar]
- Tsoukala, E.; Bartsiokas, A. New Mesopithecus pentelicus specimens from Kryopigi, Macedonia, Greece. J. Hum. Evol. 2008, 54, 448–451. [Google Scholar] [CrossRef]
- Lazaridis, G. Contribution to the Study of the Neogene Perissodactyls from Kryopigi (Kassandra, Chalkidiki, Greece). Master’s Thesis, Aristotle University of Thessaloniki, Thessaloniki, Greece, 2010. [Google Scholar]
- Lazaridis, G.; Tsoukala, E. Tetralophodon longirostris (Kaup, 1832) from Late Miocene of the Kassandra peninsula (Chalkidiki, Greece). In Proceedings of the Scientific Annals, School of Geology, Aristotle University of Thessaloniki, Greece VIth International Conference on Mammoths and their Relatives, Grevena–Siatista, Thessaloniki, Greece, 5–12 May 2014; Volume 102, p. 101. [Google Scholar]
- Tsoukala, E.; Melentis, J.K. Deinotherium giganteum Kaup (Proboscidea) from Kassandra Peninsula (Chalkidiki, Macedonia, Greece). Geobios 1994, 27, 633–640. [Google Scholar] [CrossRef]
- Lazaridis, G.; Kostopoulos, D.S.; Lyras, G.A.; Roussiakis, S. A new Late Miocene ovibovine-like bovid (Bovidae, Mammalia) from the Kassandra Peninsula (Chalkidiki, Northern Greece) and implications to the phylogeography of the group. Paläontologische Z. 2017, 91, 427–437. [Google Scholar] [CrossRef]
- Ríos, M.; Cantero, E.; Estraviz-López, D.; Solounias, N.; Morales, J. Anterior ossicone variability in Decennatherium rex Ríos; et al. 2017 (Late Miocene, Iberian Peninsula). Earth Environ. Sci. Trans. R. Soc. Edinb. 2023, 114, 159–165. [Google Scholar] [CrossRef]
- Maddison, W.P.; Maddison, D.R. Mesquite: A Modular System for Evolutionary Analysis (Version 3.61). 2019. Available online: http://www.mesquiteproject.org (accessed on 27 December 2019).
- Goloboff, P.A.; Farris, J.S.; Nixon, K.C. TNT, a free program for phylogenetic analysis. Cladistics 2008, 24, 774–786. [Google Scholar] [CrossRef]
- Hou, S.; Danowitz, M.; Sammis, J.; Solounias, N. Dead ossicones, and other characters describing Palaeotraginae (Giraffidae; Mammalia) based on new material from Gansu, Central China. Zitteliana 2014, B32, 91–98. [Google Scholar]
- Laskos, K.; Kostopoulos, D.S. On the last European giraffe, Palaeotragus inexspectatus (Mammalia: Giraffidae); new remains from the Early Pleistocene of Greece and a review of the species. Zool. J. Linn. Soc. 2025, 203, zlae056. [Google Scholar] [CrossRef]
- Solounias, N. Family Giraffidae. In The Evolution of Artiodactyls; Prothero, D.R., Foss, S.E., Eds.; The Johns Hopkins University Press: Baltimore, MD, USA, 2007; pp. 257–277. [Google Scholar]
- Parizad, E.; Ataabadi, M.M.; Mashkour, M.; Kostopoulos, D.S. Samotherium Major, 1888 (Giraffidae) skulls from the late Miocene Maragheh fauna (Iran) and the validity of Alcicephalus Rodler & Weithofer, 1890. Comptes Rendus Paleovol 2019, 19, 153–172. [Google Scholar]
- Kargopoulos, N.; Marugán-Lobón, J.; Chinsamy, A.; Agwanda, B.R.; Brown, M.B.; Fennessy, S.; Ferguson, S.; Hoffman, R.; Lala, F.; Muneza, A.; et al. Heads up–Four Giraffa species have distinct cranial morphology. PLoS ONE 2024, 19, e0315043. [Google Scholar] [CrossRef] [PubMed]
- Kargopoulos, N.; Marugán-Lobón, J.; Chinsamy, A.; Brown, M.B.; Fennessy, S.; Ferguson, S.; Petzold, A.; Winter, S.; Zabeirou, A.R.M.; Fennessy, J. A Reassessment of the Cranial Diversity of the West African Giraffe. Int. J. Zool. 2025, 2025, 8816347. [Google Scholar] [CrossRef]
- Singer, R.; Boné, E. Modern giraffes and the fossil giraffids of Africa. Ann. S. Afr. Mus. 1960, 45, 375–548. [Google Scholar]
- Lydekker, B.A. Indian Tertiaty and post-Tertiary Vertebrata. Mem. Geol. Surv. India 1876, 1, 19–83. [Google Scholar]
- Lydekker, R. Notices of Siwalik Mammals. Rec. Geol. Surv. India 1878, 11, 64–104. [Google Scholar]
- Pilgrim, G.E. Notices of new Mammalian genera and species from the tertiaries of India–Calcutta. Rec. Geol. Surv. India 1910, 40, 63–71. [Google Scholar]
- Pilgrim, G.E. The Fossil Giraffidae of India. Mem. Geol. Surv. India 1911, 4, 1–29. [Google Scholar]
- Bhatti, Z.H.; Khan, M.A.; Akthar, M.; Khsan, A.M.; Ghaffar, A.; Iqbal, M.; Ikram, T. Giraffokeryx (Artiodactyla: Mammalia) remains from the Lower Siwaliks of Pakistan. Pak. J. Zool. 2013, 44, 1623–1631. [Google Scholar]
- Harris, J.M. Orientation and variability in the ossicones of African Sivatheriinae (Mammalia: Giraffidae). Ann. S. Afr. Mus. 1974, 65, 189–198. [Google Scholar]
- Ríos, M.; Morales, J. A new skull of Decennatherium rex Ríos, Sánchez and Morales, 2017 from Batallones-4 (upper Vallesian, MN10, Madrid, Spain). Palaeontol. Electrónica 2019, 22, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Mahmood, K.; Khan, M.A.; Babar, M.A.; Akhtar, M. Siwalik Giraffidae (Mammalia, Artiodactyla): A review. Punjab Univ. J. Zool. 2015, 30, 31–36. [Google Scholar]
- Draz, O.; Ni, X.; Samiullah, K.; Yasin, R.; Fazal, R.M.; Naz, S.; Akhtar, S.; Gillani, M.; Ejaz, M. New Fossil Remains of Artiodactyla from Dhok Pathan Formation, Middle Siwaliks of Punjab, Pakistan. Pak. J. Zool. 2020, 52, 1631–2026. [Google Scholar] [CrossRef]
- Patnaik, R. Diet and habitat changes among Siwalik herbivorous mammals in response to Neogene and Quaternary climate changes: An appraisal in the light of new data. Quat. Int. 2015, 371, 232–243. [Google Scholar] [CrossRef]
- Merceron, G.; Colyn, M.; Geraads, D. Browsing and non-browsing extant and extinct giraffids: Evidence from dental microwear textural analysis. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2018, 505, 128–139. [Google Scholar] [CrossRef]
- Yohe, L.R.; Solounias, N. The five digits of the giraffe metatarsal. Biol. J. Linn. Soc. 2020, 131, 699–705. [Google Scholar] [CrossRef]
- Geraads, D.; Spassov, N.; Hristova, L.; Markov, G.N.; Tzankov, T. Upper Miocene mammals from Strumyani, South-Western Bulgaria. Geodiversitas 2011, 33, 451–484. [Google Scholar] [CrossRef]
- Solounias, N.; Danowitz, M. Astragalar morphology of selected Giraffidae. PLoS ONE 2016, 11, e0151310. [Google Scholar] [CrossRef]
- Ríos, M.; Abbas, S.G.; Khan, M.A.; Solounias, N. A new giraffid Bramiscus micros nov. gen. nov. sp. (Ruminantia, Giraffidae) from the Miocene of northern Pakistan. Palaeontol. Electron. 2024, 27, a29. [Google Scholar] [CrossRef]
- Wimberly, A.N. Predicting body mass in Ruminantia using postcranial measurements. J. Morphol. 2023, 284, e21636. [Google Scholar] [CrossRef] [PubMed]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).