
Infective Larvae of Haemonchus contortus (Nematoda: Trichostrongylidae) Are Captured and Destroyed by Nematode-Trapping Fungi Dactylellina spp. (Fungi: Orbiliales)

 ,
,  , ,
, ,  , , and
, , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Allocation
2.2. Biological Material
2.2.1. Panagrellus Redivivus
2.2.2. Haemonchus Contortus
2.2.3. Obtaining Nematophagous Fungi Isolate
2.3. Fungal Morphometric Taxonomical Identification
2.4. Molecular Identification of Nematophagous Fungi
2.5. Phylogenetic Analysis
2.6. Assessment of Predatory Activity of Fungal Isolates
2.7. Nematophagous Fungi Liquid Cultures
2.8. Obtaining Liquid Culture Filtrates
2.9. Assessing In Vitro Nematocidal Activity of Fungal Liquid Culture Filtrates Against Haemonchus contortus Infective Larvae
2.10. Statistical Analysis
3. Results
3.1. Isolation and Morphological Identification of Fungi Isolates
3.1.1. Isolate D1
3.1.2. Isolate D2
3.2. Molecular Identification of Fungi Isolates
3.3. Predatory Activity of Isolated Fungi
3.4. Nematocidal Effect of Liquid Culture Filtrates of Fungal Isolates
4. Discussion
4.1. Morphological and Molecular Identification
4.2. Assessment of Predatory Activity of Fungal Isolates Against Haemonchus contortus Infective Larvae
4.3. Assessment of In Vitro Nematocidal Activity of Fungal Isolates Against Haemonchus contortus Infective Larvae
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Van der Hoogen, J.; Geisen, S.; Routh, D.; Ferris, H.; Traunspurger, W.; Wardle, D.; De Goede, R.G.M.; Adams, B.J.; Ahmad, W.; Andriuzzi, W.; et al. Soil nematode abundance and functional group composition at a global scale. Nature 2019, 572, 194–198. [Google Scholar] [CrossRef]
- Delgado-Baquerizo, M.; Eldridge, D.J.; Liu, Y.R.; Liu, Z.W.; Coleine, C.; Trivedi, P. Soil biodiversity and function under global change. PLoS Biol. 2025, 23, e3003093. [Google Scholar] [CrossRef]
- Lawal, I.; Fardami, A.Y.; Ahmad, F.I.I.; Yahaya, S.; Abubakar, A.S.; Sa’id, M.A.; Maiyadi, K.A. A review on nematophagus fungi: A potential nematicide for the biocontrol of nematodes. J. Environ. Bioremediat. Toxicol. 2022, 5, 26–31. [Google Scholar] [CrossRef]
- Nordbring-Hertz, B. Morphogenesis in the nematode-trapping fungus Arthrobotrys oligospora: An extensive plasticity of infection structures. Mycologist 2004, 18, 125–133. [Google Scholar] [CrossRef]
- Zhu, J.; Zhang, W.; Fan, Y.; Deng, W.; Zhang, L.; Wang, S.; Xiang, M. Septin AoCDC11 is involved in trap morphogenesis, conidiation, and vegetative growth in carnivorous Arthrobotrys oligospora. Fungal Genet. Biol. 2025, 177, 103971. [Google Scholar] [CrossRef]
- Hsueh, Y.P.; Gronquist, M.R.; Schwarz, E.M.; Nath, R.D.; Lee, C.H.; Gharib, S.; Sternberg, P.W. Nematophagous fungus Arthrobotrys oligospora mimics olfactory cues of sex and food to lure its nematode prey. Elife 2017, 6, e20023. [Google Scholar] [CrossRef]
- Rahman, M.U.; Chen, P.; Zhang, X.; Fan, B. Predacious strategies of nematophagous fungi as bio-control agents. Agronomy 2023, 13, 2685. [Google Scholar] [CrossRef]
- Zhang, J.; Wei, L.; Zhang, H.; Ma, X.; Sun, Y.; Li, R.; Meng, Q. Proteomic insights into nematode-trapping fungi Arthrobotrys oligospora after their response to chitin. J. Vet. Res. 2025, 69, 71. [Google Scholar] [CrossRef]
- De Hoog, G.S.; van Oorschot, C.A.N. Taxonomy of the Dactylaria complex VI. Key to the genera and check-list of epithets. Stud. Mycol. 1985, 26, 97–122. [Google Scholar]
- Kumar, D. Evaluation of nematophagous and biocontrol efficacy of Dactylellina phymatopaga against Meloidogyne graminicola on rice (Oryza sativa L.). Biol. Control 2024, 188, 105425. [Google Scholar] [CrossRef]
- Xue, Y.J.; Li, E.L.; Wang, A.H.; Cai, K.Z. Predatory activity and passage of six nematophagous fungi species in gastrointestinal tract of trichostrongylide-infected sheep. Biocontrol Sci. Technol. 2018, 28, 654–662. [Google Scholar] [CrossRef]
- Abosse, J.S.; Terefe, G.; Teshale, B.M. Comparative study on pathological changes in sheep and goats experimentally infected with Haemonchus contortus. Surg. Exp. Pathol. 2022, 5, 14. [Google Scholar] [CrossRef]
- Khalifa, H.O.; Shikoray, L.; Mohamed, M.Y.I.; Habib, I.; Matsumoto, T. Veterinary drug residues in the food chain as an emerging public health threat: Sources, analytical methods, health impacts, and preventive measures. Foods 2024, 13, 1629. [Google Scholar] [CrossRef]
- Vokřál, I.; Podlipná, R.; Matoušková, P.; Skálová, L. Anthelmintics in the environment: Their occurrence, fate, and toxicity to non-target organisms. Chemosphere 2023, 345, 140446. [Google Scholar] [CrossRef]
- De Lara, R.; Castro, T.; Castro, J.; Castro, G. Cultivo del nematodo Panagrellus redivivus (Goodey, 1945) en un medio de avena enriquecida con Spirulina sp. Rev. Biol. Mar. Oceanogr. 2007, 42, 29–36. [Google Scholar] [CrossRef]
- Olmedo-Juárez, A.; Delgado-Núñez, E.J.; Bahena-Vicencio, A.; Villa-Mancera, A.; Zamilpa, A.; González-Cortázar, M.; Mendoza de Gives, P. In vitro nematocidal properties from two extracts: Lippia graveolens leaves and Delonix regia flowers against eggs and infective larvae of Haemonchus contortus. J. Med. Food 2022, 25, 329–337. [Google Scholar] [CrossRef]
- Valcarcel-Sancho, F.V.; Vázquez, F.A.R.; García, Á.S.O.; Novillo, M.B.A.; Sopeña, L.M. Atlas de Parasitología Ovina; Servet: Fethiye/Muğla, Türkiye, 2009; 137p, ISBN 978-84-92569-05-2. [Google Scholar]
- Liébano-Hernández, E. Ecología de larvas de nematodos gastrointestinales de bovinos, ovinos y caprinos. Epidemiol. Enferm. Paras. Anim. Domést. 2011, 1, 267–268. [Google Scholar]
- Olmedo-Juárez, A.; Rojo-Rubio, R.; Zamilpa, A.; Mendoza de Gives, P.; Arece-García, J.; López-Arellano, M.E.; von Son-de Fernex, E. In vitro larvicidal effect of a hydroalcoholic extract from Acacia cochliacantha leaf against ruminant parasitic nematodes. Vet. Res. Commun. 2017, 41, 227–232. [Google Scholar] [CrossRef]
- ANP Texcal. Secretaría de Desarrollo Sustentable. Available online: https://sustentable.morelos.gob.mx/anp/el-texcal (accessed on 26 January 2017).
- Berhanu, M.; Gebeyaw, D.T.; Kefale, D.; Kang, Y. Overview of nematophagous fungi, isolation techniques, and their role in biological control of helminthic parasites: A literature review. Acta Entomol. Zool. 2024, 5, 133–143. [Google Scholar] [CrossRef]
- Bahena-Nuñez, D.S.; Ocampo-Gutiérrez, A.Y.; Mendoza-de Gives, P.; González-Cortázar, M.; Zamilpa, A.; Higuera-Piedrahita, R.I.; Hernández-Romano, J. Arthrobotrys oligospora (Fungi: Orbiliales) and its liquid culture filtrate myco-constituents kill Haemonchus contortus infective larvae (Nematoda: Trichostrongylidae). Biocontrol Sci. Technol. 2024, 34, 754–775. [Google Scholar] [CrossRef]
- Noman, E.; Al-Gheethi, A.A.; Rahman, N.K.; Talip, B.; Mohamed, R.; Kadir, O.A. Single spore isolation as a simple and efficient technique to obtain fungal pure culture. In IOP Conference Series: Earth and Environmental Science; IOP: Bristol, UK, 2018; Volume 140, p. 12055. [Google Scholar]
- Campbell, C.K.; Johnson, E.M. Identification of Pathogenic Fungi: Campbell/Identification of Pathogenic Fungi; Warnock, D.W., Ed.; Wiley-Blackwell: Chichester, UK, 2013; ISBN 9781444330700. [Google Scholar]
- Drechsler, C. Three fungi destructive to free-living terricolous nematodes. J. Wash. Acad. Sci. 1940, 30, 240–254. [Google Scholar]
- Yu, Z.; Mo, M.; Zhang, Y.; Zhang, K.Q. Taxonomy of nematode-trapping fungi from Orbiliaceae, Ascomycota. In Nematode-Trapping Fungi; Zhang, K.-Q., Hyde, K.D., Eds.; Springer Science & Business: Berlin/Heidelberg, Germany, 2014; Volume 23, pp. 41–210. [Google Scholar]
- White, T.J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Academic Press: San Diego, CA, USA, 1990; pp. 315–322. [Google Scholar]
- Stielow, J.; Lévesque, C.; Seifert, K.; Meyer, W.; Irinyi, L.; Smits, D.; Renfurm, R.; Verkley, G.; Groenewald, M.; Chaduli, D.; et al. One fungus, which genes? Development and assessment of universal primers for potential secondary fungal DNA barcodes. Persoonia Mol. Phylogeny Evol. Fungi 2015, 35, 242–263. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Anzúrez, G.; Olmedo-Juárez, A.; Von-Son de Fernex, E.; Alonso-Díaz, M.Á.; Delgado-Núñez, E.J.; López-Arellano, M.E.; Mendoza-de Gives, P. Arthrobotrys musiformis (Orbiliales) kills Haemonchus contortus infective larvae (Trichostronylidae) through its predatory activity and its fungal culture filtrates. Pathogens 2022, 11, 1068. [Google Scholar] [CrossRef]
- Ocampo-Gutiérrez, A.Y.; Hernández-Velázquez, V.M.; Aguilar-Marcelino, L.; Cardoso-Taketa, A.; Zamilpa, A.; López-Arellano, M.E.; Mendoza-de Gives, P. Morphological and molecular characterization, predatory behavior and effect of organic extracts of four nematophagous fungi from Mexico. Fungal Ecol. 2021, 49, 101004. [Google Scholar] [CrossRef]
- Jaramillo-Tlalapango, J.; Gives, P.M.-D.; Isabel-Higuera-Piedrahita, R.; Ocampo-Gutiérrez, A.Y.; Eugenia-López-Arellano, M.; Pérez-Anzúrez, G.; Olmedo-Juárez, A.; Hernández-Romano, J.; Maza-Lopez, J.; Delgado-Núñez, E.J.; et al. Study of a Mexican isolate of Arthrobotrys musiformis (Orbiliales): Predatory behavior and nematocidal activity of liquid culture filtrates against Haemonchus contortus (Trichostrongylidae), protein profile and myco-constituent groups. Fungal Biol. 2023, 127, 1345–1361. [Google Scholar] [CrossRef]
- Campos, A.K.; Valadão, M.C.; Carvalho, L.M.; Araújo, J.V.d.; Guimarães, M.P. In vitro nematophagous activity of predatory fungi on infective larvae of Strongyloides papillosus. Acta Vet. Bras. 2017, 11, 213–218. [Google Scholar] [CrossRef]
- Wairimu, W.J.; Kimenjul, J.W.; Muiru, W.M.; Wachira, P.M. Interactions between soil additives and a variety of naturally occurring nematode-demolishing fungi in banana fields of Meru and Embu Counties, Kenya. Cell Biol. Dev. 2022, 6. [Google Scholar] [CrossRef]
- Wen, X.F.; Shi, T.T.; Zhang, Y.Q.; Wang, S.H.; Xiang, C.M.; Zhao, P.J. DHXT1, a Virulence Factor of Dactylellina haptotyla, Regulates Pathogenicity by Participating in Trap Formation and Metabolite Synthesis. Int. J. Mol. Sci. 2024, 25, 7384. [Google Scholar] [CrossRef]
- Pérez-Anzúrez, G.; Mendoza-de Gives, P.; Alonso-Díaz, M.Á.; von Son-de Fernex, E.; Paz-Silva, A.; López-Arellano, M.E.; Olmedo-Juárez, A. Lecanicillium psalliotae (Hypocreales: Cordycipitaceae) Exerts Ovicidal and Larvicidal Effects against the Sheep Blood-Feeding Nematode Haemonchus contortus through Its Liquid Culture Filtrates. Pathogens 2024, 13, 588. [Google Scholar] [CrossRef]
- Sharma, A.; Sharma, S.; Mittal, A.; Naik, S.N. Evidence for the involvement of nematocidal toxins of Purpureocillium lilacinum 6029 cultured on Karanja deoiled cake liquid medium. World J. Microbiol. Biotechnol. 2016, 32, 82. [Google Scholar] [CrossRef]
- Kuo, T.H.; Yang, C.T.; Chang, H.Y.; Hsueh, Y.P.; Hsu, C.C. Nematode-trapping fungi produce diverse metabolites during predator–prey interaction. Metabolites 2020, 10, 117. [Google Scholar] [CrossRef] [PubMed]
- Mendoza-de Gives, P.; Braga, F.R.; Araújo, J.V.d. Nematophagous fungi, an extraordinary tool for controlling ruminant parasitic nematodes and other biotechnological applications. Biocontrol Sci. Technol. 2022, 32, 777–793. [Google Scholar] [CrossRef]
- Degenkolb, T.; Vilcinskas, A. Metabolites from nematophagous fungi and nematicidal natural products from fungi as an alternative for biological control. Part I: Metabolites from nematophagous ascomycetes. Appl. Microbiol. Biotechnol. 2016, 100, 3799–3812. [Google Scholar] [CrossRef] [PubMed]
- Wan, J.; Dai, Z.; Zhang, K.; Li, G.; Zhao, P. Pathogenicity and metabolites of endoparasitic nematophagous fungus Drechmeria coniospora YMF1.01759 against nematodes. Microorganisms 2021, 9, 1735. [Google Scholar] [CrossRef]
- Dai, Z.; Gan, Y.; Zhao, P.; Li, G. Secondary metabolites from the endoparasitic nematophagous fungus Harposporium anguillulae YMF 1.01751. Microorganisms 2022, 10, 1553. [Google Scholar] [CrossRef]
- Lei, H.-M.; Wang, J.-T.; Hu, Q.-Y.; Li, C.-Q.; Mo, M.-H.; Zhang, K.-Q.; Li, G.-H.; Zhao, P.-J. 2-Furoic acid associated with the infection of nematodes by Dactylellina haptotyla and its biocontrol potential on plant root-knot nematodes. Microbiol. Spectr. 2023, 11, e01896-23. [Google Scholar] [CrossRef]
- Rodrigues, J.A.; Alvares, F.B.V.; Silva, J.T.; Ferreira, L.C.; da Costa, P.W.L.; Sarmento, W.F.; Feitosa, T.F.; de Araújo, J.V.; Braga, F.R.; Vilela, V.L.R. Predatory effects of the fungus Arthrobotrys cladodes on sheep gastrointestinal nematodes. Biocontrol Sci. Technol. 2020, 30, 830–839. [Google Scholar] [CrossRef]
- Mendoza-de Gives, P. Soil-Borne nematodes: Impact in agriculture and livestock and sustainable strategies of prevention and control with special reference to the use of nematode natural enemies. Pathogens 2022, 11, 640. [Google Scholar] [CrossRef]
- Nolinda, N.; Ikusika, O.O.; Akinmoladun, O.F.; Mpendulo, C.T. Impact of nematode infestation in livestock production and the role of natural feed additives–A review. Open Agric. 2024, 9, 20220234. [Google Scholar] [CrossRef]
- EFSA Panel on Additives and Products or Substances used in Animal Feed (FEEDAP); Bampidis, V.; Azimonti, G.; Bastos, M.d.L.; Christensen, H.; Durjava, M.; Dusemund, B.; Kouba, M.; López-Alonso, M.; Puente, S.L.; et al. Safety of a feed additive consisting of Duddingtonia flagrans NCIMB 30336 (BioWorma®) for all grazing animals (International Animal Health Products Pty Ltd.). EFSA J. 2023, 21, e8465. [Google Scholar]
- Ma, Y.; Jiang, L.; Fan, Z.; Hao, L.; Li, Z.; Zhang, Y.; Li, Q.; Wang, R.; Luo, H. Nematode controlling effects and safety tests of Duddingtonia flagrans biological preparation in sheep. Sci. Rep. 2025, 15, 1843. [Google Scholar] [CrossRef] [PubMed]
- Chavarría-Joya, L.; Alonso-Díaz, M.Á.; Olmedo-Juárez, A.; von Son-de Fernex, E.; Mendoza-de-Gives, P. Assessing the individual and combined use of Caesalpinia coriaria (Plantae: Fabaceae) and Duddingtonia flagrans (Fungi: Orbiliaceae) as sustainable alternatives of control of sheep parasitic nematodes. Biocontrol Sci. Technol. 2022, 32, 1260–1274. [Google Scholar] [CrossRef]
- Liu, H.; Zhang, Y.; Li, J.; Liu, F.; Ye, L.; Liu, X.; Hu, M. Identification and validation of protective glycoproteins in Haemonchus contortus H11. Front. Immunol. 2025, 16, 1521022. [Google Scholar] [CrossRef] [PubMed]
- Forbes, A.; Ellis, K. Parasite control in regenerative livestock farming. Livestock 2023, 28, 112–120. [Google Scholar] [CrossRef]
- Siddique, A.; Whitley, N.; Samples, O.M.; Burke, J.M.; Terrill, T.H.; Miller, J.E.; Connell, T. Exploring the Role of Duddingtonia flagrans in Integrated Parasite Management: A Systematic Review of Current Research and Applications. preprints 2025, 2025021078. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Measurement | Mean (μm) | Range (μm) |
|---|---|---|
| Conidia length | 39.42 | 31.17–48.84 |
| Conidia width | 2.74 | 2.09–3.21 |
| Distal cell length | 4.4 | 4.01–5.07 |
| Distal cell width | 3.31 | 2.5–3.98 |
| Conidiophore length | 30.33 | 15.43–66.2 |
| Conidiophore width | 2.83 | 2.11–3.85 |
| Chlamydospores | Concatenated | |
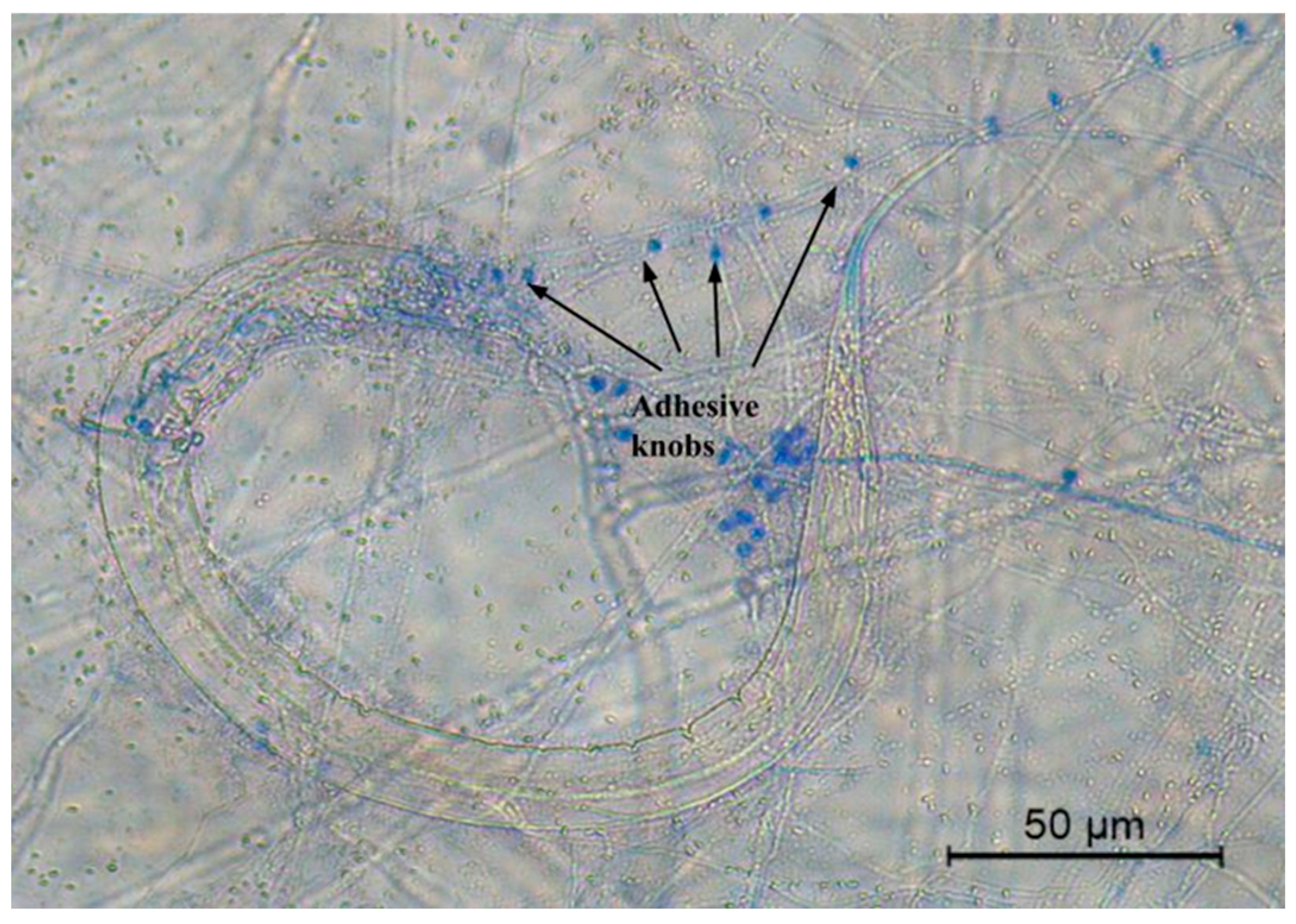
| Type of trapping devices | Adhesive knobs and sessile conidia | |
| Isolate | % Coverage | % Identity | NCBI Access Code |
|---|---|---|---|
| Dactyllelina haptospora | 100% | 93.32% | DQ999820.1 |
| D. haptospora | 98% | 93.43% | U51946.1 |
| D. thibetensis | 98% | 89.41% | DQ999833.1 |
| Dactylellina sp. | 98% | 89.62% | JX043193.1 |
| Dactylellina sp. | 99% | 89.01% | ON408470.1 |
| Isolate | % Coverage | % Identity | NCBI Access Code |
|---|---|---|---|
| Dactyllelina phymatopaga | 99% | 97.52% | KT215203.1 |
| D. phymatopaga | 98% | 96.22% | U51970.1 |
| D. phymatopaga | 97% | 95.48% | OR859126.1 |
| D. parvicollis | 89% | 95.11% | OQ398138.1 |
| D. parvicollis | 92% | 93.40% | AY804215.1 |
| Isolate | Mean of Recovered Larvae ± SE (Control Group) | Mean of Recovered Larvae ± SE (Larvae/Fungi Interaction Group) | Larval Reduction (%) |
|---|---|---|---|
| Dactyllelina haptospora | 265.3 ± 56.3 | 14 ± 10.5 | 94.72 |
| Dactylellina phymatopaga | 141.4 ± 20.7 | 44 ± 17.2 | 68.88 |
| Liquid Culture Filtrate Concentration (mg/mL) | Isolate | Dead/Total Larvae | Larval Mortality (%) (Mean ± SE) |
|---|---|---|---|
| 0 | D. haptospora | 2/110 | 1.25 ± 0.40 a |
| D. phymatopaga | 2/110 | 1.25 ± 0.40 a | |
| Control (Without fungi) | 2/110 | 1.25 ± 0.43 a | |
| 25 | D. haptospora | 4/110 | 3.51 ± 0.61 a |
| D. phymatopaga | 3/110 | 2.94 ± 0.45 a | |
| Control (Without fungi) | 2/110 | 2.20 ± 0.45 a | |
| 50 | D. haptospora | 10/110 | 8.89 ± 1.12 a |
| D. phymatopaga | 8/110 | 7.55 ± 1.36 a | |
| Control (Without fungi) | 2/110 | 3.56 ± 0.58 a | |
| 100 | D. haptospora | 31/110 | 27.83 ± 2.93 b |
| D. phymatopaga | 37/110 | 33.9 ± 2.54 b | |
| Control (Without fungi) | 2/110 | 5.54 ± 1.24 a |
| Genus/Species | Target Nematode | Conditions | Efficacy | Country | Author |
|---|---|---|---|---|---|
| D. leptospora | Various species of gastrointestinal parasitic nematodes (L3) | In vitro | 75.72% | China | [11] |
| D. phymatopaga | Unspecified genus/species | Banana | 63% | Kenya | [33] |
| D. phymatopaga | Meloidogyne graminicola | Rice | 98.8% | India | [10] |
| D. haptotyla YMF1.03409 (A WT mutant strain) | Caenorhabditis elegans | In vitro | 82.35% (after 24 h), 99.4% (After 48 h) | China | [34] |
| D. haptospora D. phymatophaga | Haemonchus contortus | In vitro | 94.79% 68.88% | Mexico | Present study |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Balanzar-Aguilera, M.S.; Gutiérrez-Medina, E.; Pérez-Anzúrez, G.; Delgado-Núñez, E.J.; López-Arellano, M.E.; Ocampo-Gutiérrez, A.Y.; Mendoza-de Gives, P. Infective Larvae of Haemonchus contortus (Nematoda: Trichostrongylidae) Are Captured and Destroyed by Nematode-Trapping Fungi Dactylellina spp. (Fungi: Orbiliales). Parasitologia 2025, 5, 26. https://doi.org/10.3390/parasitologia5020026
Balanzar-Aguilera MS, Gutiérrez-Medina E, Pérez-Anzúrez G, Delgado-Núñez EJ, López-Arellano ME, Ocampo-Gutiérrez AY, Mendoza-de Gives P. Infective Larvae of Haemonchus contortus (Nematoda: Trichostrongylidae) Are Captured and Destroyed by Nematode-Trapping Fungi Dactylellina spp. (Fungi: Orbiliales). Parasitologia. 2025; 5(2):26. https://doi.org/10.3390/parasitologia5020026
Chicago/Turabian StyleBalanzar-Aguilera, Manuel Salvador, Enrique Gutiérrez-Medina, Gustavo Pérez-Anzúrez, Edgar Jesús Delgado-Núñez, María Eugenia López-Arellano, Ana Yuridia Ocampo-Gutiérrez, and Pedro Mendoza-de Gives. 2025. "Infective Larvae of Haemonchus contortus (Nematoda: Trichostrongylidae) Are Captured and Destroyed by Nematode-Trapping Fungi Dactylellina spp. (Fungi: Orbiliales)" Parasitologia 5, no. 2: 26. https://doi.org/10.3390/parasitologia5020026
APA StyleBalanzar-Aguilera, M. S., Gutiérrez-Medina, E., Pérez-Anzúrez, G., Delgado-Núñez, E. J., López-Arellano, M. E., Ocampo-Gutiérrez, A. Y., & Mendoza-de Gives, P. (2025). Infective Larvae of Haemonchus contortus (Nematoda: Trichostrongylidae) Are Captured and Destroyed by Nematode-Trapping Fungi Dactylellina spp. (Fungi: Orbiliales). Parasitologia, 5(2), 26. https://doi.org/10.3390/parasitologia5020026