


Towards Sustainable Maize Production: Understanding the Morpho-Physiological, Genetics, and Molecular Mechanisms for Tolerance to Low Soil Nitrogen, Phosphorus, and Potassium



Abstract
1. Introduction
2. Maize Morphology and Physiology for Tolerance to Low Nitrogen (N), Phosphorus (P), Potassium (K)
2.1. Maize Shoot Morphology and Physiology for Tolerance to Low Nitrogen (N)
2.2. Morphological and Physiological Mechanisms Employed by the Maize Shoot System during Phosphorus (P) Deficiency
2.3. Morphological and Physiological Mechanisms of Maize Growth under Low Potassium (K) Conditions
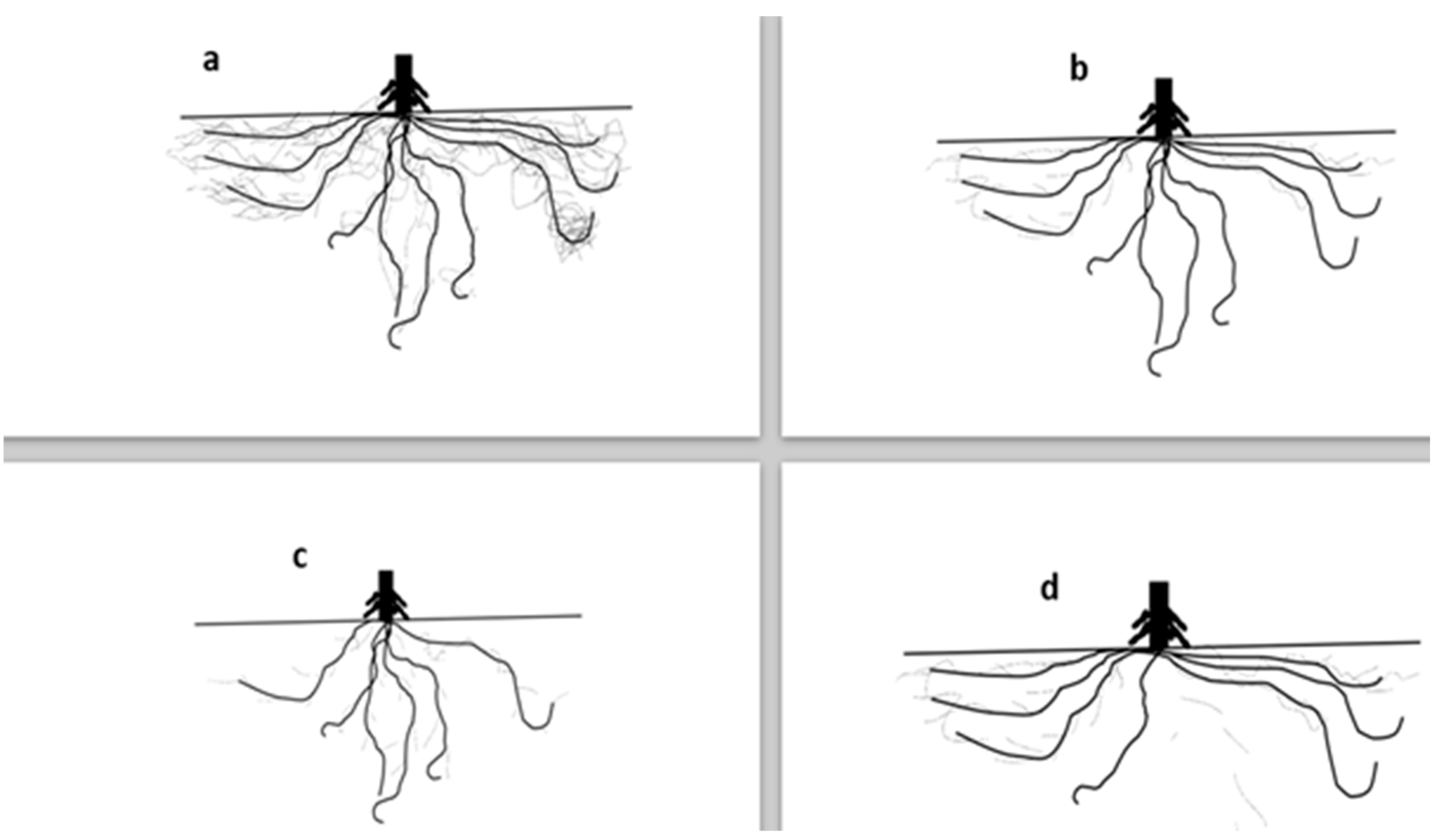
2.4. Root Morphology and Physiology for Tolerance to Low Nitrogen (N), Phosphorus (P), Potassium (K)
3. Genetics and Genomic Components of Maize for Tolerance to Low Soil Nitrogen (N), Phosphorus (P), and Potassium (K)
4. Genes, Proteins, and Metabolites Associated with Maize Response to Low Nitrogen (N), Phosphorus (P), and Potassium (K)
5. Conclusions and the Way Forward
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Sandhu, N.; Sethi, M.; Kumar, A.; Dang, D.; Singh, J.; Chhuneja, P. Biochemical and genetic approaches improving nitrogen use efficiency in cereal crops: A review. Front. Plant Sci. 2021, 12, 657629. [Google Scholar] [CrossRef]
- Torres-Rodríguez, J.; Salazar-vidal, M.; Montes, R.; Massange-Sánchez, J.A.; Gillmor, C.; Sawers, R. Low nitrogen availability inhibits the phosphorus starvation response in maize (Zea mays ssp. mays L.). BMC Plant Biol. 2021, 5, 259. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Yan, L.; Zhang, H. Morphological and physiological responses of winter wheat seedlings to nitrogen and phosphorus deficiency. J. Plant Nutr. 2013, 36, 1234–1246. [Google Scholar] [CrossRef]
- Yang, W.; Yoon, J.; Choi, H.; Fan, Y.; Chen, R.; An, G. Transcriptome analysis of nitrogen-starvation- responsive genes in rice. BMC Plant Biol. 2015, 15, 31. [Google Scholar] [CrossRef]
- Ao, X.; Guo, X.H.; Zhu, Q.; Zhang, H.J.; Wang, H.Y.; Ma, Z.H.; Han, X.R.; Zhao, M.H.; Xie, F.T. Effect of phosphorus fertilization to P uptake and dry matter accumulation in soybean with different P efficiencies. J. Integr. Agric. 2014, 13, 326–334. [Google Scholar] [CrossRef]
- Roberts, T.L.; Johnston, A.E. Phosphorus use efficiency and management in agriculture. Resour. Conserv. Recycl. 2015, 105, 275–281. [Google Scholar] [CrossRef]
- Wen, Z.; Li, H.; Shen, J.; Rengel, Z. Maize responds to low shoot P concentration by altering root morphology rather than increasing root exudation. Plant Soil 2017, 416, 377–389. [Google Scholar] [CrossRef]
- Dhillon, J.; Torres, G.; Driver, E.; Figueiredo, B.; Raun, W.R. World phosphorus use efficiency in cereal crops. Agron. J. 2017, 109, 1670–1677. [Google Scholar] [CrossRef]
- Pereira, N.C.M.; Galindo, F.S.; Gazola, R.P.D.; Dupas, E.; Rosa, P.A.L.; Mortinho, E.S.; Filho, M.C.M.T. Corn yield and phosphorus use efficiency response to phosphorus rates associated with plant growth promoting bacteria. Front. Environ. Sci. 2020, 8, 40. [Google Scholar] [CrossRef]
- Pettigrew, W.T. Potassium influences on yield and quality production for maize, wheat, soybean and cotton. Physiol. Plant 2008, 133, 670–681. [Google Scholar] [CrossRef]
- Jordan-Meille, L.; Pellerin, S. Shoot and root growth of hydroponic maize (Zea mays L.) as influenced by K deficiency. Plant Soil 2008, 304, 157–168. [Google Scholar] [CrossRef]
- Demari, G.; Carvalho, I.; Nardino, M.; JSzareski, V.; Dellagostin, S.; da Rosa, T.; Follmann, N.; Monteiro, M.; Basso, C.; Pedó, T.; et al. Importance of nitrogen in maize production. Int. J. Curr. Res. 2016, 8, 36629–36634. [Google Scholar]
- Zhu, X.-K.; Li, C.-Y.; Jiang, Z.-Q.; Huang, L.-L.; Feng, C.-N.; Guo, W.-S.; Peng, Y.-X. Responses of phosphorus use efficiency, grain yield, and quality to phosphorus application amount of weak-gluten wheat. J. Integr. Agric. 2012, 11, 1103–1110. [Google Scholar] [CrossRef]
- Li, D.; Wang, H.; Wang, M.; Li, G.; Chen, Z.; Leiser, W.L.; Wei, T.M.; Lu, X.; Wang, M.; Chen, S.; et al. Genetic dissection of phosphorus use efficiency in a maize association population under two p levels in the field. Int. J. Mol. Sci. 2021, 22, 9311. [Google Scholar] [CrossRef]
- Ertiro, B.T.; Labuschagne, M.; Olsen, M.; Das, B.; Prasanna, B.M.; Gowda, M. Genetic dissection of nitrogen use efficiency in tropical maize through genome-wide association and genomic prediction. Front. Plant Sci. 2020, 11, 474. [Google Scholar] [CrossRef]
- Singh, P.; Kumar, K.; Jha, A.K.; Yadava, P.; Pal, M.; Rakshit, S.; Singh, I. Global gene expression profiling under nitrogen stress identifies key genes involved in nitrogen stress adaptation in maize (Zea mays L.). Sci. Rep. 2022, 12, 4211. [Google Scholar] [CrossRef]
- Mi, G.; Chen, F.; Zhang, F. Physiological and genetic mechanisms for nitrogen-use efficiency in maize. J. Crop Sci. Biotech. 2005, 10, 57–63. [Google Scholar]
- Hirel, B.; Bertin, P.; Quilleré, I.; Bourdoncle, W.; Attagnant, C.; Dellay, C.; Gouy, A.; Cadiou, S.; Retailliau, C.; Falque, M.; et al. Towards a better understanding of the genetic and physiological basis for nitrogen use efficiency in maize. Plant Physiol. 2001, 125, 1258–1270. [Google Scholar] [CrossRef]
- Schlüter, U.; Mascher, M.; Colmsee, C.; Scholz, U.; Bräutigam, A.; Holger, F.; Sonnewald, U. Maize source leaf adaptation to nitrogen deficiency affects not only nitrogen and carbon metabolism but also control of phosphate homeostasis. Plant Physiol. 2012, 160, 1384–1406. [Google Scholar] [CrossRef]
- Guo, T.; Wang, D.; Fang, J.; Zhao, J.; Yuan, S.; Xiao, L.; Li, X. Mutations in the rice OsCHR4 gene, encoding a CHD3 family chromatin remodeler, induce narrow and rolled leaves with increased cuticular wax. Int. J. Mol. Sci. 2019, 20, 2567. [Google Scholar] [CrossRef]
- Lu, C.; Zhang, J. Photosynthetic CO2 assimilation, chlorophyll fluorescence and photoinhibition as affected by nitrogen deficiency in maize plants. Plant Sci. 2000, 151, 135–143. [Google Scholar] [CrossRef]
- Martin, A.; Lee, J.; Kichey, T.; Gerentes, D.; Zivy, M.; Tatout, C.; Dubois, F.; Balliau, T.; Valot, B.; Davanture, M.; et al. Two cytosolic glutamine synthetase isoforms of maize are specifically involved in the control of grain production. Plant Cell 2006, 18, 3252–3274. [Google Scholar] [CrossRef] [PubMed]
- Uauy, C.; Brevis, J.C.; Dubcovsky, J. The high grain protein content gene Gpc-B1 accelerates senescence and has pleiotropic effects on protein content in wheat. J. Exp. Bot. 2006, 57, 2785–2794. [Google Scholar] [CrossRef]
- Masclaux-Daubresse, C.; Daniel-Vedele, F.; Dechorgnat, J.; Chardon, F.; Gaufichon, L.; Suzuki, A. Nitrogen uptake, assimilation and remobilization in plants: Challenges for sustainable and productive agriculture. Ann. Bot. 2010, 105, 1141–1157. [Google Scholar] [CrossRef]
- Yao, Q.; Yang, K.; Pan, G.; Rong, T. The effects of low phosphorus stress on morphological and physiological characteristics of maize (Zea mays L.) landraces. Agric. Sci. China 2007, 6, 559–566. [Google Scholar] [CrossRef]
- Rodriguez, D.; Andrade, F.; Goudriaan, J. Does assimilate supply limit leaf expansion in wheat grown in the field under low phosphorus availability? Field Crops Res. 2000, 67, 227–238. [Google Scholar] [CrossRef]
- Høgh-Jensen, H.; Schjoerring, J.K.; Soussana, J.F. The influence of phosphorus deficiency on growth and nitrogen fixation of white clover plants. Ann. Bot. 2002, 90, 745–753. [Google Scholar] [CrossRef]
- Chun, S.C.; Paramasivan, M.; Chandrasekaran, M. Proline accumulation influenced by osmotic stress in arbuscular mycorrhizal symbiotic plants. Front. Microbiol. 2019, 9, 2525. [Google Scholar] [CrossRef]
- Liang, X.; Zhang, L.; Natarajan, S.K.; Becker, D.F. Proline mechanisms of stress survival. Antioxid. Redox Signal. 2013, 19, 998–1011. [Google Scholar] [CrossRef]
- Siddique, A.; Kandpal, G.; Kumar, P. Proline accumulation and its defensive role under diverse stress condition in plants: An overview. J. Pure Appl. Microbiol. 2018, 12, 1655–1659. [Google Scholar] [CrossRef]
- Plenet, D.; Etchebest, S.; Mollier, A.; Pellerin, S. Growth analysis of maize field crops under phosphorus deficiency. Plant Soil 2000, 223, 117–130. [Google Scholar] [CrossRef]
- Assuero, S.G.; Mollier, A.; Pellerin, S. The decrease in growth of phosphorus-deficient maize leaves is related to a lower cell production. Plant Cell Environ. 2004, 27, 887–895. [Google Scholar] [CrossRef]
- Thornburg, T.E.; Liu, J.; Li, Q.; Xue, H.; Wang, G.; Li, L.; Julia Elise, F.; Kyele, E.D.; Wanying, L.; Baohong, Z.; et al. Potassium deficiency significantly affected plant growth and development as well as microRNA- mediated mechanism in wheat (Triticum aestivum L.). Front. Plant Sci. 2020, 11, 1219. [Google Scholar] [CrossRef]
- Amanullah; Iqbal, A.; Irfanullah; Hidayat, Z. Potassium management for improving growth and grain yield of maize (Zea mays L.) under moisture stress condition. Sci. Rep. 2016, 6, 34627. [Google Scholar] [CrossRef] [PubMed]
- Bukhsh, M.; Ahmad, R.; Ishaque, M.; Malik, A.U. Response of maize hybrids to varying potassium application in Pakistan. Pak. J. Agri. Sci. 2009, 46, 179–184. [Google Scholar]
- Jan, M.F.; Khan, A.A.; Liaqat, W.; Ahmad, H.; Ahmadzai, M.D.; Rehan, W. Response of phenology, growth and productivity of maize hybrids to integrated potassium management. Pak. J. Agric. Res. 2018, 31, 306–312. [Google Scholar] [CrossRef]
- Ul-Allah, S.; Ijaz, M.; Nawaz, A.; Sattar, A.; Sher, A.; Naeem, M.; Shahzad, U.; Farooq, U.; Nawaz, F.; Mahmood, K. Potassium application improves grain yield and alleviates drought susceptibility in diverse maize hybrids. Plants 2020, 9, 75. [Google Scholar] [CrossRef]
- Sustr, M.; Soukup, A.; Tylova, E. Potassium in root growth and development. Plants 2019, 8, 435. [Google Scholar] [CrossRef]
- Hodge, A.; Berta, G.; Doussan, C.; Merchan, F.; Crespi, M. Plant root growth, architecture and function. Plant Soil 2009, 321, 153–187. [Google Scholar] [CrossRef]
- Ryan, P.R.; Delhaize, E.; Watt, M.; Richardson, A.E. Plant roots: Understanding structure and function in an ocean of complexity. Ann. Bot. 2016, 118, 555–559. [Google Scholar] [CrossRef]
- Neumann, G.; Martinoia, E. Cluster roots—An underground adaptation for survival in extreme environments. Trends Plant Sci. 2002, 7, 162–167. [Google Scholar] [CrossRef]
- Mpanga, I.K.; Ludewig, U.; Dapaah, H.K.; Neumann, G. Acquisition of rock phosphate by combined application of ammonium fertilizers and Bacillus amyloliquefaciens FZB42 in maize as affected by soil pH. J. Appl. Microbiol. 2020, 129, 947–957. [Google Scholar] [CrossRef] [PubMed]
- Mpanga, I.K.; Nkebiwe, P.M.; Kuhlmann, M.; Cozzolino, V.; Piccolo, A.; Geistlinger, J.; Berger, N.; Ludewig, U.; Neumann, G. The form of N supply determines plant growth promotion by P-solubilizing microorganisms in maize. Microorganisms 2019, 7, 38. [Google Scholar] [CrossRef] [PubMed]
- Mpanga, I.K. Fertilization Strategies to Improve the Plant Growth-Promoting Potential of Microbial Bio-Effectors. Ph.D. Dissertation, University of Hohenheim, Stuttgart, Germany, 2020. [Google Scholar]
- Mpanga, I.K.; Dapaah, H.K.; Geistlinger, J.; Ludewig, U.; Neumann, G. Soil type-dependent interactions of P-solubilizing microorganisms with organic and inorganic fertilizers mediate plant growth promotion in tomato. Agronomy 2018, 8, 213. [Google Scholar] [CrossRef]
- Mpanga, I.K.; Gomez-Genao, N.; Moradtalab, N.; Wanke, D.; Chrobaczek, V.; Ahmed, A.; Windisch, S.; Geistlinger, J.; Hafiz, B.F.; Walker, F.; et al. The role of N form supply for PGPM-host plant interactions in maize. J. Plant Nut. Soil Sci. 2019, 182, 908–920. [Google Scholar] [CrossRef]
- Nkebiwe, P.M.; Weinmann, M.; Bar-Tal, A.; Müller, T. Fertilizer placement to improve crop nutrient acquisition and yield: A review and meta-analysis. Field Crops Res. 2016, 196, 389–401. [Google Scholar] [CrossRef]
- Amegbor, I.K.; Abe, A.; Adjebeng-Danquah, J.; Adu, G.B. Genetic analysis and yield assessment of maize hybrids under low and optimal nitrogen environments. Heliyon 2022, 8, e09052. [Google Scholar] [CrossRef]
- Das, B.; Atlin, G.N.; Olsen, M.; Burgueño, J.; Tarekegne, A.; Babu, R.; Ndou, E.N.; Mashingaidze, K.; Moremoholo, L.; Ligeyo, D.; et al. Identification of donors for low-nitrogen stress with maize lethal necrosis (MLN) tolerance for maize breeding in Sub-Saharan Africa. Euphytica 2019, 215, 80. [Google Scholar] [CrossRef]
- Sunday, I.; Babatunde, A.; Stephen, A.; Charity, A.; Kayode, O. Gene action in low nitrogen tolerance and implication on maize grain yield and associated traits of some tropical maize populations. Open Agric. 2020, 5, 801–805. [Google Scholar] [CrossRef]
- Mastrodomenico, A.T.; Bohn, M.O.; Lipka, A.E.; Below, F.E. Genomic selection using maize ex-plant variety protection germplasm for the prediction of nitrogen-use traits. Crop Sci. 2019, 59, 212–220. [Google Scholar] [CrossRef]
- Liu, Z.; Liu, X.; Craft, E.J.; Yuan, L.; Cheng, L.; Mi, G.; Chen, F. Physiological and genetic analysis for maize root characters and yield in response to low phosphorus stress. Breed. Sci. 2018, 68, 268–277. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Yuan, Y.; Liao, Z.; Jiang, Y.; Wang, Q.; Zhang, L.; Gao, S.; Wu, F.; Li, M.; Xie, W.; et al. Genome-wide association study of 13 traits in maize seedlings under low phosphorus stress. Plant Genome 2019, 12, 190039. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.; Zhang, H.; Sun, J.; Guo, Z.; Zou, C.; Li, W.X.; Xie, C.; Huang, C.; Xu, R.; Liao, H.; et al. Genome-wide association study dissects yield components associated with low-phosphorus stress tolerance in maize. Theor. Appl. Genet. 2018, 131, 1699–1714. [Google Scholar] [CrossRef] [PubMed]
- Minjian, C.; Haiqiu, Y.; Hongkui, Y.; Chunji, J. Difference in tolerance to potassium deficiency between two maize inbred lines. Plant Prod. Sci. 2007, 10, 42–46. [Google Scholar] [CrossRef]
- Zhao, X.H.; Yu, H.Q.; Wen, J.; Wang, X.G.; Du, Q.; Wang, J.; Wang, Q. Response of root morphology, physiology and endogenous hormones in maize (Zea mays L.) to potassium deficiency. J. Integr. Agric. 2016, 15, 785–794. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, N.; Liu, S.; Dong, A.; Zenda, T.; Liu, X.; Li, J.; Duan, H. Comparative proteomic analysis of two contrasting maize hybrids’ responses to low nitrogen stress at the twelve leaf stage and function verification of ZmTGA gene. Genes 2022, 13, 670. [Google Scholar] [CrossRef]
- Sun, P.; Mu, C.; Chen, Y.; Kong, X.; Xu, Y.; Zheng, H.; Zhang, H.; Wang, Q.; Xue, Y.; Li, Z.; et al. Comparative transcript profiling of maize inbreds in response to long-term phosphorus deficiency stress. Plant Physiol. Biochem. 2016, 109, 467–481. [Google Scholar] [CrossRef]
- Xiong, W.; Wang, Y.; Guo, Y.; Tang, W.; Zhao, Y.; Yang, G.; Pei, Y.; Chen, J.; Song, X.; Sun, J. Transcriptional and metabolic responses of maize shoots to long-term potassium deficiency. Front. Plant Sci. 2022, 13, 922581. [Google Scholar] [CrossRef]


{kind=link}
| Soil Nutrient | Traits | Heritability | Type of Inheritance | Population | Number of Environments | Method | No. of QTLs/SNPs | Authors |
|---|---|---|---|---|---|---|---|---|
| Nitrogen (N) | Grain yield, days to silking and anthesis, anthesis-silking interval, plant and ear height, stay green, and other agronomic traits | - | Additive, non-additive | 100 tropical hybrids | 2 years | Classical (GCA, SCA) | - | Amegbor et al. [48] |
| Grain yield, days to silking and anthesis, anthesis-silking interval, plant height, protein content | 0.12–0.92 | Polygenic a | 400 tropical elite lines | 10 locations | Per se; testcross analyses | - | Das et al. [49] | |
| Grain yield, days to silking and anthesis, anthesis-silking interval, plant and ear height, and other agronomic traits | - | Additive and non-additive | 45 tropical lines | 6 environments | Classical (GCA, SCA) | - | Sunday et al. [50] | |
| N use efficiency traits (grain yield, flowering time, plant height, etc.) | 0.33–0.95 | Polygenic a | 411 testcrosses (tropical) | 13 sites | Genomics (GWAS, GP) | 45 SNPs | Ertiro et al. [15] | |
| 12 N-use traits (e.g., yield, harvest index, N use efficiency, etc.) | 0.11–0.77 | Additive, non-additive | 89 ex-plant variety protection germplasm & two public lines (B73 and Mo17) | 11 environments (location-year) | Classical (GCA, SCA) & genomics (GP) | - | Mastrodomenico et al. [51] | |
| Phosphorus (P) | Root system architecture-related traits (e.g., root number and length, root biomass, etc.); PUE-related traits (e.g., grain yield, total P uptake, etc.) | - | Additive, non-additive | 6 inbred lines and their 15 F1 progenies | 1 year | Classical (GCA, SCA) | - | Liu et al. [52] |
| 13 traits at seedling stage (e.g., total root length, total root surface area, total root volume, total number of root tips, root forks, root diameter, longest root length, root dry weight, shoot dry weight, etc.) | 0.59–0.90 | Polygenic a | 356 inbred lines (non-stiff stalk, stiff stalk, tropical/sub-tropical) | 2 years | Genomics (GWAS) | 20–580, depending on trait | Wang et al. [53] | |
| P-use efficiency traits (yield per plant, shoot dry weight per plant, P content and concentrations in seeds and shoot, P utilization efficiency, etc.) | 0.47–0.77 | Polygenic a | 359 lines (non-stiff stalk, stiff stalk, tropical/subtropical) | 1 year | Genomics (GWAS, GP) | 49 (in low-P) | Li et al. [14] | |
| Five biomass- and development-related traits; six yield-related traits, | 0.33–0.87, depending on the trait | Polygenic a | 885 temperate, tropical/subtropical (475 for GWAS) | 2 years | Genomics (GWAS, GP) | 67 in 2014, 97 in 2015, 19 in combined years | Xu et al. [54] | |
| Potassium (K) | number of leaves, symptom of K deficiency, tap and lateral root length, K-content | - | - | 2 inbreds | 1 (hydroponic) | Classical (line evaluation) | - | Minjian et al. [55] |
| Root morphological traits e.g., root length, volume, surface area, diameter; Lipid peroxidation, Superoxide dismutase and endogenous hormones | - | - | 2 inbreds | 1 year | Classical (line evaluation) | - | Zhao et al. [56] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gaikpa, D.S.; Opata, J.; Mpanga, I.K. Towards Sustainable Maize Production: Understanding the Morpho-Physiological, Genetics, and Molecular Mechanisms for Tolerance to Low Soil Nitrogen, Phosphorus, and Potassium. Stresses 2022, 2, 395-404. https://doi.org/10.3390/stresses2040028
Gaikpa DS, Opata J, Mpanga IK. Towards Sustainable Maize Production: Understanding the Morpho-Physiological, Genetics, and Molecular Mechanisms for Tolerance to Low Soil Nitrogen, Phosphorus, and Potassium. Stresses. 2022; 2(4):395-404. https://doi.org/10.3390/stresses2040028
Chicago/Turabian StyleGaikpa, David Sewordor, John Opata, and Isaac Kwadwo Mpanga. 2022. "Towards Sustainable Maize Production: Understanding the Morpho-Physiological, Genetics, and Molecular Mechanisms for Tolerance to Low Soil Nitrogen, Phosphorus, and Potassium" Stresses 2, no. 4: 395-404. https://doi.org/10.3390/stresses2040028
APA StyleGaikpa, D. S., Opata, J., & Mpanga, I. K. (2022). Towards Sustainable Maize Production: Understanding the Morpho-Physiological, Genetics, and Molecular Mechanisms for Tolerance to Low Soil Nitrogen, Phosphorus, and Potassium. Stresses, 2(4), 395-404. https://doi.org/10.3390/stresses2040028