
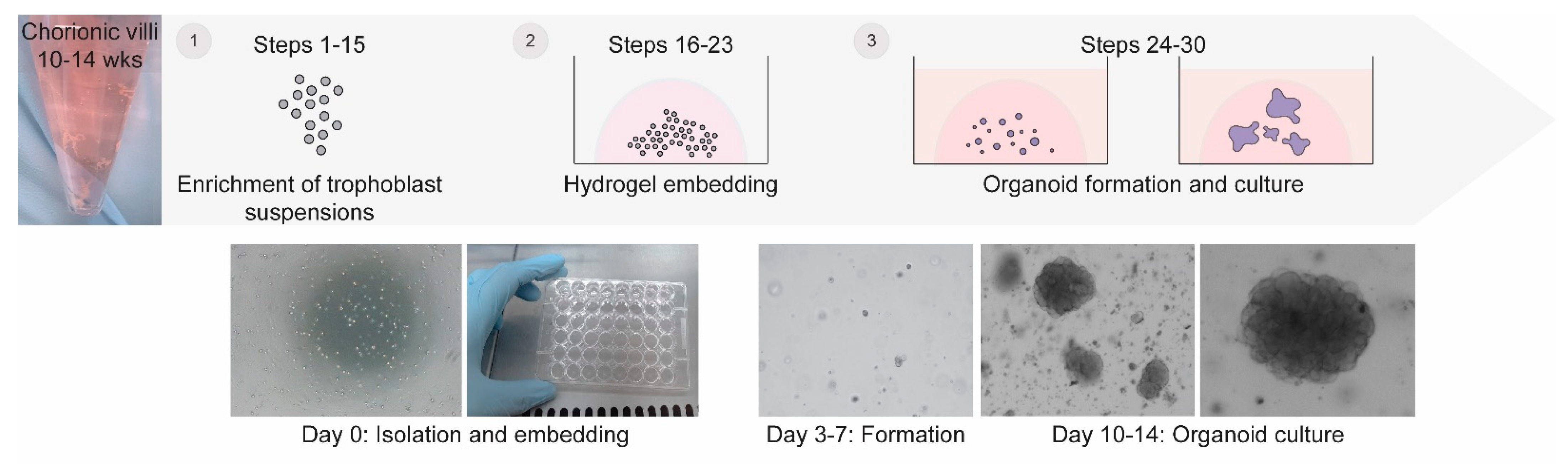
Generation of Trophoblast Organoids from Chorionic Villus Sampling



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Experimental Design
2.1. Starting Material
2.2. Dissociation
2.3. Culture
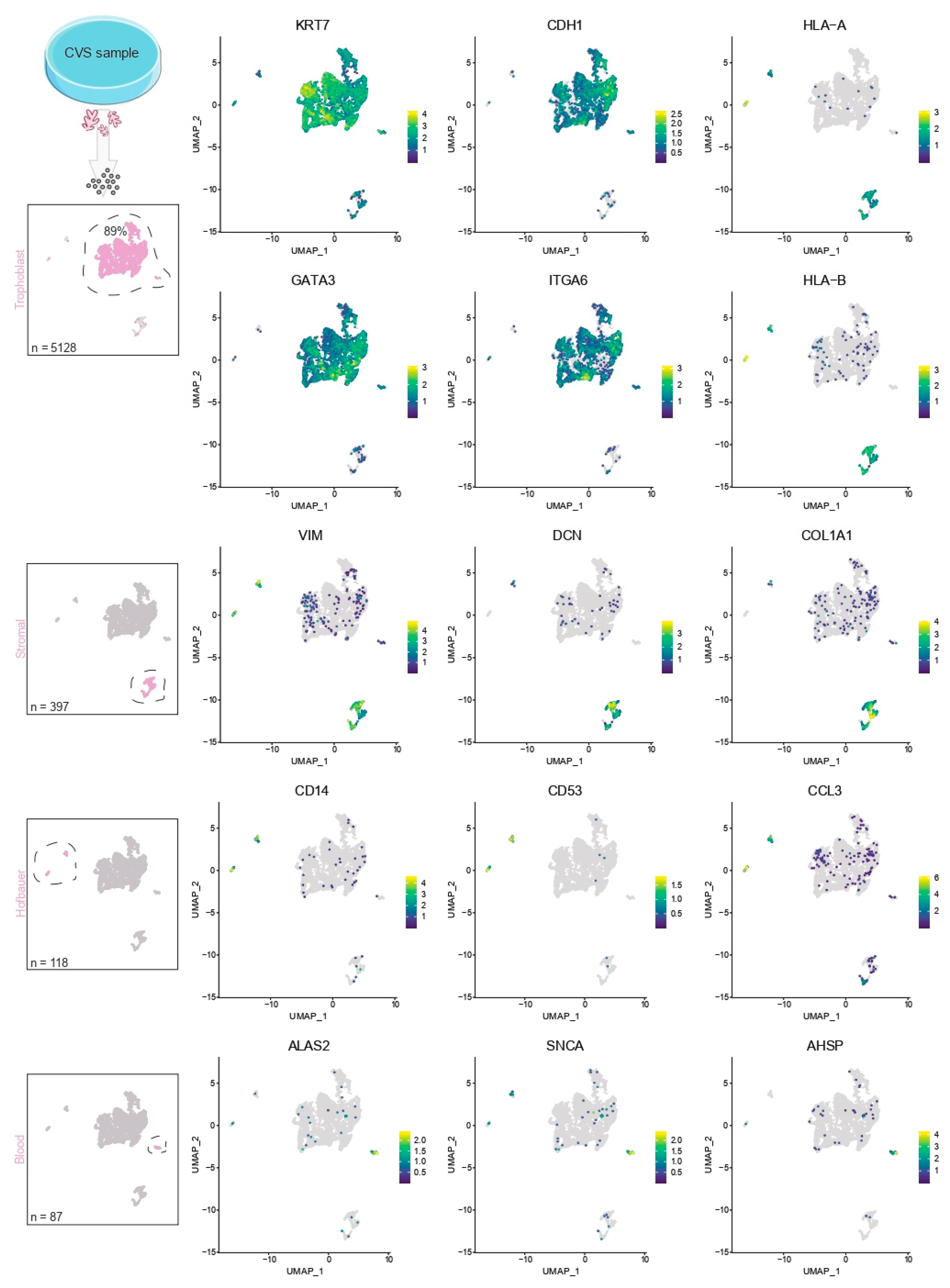
2.4. Contamination of Decidual Cells
2.5. Validation
2.6. EVT Differentiation
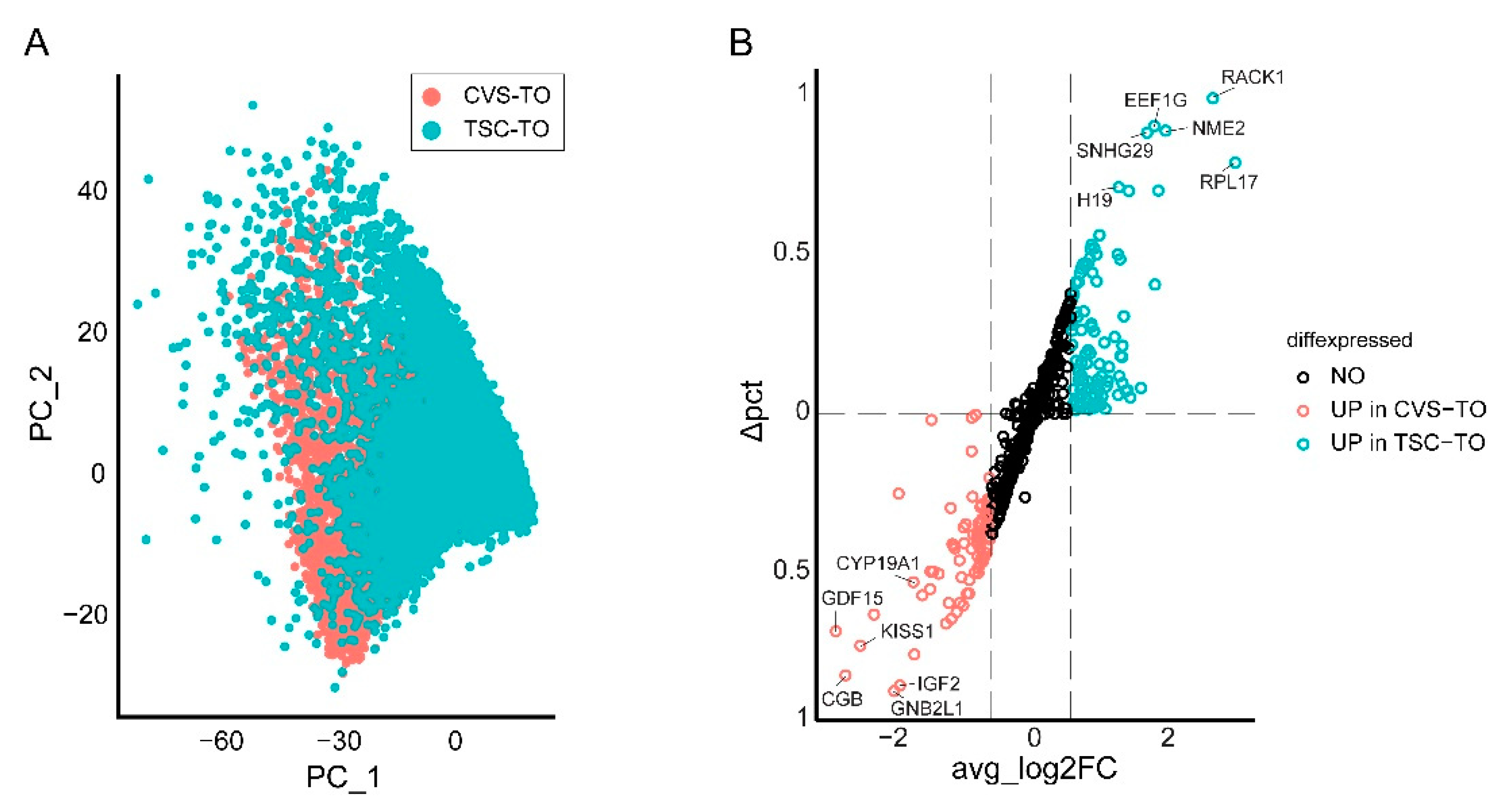
2.7. Comparison to Stem Cell-Derived Trophoblast Organoids
3. Materials
3.1. Biological Materials
- -
- Samples of chorionic villi from 10–14 weeks gestation obtained by CVS following standard procedures. Samples can be collected in PBS or culture medium (DMEM/RPMI 1640) without additives. Tissue should be recognized as a potential risk for bloodborne pathogens and should be handled with appropriate personal protective equipment. Samples obtained by transvaginal approaches have a higher likelihood to contain yeast infections. CVS tissues that are a potential hazard for emerging viruses should only be used according to specific biosafety guidelines (BSL-3/4) and require specialized equipment.
3.2. Reagents
- -
- DMEM (Gibco, Waltham, MA, USA; cat.no. 31966021)
- -
- Fetal bovine serum (Capricorn Scientific, Ebsdorfergrund, Germany; cat.no. FBS-12A)
- -
- Penicillin-Streptomycin (Sigma-Aldrich, St. Louis, MO, USA; cat.no. P0781)
- -
- Trypsin-EDTA (Sigma-Aldrich; cat.no. T3924)
- -
- DPBS (Sigma-Aldrich; cat.no. D8537)
- -
- Bovine serum albumin (Sigma-Aldrich; cat.no. A3294)
- -
- ACK lysing buffer (Gibco; cat.no. A1049201)
- -
- Cultrex UltiMatrix RGF BME (R&D Systems, Minneapolis, MN, USA; cat.no. BME00110)
- -
- Advanced DMEM/F12 (Gibco; cat.no. 12634028)
- -
- Knockout serum replacement (Gibco; cat.no. 10828010)
- -
- GlutaMAX (Gibco; cat.no. 35050038)
- -
- N-Acetyl-L-cysteine (Sigma-Aldrich; cat.no. A9165)
- -
- Nicotinamide (Sigma-Aldrich; cat.no. N0636)
- -
- N-2 supplement (Gibco; cat.no. 17502048)
- -
- B-27 supplement, minus vitamin A (Gibco; cat.no. 12587010)
- -
- Primocin (Invivogen, San Diego, CA, USA; cat.no. ant-pm-1)
- -
- Recombinant human EGF (Peprotech, London, UK; cat.no. AF-100-15)
- -
- GSK inhibitor CHIR99021 (Tocris, Bristol, UK; cat.no. 4423)
- -
- Recombinant human R-spondin-1 (Peprotech; cat.no. 120-38)
- -
- Recombinant human HGF (Peprotech; cat.no. 100-39)
- -
- Recombinant human FGF (Peprotech; cat.no. 100-18B)
- -
- Rock inhibitor Y-27632 (Millipore, Burlington, MA, USA; cat.no. 688000)
- -
- Prostaglandin E2 (Sigma-Aldrich; cat.no. P0409)
- -
- A83-01 (Tocris; cat.no. 2939)
- -
- Cultrex organoid harvesting solution (R&D Systems; cat.no. 3700-100-01)
- -
- Cellbanker 2 cryopreservation medium (Amsbio, Abingdon, UK; cat.no. 11914)
- -
- Paraformaldehyde (Sigma-Aldrich; cat.no. 158127)
- -
- Sucrose (Sigma-Aldrich; cat.no. S9378)
- -
- Anti-CDH1 antibody (Cell Signaling Technology, Danvers, MA, USA; cat.no. 3195, dilution 1:200)
- -
- Anti-KRT7 antibody (Agilent Technologies, Santa Clara, CA, USA; cat.no. M7018, dilution 1:100)
- -
- Anti-GDF15 antibody (Sigma-Aldrich; cat.no. HPA011191, dilution 1:150)
- -
- Anti-HLA-G antibody (Exbio, Vestec, Czechia; cat.no. 11-431-C100, dilution 1:100)
- -
- Alexa Fluor 488 antibody (Invitrogen, Waltham, MA, USA; cat.no. A32723, dilution 1:500)
- -
- Alexa Fluor 647 antibody (Invitrogen; cat.no. A32733, dilution 1:500)
3.3. Equipment
- -
- Microbiological class II safety cabinet (Scala Scientific, Ede, The Netherlands; cat.no. BIO 130 A2)
- -
- Incubator (PHCbi, Etten-Leur, The Netherlands; cat.no. MCO-170 AIC-UVD PE)
- -
- Centrifuge (ThermoFisher Scientific, Waltham, MA, USA; cat.no. 75004210)
- -
- Benchtop centrifuge (ThermoFisher Scientific; cat.no. 75002421)
- -
- Digital microscope (ThermoFisher Scientifc; cat.no. AMF5000)
- -
- Nitrile gloves (Ansell, Richmond, Australia; cat.no. 6034153)
- -
- Serological pipette controller (DLAB Scientific Co., Beijing, China; cat.no. 861051B)
- -
- Serological pipettes (Greiner Bio-One, Kremsmünster, Austria; cat.no. 607180)
- -
- Single-channel pipettes (Mettler Toledo, Hong Kong, China; cat.no. 17014391/17014382)
- -
- Pipette tips (Mettler Toledo; cat.no. 30389276/30389272)
- -
- Cell culture 48 well plate (Greiner Bio-One; cat.no. 677180)
- -
- Conical 15mL tubes (Greiner Bio-One; cat.no. 188271-N)
- -
- Microcentrifuge 1.5 mL tubes (Greiner Bio-One; cat.no. 616201)
- -
- Cell culture cryogenic tubes (ThermoFisher Scientific; cat.no. 366656)
- -
- Cell strainer 30 µm (Miltenyi Biotec, Bergisch Gladbach, Germany; cat.no. 130-098-458)
3.4. Reagent Setup
4. Method
4.1. Preparation
- -
- Thaw the desired number of Cultrex UltiMatrix RGF BME aliquots on ice for at least 1 h. Keep on ice at all times.
- -
- Thaw and/or warm Trypsin/EDTA.
- -
- Prepare and/or warm washing medium.
- -
- Prepare TOM.
- -
- Prepare coating solution.
- -
- Start cooling the benchtop centrifuge to 4 °C.
- -
- Warm a 48-wells plate in the incubator (37 °C and 5% CO2).
4.2. Enzymatic Digestion and Preparation of Cell Suspension
- Collect surplus CVS tissue.
- Villi can be stored for up to 2 or 3 nights in washing medium in the incubator (37 °C and 5% CO2).
- Prewash pipettes with coating solution to prevent sticking of CVS tissue.
- Transfer all CVS tissue to a 15 mL conical tube.
- Let the villi settle down or centrifuge at 300× g for 5 min at room temperature.
- Repeat until you have collected all the tissue.
- Aspirate supernatant.
- Incubate villi in 2 mL Trypsin-EDTA for 30 min in the incubator (37 °C and 5% CO2) or water bath (37 °C). Longer incubation will result in lower cell viability.
- Prewash a 30 µm cell strainer with 5 mL washing medium (collect in 15 mL tube).
- Prewash pipettes with coating solution to prevent sticking of tissue.
- Pipette villi up and down (~50/100 times) with a coated P1000 pipette and apply to cell strainer.
- Repeat until all tissue is processed.
- Wash the cell strainer with 5 mL washing medium.
- Centrifuge at 300× g for 5 min at room temperature.
- If the pellet contains large amounts of blood, follow steps 12a–c. Otherwise, proceed with step 13.
- Resuspend pellet in 1–2 mL ACK lysing buffer (1:5 ratio of cell pellet: ACK lysing buffer).
- Incubate for 5–10 min at room temperature while gently flicking the tube every 2 min. Duration of the incubation depends on the severity of red blood cell contamination; lysis should be visually assessed.
- Centrifuge at 300× g for 5 min at room temperature.
- Aspirate supernatant.
- Resuspend the pellet in 1 mL ice-cold Advanced DMEM/F12 and transfer to a 1.5 mL microcentrifuge tube.
- Count the cells.
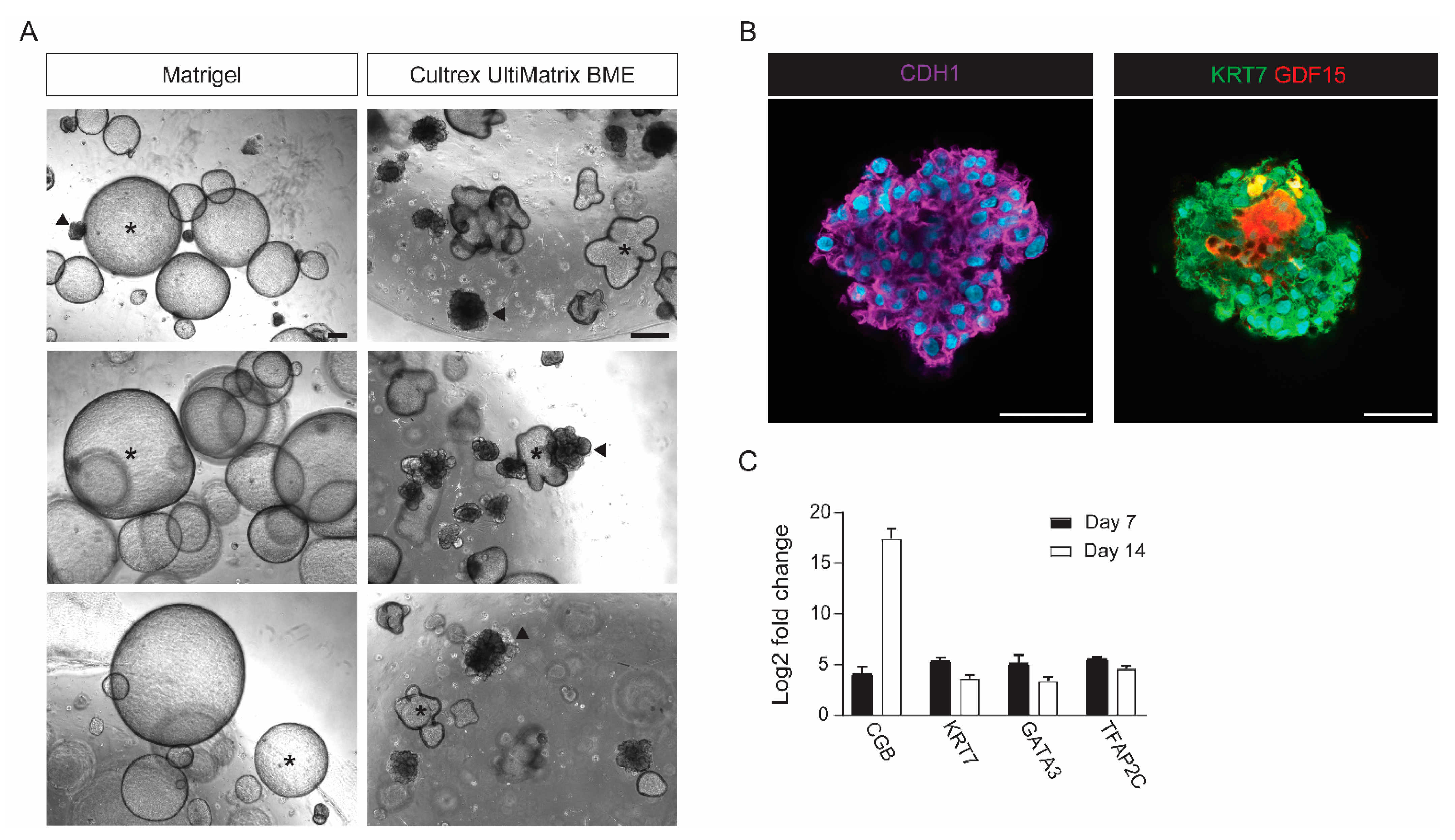
4.3. Initiation of Trophoblast Organoid Cultures
- 16.
- Centrifuge the isolated cells at 300× g for 5 min at 4 °C.
- 17.
- Carefully aspirate as much medium as possible without disturbing the pellet and gently flick the tube to loosen the pellet.
- 18.
- Put cells on ice.
- 19.
- Add Cultrex UltiMatrix RGF BME to establish a concentration of 1 × 104–1 × 105 cells/25 µL of hydrogel and carefully mix by pipetting. Prevent the formation of air bubbles.
- 20.
- Put cells on ice.
- 21.
- Pipette small hydrogel domes (20–25 µL) in the middle of every well. Prevent the formation of air bubbles.
- 22.
- Let the hydrogel domes settle for 1–2 min at room temperature and carefully transfer the plate to the incubator (37 °C and 5% CO2).
- 23.
- After 2 min, invert the plate upside down in one movement and incubate for 10–13 min. Inverting the plate facilitates spreading of the cells within the hydrogel domes. In the meantime, warm TOM medium.
- 24.
- Add 250 µL pre-warmed TOM medium to each well.
4.4. Maintenance of Trophoblast Organoids Cultures
- 25.
- Refresh TOM every 2–4 days. Organoid structures should appear after 5 to 7 days.
- 26.
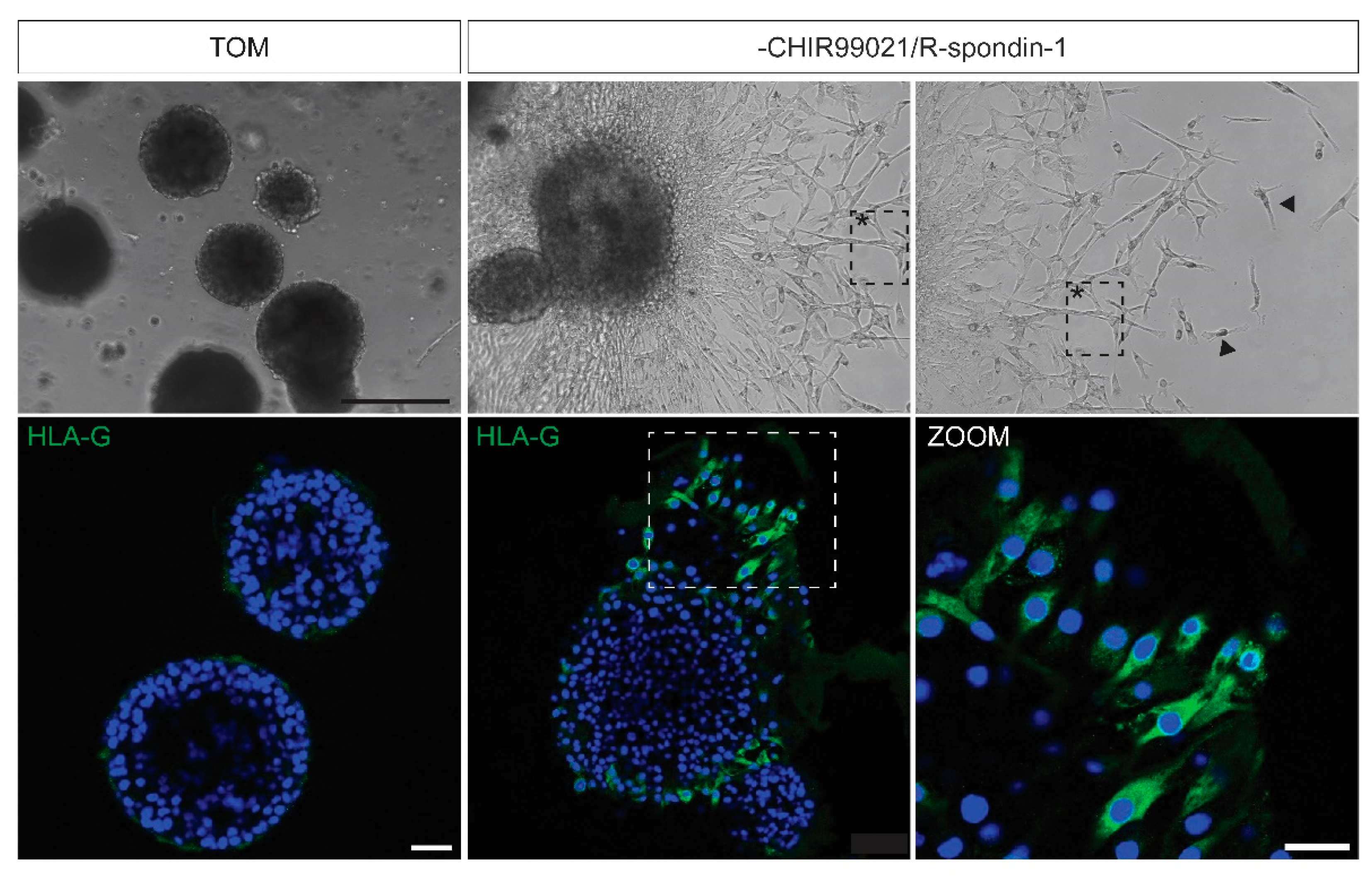
- Induction of EVT differentiation:
- Culture trophoblast organoids in TOM for 7 to 10 days (small structures should be developed).
- Aspirate TOM.
- Replace TOM for TOM without CHIR99021 and R-spondin-1 (TOM-EVT). Refresh TOM-EVT every 2–4 days.
- Outgrowth of EVT-like cells can be observed after 3–5 days. Invasive cells can be observed after 7–10 days.
- 27.
- Passaging: In order to establish long-term cultures, trophoblast organoids need to be passaged every 5 to 10 days depending on their size, density, and growth rate. Organoid cultures should be passaged when at least half of the structures reach a diameter of 200 µm.
- Break down the hydrogel domes by scraping using a pipette tip.
- Transfer the content (TOM with broken hydrogel) of the well to a 1.5 mL microcentrifuge tube. Collect up to 4 wells per tube. Prewash pipettes with coating solution to prevent sticking of organoids.
- Wash every well with 250 µL of ice-cold Advanced DMEM/F12 and add to the tube. Prewash pipettes with coating solution to prevent sticking of organoids.
- Centrifuge at 300× g for 5 min at room temperature.
- Aspirate supernatant and add 200 µL of ice-cold Advanced DMEM/F12.
- Mechanically dissociate the organoids by extensive pipetting (200–500 times) using small pipette tips until organoids are broken into hardly visible particles. Prewash pipettes with coating solution to prevent sticking of organoids.
- Add 1 mL Advanced DMEM/F12 and centrifuge at 300× g for 5 min at room temperature.
- Proceed with step 16. Residual hydrogel should be taken into account when adding new Cultrex UltiMatrix RGF BME. Organoid cultures should be passaged at a 1:2 ratio. For cryopreservation, proceed with Step 29.
- 28.
- Harvesting:
- Aspirate TOM.
- Dissolve the organoid-containing drops in 500 µL ice-cold Cell Recovery solution for 30 min at 4 °C.
- Break down any residual hydrogel domes by scraping using a pipette tip.
- Collect the content of the well in a 1.5 mL microcentrifuge tube. Prewash pipettes with coating solution to prevent sticking of organoids.
- Wash every well with 500 µL of ice-cold Advanced DMEM/F12 and add to the tube. Prewash pipettes with coating solution to prevent sticking of organoids.
- 29.
- Cryopreservation:
- Perform Step 27: Passaging.
- Aspirate supernatant.
- Resuspend in 1 mL of Cellbanker-2 cryopreservation medium and transfer to labeled cryovials.
- Store cryovials at 80 °C for 24–48 h.
- Transfer cryovials to liquid nitrogen storage. Organoid cultures can be stored for at least 3 years.
- 30.
- Thawing:
- Transfer cryovial to a water bath (37 °C) and thaw as quickly as possible.
- Add 10 mL of Advanced DMEM/F12 and transfer to a 15 mL conical tube.
- Centrifuge at 300× g for 5 min at room temperature.
- Aspirate supernatant and proceed with Step 15.
4.5. Single-Cell Isolation
- 31.
- Pool organoids for dissociation into single cells.
- 32.
- Perform Step 28: Harvesting.
- 33.
- Centrifuge at 300× g for 5 min at room temperature.
- 34.
- Aspirate supernatant and resuspend in 1 mL Trypsin-EDTA. Prewash pipettes with coating solution to prevent sticking of CVS tissue.
- 35.
- Incubate for 15 min at 37 °C.
- 36.
- Mechanically dissociate the organoids by extensive pipetting (200–500 times) using small pipette tips.
- 37.
- Prewash a 30 µm cell strainer with 1 mL Advanced DMEM/F12 (collect in 15 mL tube).
- 38.
- Filter the cell suspension of the cell strainer and wash the cell strainer with 4 mL of Advanced DMEM/F12.
- 39.
- Centrifuge at 300× g for 5 min at room temperature.
- 40.
- Count cells and check viability. If proceeding with single-cell RNA sequencing using the 10× Chromium platform, resuspend cells in Advanced DMEM/F12 to a final concentration of 700–1000 cells/µL. We pooled roughly 50 Day-14 organoids to obtain 10,000–15,000 high-quality cells for downstream analysis.
5. Limitations
6. Summary
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Turco, M.Y.; Gardner, L.; Kay, R.G.; Hamilton, R.S.; Prater, M.; Hollinshead, M.S.; McWhinnie, A.; Esposito, L.; Fernando, R.; Skelton, H.; et al. Trophoblast organoids as a model for maternal-fetal interactions during human placentation. Nature 2018, 564, 263–267. [Google Scholar] [CrossRef]
- Haider, S.; Meinhardt, G.; Saleh, L.; Kunihs, V.; Gamperl, M.; Kaindl, U.; Ellinger, A.; Burkard, T.R.; Fiala, C.; Pollheimer, J.; et al. Self-Renewing Trophoblast Organoids Recapitulate the Developmental Program of the Early Human Placenta. Stem Cell Rep. 2018, 11, 537–551. [Google Scholar] [CrossRef]
- Karvas, R.M.; Khan, S.A.; Verma, S.; Yin, Y.; Kulkarni, D.; Dong, C.; Park, K.M.; Chew, B.; Sane, E.; Fischer, L.A.; et al. Stem-cell-derived trophoblast organoids model human placental development and susceptibility to emerging pathogens. Cell Stem Cell 2022, 29, 810–825.e8. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Semmes, E.C.; Ovies, C.; Megli, C.; Permar, S.; Gilner, J.B.; Coyne, C.B. Innate immune signaling in trophoblast and decidua organoids defines differential antiviral defenses at the maternal-fetal interface. Elife 2022, 11, e79794. [Google Scholar] [CrossRef] [PubMed]
- Schäffers, O.J.M.; Dupont, C.; Bindels, E.M.; Van Opstal, D.; Dekkers, D.H.W.; Demmers, J.A.A.; Gribnau, J.; van Rijn, B.B. Single-Cell Atlas of Patient-Derived Trophoblast Organoids in Ongoing Pregnancies. Organoids 2022, 1, 106–115. [Google Scholar] [CrossRef]
- Hao, Y.; Stuart, T.; Kowalski, M.H.; Choudhary, S.; Hoffman, P.; Hartman, A.; Srivastava, A.; Molla, G.; Madad, S.; Fernandez-Granda, C.; et al. Dictionary learning for integrative, multimodal and scalable single-cell analysis. Nat. Biotechnol. 2023, 42, 293–304. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Liang, P.; Yang, H.; Coyne, C.B. Trophoblast organoids with physiological polarity model placental structure and function. J. Cell Sci. 2023, 137, jcs261528. [Google Scholar] [CrossRef] [PubMed]
- Co, J.Y.; Margalef-Català, M.; Monack, D.M.; Amieva, M.R. Controlling the polarity of human gastrointestinal organoids to investigate epithelial biology and infectious diseases. Nat. Protoc. 2021, 16, 5171–5192. [Google Scholar] [CrossRef] [PubMed]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
van Rijn, B.; Van Opstal, D.; van Koetsveld, N.; Knapen, M.; Gribnau, J.; Schäffers, O. Generation of Trophoblast Organoids from Chorionic Villus Sampling. Organoids 2024, 3, 54-66. https://doi.org/10.3390/organoids3010005
van Rijn B, Van Opstal D, van Koetsveld N, Knapen M, Gribnau J, Schäffers O. Generation of Trophoblast Organoids from Chorionic Villus Sampling. Organoids. 2024; 3(1):54-66. https://doi.org/10.3390/organoids3010005
Chicago/Turabian Stylevan Rijn, Bas, Diane Van Opstal, Nicole van Koetsveld, Maarten Knapen, Joost Gribnau, and Olivier Schäffers. 2024. "Generation of Trophoblast Organoids from Chorionic Villus Sampling" Organoids 3, no. 1: 54-66. https://doi.org/10.3390/organoids3010005
APA Stylevan Rijn, B., Van Opstal, D., van Koetsveld, N., Knapen, M., Gribnau, J., & Schäffers, O. (2024). Generation of Trophoblast Organoids from Chorionic Villus Sampling. Organoids, 3(1), 54-66. https://doi.org/10.3390/organoids3010005