Abstract
This study aimed to develop seed-based mass propagation techniques for the conservation and horticultural and medicinal uses of Scrophularia koraiensis Nakai, an endemic plant in the Korean Peninsula. Seeds were collected from four different locations (accessions) and subjected to untreated (control), gibberellic acid (GA)-only treatment, cold stratification-only treatment, or a cold stratification + GA treatment. Except for seeds collected from one location, the control group exhibited low germination of below 20% in all other locations. However, the GA-only and cold stratification-only treatments released seed dormancy and promoted germination compared with the control. In particular, the cold stratification-only treatment at 5 °C for 4 weeks resulted in about 80% germination in all accessions and demonstrated positive effects on germination speed and uniformity. These findings indicated that S. koraiensis seeds exhibit non-deep physiological dormancy (PD). Upon comparing the seed dormancy classes across various species of Scrophularia, native to different continents and countries, we confirmed that PD is very well conserved in the genus Scrophularia. Therefore, the study outcomes will provide fundamental and practical insights into the seed dormancy and germination characteristics of various Scrophularia species for future studies.
1. Introduction
Scrophularia L., belonging to the family Scrophulariaceae and tribe Scrophularia, comprises about 270 taxa. They are predominantly distributed in the temperate regions of the Northern Hemisphere, with a few species found in the Southern Hemisphere [1,2,3,4]. In Eastern cultures such as China and Korea, the roots of Scrophularia plants have been used as anti-inflammatory, antipyretic, and detoxifying herbal medicines [5,6]. In Korea, S. buergeriana Miq. is considered the original plant of Scrophularia radix (Hyun-Sam), a herbal medicine also known as Korean Hyun-Sam [7]; additionally, S. kakudensis Franch., S. koraiensis Nakai, and S. takesimensis Nakai are used as alternatives to this herbal medicine [8,9].
Reports suggest that plants belonging to the genus Scrophularia L., which is native to the Korean Peninsula, are classified into six species and one variety (seven taxa). This classification is based on external morphology such as flowering time; growth traits of inflorescence, peduncle, floret, calyx, stem, and leaf; characteristics of root and seed; and internal transcribed spacer (ITS) DNA sequence data [4,10,11]. Among them, three species, including S. koraiensis Nakai, S. cephalantha Nakai, and S. takesimensis Nakai, were reported to be endemic only to the Korean Peninsula in East Asia [12,13].
S. koraiensis is a perennial herb growing in the semi-shade of mountainous areas, reaching a height of approximately 1.5 m, and flowering in the month of July. Glandular trichomes develop throughout the plant during the early growth period but nearly disappear by the flowering period [4]. Its fruit is an ovoid capsule that matures from August to September. Recent studies have shown the preventive effects of S. koraiensis on bone disorders, particularly through inhibiting osteoclast differentiation and bone resorption [9]; additionally, its extracts could be used as a potential therapeutic agent for allergic airway inflammation [14]. As a member of the family Scrophulariaceae, S. koraiensisis has been designated as a “biological resource subject to approval for overseas export” in Korea. This designation reflects the high protection value of these plants for biodiversity conservation, thereby requiring approval from the Minister of Environment in accordance with the provisions of the Ministry of Environment Ordinance while exporting them [15]. Additionally, S. koraiensisis has been identified as a target plant to monitor mid- to long-term adaptability of plant resources to climate change on the Korean Peninsula [16]. Therefore, developing mass propagation technologies is essential for the sustainable conservation and utilization of this plant; however, research in this area remains limited.
Seed-based mass propagation of plants is essential for preserving genetic diversity and promoting its sustainable use. Studies have reported characteristics of seed germination for several Scrophularia plants. When seeds were treated with 400 ppm GA3 for 24 h, the seeds of S. striata, a naturally growing plant in the western parts of Iran, showed the best germination [17]. When seeds were soaked in 200 mgL−1 GA3 for 24 h, the seeds of S. takesimensis, a plant native to the Korean Peninsula, exhibited approximately 80% germination [18]. The seeds of S. marilandica, a plant native to Canada, exhibited nearly complete germination following treatment with 3 × 10−4 M GA3 [19]. However, there are no reports on the seed propagation of S. koraiensis. We hypothesized that GA treatment would break dormancy in S. koraiensis seeds, as reported for other Scrophularia species. Given the known interaction between GA and cold stratification [17,19], additional experiments were conducted to examine this relationship. Therefore, this study aimed to improve the seed propagation efforts by determining whether seed dormancy is present in S. koraiensisis seeds and, if so, identifying its type. In particular, (1) the water imbibition of seeds; (2) the morphology of seeds and embryo; and (3) the effects of cold stratification, gibberellic acid (GA) treatment, and seed collection sites were investigated.
2. Materials and Methods
2.1. Seed Collection
The seeds investigated in this study were collected between 2 and 4 September 2014 from S. koraiensis Nakai growing on Mt. Gariwang (Kari, Figure 1), Balwang (Balw), Gyebang (Gyeb), and Deokhang (Deok) in the Gangwon Province, Republic of Korea. Fully mature fruits were collected and brought to the laboratory, and only fresh seeds were selected for experiments. The fresh seeds were dried for 2 weeks and then refrigerated at 5 °C for 6 months (WIM-RL4, Daihan Scientific Co., Wonju, Republic of Korea) until further use.

Figure 1.
Scrophularia koraiensis inhabiting Mt. Kariwang, Gangwon Province, Republic of Korea.
2.2. Seed Water Absorption
The degree of moisture absorption of seeds was determined by placing the seeds in Petri dishes (90 × 15 mm), lined with two sheets of filter paper (Whatman No. 1, GE Healthcare, Buckinghamshire, UK) and containing at least 10 mL distilled water. Thereafter, the weight of the seeds was measured at room temperature (22–25 °C) at predetermined time intervals of 0, 2, 4, 8, 12, 24, and 48 h. The moisture on the surface of the seeds was wiped off with a tissue, and the weight of 40 seeds was measured at each time point in triplicate. The increase in the mass of the seeds was calculated using the following formula:
%Ws = [(Wh − Wi)/Wi] × 100
Here, Ws denotes the increase in the mass of the seeds; Wh refers to the mass of seeds measured after imbibition at a specific time point; and Wi indicates the initial mass of the seeds.
2.3. Seed Germination
The seeds were collected from four different habitats, stored for 6 months at 5 °C and tested in four treatment groups: (1) no treatment (control), (2) GA-only treatment, (3) cold stratification-only treatment, and (4) cold stratification + GA treatment. For the GA-only treatment, seeds were soaked in a solution containing 800 mg∙L−1 GA3 for 24 h at room temperature (22–25 °C) prior to sowing. For the cold stratification-only treatment, seeds were placed in Petri dishes (90 × 15 mm) lined with two sheets of filter paper (Whatman No. 1, GE Healthcare, Buckinghamshire, UK) and containing 5 mL distilled water. To prevent moisture evaporation from the Petri dishes, the Petri dishes was sealed with parafilm, and additional distilled water was supplied when moisture was insufficient. The seeds were then incubated in a cold lab chamber (WIM-RL4, Daihan Scientific Co., Wonju, Republic of Korea) at 5 °C for 4 weeks of cold stratification before sowing. For the cold stratification + GA treatment, the seeds were first cold stratified using the same method and then soaked in the GA solution for 24 h prior to sowing. For the control group, untreated seeds were sown.
Treated seeds of the three groups were sown, one seed per hole, in 128-hole plug trays (27.5 × 54.0 × 5.0 cm), which was filled with horticultural soil (TKS-2, Floragard, Oldenburg, Germany). These plug trays were placed in an experimental greenhouse at the Useful Plant Resources Center of the Korea National Arboretum in Yangpyeong-gun, Gyeonggi-do, Republic of Korea. The average temperature in the greenhouse was 18 ± 3.5 °C, and the natural day length was maintained without any artificial control. All experimental treatments were performed with 40 seeds in triplicate.
2.4. Data Collection and Statistical Analysis
Following each experimental treatment, germinated seeds were counted daily for 4 weeks, and germination percentage (GP), days to 50% of germination of final GPs (T50), mean germination time (MGT), germination uniformity (GU), and germination speed (GS) were calculated using the following equations [20,21,22]. Germination was considered to have occurred when the radicle protruded through the seed coat and reached a length of approximately 1 mm.
GP = (N/S) × 100
T50 = Ti + (Tj − Ti) × (N/2 − Ni)/(Nj − Ni)
MGT = ∑(Tx∙Nx)/N
GU = ∑[(MGT − Tx)2∙Nx]/N − 1
GS = ∑(Nx/Tx)
Here, N represents the total number of seeds germinated; S denotes the total number of seeds tested; Ni refers to the total number of seeds germinated just before 50% germination for N was reached; Nj denotes the total number of seeds germinated just after 50% germination for N was reached; Ti refers to the germination period until Ni; Tj indicates the germination period until Nj; Nx is the number of seeds germinated on the day of irradiation; and Tx represents the number of days after irradiation.
The initial seed morphology and changes during the germination process were observed using a USB microscope (AM 3111 Dino-Lite premier, AnMo Electronics Co., New Taipei City, Taiwan) at 50–60× magnification.
Statistical analyses were performed using SPSS version 12 (SPSS Inc., Chicago, IL, USA), and ANOVA was performed to analyze the results of each treatment group and the interactions among factors including habitat and treatment strategies, particularly the cold stratification-only treatment and the GA-only treatment. Duncan’s multiple range test (p < 0.05) was also used as a post hoc test to compare treatment means. Graphs were created using SigmaPlot 10.0 (SPSS Inc., Chicago, IL, USA).
3. Results
3.1. Seed Water Absorption
The seeds of S. koraiensis Nakai exhibited rapid water intake, reaching 84% of their initial mass within 3 h and exceeding 100% by 6 h, after which the hydration level remained constant for up to 48 h.
3.2. ANOVA Analysis of Germination Percentage
We observed that the cold stratification-only and the GA-only treatments significantly affected the germination process of S. koraiensis Nakai seeds (p < 0.001, Table 1). However, the effect of the seed collection location (habitat) was not statistically significant (p = 0.0767). Additionally, the interactions of the treatment type and habitat, except the interaction of the cold stratification-only treatment and habitat, significantly affected the germination process.

Table 1.
Three-way ANOVA (degrees of freedom, d.f., sum of squares, SS, mean square, MS, and level of significance, p-value) of the effects of habitat, cold stratification-only treatment, gibberellic acid (GA)-only treatment, and their interactions on the germination process of Scrophularia koraiensis.
3.3. Germination Percentage (GP)
For seeds collected from Mt. Gariwang, we recorded 11.7% germination in the control group; however, germination increased to 70.7% and 93.6% in the GA-only and cold stratification-only treatment groups, respectively. The GP was 84.4% for the cold stratification + GA treatment group (Figure 2). For seeds collected from Mt. Balwang, we observed an 8.5% GP in the control group that increased to 65.8% and 89.3% in the GA-only and cold stratification-only treatment groups, respectively. The GP was 92.2% in the cold stratification + GA treatment group. For seeds collected from Mt. Gyebang, we recorded a 35.3% GP in the control group, 57.2% in the GA-only treatment group, and 90.4% in the cold stratification-only treatment group. The GP was 81% for the cold stratification + GA treatment group. For seeds collected from Mt. Deokhang, we observed a 12.8% GP in the control group that increased to 54.0% and 76.3% in the GA-only and cold stratification-only treatment groups, respectively. The GP was 89.4% for the cold stratification + GA treatment group. The seeds of S. koraiensis Nakai exhibited fully developed embryos at the time of maturation, and the germinated seeds subsequently formed normal cotyledons (Figure 3).
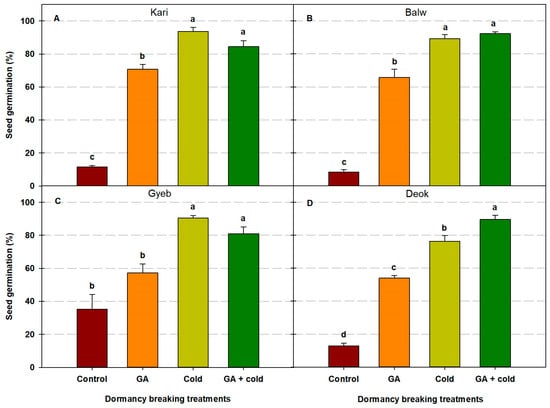
Figure 2.
Effects of seed collection location, gibberellic acid (GA)-only treatment, cold stratification-only treatment, and cold stratification + GA treatment on the seed germination of Scrophularia koraiensis. The seeds collected from Mt. Gariwang (Kari) (A), Balwang (Balw) (B), Gyebang (Gyeb) (C), and Deokhang (Deok) (D) in the Gangwon Province, Republic of Korea, were used for this experiment. Forty seeds were assessed in triplicate. Different letters indicate significant differences based on Duncan’s multiple range test (p < 0.05).
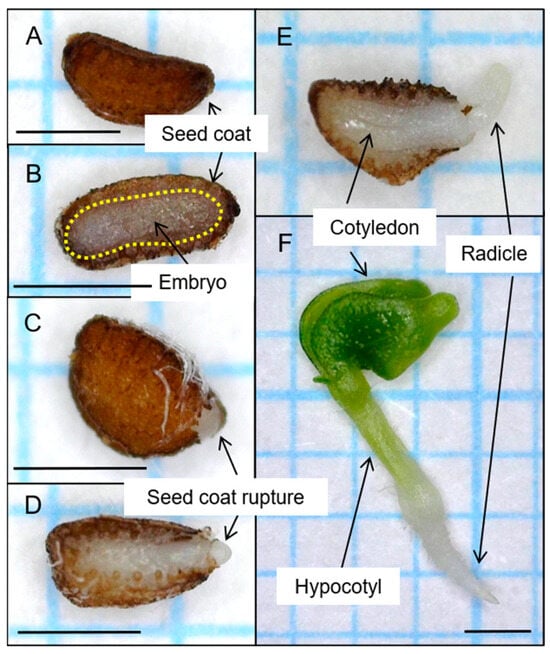
Figure 3.
Morphology, germination, and seedling growth of Scrophularia koraiensis seeds. (A) Intact seed. (B) Fully developed embryo. (C,D) Seed coat rupture. (E) Germination. (F) Seedling. Scale bar indicates 1 cm.
3.4. Germination Parameters
We observed a statistically significant effect of the cold stratification-only treatment on the germination parameters including MGT, GU, GS, and T50 (Table 2). However, the effect of the GA-only treatment was only statistically significant for GS. Additionally, the interaction between the two factors was statistically significant for all parameters except for GU.
Table 2.
Effects of habitats, cold stratification-only treatment, and GA-only treatment on mean germination time (MGT), germination uniformity (GU), germination speed (GS), and days to 50% of germination (T50) in Scrophularia koraiensis seeds.
4. Discussion
Seeds collected from Mt. Gyebang exhibited germination percentage (GP) of 35.3%, while seeds collected from the remaining three regions exhibited GPs below 20% (Figure 2). The Nikolaeva–Baskin seed dormancy classification system classifies dormancy into the following five types: physiological dormancy (PD), morphological dormancy (MD), morphophysiological dormancy (MPD), physical dormancy (PY), and combined dormancy (PY + PD) [23,24,25]. The seeds collected from all four regions had fully developed embryos at the time of detachment from the mother plant (Figure 3). Furthermore, fresh seeds absorbed more than 100% of their initial weight. These observations suggest that seed dormancy in S. koraiensis seeds is neither MD nor MPD, owing to the underdeveloped embryos, nor PY nor PY + PD, due to the impermeability of the seed coat. Instead, the results indicate that the seeds of this plant exhibit PD.
Based on the depth of PD, it has three levels, namely, non-deep, intermediate, and deep [26]. Cold (days or up to several months) or warm stratification can release the non-deep PD in seeds; additionally, reports suggest the potential of the GA-only treatment in effectively breaking seed dormancy [24,26]. Consequently, we used the cold stratification-only and GA-only treatments to break seed dormancy, which promoted germination in seeds collected from all four regions. However, for seeds collected from three regions excluding Mt. Deokhang, we observed no difference in the GP between the cold stratification-only and cold stratification + GA treatments. Therefore, the cold stratification-only treatment could sufficiently break seed dormancy, and the additional GA treatment proved unnecessary. Furthermore, the outcomes suggest the dormancy type of S. koraiensis seeds as non-deep PD. Reports suggest that dry after-ripening (DAR) positively affects breaking seed dormancy in plants with a non-deep PD, which includes seeds of Osteomeles schwerinae [27], Primula poissonii [28], Rhodiola crenulate [29], Rheum nobile [30], Lamiophlomis rotata, and Marmoritis complanatum [31], where DAR helped in reducing the depth of PD and facilitated seed germination. In this study, seeds that had been dried and stored at 5 °C for 6 months after seed collection were used. Therefore, further research is essential to determine the dormancy-breaking effect of DAR treatment on S. koraiensis seeds.
Upon summarizing the outcomes of previous studies on the genus Scrophularia, Baskin and Baskin [24] reported that the seeds of S. glabrata, S. incisa, S. marilandica, S. nodosa, and S. glabrata exhibit PD. Additionally, seeds of S. takesimensis, native to the Korean Peninsula [18], and S. stricta, native to Iran [17], also reported to demonstrate PD. The outcomes of the present study also indicated that S. koraiensis seeds exhibit PD, thereby aligning with the previous studies and indicating that PD is well conserved in plants belonging to the genus Scrophularia. The phenomenon in which ecological and physiological characteristics, such as seed dormancy, remain conserved or exhibit little change despite plants of the same genus being distributed across different continents and long-term geographically separation is referred to as “trait stasis”. In contrast, when these characteristics change gradually over geologically timescales as plants adapt to new habitats, including changes in seed dormancy and germination requirements, is known as “trait divergence (adaptation)” [32,33,34]. Accordingly, the findings of this study suggest that plants belonging to the genus Scrophularia may represent a case of “trait stasis” with respect to seed dormancy/germination.
In this experiment, the GA-only and cold stratification-only treatments effectively inhibited seed dormancy and promoted germination; however, the latter proved to be more effective (Figure 2). Adding GA treatment to the cold stratification-only treatment did not further promote germination. In contrast, adding cold stratification treatment to the GA-only treatment further promoted germination. Additionally, the cold stratification-only treatment demonstrated a positive effect on germination parameters, including MGT, GU, GS, and T50, in seeds collected from all four sites (Table 2). Previous studies conducted in S. striata [17], S. takesimensis [18], and S. marilandica [19] also report similar germination-promoting effect through the cold stratification-only or GA-only treatments. For seeds with PD, factors including the content, synthesis, and sensitivity of plant hormones GA and ABA considerably affect the breaking of seed dormancy and promotion of germination. The sensitivity of the seeds to GA increases with the progressive release of non-deep PD. Additionally, a decrease in the sensitivity of ABA influences the transition of seeds from a dormant to a nondormant state [35,36]. This implies that the cold stratification-only treatment in the present study affected the content of GA and ABA, along with their sensitivity. While exogenous application of IAA alone does not significantly affect seed germination in a wild-type Arabidopsis species, its co-application with ABA markedly inhibits germination, suggesting that ABA may suppress seed germination through the auxin signaling pathway [37,38]. Seed germination is regulated by the formation of an endogenous indole-3-acetic acid (IAA) gradient; therefore, exogenous application of IAA does not significantly affect the germination process. Considerable research has been devoted to understanding the effects of after-ripening and low-temperature stratification treatments among the various types of seed dormancy, with particular emphasis on elucidating the molecular mechanisms underlying physiological dormancy (PD) [39,40,41]. Additional research is necessary to clarify the molecular mechanisms of diverse nondeep PD types in seeds.
5. Conclusions
The primary aim of our research was to develop a seed-based mass propagation technique for breaking seed dormancy and promoting germination in S. koraiensisis seeds. The results indicated that the cold stratification-only treatment at 5 °C for 4 weeks or immersion in 800 mg∙L−1 GA3 for 24 h effectively inhibited seed dormancy and increased germination. In particular, the cold stratification-only treatment exhibited positive effects across seeds collected from four different locations, suggesting the relevance of the technique regardless of the collection location. The findings of this study suggest that S. koraiensis seeds possess non-deep PD. Furthermore, the study demonstrated that PD is highly conserved in Scrophularia species, native to various regions worldwide. Consequently, the cold stratification-only treatment can be consistently applied in future seed research and for practical mass propagation of plants within the genus Scrophularia.
Author Contributions
Conceptualization, C.H.K. and S.Y.L.; Investigation and Formal analysis, K.S.C. and S.Y.L.; Data curation, K.S.C. and S.Y.L.; Funding acquisition, C.H.K. and Y.H.R.; Methodology, Supervision, and Project administration, S.Y.L. and Y.H.R.; Writing—original draft preparation, S.Y.L. and K.S.C.; Writing—review and editing, C.H.K. and Y.H.R. All authors have read and agreed to the published version of the manuscript.
Funding
This study was supported by the Korea National Arboretum of the Korea Forest Service (Development of propagation protocols for establishing a sustainable utilization base for forest useful resource plants, grant no. KNA1-2-40-21-3) and the Basic Science Research Program through the National Research Foundation of Korea (NRF) funded by the Ministry of Education (grant number NRF-2018R1A6A1A03024862).
Data Availability Statement
The original contributions presented in this study are included in the article. Further inquiries can be directed to the corresponding authors.
Acknowledgments
Thanks to every member of Useful Plant Resources Center in Korea National Arboretum.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Stiefelhagen, H. Systematische und pflanzengeographische studien zur kenntnis der Gattung Scrophularia. Bot. Jahrb. Syst. 1910, 44, 406–496. [Google Scholar]
- Hong, D.Y. The distribution of Scrophulariaceae in the holarctic with special reference to the floristic relationships between eastern Asia and eastern north America. Ann. Missouri Bot. Gard. 1983, 70, 701–712. [Google Scholar]
- Yamazaki, T. Scrophulariaceae. In Flora of Japan; Iwatsuki, K., Yamazaki, T., Boufford, D.E., Ohba, H., Eds.; Kodansha: Tokyo, Japan, 1993; Volume IIIa, pp. 326–331. [Google Scholar]
- Jang, H.D.; Oh, B.U. A taxonomic study of Korean Scrophularia L. (Scrophulariaceae) based on morphological characters. Korean J. Plant Res. 2013, 26, 271–283. [Google Scholar] [CrossRef]
- Woo, W.S. Active principle, ρ-methoxycinnamic acid, of scrophulariae radix. Identification of ρ-methoxycinnamic acid and its antipyretic action. Korean J. Pharmacogn. 1963, 7, 55–59. [Google Scholar]
- Yook, C.S. Coloured Medicinal Plants of Korea; Academy Publishing Co.: Seoul, Republic of Korea, 1989; pp. 494–495. (In Korean) [Google Scholar]
- Lee, J.H.; Jo, I.H.; Lee, J.W.; Park, C.G.; Bang, K.H.; Kim, H.S.; Park, C.B. Molecular authentication of Scrophularia herbs by PCR-RFLP based on rpl-5 region of mitochondrial DNA. Korean J. Med. Crop Sci. 2010, 18, 173–179. (In Korean) [Google Scholar]
- Hwang, S.Y.; Byun, J.H.; Song, H.J.; Shin, M.K. A herbalogical study on the plants of Scrophulariaceae in Korea. Kor. J. Herbol. 1998, 13, 241–270. (In Korean) [Google Scholar]
- Nam, H.-H.; Lee, A.Y.; Seo, Y.-S.; Park, I.; Yang, S.; Chun, J.M.; Moon, B.C.; Song, J.-H.; Kim, J.-S. Three Scrophularia species (S. buergeriana, S. koraiensis, and S. takesimensis) inhibit RANKL-induced osteoclast differentiation in bone marrow-derived macrophages. Plants 2020, 9, 1656. [Google Scholar] [CrossRef]
- Han, K.S.; So, S.K.; Lee, C.H.; Kim, M.Y. Taxonomy of the genus Scrophularia (Scrophulariaceae) in Korea. Korean J. Plant Taxon. 2009, 39, 237–246. [Google Scholar] [CrossRef]
- Jang, H.D.; Kim, T.H.; Oh, B.U. A taxonomic review of Scrophularia kakudensis Franch. and its relatives. Korean J. Plant Taxon. 2011, 41, 345–352. (In Korean) [Google Scholar] [CrossRef]
- Chung, G.Y.; Chang, K.S.; Chung, J.-M.; Choi, H.J.; Paik, W.-K.; Hyun, J.-O. A checklist of endemic plants on the Korean Peninsula. Korean J. Plant Taxon. 2017, 47, 264–288. [Google Scholar] [CrossRef]
- Chung, G.Y.; Jang, H.-D.; Chang, K.S.; Choi, H.J.; Kim, Y.-S.; Kim, H.-J.; Son, D.C. A checklist of endemic plants on the Korean Peninsula II. Korean J. Plant Taxon. 2023, 53, 79–101. [Google Scholar] [CrossRef]
- Jung, T.-Y.; Lee, A.Y.; Song, J.-H.; Lee, M.Y.; Lim, J.-O.; Lee, S.-J.; Ko, J.-W.; Shin, N.-R.; Kim, J.-C.; Shin, I.-S.; et al. Scrophularia koraiensis Nakai attenuates allergic airway inflammation via suppression of NF-κB and enhancement of Nrf2/HO-1 signaling. Antioxidants 2020, 9, 99. [Google Scholar] [CrossRef] [PubMed]
- National Institute of Biological Resources (NIBR). Biodiversity of the Korean Peninsula. Available online: https://wims.me.go.kr/wims/minwon/cms/cmsCont.do?cntnts_sn=8 (accessed on 5 June 2025).
- Korea National Arboretum. 300 Target Plants Adaptable to Climate Change in the Korean Peninsula; Korea National Arboretum: Pocheon, Republic of Korea, 2010; p. 104. (In Korean)
- Karavani, B.; Afshari, R.T.; Hosseini, N.M.; Oveisi, M.; Miranshahi, B. Assessment of seed dormancy in Scrophularia striata. Seed Sci. Technol. 2016, 44, 218–223. [Google Scholar] [CrossRef]
- Kang, J.H.; Choi, K.O.; Ahn, S.Y.; Kim, D.S.; Chon, Y.S.; Yun, J.G. Improvement of seed germination in Scrophularia takesimensis, Korean native plant. Korean J. Hortic. Sci. Technol. 2009, 27, 535–539. [Google Scholar]
- Nurse, R.E.; Cavers, P.B. The germination characteristics of Scrophularia marilandica L. (Scrophulariaceae) seeds. Plant Ecol. 2008, 196, 185–196. [Google Scholar] [CrossRef]
- Gordon, A.G. The germination resistance test—A new test for measuring germination quality of cereals. Can. J. Plant Sci. 1971, 51, 181–183. [Google Scholar] [CrossRef]
- Coolbear, P.; Francis, A.; Grierson, D. The effect of low temperature pre-sowing treatment on the germination performance and membrane integrity of artificially aged tomato seeds. J. Exp. Bot. 1984, 35, 1609–1617. [Google Scholar] [CrossRef]
- Andalibi, B.; Zangani, E.; Haghnazari, A. Effects of water stress on germination indices in six rapeseed cultivars (Brassica napus L.). Iran J. Agric. Sci. 2005, 36, 457–463. [Google Scholar]
- Baskin, J.M.; Baskin, C.C. A classification system for seed dormancy. Seed Sci. Res. 2004, 14, 1–16. [Google Scholar] [CrossRef]
- Baskin, C.C.; Baskin, J.M. Seeds: Ecology, Biogeography, and Evolution of Dormancy and Germination, 2nd ed.; Elsevier Academic Press: San Diego, CA, USA, 2014. [Google Scholar]
- Baskin, C.C.; Baskin, J.M. Germinating seeds of wildflowers, an ecological perspective. HortTechnology 2004, 14, 467–473. [Google Scholar] [CrossRef]
- Baskin, C.C.; Baskin, J.M. Mimicking the natural thermal environments experienced by seeds to break physiological dormancy to enhance seed testing and seedling production. Seed Sci. Technol. 2022, 50, 22–29. [Google Scholar] [CrossRef]
- Geng, B.; Peng, D. Seed dormancy and germination characteristics of Osteomeles schwerinae, a riparian plant endemic to the Hengduan Mountains and adjacent regions. Seed Sci. Technol. 2022, 50, 339–344. [Google Scholar] [CrossRef]
- Peng, D.; Hu, X.; Sun, H.; Li, Z. Dry after-ripening, light, cold stratification and temperature effects on seed germination of Primula poissonii from Yunnan, China. Seed Sci. Technol. 2019, 47, 301–306. [Google Scholar] [CrossRef]
- Peng, D.; Geng, B.; Qin, Y.; Yang, L.-E.; Baskin, J.M.; Baskin, C.C. Non-deep physiological dormancy in seeds of two endangered medicinal alpine species of Rhodiola from the Hengduan Mountains of southwest China. Seed Sci. Technol. 2023, 51, 297–303. [Google Scholar] [CrossRef]
- Peng, D.L.; Chen, Z.; Hu, X.J.; Li, Z.M.; Song, B.; Sun, H. Seed dormancy and germination characteristics of two Rheum species in the Himalaya-Hengduan Mountains. Plant Divers. 2017, 39, 180–186. [Google Scholar] [CrossRef] [PubMed]
- Peng, D.L.; Hu, X.J.; Yang, J.; Sun, H. Seed dormancy, germination and soil seed bank of Lamiophlomis rotata and Marmoritis complanatum (Labiatae), two endemic species from Himalaya–Hengduan Mountains. Plant Biosyst. 2017, 152, 642–648. [Google Scholar] [CrossRef]
- Adams, C.A.; Baskin, J.M.; Baskin, C.C. Trait stasis versus adaptation in disjunct relict species: Evolutionary changes in seed dormancy-breaking and germination requirements in a subclade of Aristolochia subgenus Siphisia (Piperales). Seed Sci. Res. 2005, 15, 161–173. [Google Scholar] [CrossRef]
- Walck, J.L.; Hidayati, S.N.; Okagami, N. Seed germination ecophysiology of the Asian species Osmorhiza aristata (Apiaceae): Comparison with its North American congeners and implications for evolution of types of dormancy. Am. J. Bot. 2002, 89, 829–835. [Google Scholar] [CrossRef]
- Hidayati, S.N.; Walck, J.L.; Taylor, J. Seed germination ecophysiology of two western North American species of Sambucus (Caprifoliaceae sensu lato), and comparisons with eastern North American and European congeners. Plant Species Biol. 2010, 25, 203–213. [Google Scholar] [CrossRef]
- Finch-Savage, W.E.; Leubner-Metzger, G. Seed dormancy and the control of germination. New Phytol. 2006, 171, 501–523. [Google Scholar] [CrossRef]
- Kucera, B.; Cohn, M.A.; Leubner-Metzger, G. Plant hormone interactions during seed dormancy release and germination. Seed Sci. Res. 2005, 15, 281–307. [Google Scholar] [CrossRef]
- Brady, S.M.; Sarkar, S.F.; Bonetta, D.; McCourt, P. The ABSCISIC ACID INSENSITIVE 3 (ABI3) gene is modulated by farnesylation and is involved in auxin signaling and lateral root development in Arabidopsis. Plant J. 2003, 34, 67–75. [Google Scholar] [CrossRef]
- Liu, X.; Zhang, H.; Zhao, Y.; Feng, Z.; Li, Q.; Yang, H.Q.; Luan, S.; Li, J.; He, Z.H. Auxin controls seed dormancy through stimulation of abscisic acid signaling by inducing ARF-mediated ABI3 activation in Arabidopsis. Proc. Natl. Acad. Sci. USA 2013, 110, 15485–15490. [Google Scholar] [CrossRef] [PubMed]
- Loades, E.; Pérez, M.; Turečková, V.; Tarkowská, D.; Strnad, M.; Seville, A.; Nakabayashi, K.; Leubner-Metzger, G. Distinct Hormonal and Morphological Control of Dormancy and Germination in Chenopodium album Dimorphic Seeds. Front. Plant Sci. 2023, 14, 1156794. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Bezodis, W.; González-Suárez, P.; Knitlhoffer, V.; Goldson, A.; Lister, A.; Macaulay, I.; Penfield, S. Adaptation of Seed Dormancy to Maternal Climate Occurs via Intergenerational Transport of Abscisic Acid. Proc. Natl. Acad. Sci. USA 2025, 122, e2519319122. [Google Scholar] [CrossRef] [PubMed]
- Sajeev, N.; Koornneef, M.; Bentsink, L. A Commitment for Life: Decades of Unraveling the Molecular Mechanisms behind Seed Dormancy and Germination. Plant Cell 2024, 36, 1358–1376. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).