
Surveillance of Host-Seeking Ticks in the Flint Hills Region (USA) and Associations with Environmental Determinants

 , ,
, , 
Abstract
:1. Introduction
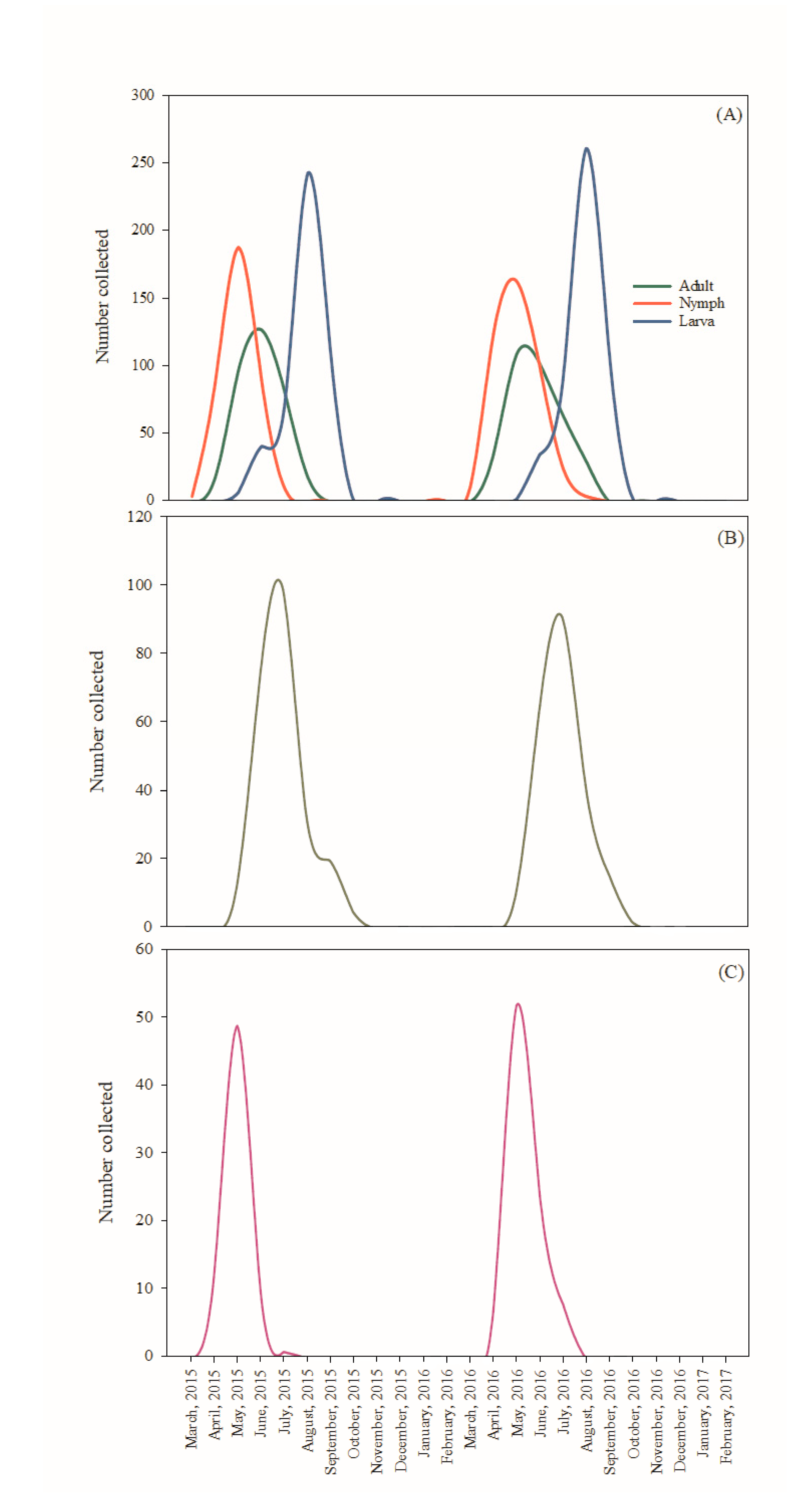
2. Results
3. Discussion
4. Materials and Methods
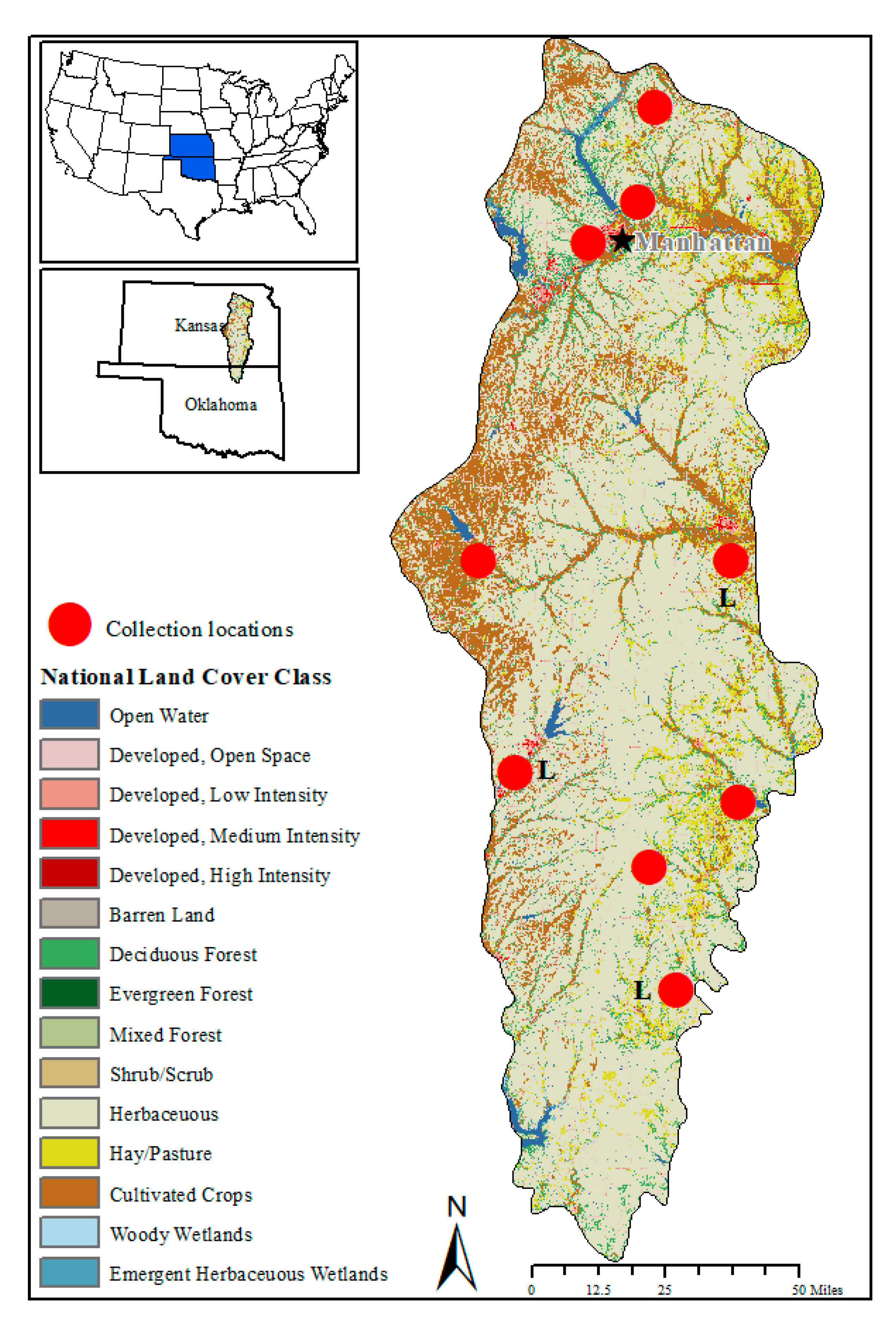
4.1. Study Area
4.2. Tick Survey and Classification
4.3. Sample Handling and DNA Extraction
4.3.1. Positive Control Construction
4.3.2. Real-Time PCR
4.4. Environmental Data
4.5. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Goodger, W.J.; Carpenter, T.; Riemann, H. Estimation of economic loss associated with anaplasmosis in California beef cattle. J. Am. Vet. Med. Assoc. 1979, 174, 1333–1336. [Google Scholar]
- Alderink, F.J.; Dietrich, R.A. Economic and Epidemiological Implications of Anaplasmosis in Texas Beef Cattle Herds. Available online: https://core.ac.uk/download/pdf/18488711.pdf (accessed on 6 August 2021).
- Okafor, C.C.; Collins, S.L.; Daniel, J.A.; Harvey, B.; Coetzee, J.; Whitlock, B. Factors associated with seroprevalence of bovine anaplasmosis in Texas. Vet. Parasitol. Reg. Stud. Rep. 2018, 14, 32–40. [Google Scholar] [CrossRef] [PubMed]
- Raghavan, R.K.; Harrington, J.; Anderson, G.A.; Hutchinson, J.S.; Debey, B.M. Environmental, climatic, and residential neighborhood determinants of feline tularemia. Vector Borne Zoonotic Dis. 2013, 13, 449–456. [Google Scholar] [CrossRef] [Green Version]
- Raghavan, R.K.; Neises, D.; Goodin, U.G.; Andresen, D.A.; Ganta, R.R. Bayesian spatio-temporal analysis and geospatial risk factors of human monocytic ehrlichiosis. PLoS ONE 2014, 9, e100850. [Google Scholar] [CrossRef] [Green Version]
- Raghavan, R.K.; Almes, K.; Goodin, D.G.; Harrington, J.A., Jr.; Stackhouse, P.W., Jr. Spatially heterogeneous land cover/land use and climatic risk factors of tick-borne feline cytauxzoonosis. Vector Borne Zoonotic Dis. 2014, 14, 486–495. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raghavan, R.K.; Goodin, D.G.; Neises, D.; Anderson, G.A.; Ganta, R.R. Hierarchical Bayesian spatio–temporal analysis of climatic and socio–economic determinants of Rocky Mountain spotted fever. PLoS ONE 2016, 11, e0150180. [Google Scholar] [CrossRef] [Green Version]
- Hanzlicek, G.A.; Raghavan, R.K.; Ganta, R.R.; Anderson, G.A. Bayesian space-time patterns and climatic determinants of bovine anaplasmosis. PLoS ONE 2016, 11, e0151924. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Savage, H.M.; Godsey, M.S., Jr.; Tatman, J.; Burkhalter, K.L.; Hamm, A.; Panella, N.A.; Ghosh, A.; Raghavan, R.K. Surveillance for Heartland and Bourbon viruses in eastern Kansas, June 2016. J. Med. Entomol. 2018, 55, 1613–1616. [Google Scholar] [CrossRef] [Green Version]
- Savage, H.M.; Godsey, M.S., Jr.; Panella, N.A.; Burkhalter, K.L.; Manford, J.; Trevino-Garrison, I.C.; Straily, A.; Wilson, S.; Bowen, J.; Raghavan, R.K. Surveillance for tick-borne viruses near the location of a fatal human case of Bourbon virus (Family Orthomyxoviridae: Genus Thogotovirus) in eastern Kansas, 2015. J. Med. Entomol. 2018, 55, 701–705. [Google Scholar] [CrossRef]
- Raghavan, R.K.; Goodin, D.G.; Dryden, M.W.; Hroobi, A.; Gordon, D.M.; Cheng, C.; Nair, A.D.; Jakkula, L.U.; Hanzlicek, G.A.; Anderson, G.A.; et al. Heterogeneous associations of ecological attributes with tick-borne Rickettsial pathogens in a periurban landscape. Vector Borne Zoonotic Dis. 2016, 16, 569–576. [Google Scholar] [CrossRef] [Green Version]
- Halos, L.; Bord, S.; Cotté, V.; Gasqui, P.; Abrial, D.; Barnouin, J.; Boulouis, H.-J.; Vayssier-Taussat, M.; Vourc, G. Ecological factors characterizing the prevalence of bacterial tick-borne pathogens in Ixodes ricinus ticks in pastures and woodlands. Appl. Environ. Microbiol. 2010, 76, 4413–4420. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, T.L.; Boegler, K.A.; Clark, R.J.; Delorey, M.J.; Bjork, J.K.H.; Dorr, F.M.; Schiffman, E.K.; Neitzel, D.F.; Monaghan, A.J.; Eisen, R.J. An acarological risk model predicting the density and distribution of host-seeking Ixodes scapularis nymphs in Minnesota. Am. J. Trop. Med. Hyg. 2018, 98, 1671–1682. [Google Scholar] [CrossRef] [PubMed]
- Polito, V.J.; Baum, K.; Payton, M.E.; Little, S.E.; Fuhlendorf, S.D.; Reichard, M.V. Tick abundance and levels of infestation on cattle in response to patch burning. Rangel. Ecol. Manag. 2013, 66, 545–552. [Google Scholar] [CrossRef]
- Gleim, E.R.; Zemtsova, G.E.; Berghaus, R.; Levin, M.L.; Conner, M.; Yabsley, M.J. Frequent prescribed fires can reduce risk of tick-borne diseases. Sci. Rep. 2019, 9, 1–10. [Google Scholar] [CrossRef]
- Barnard, D.R. Aspects of the bovine host-lone star tick interaction process in forage areas. In Morphology, Physiology and Behavioral Ecology of Ticks; Sauer, J.R., Hair, J.A., Eds.; Horwood: Chichester, UK, 1986; pp. 428–444. [Google Scholar]
- Hair, J.A.; Bowman, J. Behavioral ecology of Amblyomma americanum (L.). In Morphology, Physiology and Behavioral Ecology of Ticks; Sauer, J.R., Hair, J.A., Eds.; Horwood: Chichester, UK, 1986; pp. 406–427. [Google Scholar]
- White, S.C.; Mock, D.E. Ixodes scapularis (Acari: Ixodidae) in Kansas: A new distribution record. J. Med. Èntomol. 1991, 28, 872–873. [Google Scholar] [CrossRef] [PubMed]
- Kollars, T.M.; Oliver, J.H.; Kollars, P.G.; Durden, L.A. Seasonal activity and host associations of Ixodes scapularis (Acari: Ixodidae) in southeastern Missouri. J. Med. Èntomol. 1999, 36, 720–726. [Google Scholar] [CrossRef] [PubMed]
- Hroobi, A.; Boorgula, G.D.; Gordon, D.; Bai, J.; Goodin, D.; Anderson, G.; Wilson, S.; Staggs, A.; Raghavan, R.K. Diversity and seasonality of host-seeking ticks in a periurban environment in the central Midwest (USA). PLoS ONE 2021, 16, e0250272. [Google Scholar] [CrossRef] [PubMed]
- Remesar, S.; Fernández, P.D.; Venzal, J.M.; Pérez-Creo, A.; Prieto, A.; Estrada-Peña, A.; López, C.M.; Panadero, R.; Fernández, G.; Díez-Baños, P.; et al. Tick species diversity and population dynamics of Ixodes ricinus in Galicia (north-western Spain). Ticks Tick Borne Dis. 2018, 10, 132–137. [Google Scholar] [CrossRef]
- Schulze, T.L.; Jordan, R.A.; Hung, R.W. Effects of selected meteorological factors on diurnal questing of Ixodes scapularis and Amblyomma americanum (Acari: Ixodidae). J. Med. Èntomol. 2001, 38, 318–324. [Google Scholar] [CrossRef] [PubMed]
- Harlan, H.J.; Foster, W.A. Micrometeorologic factors affecting field host-seeking activity of adult Dermacentor variabilis (Acari: Ixodidae). J. Med. Èntomol. 1990, 27, 471–479. [Google Scholar] [CrossRef]
- Sonenshine, D.E.; Haines, G. A convenient method for controlling populations of the american dog tick, Dermacentor variabilis (Acari: Ixodidae) in the natural environment. J. Med. Èntomol. 1985, 22, 577–583. [Google Scholar] [CrossRef] [PubMed]
- Haile, D.G.; Mount, G.A. Computer simulation of population dynamics of the lone star tick, Amblyomma americanum (Acari: Ixodidae). J. Med. Èntomol. 1987, 24, 356–369. [Google Scholar] [CrossRef]
- Yoder, J.A.; Rosendale, A.J.; Benoit, J.B. Short day-triggered quiescence promotes water conservation in the American dog tick, Dermacentor variabilis. J. Comp. Physiol. B 2015, 186, 287–296. [Google Scholar] [CrossRef]
- Yoder, J.A.; Hedges, B.Z.; Benoit, J.B. Water balance of the American dog tick, Dermacentor variabilis, throughout its development with comparative observations between field-collected and laboratory-reared ticks. Int. J. Acarol. 2012, 38, 334–343. [Google Scholar] [CrossRef]
- Campbell, A.; Harris, D.L. Reproduction of the American dog tick, Dermacentor variabilis, under laboratory and field conditions. Environ. Èntomol. 1979, 8, 734–739. [Google Scholar] [CrossRef]
- Civitello, D.J.; Flory, S.L.; Clay, K. Exotic grass invasion reduces survival of Amblyomma americanum and Dermacentor variabilis ticks (Acari: Ixodidae). J. Med. Èntomol. 2008, 45, 867–872. [Google Scholar] [CrossRef] [PubMed]
- Raghavan, R.K.; Peterson, A.T.; Cobos, M.E.; Ganta, R.; Foley, D. Current and future distribution of the lone star tick, Amblyomma americanum (L.) (Acari: Ixodidae) in north America. PLoS ONE 2019, 14, e0209082. [Google Scholar] [CrossRef] [PubMed]
- Boorgula, G.D.Y.; Peterson, A.T.; Foley, D.H.; Ganta, R.R.; Raghavan, R.K. Assessing the current and future potential geographic distribution of the American dog tick, Dermacentor variabilis (Say) (Acari: Ixodidae) in north America. PLoS ONE 2020, 15, e0237191. [Google Scholar] [CrossRef] [PubMed]
- Raghavan, R.K.; Barker, S.; Cobos, M.E.; Barker, D.; Teo, E.J.M.; Foley, D.H.; Nakao, R.; Lawrence, K.; Heath, A.C.G.; Peterson, A.T. Potential spatial distribution of the newly introduced long-horned tick, Haemaphysalis longicornis in north America. Sci. Rep. 2019, 9, 1–8. [Google Scholar] [CrossRef]
- Randolph, S.E.; Storey, K. Impact of microclimate on immature tick-rodent host interactions (Acari: Ixodidae): Implications for parasite transmission. J. Med. Èntomol. 1999, 36, 741–748. [Google Scholar] [CrossRef]
- Smith, C.N.; Cole, M.M. Effect of length of day on the activity and hibernation of the American dog tick, Dermacentor variabilis (Say) (Acarina: Ixodidae). Ann. Èntomol. Soc. Am. 1941, 34, 426–431. [Google Scholar] [CrossRef]
- Childs, J.E.; Paddock, C.D. The ascendancy of Amblyomma americanum as a vector of pathogens affecting humans in the United States. Annu. Rev. Entomol. 2003, 48, 307–337. [Google Scholar] [CrossRef] [Green Version]
- Savage, H.M.; Godsey, M.S.G., Jr.; Lambert, A.; Panella, N.A.; Burkhalter, K.L.; Harmon, J.R.; Lash, R.R.; Ashley, D.C.; Nicholson, W.L. First detection of Heartland virus (Bunyaviridae: Phlebovirus) from field collected arthropods. Am. J. Trop. Med. Hyg. 2013, 89, 445–452. [Google Scholar] [CrossRef] [PubMed]
- Raghavan, R.K.; Goodin, D.G.; Hanzlicek, G.A.; Zolnerowich, G.; Dryden, M.W.; Anderson, G.A.; Ganta, R.R. Maximum entropy-based ecological niche model and bio-climatic determinants of lone star tick (Amblyomma americanum) niche. Vector Borne Zoonotic Dis. 2016, 16, 205–211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bishopp, F.C.; Trembley, H.L. Distribution and hosts of certain north American ticks. J. Parasitol. 1945, 31. [Google Scholar] [CrossRef]
- Tugwell, P.; Lancaster, J.L., Jr. Notes on the seasonal occurrence of six tick species in northwest Arkansas. J. Kans. Entomol. Soc. 1963, 36, 167–171. [Google Scholar]
- Kocan, K.M.; Hair, J.A.; Ewing, S.A. Ultrastructure of Anaplasma marginale Theiler in Dermacentor andersoni stiles and Dermacentor variabilis (Say). Am. J. Vet. Res. 1980, 41, 1966–1976. [Google Scholar] [PubMed]
- Logan, T.M.; Kocan, K.M.; Edwards, W.; Hair, J.A.; Claypool, P.L.; Ewing, S.A. Persistence of colonies of Anaplasma marginale in overwintering Dermacentor variabilis. Am. J. Vet. Res. 1987, 48. [Google Scholar]
- Stich, R.; Kocan, K.M.; Palmer, G.H.; Ewing, S.A.; Hair, J.A.; Barron, S.J. Transstadial and attempted transovarial transmission of Anaplasma marginale by Dermacentor variabilis. Am. J. Vet. Res. 1989, 50, 1377–1380. [Google Scholar]
- Coley, K. Identification guide to larval stages of ticks of medical importance in the USA. University Honors. Program Thesis, Georgia Southern University, Statesboro, GA, USA, 2015. [Google Scholar]
- Stich, R.W.; Blagburn, B.L.; Bowman, D.D.; Carpenter, C.; Cortinas, M.R.; Ewing, S.A.; Foley, D.; Foley, J.E.; Gaff, H.; Hickling, G.J.; et al. Quantitative factors proposed to influence the prevalence of canine tick-borne disease agents in the United States. Parasites Vectors 2014, 7, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Knudson, C. An Introduction to Model-Fitting with the R Package GLMM. Available online: https://mran.microsoft.com/snapshot/2015-04-08/web/packages/glmm/vignettes/intro.pdf (accessed on 6 August 2021).
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2018; Available online: https://www.R-project.org/ (accessed on 6 August 2021).

{kind=link}
{kind=link}
| Species | Year | Male | Female | Nymph | Larvae | Total |
|---|---|---|---|---|---|---|
| Amblyomma americanum | Year 1 | 518 | 782 | 1265 | 1309 | 3874 |
| Year 2 | 506 | 887 | 1447 | 1369 | 4209 | |
| Amblyomma maculatum | Year 1 | 222 | 269 | - | - | 491 |
| Year 2 | 203 | 271 | - | - | 474 | |
| Dermacentor variabilis | Year 1 | 321 | 450 | - | - | 771 |
| Year 2 | 286 | 424 | - | - | 710 | |
| Ixodes scapularis | Year 1 | 2 | 6 | - | - | 8 |
| Year 2 | 5 | 15 | - | - | 20 |
| Covariate | Estimate | Std. Error | Pr (>|z|) | 95% CI |
|---|---|---|---|---|
| Accumulated temperature | 1.232 | 0.012 | 0.000 | 1.209, 1.256 |
| Photoperiod | 1.079 | 0.031 | 0.000 | 1.019, 1.140 |
| NDVI | 0.121 | 0.010 | 0.017 | 0.101, 0.1406 |
| Location | 1.553 | 0.496 | 0.015 | 0.580, 2.525 |
| Month | 0.811 | 0.213 | 0.007 | 0.039, 1.228 |
| Covariate | Estimate | Std. Error | Pr (>|z|) | 95% CI |
|---|---|---|---|---|
| Accumulated temperature | 1.421 | 0.119 | 0.000 | 1.877, 1.654 |
| Photoperiod | 1.549 | 0.088 | 0.001 | 1.376, 1.721 |
| Saturation deficit | −1.388 | 0.192 | 0.038 | −1.764, −1.011 |
| Location | 1.321 | 0.622 | 0.048 | 0.101, 2.540 |
| Month | 0.422 | 0.183 | 0.001 | 0.063, 0.780 |
| Covariate | Estimate | Std. Error | Pr (>|z|) | 95% CI |
|---|---|---|---|---|
| Accumulated temperature | 2.076 | 0.083 | 0.000 | 1.913, 2.238 |
| Photoperiod | 0.373 | 0.108 | 0.000 | 0.165, 0.588 |
| Mean relative humidity | −0.274 | 0.187 | 0.041 | −0.640, 0.092 |
| Location | 1.118 | 0.288 | 0.038 | 0.553, 1.682 |
| Month | 1.027 | 0.261 | 0.001 | 0.514, 1.538 |
| Species | Target Gene | Product Size | Primes /Probes | Sequence (5’-3’) |
|---|---|---|---|---|
| Amblyomma americanum | 16S rRNA | 150bp | * FP * RP * Pr | TTTAATTGGGGCGATTTAACTA CATCGAGGTCGCAAACTATT FAM-GAACCGTTATTAACGGACACTTGGA-BHQ-1 |
| Amblyomma maculatum | 16S rRNA | 178bp | * FP * RP * Pr | AAGGACAAGAAGACCCTAAGAATTT ATTACGCTGTTATCCCTAGAGTATTT CAL Fluor Red 610-TGAAATTTTTTAATTGGGGCGA-BHQ-2 |
| Dermacentor variabilis | 16S rRNA | 138bp | *FP * RP * Pr | TGGTATTTTGACTATACAAAGGTATT CCTTAATTTTAATAATTGTTTCTTCAC CAL Fluor Gold 540- TGCTAAGAGAATGGAATTACAGGGAATA-BHQ-1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Spare, M.; Boorgula, G.D.; Thomson, D.; Bai, J.; Goodin, D.; Anderson, G.; Stich, R.W.; Hroobi, A.; Wilson, S.; Staggs, A.; et al. Surveillance of Host-Seeking Ticks in the Flint Hills Region (USA) and Associations with Environmental Determinants. Parasitologia 2021, 1, 137-147. https://doi.org/10.3390/parasitologia1030015
Spare M, Boorgula GD, Thomson D, Bai J, Goodin D, Anderson G, Stich RW, Hroobi A, Wilson S, Staggs A, et al. Surveillance of Host-Seeking Ticks in the Flint Hills Region (USA) and Associations with Environmental Determinants. Parasitologia. 2021; 1(3):137-147. https://doi.org/10.3390/parasitologia1030015
Chicago/Turabian StyleSpare, Mark, Gunavanthi D. Boorgula, Dan Thomson, Jianfa Bai, Doug Goodin, Gary Anderson, Roger W. Stich, Ali Hroobi, Savannah Wilson, Alexander Staggs, and et al. 2021. "Surveillance of Host-Seeking Ticks in the Flint Hills Region (USA) and Associations with Environmental Determinants" Parasitologia 1, no. 3: 137-147. https://doi.org/10.3390/parasitologia1030015
APA StyleSpare, M., Boorgula, G. D., Thomson, D., Bai, J., Goodin, D., Anderson, G., Stich, R. W., Hroobi, A., Wilson, S., Staggs, A., Bowers, A., Hamm, A., Tatman, J., & Raghavan, R. K. (2021). Surveillance of Host-Seeking Ticks in the Flint Hills Region (USA) and Associations with Environmental Determinants. Parasitologia, 1(3), 137-147. https://doi.org/10.3390/parasitologia1030015