Abstract
Propolis is a natural honeybee product, which is known for its beneficial pharmacological effects and use in traditional medicine. This study aspired to investigate the chemical and antioxidant properties of five propolis specimens, followed by the analysis of their in vitro impact on bovine spermatozoa vitality and oxidative profile. Semen samples from 10 breeding bulls were incubated in the absence or presence of 50 µg/mL propolis extracts, and selected sperm quality parameters including motility, mitochondrial activity, cAMP concentration, NAD+/NADH ratio, reactive oxygen species (ROS) and superoxide production were assessed at 2 h and 24 h. Sperm motion behavior and mitochondrial function were stabilized particularly in the presence of propolis collected from Hrabské and Litpovský Hrádok, Slovakia. At the same time, all propolis specimens acted as significant ROS quenchers and stabilized the oxidative milieu of the sperm cultures. Our data suggest that propolis may stabilize the mitochondrial function of spermatozoa while preventing excessive oxidative insults, thereby showing the potential to sustain the sperm motility of extended semen.
1. Introduction
Artificial reproduction, considered to be a main driving force of animal breeding programs, basically relies on extended ejaculates [1]. Nevertheless, full benefits of semen handling may be complicated by the loss of sperm cell viability due to increased ex vivo stress conditions. At the same time, since the seminal plasma is diluted with an extender, male gametes lose an important source of nutrients and antioxidants [2]. To compensate for any potential negative consequences of high reactive oxygen species (ROS) levels on the sperm survival, several papers have studied the impact of various biomolecules and compounds that could serve as bovine semen extender supplements, with often contradictory outcomes [3,4]. The ultimate supplement for sperm cultures should ideally stimulate the mitochondrial apparatus in charge of energy synthesis for a proper sperm motility, while at the same time, act as an antioxidant to protect critical sperm structures against oxidative stress [5]. More and more evidence suggests that both these goals could be achieved by taking advantage of natural products such as plant extracts, herbs or honeybee products [6].
Propolis is a term referring to a brown or red resinous substance collected by honeybees from tree buds, sap flows or other plant sources. This honeybee product has been used in folk medicine for centuries due to its great variety of its beneficial pharmacological effects, encompassing antiviral, antibacterial, antifungal, anti-inflammatory, immunomodulatory, chemo-preventive, antioxidant and antitumor activities (reviewed by [7]). As such, propolis has been subjected to a variety of medical studies over previous decades.
Several recent in vivo studies have found that propolis could be effective in the stimulation and protection of the male reproductive system. Capucho et al. [8] suggest that green Brazilian propolis administration to rats resulted in an increased spermatogenesis, accompanied by lower oxidative stress indices. Further studies have unraveled that propolis stimulates testosterone production [9], while at the same time, preventing excessive germ cell apoptosis and improving testicular damage in mice exposed to toxicants [10]. In the meantime, preliminary in vitro reports have implied that propolis improves the functional efficiency of human spermatozoa collected from normozoospermic [11] as well as asthenozoospermic men [12], thereby exhibiting the potential to be used for in vitro preservation and activation of spermatozoa in assisted reproduction.
As such, this paper was designed to analyze the chemical characteristics of five propolis specimens of Slovak origin, as well as their in vitro effects on the functional characteristics and oxidative profile of bovine spermatozoa. Being a cost-effective source of antioxidant molecules of natural origin, this honeybee product could become an appealing alternative to supplements traditionally used in extenders for bovine ejaculates.
2. Materials and Methods
2.1. Propolis Collection and Extract Preparation
Five different propolis samples were obtained from local beekeepers from various Slovak locations (Table 1) in September 2017. The samples were frozen at −20 °C and ground in a mill to obtain powder, which was subsequently mixed (30 g) with 100 mL 70% ethanol (Centralchem, Bratislava, Slovakia). The mixture was subjected to a cold extraction by keeping it at room temperature and shaking the contents every 24 h for 7 days. The extract was filtered using Whatman filter paper (Sigma-Aldrich, St. Louis, MO, USA), the resulting filtrate was made up to 100 mL with 70% ethanol and kept in darkness at 4 °C. For the sperm cultures, the propolis extracts were further diluted with 70% ethanol to reach a final concentration of 50 µg/mL, with ethanol being present in the culture medium below 1%, which is considered not to be toxic to the sperm cells [13].

Table 1.
Details of the locations where propolis was collected.
2.2. Characterization of the Extract
Total phenolic content of the propolis extracts was assessed with the Folin–Ciocalteu protocol [14]. A total of 100 mL of each extract was treated with 0.1 mL of the Folin–Ciocalteu reagent (Sigma-Aldrich, St. Louis, MO, USA), 1 mL 20% sodium carbonate (Centralchem, Bratislava, Slovakia) and 8.8 mL ultrapure water. Following a 30 min incubation under dark conditions, absorbance of the mixtures was assessed with the Jenway 6405 UV/Vis spectrophotometer (Cole-Pharmer Ltd., Vernon Hills, IL, USA) at 700 nm. The results were calculated using a gallic acid (1–100 mg/mL; Sigma-Aldrich, St. Louis, MO, USA) standard curve and are expressed in mg gallic acid equivalents (GAE)/g dry extract [14,15].
A modified Willett technique was used to quantify the total content of flavonoids [16]. In total, 500 µL of each extract was treated with 0.1 mL of 10% aluminum chloride (Sigma-Aldrich, St. Louis, MO, USA) dissolved in ethanol, 0.1 mL of 1 mol/L potassium acetate (Sigma-Aldrich, St. Louis, MO, USA) and 4.3 mL ultrapure water. Following a 30 min incubation under dark conditions, absorbance of the mixtures was assessed with the Jenway 6405 UV/Vis spectrophotometer (Cole-Pharmer Ltd., Vernon Hills, IL, USA) at 415 nm. The results were calculated using a quercetin (1–400 mg/L; Sigma-Aldrich, St. Louis, MO, USA) standard curve and are expressed in mg quercetin equivalents (QUE)/g dry extract [15].
The 2,2-diphenyl-1-picrylhydrazyl (DPPH) assay was used to study the free-radical scavenging activity of the extracts [17]. Each extract (400 µL) was mixed with 3.6 mL DPPH reagent (0.025 g DPPH/100 mL methanol; Sigma-Aldrich, St. Louis, MO, USA). The reaction mixture was subjected to absorbance measurement with the Jenway 6405 UV/Vis spectrophotometer (Cole-Pharmer Ltd., Vernon Hills, IL, USA) at 515 nm. The results were calculated using a trolox (6-hydroxy-2,5,7,8-tetramethylchroman-2-carboxylic acid; 10–100 mmol/L; Sigma-Aldrich, St. Louis, MO, USA) standard curve and are expressed in mmol Trolox equivalents (TEAC)/g dry extract [15].
To quantify the selected phenolic compounds, HPLC (high performance liquid chromatography) standards, acetonitrile (HPLC grade) and phosphoric acid (ACS grade) were obtained from Sigma-Aldrich (St. Louis, MO, USA). The analysis was carried out using the Agilent 1260 Infinity high-performance liquid chromatograph (Agilent Technologies, Santa Clara, CA, USA) with a Purosphere reverse phase C18 column (Merck, Darmstadt, Germany). The analysis was carried out as previously described [15], and the collected data were analyzed with the Agilent OpenLab ChemStation software for LC 3D Systems.
2.3. Sperm Cultures and Analysis
Semen specimens were collected from 10 adult Holstein Friesian breeding bulls (Slovak Biological Services, Nitra, Slovakia) using an artificial vagina. Only ejaculates with a minimum 70% initial motility and concentration of 1 billion sperm/mL were used for subsequent cultures. Following sample transfer to the laboratory, each of them was diluted in PBS (Dulbecco’s phosphate buffered saline without Ca or Mg; Sigma-Aldrich, St. Louis, MO, USA), containing 50 μg/mL of each propolis extract at a dilution ratio of 1:40 and kept at 22–25 °C. An equal aliquot of ethanol was added to the control group. Specific sperm analyses were carried out at 2 and 24 h [15].
Sperm motility was assessed using the IVOS II computer-assisted sperm analysis (CASA; Hamilton Thorne Biosciences, Beverly, MA, USA). The fluorescent IDENT dye was used to stain the cells (Hamilton Thorne Biosciences, Beverly, MA, USA) and assessed as previously described [15].
Sperm mitochondrial function expressed through the mitochondrial succinate dehydrogenase activity was evaluated using the colorimetric metabolic activity (MTT) test, employing tetrazolium bromide (3-(4,5-dimetylthiazol-2-yl)-2,5-diphenyltetrazolium bromide; MTT). A total of 20 μL of tetrazolium (5 mg/mL) was administered to each sperm suspension. After 2 h of incubation (37 °C, shaker), the reaction was stopped with 80 μL isopropanol (Centralchem, Bratislava, Slovakia). Absorbance of the suspensions was measured at 570 nm against 620 nm as the reference using the Glomax plate spectrophotometer (Promega Corporation, Madison, WI, USA). The results are expressed as percentage of the control set to 100% [15].
The cAMP concentration was quantified using a commercial cAMP-GloTM kit (Promega, Madison, WI, USA), based on protein kinase A enhancement, followed by a decrease in adenosine triphosphate (ATP). This reaction was accompanied by a decreased luminescent signal catalyzed by luciferase, which was monitored by the Glomax Multi+ combined spectro-fluoro-luminometer (Promega Corporation, Madison, WI, USA) [18].
The NAD+/NADH ratio was assessed based to the levels of lactate and pyruvate. Lactate was assessed with the lactate assay kit (Sigma-Aldrich, St. Louis, MO, USA), while pyruvate was quantified with the pyruvate assay kit (Sigma-Aldrich, St. Louis, MO, USA). Both colorimetric reactions were measured with the Glomax Multi+ combined spectro-fluoro-luminometer (Promega Corporation, Madison, WI, USA) at 570 nm [19].
ROS production was assessed by luminol-based chemiluminescence. All samples were treated with luminol (5-amino-2,3-dihydro-1,4-phthalazinedione; Sigma-Aldrich, St. Louis, MO, USA; 5 mmol/L), and the chemiluminescent signal was monitored in fifteen 1 min cycles with the Glomax Multi+-combined spectro-fluoro-luminometer (Promega Corporation, Madison, WI, USA) [15]. Negative controls run in parallel contained the culture media without spermatozoa, while positive controls were comprised of each culture medium, luminol and hydrogen peroxide (H2O2, 30%; 8.8 M; Sigma-Aldrich, St. Louis, MO, USA). The results are expressed as relative light units (RLU)/s/106 sperm.
The nitroblue tetrazolium (NBT) test was used to measure superoxide production, based on a reduction of nitroblue tetrazolium chloride (2,20-bis(4-nitrophenyl)-5,50-diphenyl-3,30-(3,30-dimethoxy-4,40-diphenylene) ditetrazolium chloride; Sigma-Aldrich, St. Louis, MO, USA) by the superoxide radical. NBT tetrazolium was dissolved in PBS/1.5% DMSO (Sigma-Aldrich, St. Louis, MO, USA) at 1 mg/mL and administered to the suspensions (100 μL per well). Following incubation (37 °C, shaker, 1 h), the suspensions were washed with PBS and centrifuged at 305× g for 10 min twice. Finally, the samples were treated with 2 mol/L potassium hydroxide (Centralchem, Bratislava, Slovakia) in DMSO. The resulting optical density was monitored at 620 nm against 570 nm as the reference with the Glomax plate spectrophotometer (Promega Corporation, Madison, WI, USA). The data are expressed in percentage of the control set to 100% [15,19].
Statistical analysis was performed with the GraphPad Prism program (version 9.5.1. for Mac; GraphPad Software, La Jolla, CA, USA). The data are expressed as mean ± standard deviation. One-way ANOVA and Dunnett’s test were used for specific statistical evaluations. The level of significance was set at p < 0.05, p < 0.01, p < 0.001 and p < 0.0001.
3. Results
3.1. Chemical Properties of the Propolis Extract
The collected data reveal that the total phenolic content ranged between 61.76 ± 7.04 and 89.55 ± 9.47 mg GAE/g dry extract with the highest content of phenols detected in Sample 5 (Table 2). The flavonoid content varied between 20.41 ± 3.09 and 38.73 ± 5.05 mg QUE/g dry extract (Table 2). The highest concentration of flavonoids was also recorded in Sample 5. At the same time, this sample presented with the highest ROS-scavenging activity amongst all the tested samples of propolis, as indicated by the DPPH assay (Table 2).
Table 2.
Chemical properties of the propolis extracts.
With the help of HPLC, we identified compounds as caffeic acid, t-ferulic acid, p-coumaric acid, rutin, kaempferol, quercetin, apigenin and chrysin (Table 3). Although standards for other phenolic compounds such as benzoic acid, pinocembrin, vanillin, naringenin, luteolin or genistein were used for the analysis, these were not identified in any sample.
Table 3.
Analysis of the propolis extracts by high performance liquid chromatography (HPLC).
3.2. Sperm Characteristics
According to the CASA analysis, a significantly higher sperm motility followed exposure to propolis sample 1 (p < 0.05), sample 3 (p < 0.01) and sample 5 (p < 0.0001) (Figure 1) in comparison to the untreated control at 2 h. At the end of the in vitro culture (24 h), a significantly higher motility in the groups was subjected to propolis sample 1 (p < 0.0001), sample 2 (p < 0.01), sample 3 (p < 0.0001) and sample 5 (p < 0.0001) in comparison to the control.
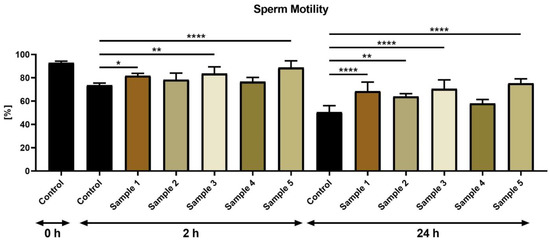
Figure 1.
Bovine sperm motility (%) in the absence (control) or presence of propolis during different time periods. * p < 0.05, ** p < 0.01, **** p < 0.0001.
The MTT test unraveled that following 2 h, the presence of propolis samples 3 (p < 0.01) and 5 (p < 0.001) led to significantly increased mitochondrial activity (Figure 2). At 24 h, a significant mitochondria-promoting effect of propolis administration to the sperm suspensions was observed, most notably in the case of sample 1 (p < 0.01), sample 3 (p < 0.0001) and sample 5 (p < 0.0001).
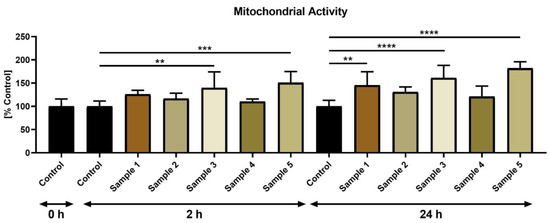
Figure 2.
Bovine sperm mitochondrial activity (%) in the absence (control) or presence of propolis during different time periods. ** p < 0.01, *** p < 0.001, **** p < 0.0001.
In the case of cAMP concentration, no significant differences were found amongst the control and experimental groups at 2 h (Figure 3). At the end of the experiment (24 h), a significantly higher cAMP concentration was observed in the experimental groups subjected to propolis sample 3 (p < 0.05) and sample 5 (p < 0.01) in comparison to the control.
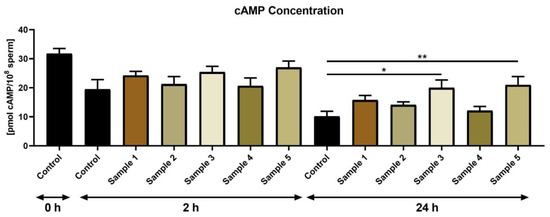
Figure 3.
cAMP concentration of bovine spermatozoa (pmol/108 sperm) in the absence (control) or presence of propolis during different time periods. * p < 0.05, ** p < 0.01.
Similar to cAMP, no significant changes were found in the case of the NAD+/NADH ratio between the control and experimental groups at 2 h (Figure 4). Assessment of the ratio at 24 h revealed significant differences amongst the control and experimental group supplemented with the propolis sample 5 (p < 0.05).
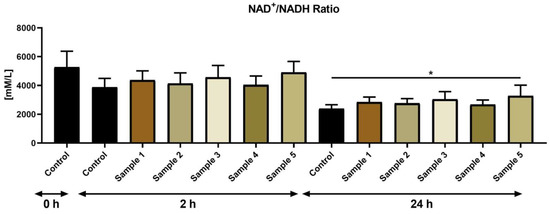
Figure 4.
NAD+/NADH of bovine spermatozoa (mM/L) in the absence (control) or presence of propolis during different time periods. * p < 0.05.
ROS-scavenging properties of all propolis samples were confirmed by the luminometric assay (Figure 5). Significantly lower ROS levels were observed in all experimental groups against the control at 2 h (p < 0.01). Significantly lower ROS production was also observed following 24 h in sperm cultures exposed to the propolis sample 1 (p < 0.05), 3 and 5 (p < 0.01) in comparison with the control.
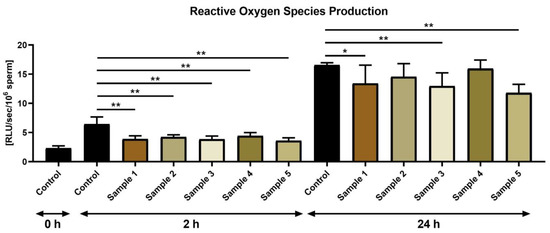
Figure 5.
ROS production by bovine spermatozoa (RLU/sec/106 sperm) in the absence (control) or presence of propolis during different time periods. * p < 0.05, ** p < 0.01.
The NBT test revealed that the propolis extracts possess superoxide-quenching properties (Figure 6). Significantly reduced superoxide levels were recorded following 2 h of the in vitro culture in the case of propolis samples 1 (p < 0.01), 2 (p < 0.05), 3 and 5 (p < 0.001), while at time 24 h, superoxide levels were significantly lower in all experimental groups (p < 0.001 with regards to sample 3; p < 0.0001 in case of samples 1, 2, 4 and 5).
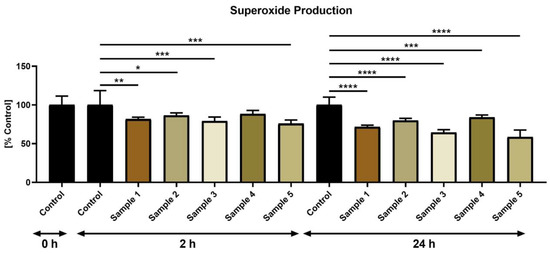
Figure 6.
Superoxide production by bovine spermatozoa (%) in the absence (control) or presence of propolis during different time periods. * p < 0.05, ** p < 0.01, *** p < 0.001, **** p < 0.0001.
4. Discussion
Since ex vivo semen handling may cause damage to cellular components with detrimental effects on sperm function, administration of synthetic antioxidants to semen extenders has become routine during sperm preservation, although it often adds to the cost of insemination doses [2,5,20]. As an alternative, natural products are increasingly gaining attention as a source of effective and affordable biomolecules that could be used as supplements during ex vivo semen processing (reviewed by [21]). Although propolis has been found to alleviate male reproductive dysfunction, limited data are available on its impact on ejaculated spermatozoa. Hence, this study aimed to characterize the chemical properties of five different propolis extracts, accompanied by an evaluation of their in vitro effects on the sperm metabolic activity and oxidative properties.
Over the past few years, propolis has received special attention as a rich source of bioactive compounds, with various beneficial health effects [7,22]. A constraining body of evidence suggests that the beneficial properties of propolis may be attributed to a complex chemical structure and antioxidant behavior of molecules, such as polyphenols and flavonoids [23,24,25]. The biochemical analysis of the propolis extracts revealed that this honeybee product is a rich source of phenolic compounds, which is in accordance with the findings by Kurek-Górecka [25], Romero et al. [26] and Kumazawa et al. [27] on propolis samples collected from different locations around the globe. On the other hand, the total flavonoid concentration found in our samples was lower when compared with other studies [25,28,29], possibly because the extraction methods or solvents differed among the studies. Furthermore, the discrepancies in the general concentrations of biomolecules may be explained by different analytical protocols selected for the experimental design of the studies. An important point to be considered is that the chemical properties of propolis depend on the time or place of collection, as well as plant material, making it a highly variable natural product [11].
Several studies addressed the chemical composition of propolis. While it is composed mainly of resin and wax, more than 300 molecules have been detected in propolis, such as essential and aromatic oils, pollen, as well as minor compounds such as terpenes, minerals, flavonoids, beta-steroids and vitamins [11,25,26,30]. Similar to our data, the phytochemical investigation by Cedikova et al. [11] revealed that amongst the most notable phytochemicals were trans-Ferulic acid, p-Coumaric acid and kaempferol. Furthermore, Kurek-Górecka et al. [25] and Volpin et al. [30] also detected caffeic acid, quercetin and apigenin in propolis samples from Poland, Turkey and Uruguay. Interestingly, rutin was not detected in the above-mentioned studies. On the other hand, vanillin was not identified in our samples. These differences may be the result of varying sets of standards chosen for the chemical analysis or by differences in the extraction protocol, time or solvents used. Other factors such as the place or time of harvest should also be considered.
During the previous decade, honeybee products have become a popular research topic, particularly because of a significant antioxidant behavior, which is associated with their chemical properties. Propolis is known to contain large amounts of polyphenols and flavonoids, which are strongly related with its ROS-scavenging properties [24,25,27]. Data collected from the DPPH assay match the reports of Rosli et al. [31] and Shehata et al. [32] on Trigona Apicalis propolis and propolis samples collected from Egypt, Saudi Arabia, Oman, China or Brazil, respectively. Finally, a higher DPPH score was observed by Kurek-Górecka et al. [25], which correlated well with the ABTS (2,2′-azino-bis(3-ethylbenzothiazoline-6-sulfonic acid) assay based on a similar experimental principle (ROS scavenging-based colorization).
In this study, propolis extracts administered to bovine sperm cultures acted as efficient motility- and metabolism-preserving supplements. Mitochondrial activity is an essential factor for critical sperm functions and the fertilization process [33]. On the other hand, mitochondria are thought to act as primary ROS producers, physiological amounts of which are important for the sperm cell to accomplish capacitation and hyperactivation. Inversely, mitochondrial dysfunction accompanied by ROS overproducion may deteriorate the sperm architecture and behavior [34]. ROS can, therefore, exhibit bipolar effects on sperm physiology, which is why an adequate oxidative milieu is crucial for an optimal state of male fertility [33]. NADH ubiquinone oxidoreductase and cytochrome bc1 complex have been acknowledged as primary sources of ROS [34]. When superoxide is produced, it will be briskly dismutated to hydrogen peroxide, both of which may contribute to the creation of the highly reactive hydroxyl radical. Oxidative overload may subsequently compromise the mitochondrial transition pores, resulting in changes of their dynamics and a disrupted membrane permeability [34,35].
Cedikova et al. [11] and Das et al. [36] reported that propolis or its phenols inhibited the depolarization of mitochondrial membranes through stabilization of the inner mitochondrial membrane. Furthermore, it was revealed that exposure of mitochondria to propolis resulted in a higher effectivity of the electron transport through the complex II. The possible molecular pathway underlying complex II activation has been proposed by Cimen et al. [37], according to whom kaempferol enhanced deacetylation and thus the activity of succinate dehydrogenase. At the same time, quercetin found in our propolis specimens has been shown to affect the activity of complex I, leading to a rise in ATP synthesis and a concomitant lower incidence of oxidative stress [38]. Finally, previous studies [39,40] reported that several phenolic acids may act as electron acceptors at the complex I, leading to increased ATP synthesis. In conclusion, we may hypothesize that the biomolecules present in propolis may stabilize the structural integrity and functional attributes of sperm mitochondria and prevent their disintegration triggered by ROS overproduction, leading to a more effective conservation of the sperm motion.
To our knowledge, two major in vitro studies have been published on the effects of propolis on male gametes. According to Cedikova et al. [11], oxygen consumption by spermatozoa determined by high-resolution respirometry was significantly higher in the samples exposed to propolis. Propolis also increased uncoupled respiration and complex IV activity, whereas the mitochondrial membrane potential remained unchanged. In the meantime, Al-Nawab et al. [12] reported a significant stimulating effect of propolis on active sperm motion behavior and morphology before and after activation using PBS and Hams F-12 media.
Our luminometric and NBT data highlight strong antioxidant abilities of biomolecules found in propolis and support the findings of Kocot et al. [23] and Kurek-Górecka [25], who suggest that the phenolic nature of biomolecules found in propolis contributes to their ROS-scavenging action by removing singlet oxygen, superoxide, hydrogen peroxide and hydroxyl radical. Moreover, it has been previously postulated that flavonoids such as quercetin and kaempferol could stimulate enzymatic antioxidants during episodes of increased oxidative insults [21]. Based on these characteristics, we may hypothesize that propolis can contribute to an increased stability of male reproductive cells subjected to stressful conditions.
The most important limitation of this paper lies in the inherently complex nature of propolis. Honeybee products are complex and heterogenous resources, comprising a variety of molecules with a significant antioxidant and cell-stimulating behavior in synergy or antagony. Information on the individual impact of different components of propolis on the sperm structure and/or function is currently limited and calls for further research. Hence, future studies should be aimed at studying the specific contribution of the biomolecules present in propolis to its protective and antioxidant effects. Furthermore, the full potential of propolis should be verified by assessing its impact on the sperm fertilization potential under conditions simulating oxidative stress.
Author Contributions
Conceptualization, E.T. and M.K.; methodology, E.T. and J.A; validation, E.T., J.Á. and M.K.; investigation, E.T., J.Á., M.Ď. and M.K.; resources, E.T. and M.K.; writing—original draft preparation, E.T. and M.Ď.; writing—review and editing, E.T.; project administration, E.T. and M.K.; funding acquisition, E.T. and M.K. All authors have read and agreed to the published version of the manuscript.
Funding
This research was financially supported by the Slovak Research and Development Agency (Grants no. APVV-15-0544, APVV-20-0058 and APVV-21-0095) and by the Operational program Integrated Infrastructure, within the project Demand-driven research for sustainable and innovative food, Drive4SIFood 313011V336, cofinanced by the European Regional Development Fund.
Institutional Review Board Statement
The animals and sample collections were carefully handled in accordance with the ethical guidelines stated in the Slovak Animal Protection Regulation RD 377/12, which conforms to European Union Regulation 2010/63. Since semen collection is routinely performed by the breeding company Slovenské biologické služby, a.s., causing no harm or discomfort, special ethical approval was not needed for this type of experiment.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data reported in this study are available on request from the corresponding author.
Acknowledgments
We wish to thank the Center for Animal Reproduction (CeRA) Team of Excellence for their support.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Barszcz, K.; Wiesetek, D.; Wasowicz, M.; Kupczynska, M. Bull semen collection and analysis for artificial insemination. J. Agric. Sci. 2007, 4, 1–10. [Google Scholar] [CrossRef]
- Khan, I.M.; Cao, Z.; Liu, H.; Khan, A.; Rahman, S.U.; Khan, M.Z.; Sathanawongs, A.; Zhang, Y. Impact of Cryopreservation on Spermatozoa Freeze-Thawed Traits and Relevance OMICS to Assess Sperm Cryo-Tolerance in Farm Animals. Front. Vet. Sci. 2021, 8, 609180. [Google Scholar] [CrossRef]
- Bucak, M.N.; Ataman, M.B.; Başpınar, N.; Uysal, O.; Taşpınar, M.; Bilgili, A.; Öztürk, C.; Güngör, Ş.; İnanç, M.E.; Akal, E. Lycopene and resveratrol improve post-thaw bull sperm parameters: Sperm motility, mitochondrial activity and DNA integrity. Andrologia 2015, 47, 545–552. [Google Scholar] [CrossRef] [PubMed]
- Riesco, M.F.; Alvarez, M.; Anel-Lopez, L.; Neila-Montero, M.; Palacin-Martinez, C.; Montes-Garrido, R.; Boixo, J.C.; de Paz, P.; Anel, L. Multiparametric Study of Antioxidant Effect on Ram Sperm Cryopreservation—From Field Trials to Research Bench. Animals 2021, 11, 283. [Google Scholar] [CrossRef] [PubMed]
- Nazari, M. A review study on the effect of various antioxidant supplements on maintaining and improving the performance of sperm parameters. Iran. J. Biol. 2021, 5, 69–77. [Google Scholar]
- Martins, R.V.L.; Silva, A.M.S.; Duarte, A.P.; Socorro, S.; Correia, S.; Maia, C.J. Natural Products as Protective Agents for Male Fertility. BioChem 2021, 1, 122–147. [Google Scholar] [CrossRef]
- Braakhuis, A. Evidence on the Health Benefits of Supplemental Propolis. Nutrients 2019, 11, 2705. [Google Scholar] [CrossRef]
- Capucho, C.; Sette, R.; de Souza Predes, F.; de Castro Monteiro, J.; Pigoso, A.A.; Barbieri, R.; Dolder, M.A.; Severi-Aguiar, G.D. Green Brazilian propolis effects on sperm count and epididymis morphology and oxidative stress. Food Chem. Toxicol. 2012, 50, 3956–3962. [Google Scholar] [CrossRef]
- Handayani, N.; Gofur, A. Does Propolis Extract Alleviate Male Reproductive Performance Through Gonadotropic Hormone Levels and Sperm Quality? IOP Conf. Ser. Earth Environ. Sci. 2019, 276, 012056. [Google Scholar] [CrossRef]
- Seven, I.; Tatli Seven, P.; Gul Baykalir, B.; Parlak Ak, T.; Ozer Kaya, S.; Yaman, M. Bee glue (propolis) improves reproductive organs, sperm quality and histological changes and antioxidant parameters of testis tissues in rats exposed to excess copper. Andrologia 2020, 52, e13540. [Google Scholar] [CrossRef]
- Cedikova, M.; Miklikova, M.; Stachova, L.; Grundmanova, M.; Tuma, Z.; Vetvicka, V.; Zech, N.; Kralickova, M.; Kuncova, J. Effects of the Czech propolis on sperm mitochondrial function. Evid. Based Complement. Altern. Med. 2014, 2014, 248768. [Google Scholar] [CrossRef]
- Al-Nawab, N.; Al-Dujaily, S.S.; Al-Obaidi, Z.F.H. Effects of Propolis Extract Medium on In Vitro Sperms Activation of Infertile Asthenozoospermic Men. Ann. Rom. Soc. Cell Biol. 2021, 25, 5050–5059. [Google Scholar]
- Barbarić, M.; Mišković, K.; Bojić, M.; Lončar, M.B.; Smolčić-Bubalo, A.; Debeljak, Z.; Medić-Šarić, M. Chemical composition of the ethanolic propolis extracts and its effect on HeLa cells. J. Ethnopharmacol. 2011, 135, 772–778. [Google Scholar] [CrossRef] [PubMed]
- Blainski, A.; Lopes, G.C.; de Mello, J.C. Application and analysis of the folin ciocalteu method for the determination of the total phenolic content from Limonium brasiliense L. Molecules 2013, 18, 6852–6865. [Google Scholar] [CrossRef] [PubMed]
- Tvrdá, E.; Michalko, J.; Árvay, J.; Vukovic, N.L.; Ivanišová, E.; Ďuračka, M.; Matušíková, I.; Kačániová, M. Characterization of the Omija (Schisandra chinensis) Extract and Its Effects on the Bovine Sperm Vitality and Oxidative Profile during In Vitro Storage. Evid. Based Complement. Altern. Med. 2020, 2020, 7123780. [Google Scholar] [CrossRef]
- Willett, W.C. Balancing life-style and genomics research for disease prevention. Science 2002, 296, 695–698. [Google Scholar] [CrossRef] [PubMed]
- Sánchés-Moreno, C.; Larrauri, A.; Saura-Calixto, F. A procedure to measure the antioxidant efficiency of polyphenols. J. Sci. Food Agric. 1998, 76, 270–276. [Google Scholar] [CrossRef]
- Benko, F.; Mohammadi-Sangcheshmeh, A.; Ďuračka, M.; Lukáč, N.; Tvrdá, E. In vitro versus cryo-induced capacitation of bovine spermatozoa, part 1: Structural, functional, and oxidative similarities and differences. PLoS ONE 2022, 17, e0276683. [Google Scholar] [CrossRef]
- Galimov, S.N.; Gromenko, J.Y.; Bulygin, K.V.; Galimov, K.S.; Galimova, E.F.; Sinelnikov, M.Y. (2021). The level of secondary messengers and the redox state of NAD+/NADH are associated with sperm quality in infertility. J. Reprod. Immunol. 2021, 148, 103383. [Google Scholar] [CrossRef]
- Ahmadi, S.; Bashiri, R.; Ghadiri-Anari, A.; Nadjarzadeh, A. Antioxidant supplements and semen parameters: An evidence based review. Int. J. Reprod. Biomed. 2016, 14, 729–736. [Google Scholar] [CrossRef]
- Tvrdá, E.; Benko, F.; Slanina, T.; du Plessis, S.S. The Role of Selected Natural Biomolecules in Sperm Production and Functionality. Molecules 2021, 26, 5196. [Google Scholar] [CrossRef] [PubMed]
- Seven, P.T.; Seven, I.; Baykalir, B.G.; Mutlu, S.I.; Salem, A.Z.M. Nanotechnology and nano-propolis in animal production and health: An overview. Ital. J. Anim. Sci. 2018, 17, 921–930. [Google Scholar] [CrossRef]
- Kocot, J.; Kiełczykowska, M.; Luchowska-Kocot, D.; Kurzepa, J.; Musik, I. Antioxidant Potential of Propolis, Bee Pollen, and Royal Jelly: Possible Medical Application. Oxid. Med. Cell. Longev. 2018, 2018, 7074209. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, M.E.E.; Alqurashi, R.M. Anti-fungal and antioxidant properties of propolis (bee glue) extracts. Int. J. Food Microbiol. 2022, 361, 109463. [Google Scholar] [CrossRef] [PubMed]
- Kurek-Górecka, A.; Keskin, Ş.; Bobis, O.; Felitti, R.; Górecki, M.; Otręba, M.; Stojko, J.; Olczyk, P.; Kolayli, S.; Rzepecka-Stojko, A. Comparison of the Antioxidant Activity of Propolis Samples from Different Geographical Regions. Plants 2022, 11, 1203. [Google Scholar] [CrossRef]
- Romero, M.; Freire, J.; Pastene, E.; García, A.; Aranda, M.; Gonzáles, C. Propolis polyphenolic compounds affect the viability and structure of Helicobacter pylori in vitro. Rev. Bras. Farmacogn. 2019, 29, 325–332. [Google Scholar] [CrossRef]
- Kumazawa, S.; Hamasaka, T.; Nakayama, T. Antioxidant activity of propolis of various geographic origins. Food Chem. 2004, 84, 329–339. [Google Scholar] [CrossRef]
- Betances-Salcedo, E.; Revilla, I.; Vivar-Quintana, A.M.; González-Martín, M.I. Flavonoid and Antioxidant Capacity of Propolis Prediction Using Near Infrared Spectroscopy. Sensors 2017, 17, 1647. [Google Scholar] [CrossRef]
- Galeotti, F.; Maccari, F.; Fachini, A.; Volpi, N. Chemical Composition and Antioxidant Activity of Propolis Prepared in Different Forms and in Different Solvents Useful for Finished Products. Foods 2018, 7, 41. [Google Scholar] [CrossRef]
- Volpi, N.; Bergonzini, G. Analysis of flavonoids from propolis by on-line HPLC-electrospray mass spectrometry. J. Pharm. Biomed. Anal. 2006, 42, 354–361. [Google Scholar] [CrossRef]
- Rosli, N.L.; Roslan, H.R.; Omar, E.A.; Mokhtar, N.; Abdul Hapit, N.H.; Asem, N. Phytochemical Analysis and Antioxidant Activities of Trigona Apicalis Propolis Extract. IOP Conf. Proc. 2016, 1791, 010001. [Google Scholar]
- Shehata, M.G.; Ahmad, F.T.; Badr, A.N.; Masry, S.H.; El-Sohaimy, S.A. Chemical analysis, antioxidant, cytotoxic and antimicrobial properties of propolis from different geographic regions. Ann. Agric. Sci. 2020, 65, 209–217. [Google Scholar] [CrossRef]
- Piomboni, P.; Focarelli, R.; Stendardi, A.; Ferramosca, A.; Zara, V. The role of mitochondria in energy production for human sperm motility. Int. J. Androl. 2012, 35, 109–124. [Google Scholar] [CrossRef] [PubMed]
- Murphy, M.P. How mitochondria produce reactive oxygen species. Biochem. J. 2009, 417, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Cadenas, E. Mitochondrial free radical production and cell signaling. Mol. Asp. Med. 2004, 25, 17–26. [Google Scholar] [CrossRef] [PubMed]
- Das, S.; Das, J.; Samadder, A.; Boujedaini, N.; Khuda-Bukhsh, A.R. Apigenin-induced apoptosis in A375 and A549 cells through selective action and dysfunction of mitochondria. Exp. Biol. Med. 2012, 237, 1433–1448. [Google Scholar] [CrossRef]
- Cimen, H.; Han, M.J.; Yang, Y.; Tong, Q.; Koc, H.; Koc, E.C. Regulation of succinate dehydrogenase activity by SIRT3 in mammalian mitochondria. Biochemistry 2010, 49, 304–311. [Google Scholar] [CrossRef]
- Silva Santos, L.F.; Stolfo, A.; Calloni, C.; Salvador, M. Catechin and epicatechin reduce mitochondrial dysfunction and oxidative stress induced by amiodarone in human lung fibroblasts. J. Arrhythm. 2017, 33, 220–225. [Google Scholar] [CrossRef]
- Parihar, P.; Jat, D.; Ghafourifar, P.; Parihar, M.S. Efficiency of mitochondrially targeted gallic acid in reducing brain mitochondrial oxidative damage. Cell. Mol. Biol. 2014, 60, 35–41. [Google Scholar]
- Semaming, Y.; Sripetchwandee, J.; Sa-Nguanmoo, P.; Pintana, H.; Pannangpetch, P.; Chattipakorn, N.; Chattipakorn, S.C. Protocatechuic acid protects brain mitochondrial function in streptozotocin-induced diabetic rats. Appl. Physiol. Nutr. Metab. 2015, 40, 1078–1081. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).