


The Role of Lignin Structure on Cellulase Adsorption and Enzymatic Hydrolysis

 ,
,  ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
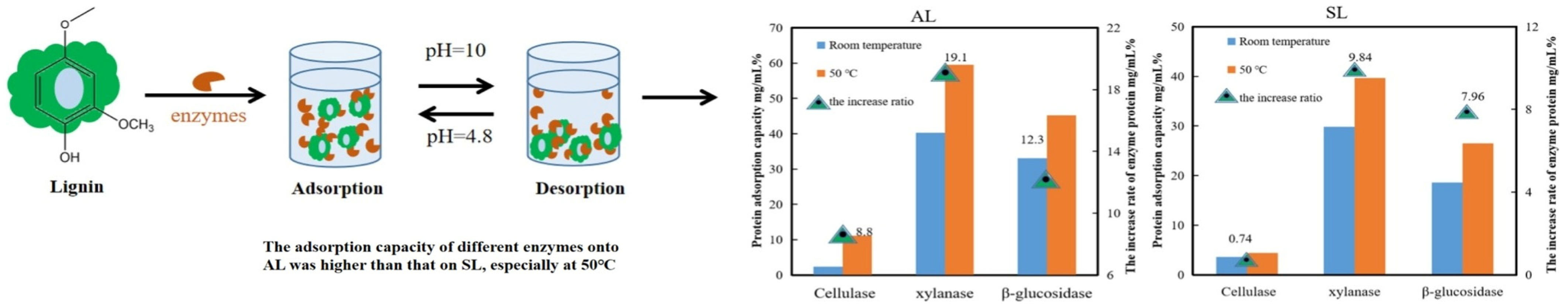{kind=link}
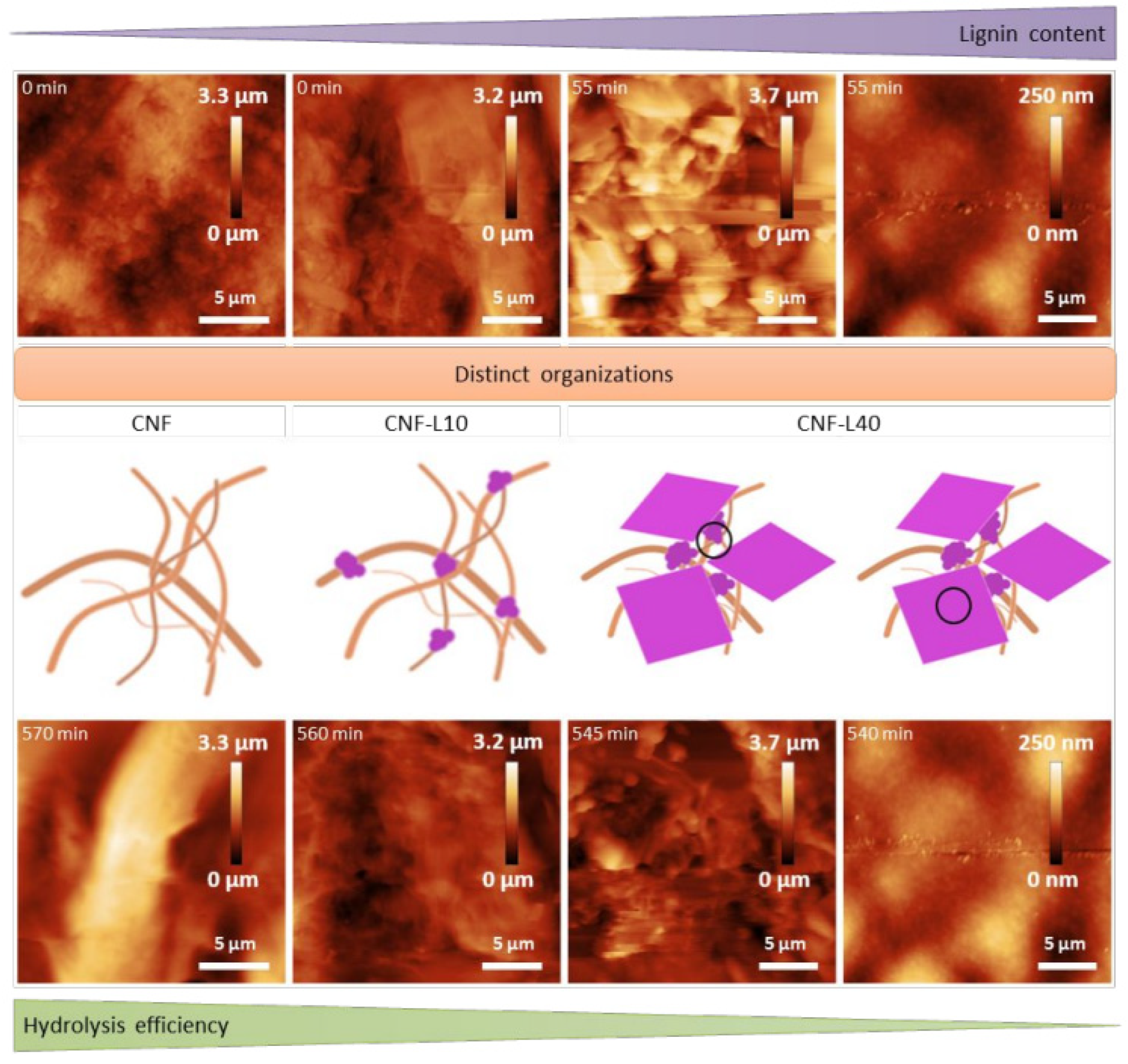{kind=link}
Abstract
1. Introduction
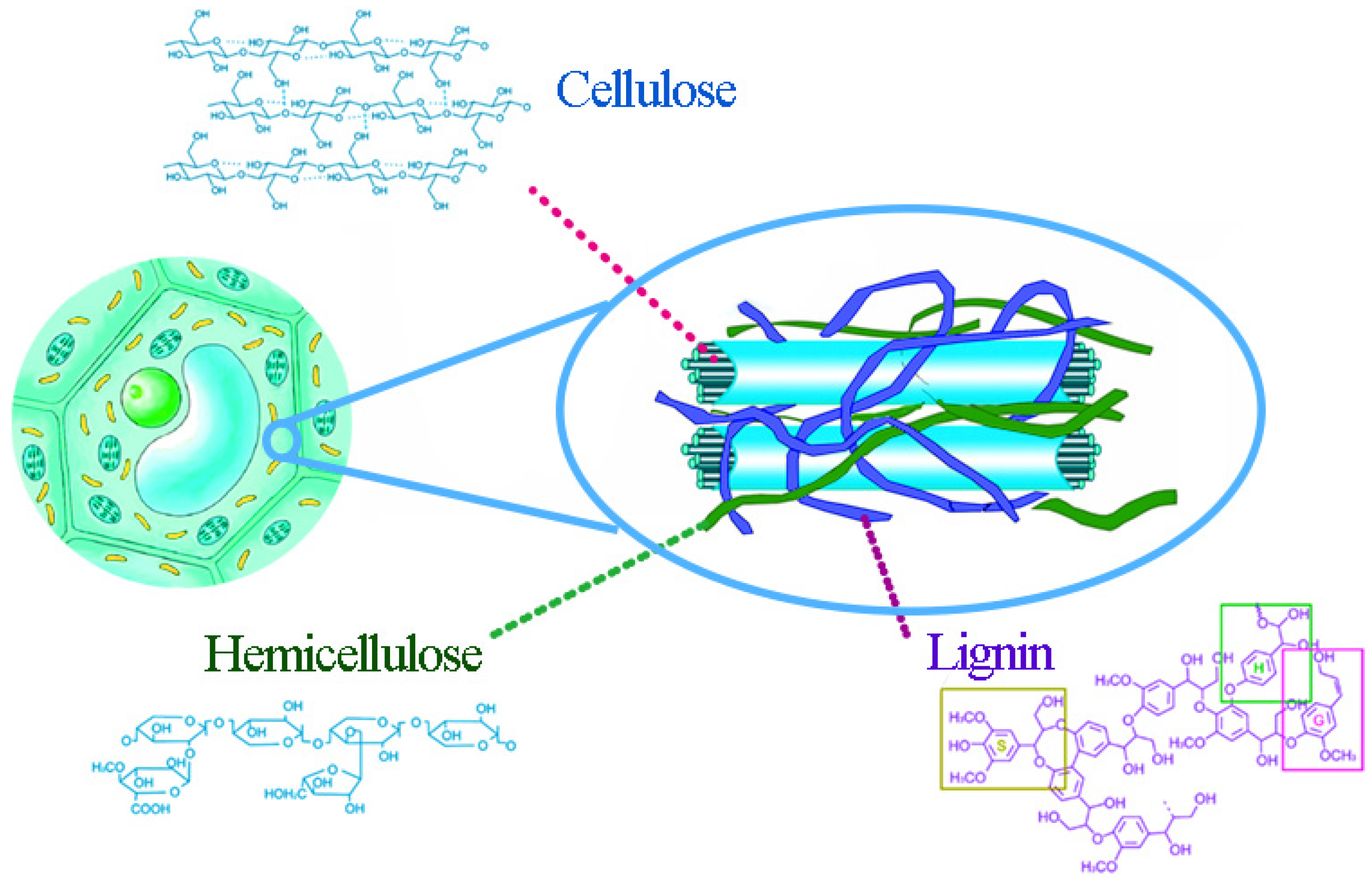
2. Lignocellulose Raw Materials
3. Efficient Saccharification of Lignocellulosic Materials
3.1. Pretreatment of Lignocellulosic Materials
3.2. Cellulase Hydrolysis
4. Factors Influencing the Hydrolysis of Cellulase
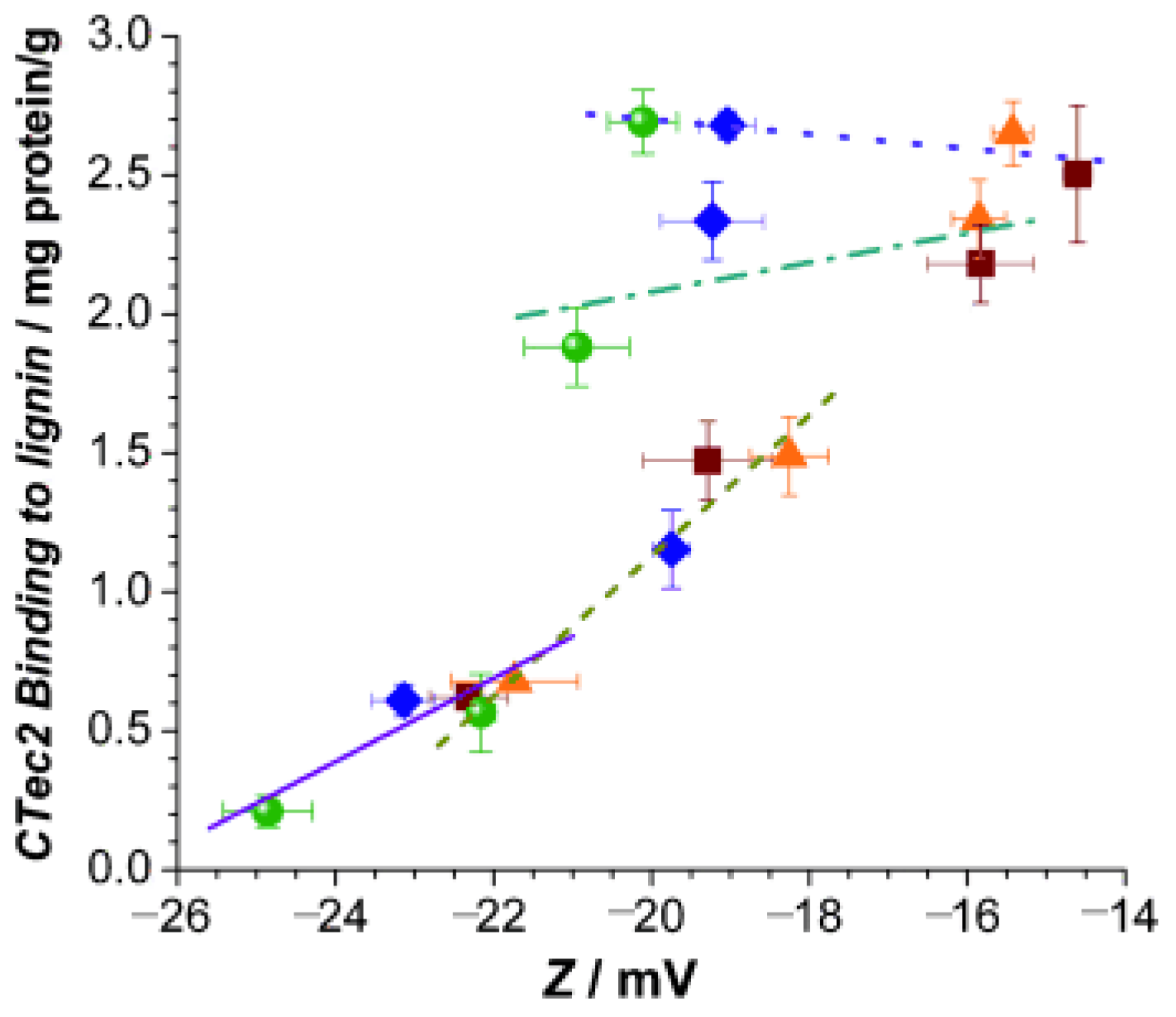
4.1. Mechanism of Lignin–Enzyme Interactions
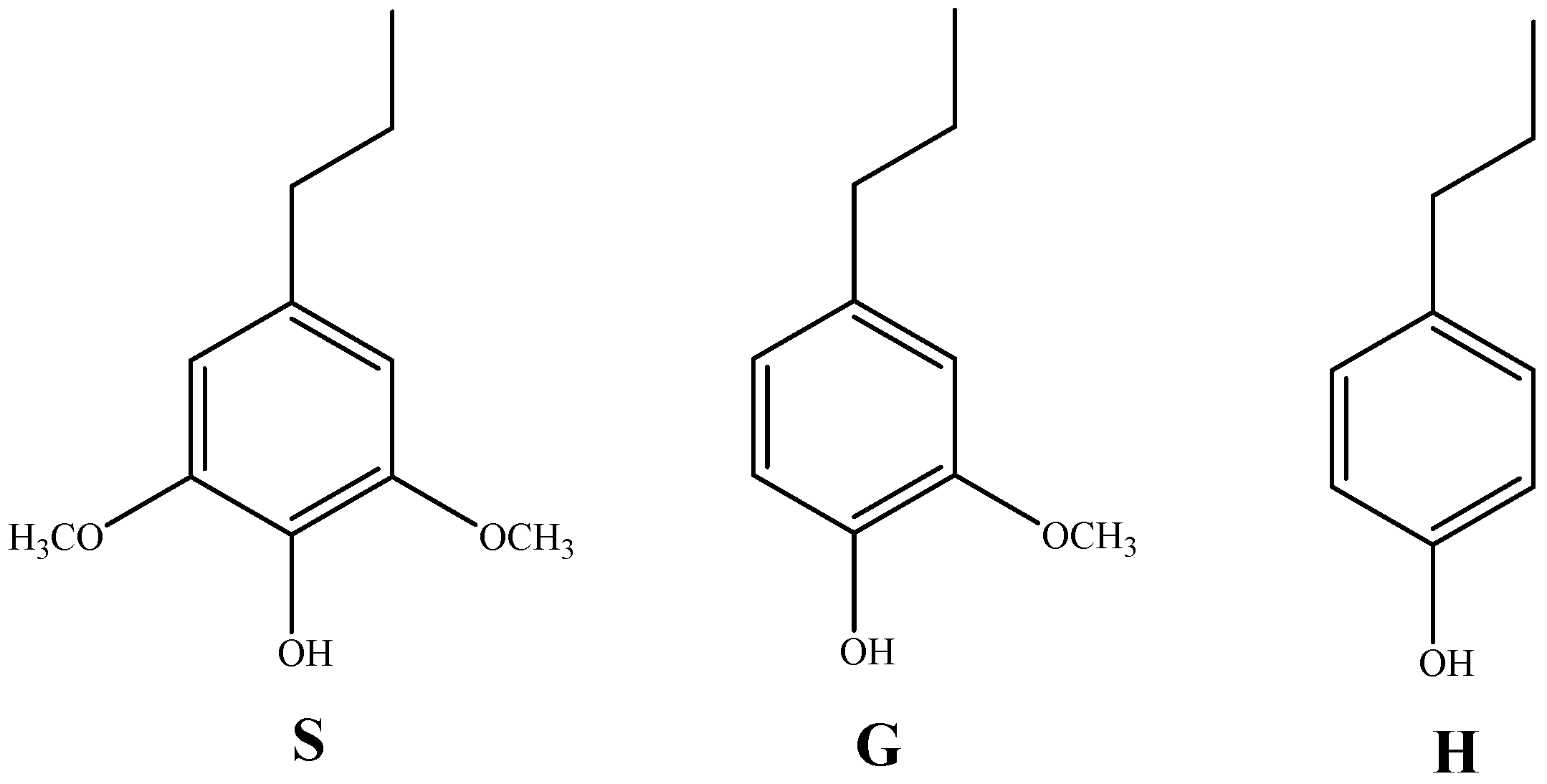
4.2. Effect of Lignin Functional Groups on Cellulase Hydrolysis
5. Conclusions and Outlook
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Devi, A.; Singh, A.; Bajar, S.; Pant, D.; Din, Z.U. Ethanol from lignocellulosic biomass: An in-depth analysis of pre-treatment methods, fermentation approaches and detoxification processes. J. Environ. Chem. Eng. 2021, 9, 105798. [Google Scholar] [CrossRef]
- Göncü, B.; Gülşen, H.; Hoşgün, E.Z. Bioethanol production from pistachio (Pistacia vera L.) shells applying ozone pretreatment and subsequent enzymatic hydrolysis. Environ. Technol. 2021, 42, 2438–2446. [Google Scholar] [CrossRef] [PubMed]
- Sun, S.N.; Sun, S.L.; Cao, X.F.; Sun, R.C. The role of pretreatment in improving the enzymatic hydrolysis of lignocellulosic materials. Bioresour. Technol. 2016, 199, 49–58. [Google Scholar] [CrossRef] [PubMed]
- Agrawal, R.; Verma, A.; Singhania, R.R.; Varjani, S.; Di Dong, C.; Patel, A.K. Current understanding of the inhibition factors and their mechanism of action for the lignocellulosic biomass hydrolysis. Bioresour. Technol. 2021, 332, 125042. [Google Scholar] [CrossRef]
- Hao, X.; Xu, F.; Zhang, J. Effect of pretreatments on production of xylooligosaccharides and monosaccharides from corncob by a two-step hydrolysis. Carbohydr. Polym. 2022, 285, 119217. [Google Scholar] [CrossRef]
- Lynd, L.R.; Weimer, P.J.; van Zyl, W.H.; Pretorius, I.S. Microbial cellulose utilization: Fundamentals and biotechnolog. Microbiol. Mol. Biol. R 2002, 66, 506–577. [Google Scholar] [CrossRef]
- Hasanov, I.; Raud, M.; Kikas, T. The Role of Ionic Liquids in the Lignin Separation from Lignocellulosic Biomass. Energies 2020, 13, 4864. [Google Scholar] [CrossRef]
- Liu, K.; Xu, T.; Du, H.S.; Zheng, T.; Liu, H.Y.; Zhang, M.; Liu, W.; Zhang, R.; Li, H.M.; Si, C.L. Lignin-based electrodes for energy storage application. Carbohyd. Polym. 2021, 259, 117740. [Google Scholar] [CrossRef]
- Sun, L.; Han, J.; Wu, J.; Huang, W.; Li, Y.; Mao, Y.; Wang, L.; Wang, Y. Cellulose pretreatment with inorganic salt hydrate: Dissolution, regeneration, structure and morphology. Ind. Crop. Prod. 2022, 180, 114722. [Google Scholar] [CrossRef]
- Chen, X.; Xiong, L.; Li, H.; Zhang, L.; Yuan, G.; Chen, X.; Wang, C.; Chen, X. The inhibitory effect of xylan on enzymatic hydrolysis of cellulose is dependent on cellulose ultrastructure. Cellulose 2020, 27, 4417–4428. [Google Scholar] [CrossRef]
- Yu, H.; Wang, J.; Yu, J.-X.; Wang, Y.; Chi, R.-A. Adsorption performance and stability of the modified straws and their extracts of cellulose, lignin, and hemicellulose for Pb2+: pH effect. Arab. J. Chem. 2020, 13, 9019–9033. [Google Scholar] [CrossRef]
- Khan, M.U.; Ahring, B.K. Lignin degradation under anaerobic digestion: Influence of lignin modifications—A review. Biomass-Bioenergy 2019, 128, 105325. [Google Scholar] [CrossRef]
- Schutyser, W.; Renders, T.; Van Den Bosch, S.; Koelewijn, S.-F.; Beckham, G.T.; Sels, B.F. Chemicals from lignin: An interplay of lignocellulose fractionation, depolymerisation, and upgrading. Chem. Soc. Rev. 2018, 47, 852–908. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, A.; Rana, M.; Park, J.-H. Advancement in technologies for the depolymerization of lignin. Fuel Process. Technol. 2018, 181, 115–132. [Google Scholar] [CrossRef]
- Zhu, Y.; Huang, J.; Wang, K.; Wang, B.; Sun, S.; Lin, X.; Song, L.; Wu, A.; Li, H. Characterization of Lignin Structures in Phyllostachys edulis (Moso Bamboo) at Different Ages. Polymers 2020, 12, 187. [Google Scholar] [CrossRef] [PubMed]
- Sheng, Y.Q.; Shiung, L.S.; Wu, Y.J.; Ge, S.B.; Wu, J.L.; Cai, L.P.; Huang, Z.H.; van Le, Q.; Sonne, C.; Xia, C.L. Simultaneous removal of nitrate and sulfate using an up-flow three-dimensional biofilm electrode reactor: Performance and microbial response. Bioresour. Technol. 2020, 324, 124631. [Google Scholar] [CrossRef]
- Cai, J.; Wang, Y.; Liu, J.; Zhang, X.; Li, F. Pretreatment enhanced structural disruption, enzymatic hydrolysis, fermentative hydrogen production from rice straw. Int. J. Hydrogen Energy 2022, 47, 11778–11786. [Google Scholar] [CrossRef]
- Li, P.; Yang, C.; Jiang, Z.; Jin, Y.; Wu, W. Lignocellulose pretreatment by deep eutectic solvents and related technologies: A review. J. Bioresour. Bioprod. 2023, 8, 33–44. [Google Scholar] [CrossRef]
- Sun, L.L.; Yue, Z.; Sun, S.C.; Sun, S.N.; Yuan, X.F.; Tong, Q.; Jia, L. Exploration of deep eutectic solvent-based biphasic system for furfural production and enhancing enzymatic hydrolysis: Chemical, topochemical, and morphological changes. Bioresour. Technol. 2022, 352, 127074. [Google Scholar] [CrossRef]
- Chandra, R.P.; Bura, R.; Mabee, W.E.; Berlin, A.; Pan, X.; Saddler, J.N. Substrate pretreatment: The key to effective enzymatic hydrolysis of lignocellulosics? Biofuels 2007, 108, 67–93. [Google Scholar]
- Wu, Y.; Ge, S.; Xia, C.; Mei, C.; Kim, K.-H.; Cai, L.; Smith, L.M.; Lee, J.; Shi, S.Q. Application of intermittent ball milling to enzymatic hydrolysis for efficient conversion of lignocellulosic biomass into glucose. Renew. Sustain. Energy Rev. 2021, 136, 110442. [Google Scholar] [CrossRef]
- Silva, G.G.D.; Couturier, M.; Berrin, J.-G.; Buléon, A.; Rouau, X. Effects of grinding processes on enzymatic degradation of wheat straw. Bioresour. Technol. 2012, 103, 192–200. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Deng, Y.; Li, S.; Xu, W.; Liu, G. Enhanced efficiency of enzymatic hydrolysis of wheat straw via freeze–thaw pretreatment. Environ. Sci. Pollut. Res. 2022, 29, 56696–56704. [Google Scholar] [CrossRef]
- Santos, V.T.D.O.; Siqueira, G.; Milagres, A.M.F.; Ferraz, A. Role of hemicellulose removal during dilute acid pretreatment on the cellulose accessibility and enzymatic hydrolysis of compositionally diverse sugarcane hybrids. Ind. Crop. Prod. 2018, 111, 722–730. [Google Scholar] [CrossRef]
- Zhu, J.Y.; Pan, X.J.; Wang, G.S.; Gleisner, R. Sulfite pretreatment (SPORL) for robust enzymatic saccharification of spruce and red pine. Bioresour. Technol. 2009, 100, 2411–2418. [Google Scholar] [CrossRef]
- Krishnan, C.; Sousa, L.D.C.; Jin, M.; Chang, L.; Dale, B.E.; Balan, V. Alkali-based AFEX pretreatment for the conversion of sugarcane bagasse and cane leaf residues to ethanol. Biotechnol. Bioeng. 2010, 107, 441–450. [Google Scholar] [CrossRef]
- Souto, L.R.F.; da Silva, I.F.; Ninow, J.L.; Collins, S.R.; Elliston, A.; Waldron, K.W. Effect of hydrothermal pre-treatment on duckweed (Landoltia punctata) biomass for simultaneous saccharification and fermentation process. Biomass-Bioenergy 2019, 127, 105259. [Google Scholar] [CrossRef]
- Saha, B.C.; Qureshi, N.; Kennedy, G.J.; Cotta, M.A. Biological pretreatment of corn stover with white-rot fungus for improved enzymatic hydrolysis. Int. Biodeterior. Biodegrad. 2016, 109, 29–35. [Google Scholar] [CrossRef]
- da Silva, M.A.; Ferraz, A. Biological pretreatment of sugarcane bagasse with basidiomycetes producing varied patterns of biodegradation. Bioresour. Technol. 2017, 225, 17–22. [Google Scholar]
- Asgher, M.; Ahmad, Z.; Iqbal, H.M.N. Alkali and enzymatic delignification of sugarcane bagasse to expose cellulose polymers for saccharification and bio-ethanol production. Ind. Crop. Prod. 2013, 44, 488–495. [Google Scholar] [CrossRef]
- Ni, J.; Wang, H.; Chen, Y.; She, Z.; Na, H.; Zhu, J. A novel facile two-step method for producing glucose from cellulose. Bioresour. Technol. 2013, 137, 106–110. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.; Zhang, R.H.; Pan, Z.L. Investigation of adsorption kinetics and isotherm of cellulase and beta-glucosidase on lignocellulosic substrates. Biomass. Bioenerg. 2016, 91, 1–9. [Google Scholar] [CrossRef]
- Yuan, Y.; Jiang, B.; Chen, H.; Wu, W.; Wu, S.; Jin, Y.; Xiao, H. Recent advances in understanding the effects of lignin structural characteristics on enzymatic hydrolysis. Biotechnol. Biofuels 2021, 14, 205. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Chi, C.; Gong, Y.; Li, H.; Wu, Y. Research progress in the fractal kinetics of lignocellulose enzymatic hydrolysis. China Pulp Pap. 2015, 34, 62–67. [Google Scholar]
- Bhat, S.; Owen, E.; Bhat, M.K. Isolation and Characterisation of a Major Cellobiohydrolase (S8) and a Major Endoglucanase (S11) Subunit from the Cellulosome of Clostridium thermocellum. Anaerobe 2001, 7, 171–179. [Google Scholar] [CrossRef]
- Ooshima, H.; Sakata, M.; Harano, Y. Adsorption of cellulase fromTrichoderma viride on cellulose. Biotechnol. Bioeng. 1983, 25, 3103–3114. [Google Scholar] [CrossRef]
- Kinnarinen, T.; Häkkinen, A. Influence of enzyme loading on enzymatic hydrolysis of cardboard waste and size distribution of the resulting fiber residue. Bioresour. Technol. 2014, 159, 136–142. [Google Scholar] [CrossRef]
- Du, R.Y.; Su, R.X.; Li, X.; Tantai, X.W.; Liu, Z.H.; Yang, J.F.; Qi, W.; He, Z.M. Controlled adsorption of cellulase onto pretreated corncob by pH adjustment. Cellulose 2012, 19, 371–380. [Google Scholar] [CrossRef]
- Gabhane, J.; Kumar, S.; Sarma, A. Effect of glycerol thermal and hydrothermal pretreatments on lignin degradation and enzymatic hydrolysis in paddy straw. Renew. Energy 2020, 154, 1304–1313. [Google Scholar] [CrossRef]
- Hou, X.-D.; Li, N.; Zong, M.-H. Significantly enhancing enzymatic hydrolysis of rice straw after pretreatment using renewable ionic liquid–water mixtures. Bioresour. Technol. 2013, 136, 469–474. [Google Scholar] [CrossRef]
- Li, C.; Cheng, G.; Balan, V.; Kent, M.S.; Ong, M.; Chundawat, S.P.; Sousa, L.D.; Melnichenko, Y.B.; Dale, B.E.; Simmons, B.A.; et al. Influence of physico-chemical changes on enzymatic digestibility of ionic liquid and AFEX pretreated corn stover. Bioresour. Technol. 2011, 102, 6928–6936. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Zhang, L.; Liu, D. Biomass recalcitrance. Part II: Fundamentals of different pre-treatments to increase the enzymatic digestibility of lignocellulose. Biofuels Bioprod. Biorefining 2012, 6, 561–579. [Google Scholar] [CrossRef]
- Chundawat, S.P.S.; Bellesia, G.; Uppugundla, N.; Sousa, L.D.C.; Gao, D.; Cheh, A.M.; Agarwal, U.P.; Bianchetti, C.M.; Phillips, J.G.N.; Langan, P.; et al. Restructuring the Crystalline Cellulose Hydrogen Bond Network Enhances Its Depolymerization Rate. J. Am. Chem. Soc. 2011, 133, 11163–11174. [Google Scholar] [CrossRef] [PubMed]
- Cheng, L.; Hu, X.; Gu, Z.; Hong, Y.; Li, Z.; Li, C. Characterization of physicochemical properties of cellulose from potato pulp and their effects on enzymatic hydrolysis by cellulase. Int. J. Biol. Macromol. 2019, 131, 564–571. [Google Scholar] [CrossRef] [PubMed]
- Satari, B.; Karimi, K.; Molaverdi, M. Structural features influential to enzymatic hydrolysis of cellulose-solvent-based pretreated pinewood and elmwood for ethanol production. Bioprocess Biosyst. Eng. 2017, 41, 249–264. [Google Scholar] [CrossRef]
- Zhang, H.; Wu, S.; Xie, J. Evaluation of the effects of isolated lignin on enzymatic hydrolysis of cellulose. Enzym. Microb. Technol. 2017, 101, 44–50. [Google Scholar] [CrossRef]
- Saini, J.K.; Patel, A.K.; Adsul, M.; Singhania, R.R. Cellulase adsorption on lignin: A roadblock for economic hydrolysis of biomass. Renew. Energy 2016, 98, 29–42. [Google Scholar] [CrossRef]
- Pareek, N.; Gillgren, T.; Jönsson, L.J. Adsorption of proteins involved in hydrolysis of lignocellulose on lignins and hemicelluloses. Bioresour. Technol. 2013, 148, 70–77. [Google Scholar] [CrossRef]
- Costaouëc, T.L.; Pakarinen, A.; Várnai, A.; Puranen, T.; Viikari, L. The role of carbohydrate binding module (CBM) at high substrate consistency: Comparison of Trichoderma reesei and Thermoascus aurantiacus Cel7A (CBHI) and Cel5A (EGII). Bioresour. Technol. 2013, 143, 196–203. [Google Scholar] [CrossRef]
- Hodgson, K.T.; Berg, J.C. The effect of surfactants on wicking flow in fiber networks. J. Colloid Interface Sci. 1988, 121, 22–31. [Google Scholar] [CrossRef]
- Lu, X.; Zheng, X.; Li, X.; Zhao, J. Adsorption and mechanism of cellulase enzymes onto lignin isolated from corn stover pretreated with liquid hot water. Biotechnol. Biofuels 2016, 9, 118. [Google Scholar] [CrossRef] [PubMed]
- Lou, H.M.; Zhu, J.Y.; Lan, T.Q.; Lai, H.R.; Qiu, X.Q. pH-induced lignin surface modification to reduce nonspecific cellulase binding and enhance enzymatic saccharification of lignocelluloses. ChemSusChem 2013, 6, 919–927. [Google Scholar] [CrossRef] [PubMed]
- Fritz, C.; Ferrer, A.; Salas, C.; Jameel, H.; Rojas, O.J. Interactions between Cellulolytic Enzymes with Native, Autohydrolysis, and Technical Lignins and the Effect of a Polysorbate Amphiphile in Reducing Nonproductive Binding. Biomacromolecules 2015, 16, 3878–3888. [Google Scholar] [CrossRef] [PubMed]
- Mou, H.; Wu, X.; Huang, J.; Liu, Y.; Fan, H. Eucalyptus lignin modification for dynamic adsorption with lignocellulose-degradation enzymes dependent on pH values. Ind. Crop. Prod. 2021, 169, 113650. [Google Scholar] [CrossRef]
- Zhang, Y.Q.; Xu, X.M.; Zhang, Y.Y.; Li, J.F. Effect of adding surfactant for transforming lignocellulose into fermentable sugars during biocatalysing. Biotechnol. Bioproc. E 2011, 16, 930–936. [Google Scholar] [CrossRef]
- Djajadi, D.T.; Jensen, M.M.; Oliveira, M.; Jensen, A.; Thygesen, L.G.; Pinelo, M.; Glasius, M.; Jørgensen, H.; Meyer, A.S. Lignin from hydrothermally pretreated grass biomass retards enzymatic cellulose degradation by acting as a physical barrier rather than by inducing nonproductive adsorption of enzymes. Biotechnol. Biofuels 2018, 11, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Donaldson, L.A.; Wong, K.K.Y.; Mackie, K.L. Ultrastructure of steam-exploded wood. Wood Sci. Technol. 1988, 22, 103–114. [Google Scholar] [CrossRef]
- Lambert, E.; Aguié-Béghin, V.; Dessaint, D.; Foulon, L.; Chabbert, B.; Paës, G.; Molinari, M. Real Time and Quantitative Imaging of Lignocellulosic Films Hydrolysis by Atomic Force Microscopy Reveals Lignin Recalcitrance at Nanoscale. Biomacromolecules 2018, 20, 515–527. [Google Scholar] [CrossRef]
- Huang, C.; He, J.; Min, D.; Lai, C.; Yong, Q. Understanding the Nonproductive Enzyme Adsorption and Physicochemical Properties of Residual Lignins in Moso Bamboo Pretreated with Sulfuric Acid and Kraft Pulping. Appl. Biochem. Biotechnol. 2016, 180, 1508–1523. [Google Scholar] [CrossRef]
- Yang, Q.; Pan, X. Correlation between lignin physicochemical properties and inhibition to enzymatic hydrolysis of cellulose. Biotechnol. Bioeng. 2015, 113, 1213–1224. [Google Scholar] [CrossRef]
- Pan, X. Role of Functional Groups in Lignin Inhibition of Enzymatic Hydrolysis of Cellulose to Glucose. J. Biobased Mater. Bioenergy 2008, 2, 25–32. [Google Scholar] [CrossRef]
- Palonen, H.; Viikari, L. Role of oxidative enzymatic treatments on enzymatic hydrolysis of softwood. Biotechnol. Bioeng. 2004, 86, 550–557. [Google Scholar] [CrossRef]
- Ying, W.; Shi, Z.; Yang, H.; Xu, G.; Zheng, Z.; Yang, J. Effect of alkaline lignin modification on cellulase–lignin interactions and enzymatic saccharification yield. Biotechnol. Biofuels 2018, 11, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.S.; Wang, W.X.; Wang, Y.; Jin, Y.C. Complete Dissolution of Ball-Milled Masson Pine Using an Aqueous Sodium Hydroxide Solvent. Bioresources 2016, 10, 7361–7371. [Google Scholar] [CrossRef]
- Wu, J.; Chandra, R.P.; Takada, M.; Liu, L.-Y.; Renneckar, S.; Kim, K.H.; Kim, C.S.; Saddler, J.N. Enhancing Enzyme-Mediated Cellulose Hydrolysis by Incorporating Acid Groups onto the Lignin during Biomass Pretreatment. Front. Bioeng. Biotechnol. 2020, 8, 608835. [Google Scholar] [CrossRef]
- Zheng, W.; Lan, T.; Li, H.; Yue, G.; Zhou, H. Exploring why sodium lignosulfonate influenced enzymatic hydrolysis efficiency of cellulose from the perspective of substrate–enzyme adsorption. Biotechnol. Biofuels 2020, 13, 1–12. [Google Scholar] [CrossRef]
- Del Rio, L.F.; Chandra, R.P.; Saddler, J.N. The effects of increasing swelling and anionic charges on the enzymatic hydrolysis of organosolv-pretreated softwoods at low enzyme loadings. Biotechnol. Bioeng. 2011, 108, 1549–1558. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, W.; Li, P.; Huang, L.; Wei, Y.; Li, J.; Zhang, L.; Jin, Y. The Role of Lignin Structure on Cellulase Adsorption and Enzymatic Hydrolysis. Biomass 2023, 3, 96-107. https://doi.org/10.3390/biomass3010007
Wu W, Li P, Huang L, Wei Y, Li J, Zhang L, Jin Y. The Role of Lignin Structure on Cellulase Adsorption and Enzymatic Hydrolysis. Biomass. 2023; 3(1):96-107. https://doi.org/10.3390/biomass3010007
Chicago/Turabian StyleWu, Wenjuan, Penghui Li, Lijing Huang, Yumeng Wei, Jiaquan Li, Lu Zhang, and Yongcan Jin. 2023. "The Role of Lignin Structure on Cellulase Adsorption and Enzymatic Hydrolysis" Biomass 3, no. 1: 96-107. https://doi.org/10.3390/biomass3010007
APA StyleWu, W., Li, P., Huang, L., Wei, Y., Li, J., Zhang, L., & Jin, Y. (2023). The Role of Lignin Structure on Cellulase Adsorption and Enzymatic Hydrolysis. Biomass, 3(1), 96-107. https://doi.org/10.3390/biomass3010007