
Exploring the Effect of the COVID-19 Zoo Closure Period on Flamingo Behaviour and Enclosure Use at Two Institutions



Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
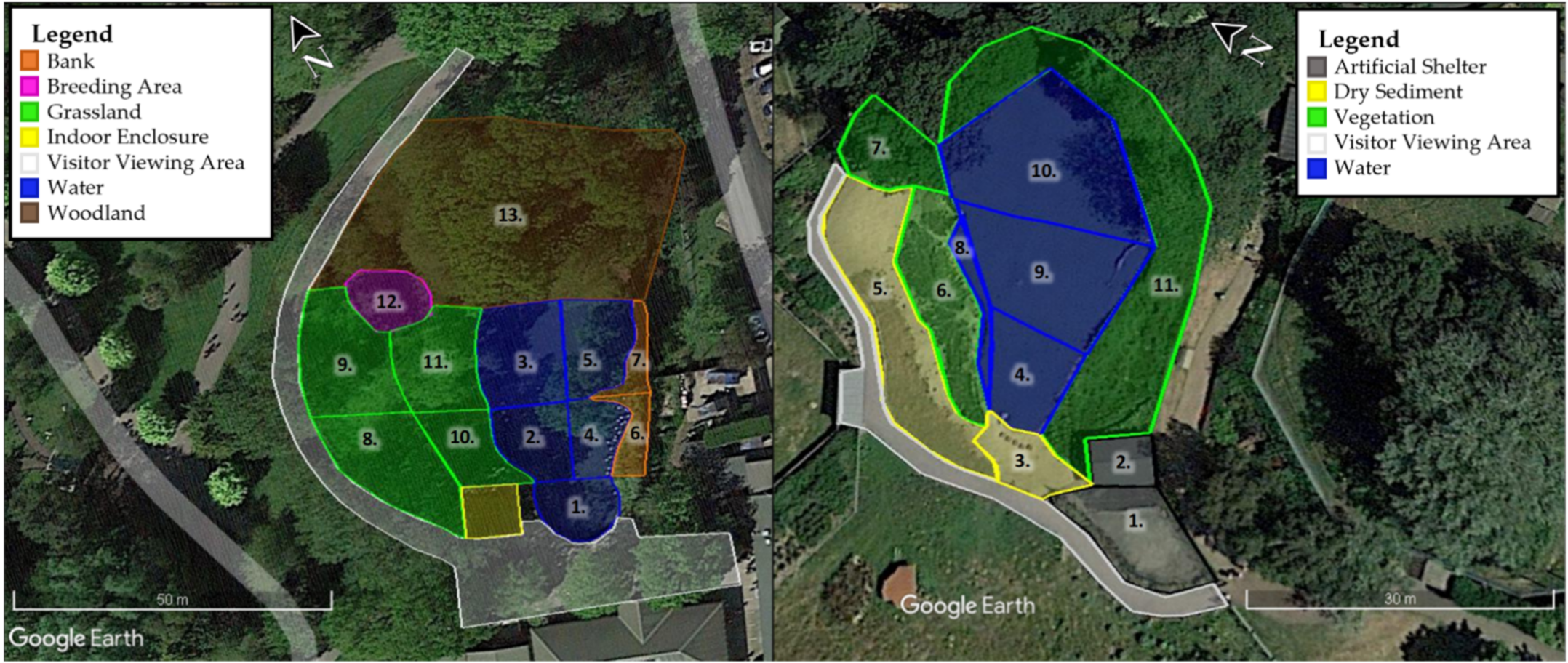
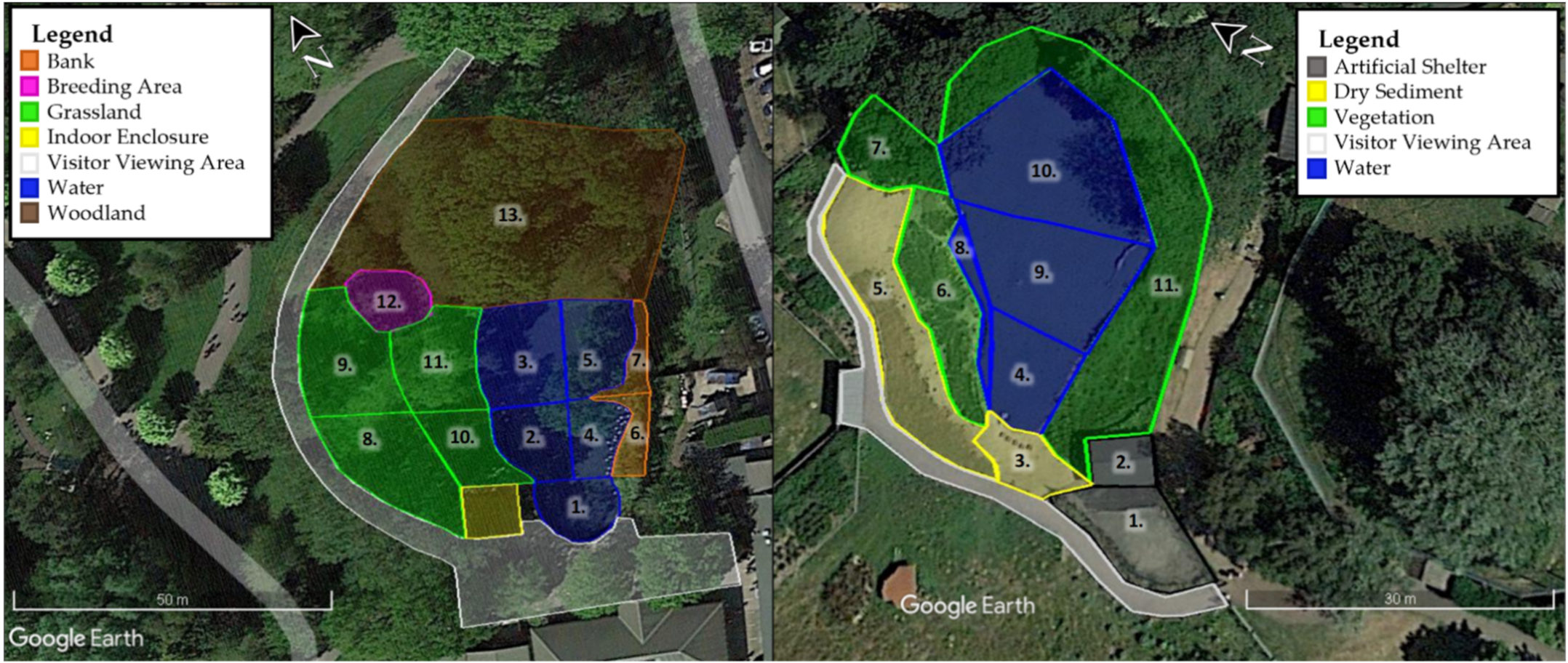
2.1. Study Species and Study Sites
2.2. Data Collection
2.3. Image Processing
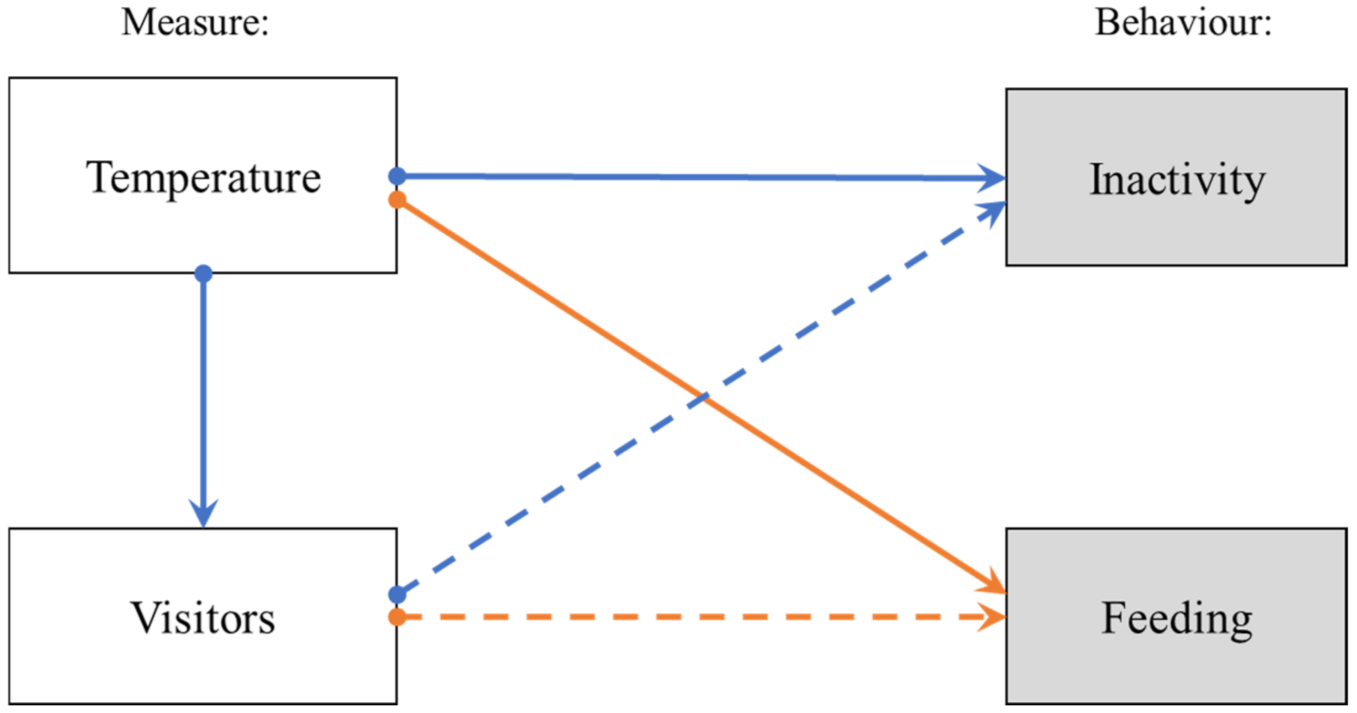
2.4. Statistical Analysis
3. Results
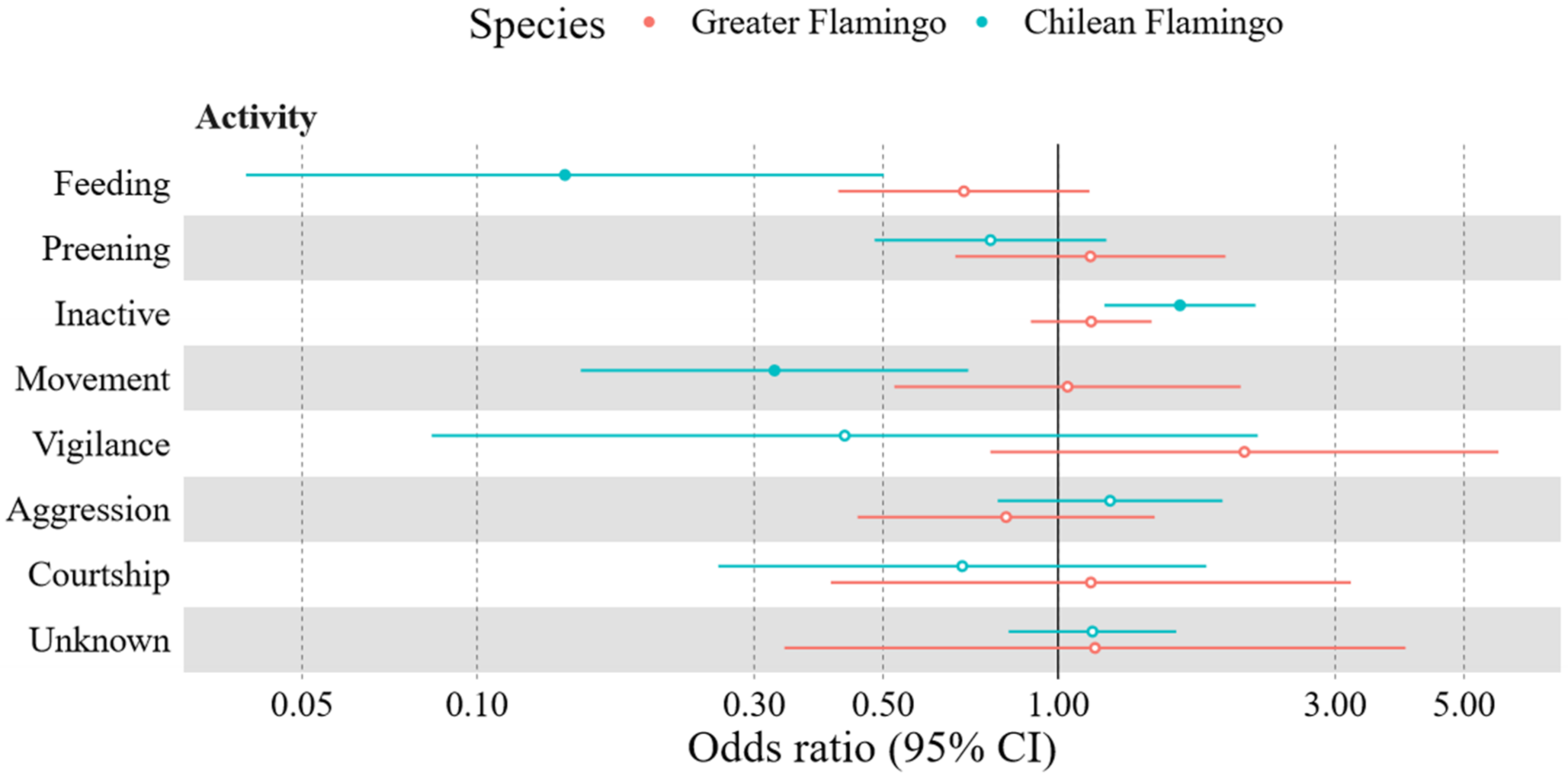
3.1. Flamingo Behaviour
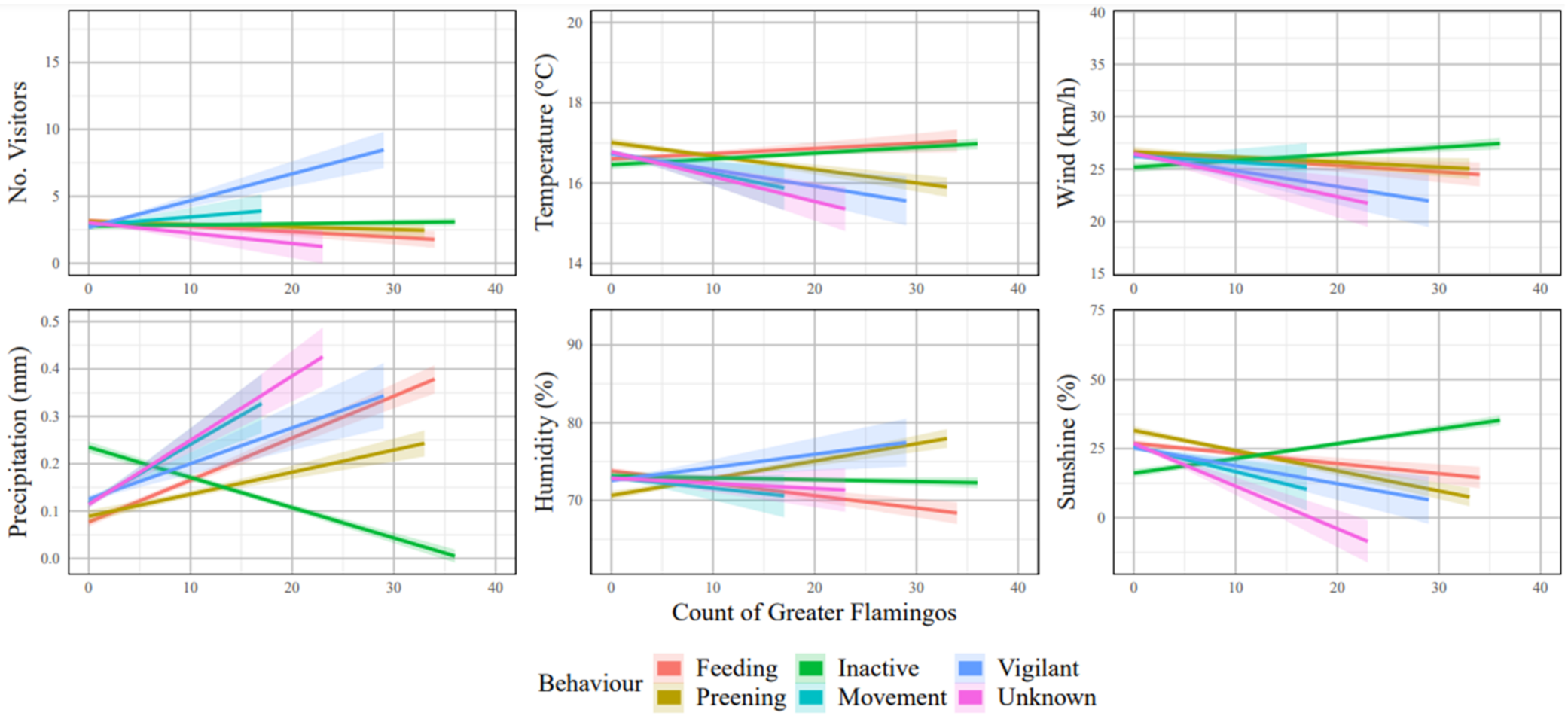
3.1.1. Greater Flamingo Behaviour
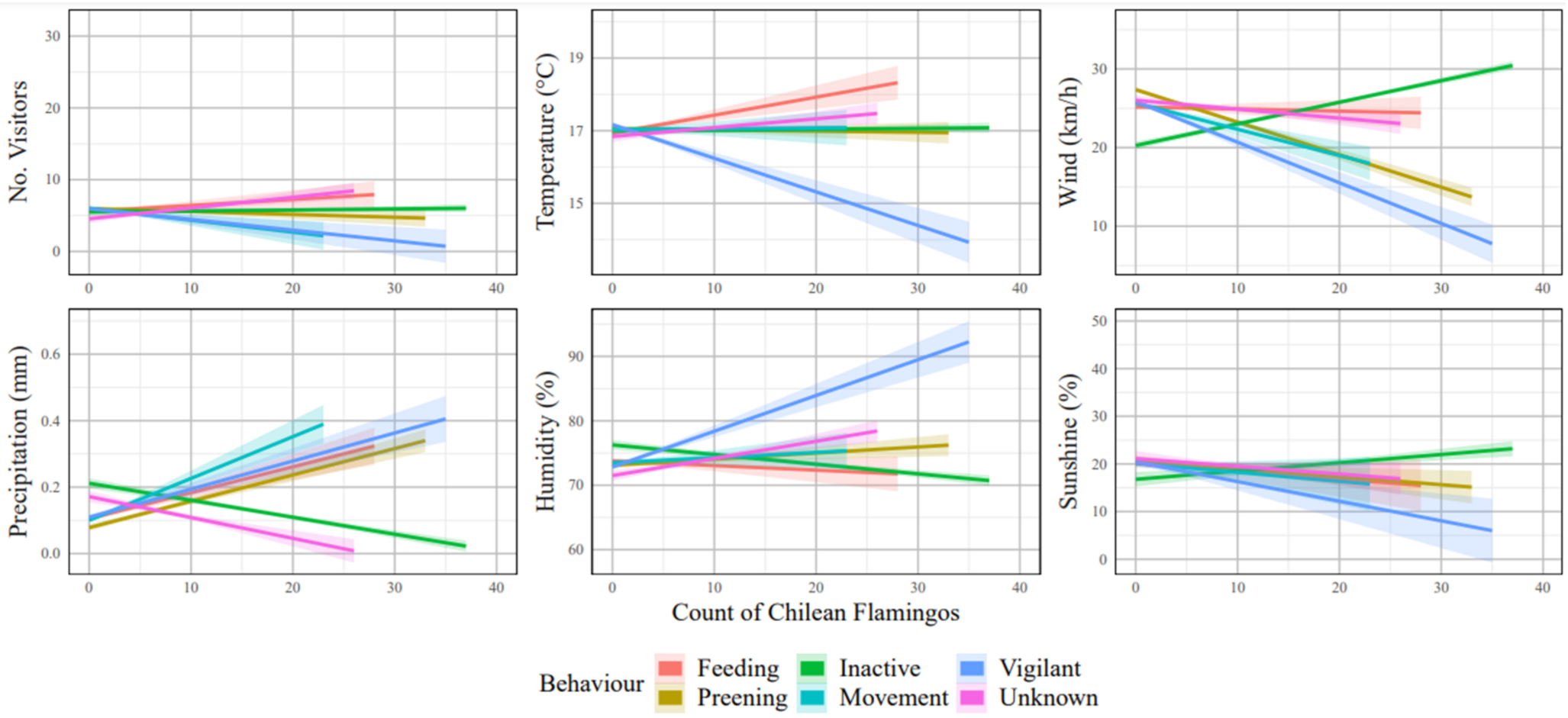
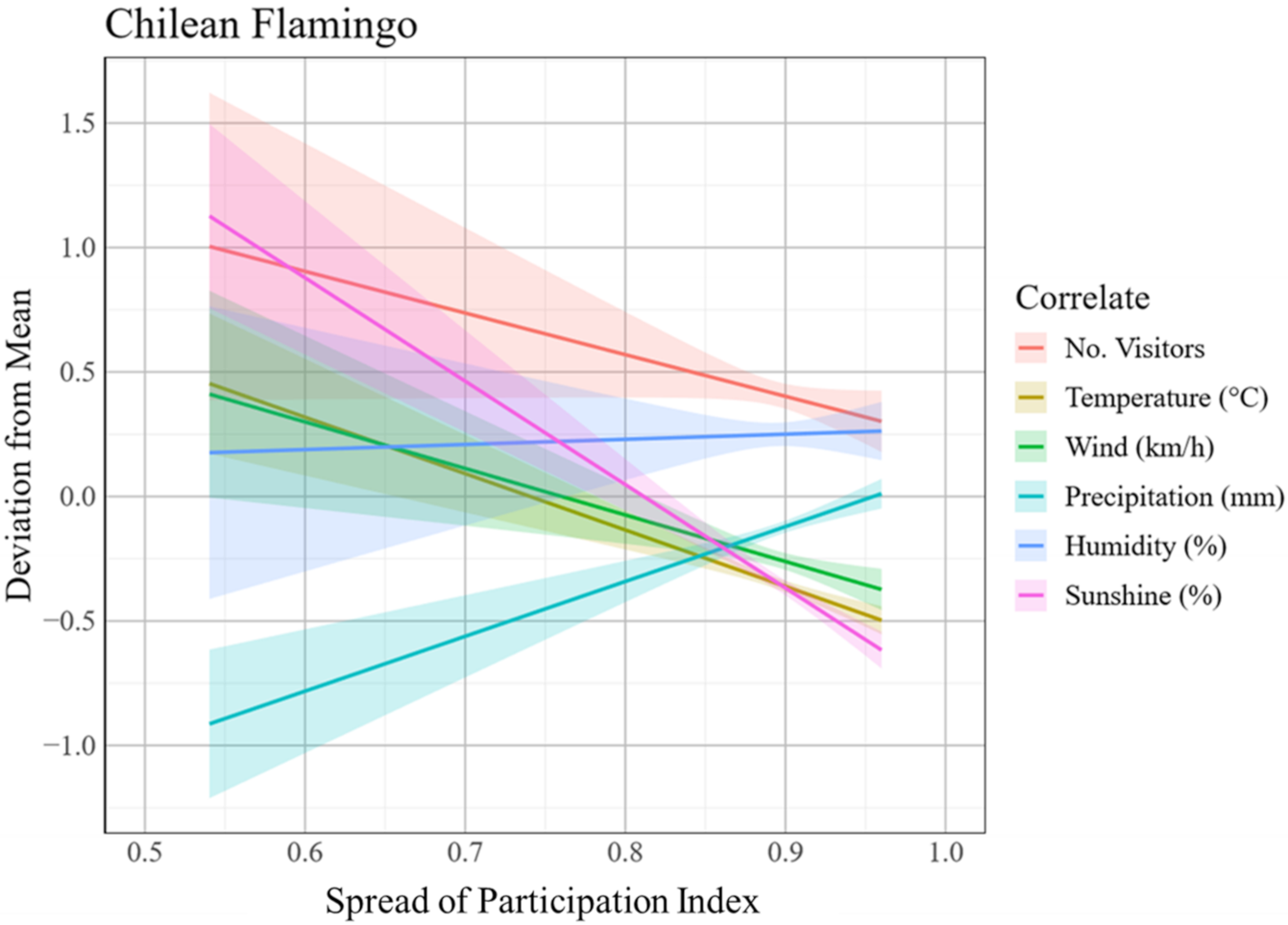
3.1.2. Chilean Flamingo Behaviour
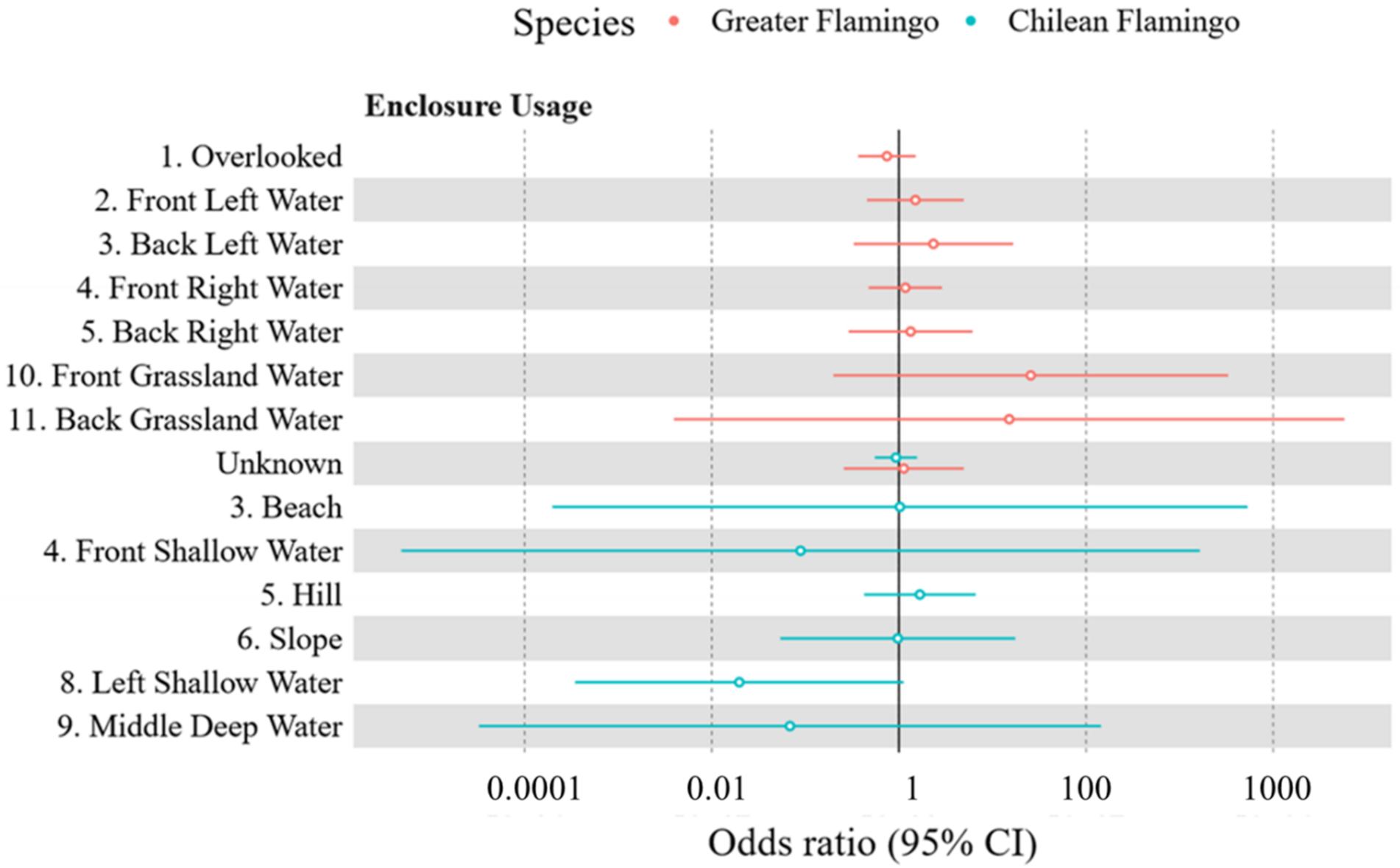
3.2. Flamingo Enclosure Usage
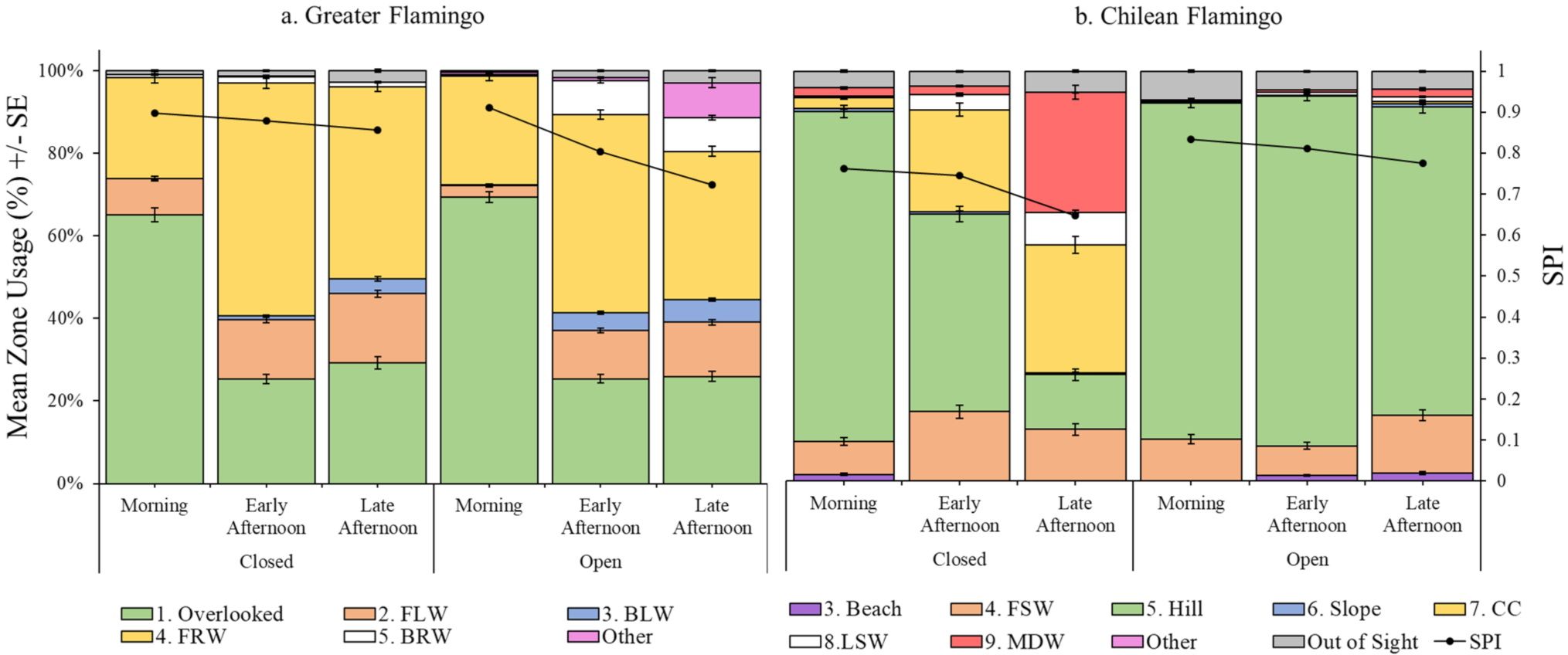
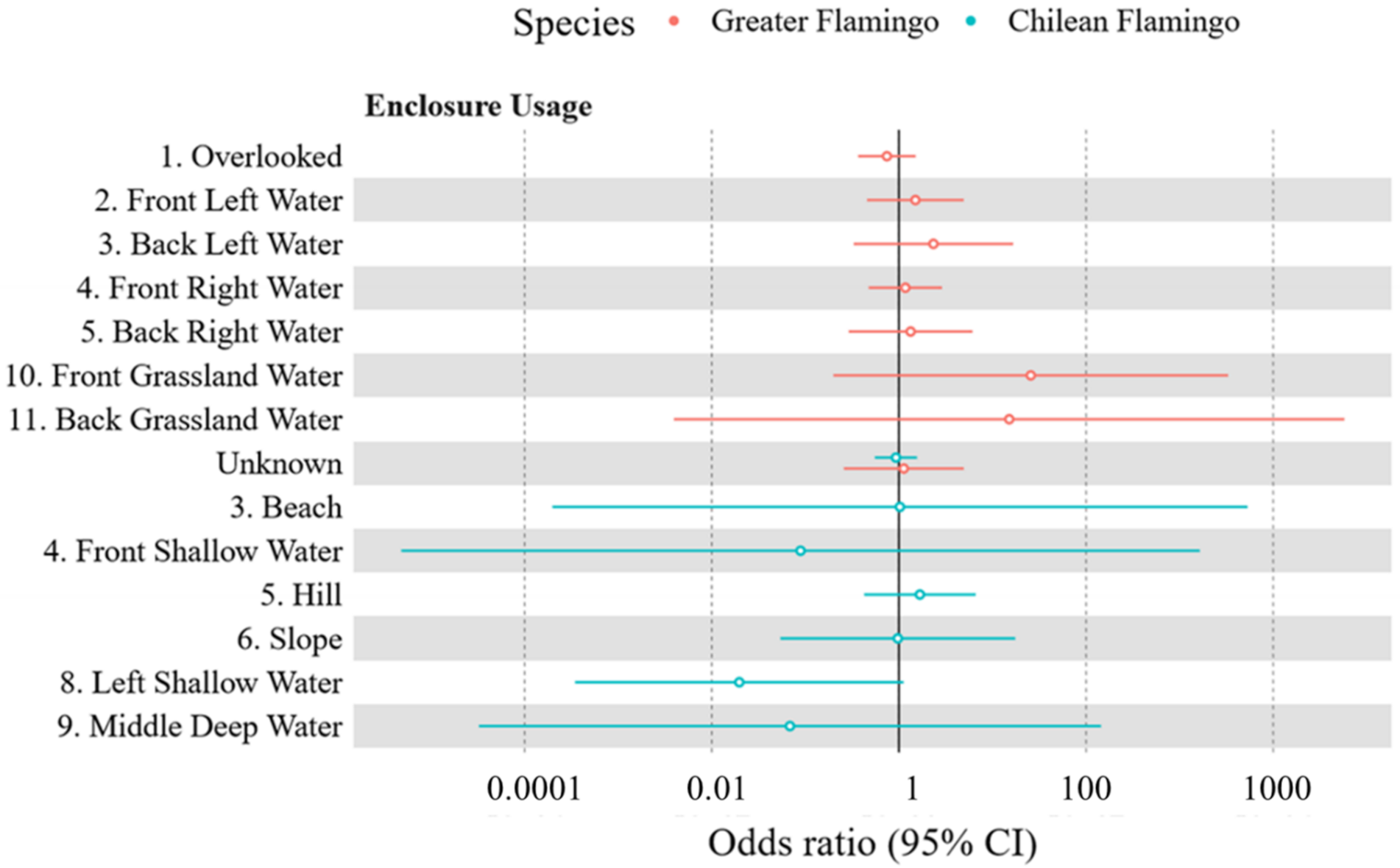
3.2.1. Greater Flamingo Enclosure Usage
3.2.2. Chilean Flamingo Enclosure Usage
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Davey, G. Visitors’ effects on the welfare of animals in the zoo: A review. J. Appl. Anim. Welf. Sci. 2007, 10, 169–183. [Google Scholar] [CrossRef]
- Fernandez, E.J.; Tamborski, M.A.; Pickens, S.R.; Timberlake, W. Animal-visitor interactions in the modern zoo: Conflicts and interventions. Appl. Anim. Behav. Sci. 2009, 120, 1–8. [Google Scholar] [CrossRef]
- Quadros, S.; Goulart, V.D.L.; Passos, L.; Vecci, M.A.M.; Young, R.J. Zoo visitor effect on mammal behaviour: Does noise matter? Appl. Anim. Behav. Sci. 2014, 156, 78–84. [Google Scholar] [CrossRef]
- Williams, E.; Carter, A.; Rendle, J.; Ward, S.J. Understanding impacts of zoo visitors: Quantifying behavioural changes of two popular zoo species during COVID-19 closures. Appl. Anim. Behav. Sci. 2021, 236, 105253. [Google Scholar] [CrossRef]
- Hosey, G. A preliminary model of human–animal relationships in the zoo. Appl. Anim. Behav. Sci. 2008, 109, 105–127. [Google Scholar] [CrossRef] [Green Version]
- Sherwen, S.L.; Hemsworth, P.H. The visitor effect on zoo animals: Implications and opportunities for zoo animal welfare. Animals 2019, 9, 366. [Google Scholar] [CrossRef] [Green Version]
- Rose, P.E.; Badman-King, A.; Hurn, S.; Rice, T. Visitor presence and a changing soundscape, alongside environmental parameters, can predict enclosure usage in captive flamingos. Zoo Biol. 2021, 40, 363–375. [Google Scholar] [CrossRef]
- Sulser, C.E.; Steck, B.L.; Baur, B. Effects of construction noise on behaviour of and exhibit use by Snow leopards Uncia uncia at Basel Zoo. Int. Zoo Yearb. 2008, 42, 199–205. [Google Scholar] [CrossRef]
- Ross, S.R. Issues of choice and control in the behaviour of a pair of captive polar bears (Ursus maritimus). Behav. Processes 2006, 73, 117–120. [Google Scholar] [CrossRef] [PubMed]
- Ozella, L.; Favaro, L.; Carnovale, I.; Pessani, D. Pond use by captive African penguins (Spheniscus demersus) in an immersive exhibit adjacent to human bathers. J. Appl. Anim. Welf. Sci. 2015, 18, 303–309. [Google Scholar] [CrossRef]
- Collins, C.K.; Quirke, T.; Overy, L.; Flannery, K.; O’Riordan, R. The effect of the zoo setting on the behavioural diversity of captive gentoo penguins and the implications for their educational potential. J. Zoo Aquar. Res. 2016, 4, 85–90. [Google Scholar]
- Blanchett, M.K.S.; Finegan, E.; Atkinson, J. The effects of increasing visitor and noise levels on birds within a free-flight aviary examined through enclosure use and behavior. Anim. Behav. Cogn. 2020, 7, 49–69. [Google Scholar] [CrossRef]
- Rose, P.E.; Scales, J.S.; Brereton, J.E. Why the “visitor effect” is complicated. Unraveling individual animal, visitor number, and climatic influences on behavior, space use and interactions with keepers—A case study on captive hornbills. Front. Vet. Sci. 2020, 7, 236. [Google Scholar] [CrossRef] [PubMed]
- Rose, P.E.; Brereton, J.E.; Croft, D.P. Measuring welfare in captive flamingos: Activity patterns and exhibit usage in zoo-housed birds. Appl. Anim. Behav. Sci. 2018, 205, 115–125. [Google Scholar] [CrossRef]
- De Azevedo, C.S.; Lima, M.F.F.; da Silva, V.C.A.; Young, R.J.; Rodrigues, M. Visitor influence on the behavior of captive greater rheas (Rhea americana, Rheidae Aves). J. Appl. Anim. Welf. Sci. 2012, 15, 113–125. [Google Scholar] [CrossRef]
- Sherwen, S.L.; Magrath, M.J.L.; Butler, K.L.; Hemsworth, P.H. Little penguins, Eudyptula minor, show increased avoidance, aggression and vigilance in response to zoo visitors. Appl. Anim. Behav. Sci. 2015, 168, 71–76. [Google Scholar] [CrossRef]
- Williams, E.; Carter, A.; Rendle, J.; Ward, S.J. Impacts of COVID-19 on animals in zoos: A longitudinal multi-species analysis. J. Zool. Bot. Gard. 2021, 2, 130–145. [Google Scholar] [CrossRef]
- Rose, P.E.; Brereton, J.E.; Rowden, L.J.; de Figueiredo, R.L.; Riley, L.M. What’s new from the zoo? An analysis of ten years of zoo-themed research output. Palgrave Commun. 2019, 5, 128. [Google Scholar] [CrossRef] [Green Version]
- Rose, P.E. Investigating the behaviour and welfare of captive flamingos (Phoenicopterformes). In College of Life & Environmental Sciences; University of Exeter: Exeter, UK, 2018. [Google Scholar]
- Mascitti, V.; Kravetz, F.O. Bill morphology of South American flamingos. Condor 2002, 104, 73–83. [Google Scholar] [CrossRef]
- Johnson, A.; Cézilly, F. The Greater Flamingo; Bloomsbury Publishing: London, UK, 2009. [Google Scholar]
- BirdLife International. Phoenicopterus roseus (Amended Version of 2018 Assessment). Available online: https://dx.doi.org/10.2305/IUCN.UK.2018-2.RLTS.T22697360A155527405.en (accessed on 7 July 2021).
- Kear, J.; Duplaix, N. (Eds.) Flamingos; T&AD Poyser: Berkhamsted, UK, 1975. [Google Scholar]
- BirdLife International. Phoenicopterus chilensis. Available online: https://doi.org/10.2305/IUCN.UK.2018-2.RLTS.T22697365A132068236.en (accessed on 7 July 2021).
- Gordo, O.; Brotons, L.; Herrando, S.; Gargallo, G. Rapid behavioural response of urban birds to COVID-19 lockdown. Proc. R. Soc. B 2021, 288, 20202513. [Google Scholar] [CrossRef]
- Seress, G.; Sándor, K.; Vincze; Pipoly, I.; Bukor, B.; Ágh, N.; Liker, A. Contrasting effects of the COVID-19 lockdown on urban birds’ reproductive success in two cities. Sci. Rep. 2021, 11, 17649. [Google Scholar] [CrossRef]
- Bateson, M.; Martin, P. Measuring Behaviour: An Introductory Guide, 4th ed.; Cambridge University Press: Cambridge, UK, 2021. [Google Scholar]
- World Weather Online. World Weather. Available online: https://www.worldweatheronline.com/ (accessed on 9 July 2020).
- Rose, P.E. Evaluating the behaviour of Andean Flamingos Phoenicoparrus andinus and James’s Flamingos, P. jamesi in captivity: Comparing species and flocks using multiple methods. Wildfowl 2019, 69, 70–92. [Google Scholar]
- Plowman, A.B. A note on a modification of the spread of participation index allowing for unequal zones. Appl. Anim. Behav. Sci. 2003, 83, 331–336. [Google Scholar] [CrossRef]
- RStudio Team. RStudio: Integrated Development for R. Available online: http://www.rstudio.com (accessed on 10 July 2021).
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting linear mixed-effects models using lme4. J. Stat. Softw. 2014, 67, 1–48. [Google Scholar]
- Lüdecke, D.; Ben-Shachar, M.S.; Patil, I.; Waggoner, P.; Makowski, D. Performance: An R package for assessment, comparison and testing of statistical models. J. Open Source Softw. 2021, 6, 3139. [Google Scholar] [CrossRef]
- Barton, K. Mu-Min-Model Interference. Available online: https://cran.r-project.org/web/packages/MuMIn/MuMIn.pdf (accessed on 1 July 2021).
- Szumilas, M. Explaining odds ratios. J. Can. Acad. Child Adolesc. Psychiatry 2010, 19, 227. [Google Scholar]
- Delfino, H.C.; Carlos, C.J. What do we know about flamingo behaviors? A systematic review of the ethological research on the Phoenicopteridae (1978–2020). Acta Ethol. 2022, 25, 1–14. [Google Scholar] [CrossRef]
- Naz, S.; Tanveer, A.; Abrar, M.; Ashraf, A. Behavioral activity patterns of adult and juvenile greater flamingos (Phoenicopterus roseus) with alteration of climatic pattern at Uchalli Lake, Punjab, Pakistan. Environ. Sci. Pollut. Res. 2018, 25, 30132–30138. [Google Scholar] [CrossRef] [PubMed]
- Rose, P.E.; Croft, D.P. Social bonds in a flock bird: Species differences and seasonality in social structure in captive flamingo flocks over a 12-month period. Appl. Anim. Behav. Sci. 2017, 193, 87–97. [Google Scholar] [CrossRef]
- Rose, P.E.; Croft, D.P. Evaluating the social networks of four flocks of captive flamingos over a five-year period: Temporal, environmental, group and health influences on assortment. Behav. Processes 2020, 175, 104118. [Google Scholar] [CrossRef]
- Rose, P.E. Evidence for aviculture: Identifying research needs to advance the role of ex situ bird populations in conservation initiatives and collection planning. Birds 2021, 2, 77–95. [Google Scholar] [CrossRef]
- Kidd, P.; Rose, P.E. Influences of rearing environment on behaviour and welfare of captive Chilean flamingos: A case study on foster-reared and parent-reared birds. J. Zool. Bot. Gard. 2021, 2, 174–206. [Google Scholar] [CrossRef]
- Riley, A.; Terry, M.; Freeman, H.; Alba, A.C.; Soltis, J.; Leeds, A. Evaluating the effect of visitor presence on Nile crocodile (Crocodylus niloticus) behavior. J. Zool. Bot. Gard. 2021, 2, 115–129. [Google Scholar] [CrossRef]
- Sherwen, S.L.; Magrath, M.J.L.; Butler, K.L.; Phillips, C.J.C.; Hemsworth, P.H. A multi-enclosure study investigating the behavioural response of meerkats to zoo visitors. Appl. Anim. Behav. Sci. 2014, 156, 70–77. [Google Scholar] [CrossRef]
- Collins, C.K.; Corkery, I.; Haigh, A.; McKeown, S.; Quirke, T.; O’Riordan, R. The effects of environmental and visitor variables on the behavior of free-ranging ring-tailed lemurs (Lemur catta) in captivity. Zoo Biol. 2017, 36, 250–260. [Google Scholar] [CrossRef] [PubMed]
- Woolway, E.E.; Goodenough, A.E. Effects of visitor numbers on captive European red squirrels (Sciurus vulgaris) and impacts on visitor experience. Zoo Biol. 2017, 36, 112–119. [Google Scholar] [CrossRef]
- Margulis, S.W.; Hoyos, C.; Anderson, M.J. Effect of felid activity on zoo visitor interest. Zoo Biol. 2003, 22, 587–599. [Google Scholar] [CrossRef]
- Galicia, E.; Baldassarre, G.A. Effects of motorized tourboats on the behavior of nonbreeding American flamingos in Yucatan, Mexico. Conserv. Biol. 1997, 11, 1159–1165. [Google Scholar] [CrossRef]
- Jezierski, T.; Jaworski, Z.; Górecka, A. Effects of handling on behaviour and heart rate in Konik horses: Comparison of stable and forest reared youngstock. Appl. Anim. Behav. Sci. 1999, 62, 1–11. [Google Scholar] [CrossRef]
- Rose, P.E. The relevance of captive flamingos to meeting the four aims of the modern zoo. Flamingo 2018, e1, 23–32. [Google Scholar]
- Burger, J. The effect of human disturbance on foraging behavior and habitat use in piping plover (Charadrius melodus). Estuaries 1994, 17, 695–701. [Google Scholar] [CrossRef]
- Burger, J.; Gochfeld, M. Effects of ecotourists on bird behaviour at Loxahatchee National Wildlife Refuge, Florida. Environ. Conserv. 1998, 25, 13–21. [Google Scholar] [CrossRef]
- Burger, J.; Gochfeld, M. Effect of human presence on foraging behavior of sandhill cranes (Grus canadensis) in Nebraska. Bird Behav. 2001, 14, 81–87. [Google Scholar]
- Burger, J.; Niles, L. Effects on five species of shorebirds of experimental closure of a beach in New Jersey: Implications for severe storms and sea-level rise. J. Toxicol. Environ. Health Part A 2014, 77, 1102–1113. [Google Scholar] [CrossRef]
- Verhulst, S.; Oosterbeek, K.; Ens, B.J. Experimental evidence for effects of human disturbance on foraging and parental care in oystercatchers. Biol. Conserv. 2001, 101, 375–380. [Google Scholar] [CrossRef]
- Brooking, Z.A.; Price, D.J. The effect of human presence on the behavioural and distributional patterns of two species of captive penguins, African (Spheniscus demersus) and gentoo (Pygoscelis papua), on being transferred into their new enclosure at Living Coasts, Devon. In Proceedings of the 6th Annual Symposium on Zoo Research, Edinburgh, UK, 8–9 July 2004. [Google Scholar]
- Boultwood, J.; O’Brien, M.; Rose, P.E. Bold frogs or shy toads? How did the COVID-19 closure of zoological organisations affect amphibian activity? Animals 2021, 11, 1982. [Google Scholar] [CrossRef]
- Rose, P.E.; Lloyd, I.; Brereton, J.E.; Croft, D.P. Patterns of nocturnal activity in captive greater flamingos. Zoo Biol. 2018, 37, 290–299. [Google Scholar] [CrossRef]
- Britton, R.H.; de Groot, E.R.; Johnson, A.R. The daily cycle of feeding activity of the Greater Flamingo in relation to the dispersion of the prey Artemia. Wildfowl 1986, 37, 151–155. [Google Scholar]
- Beauchamp, G.; McNeil, R. Vigilance in greater flamingos foraging at night. Ethology 2003, 109, 511–520. [Google Scholar] [CrossRef]
- Tindle, R.W.; Tupiza, A.; Blomberg, S.P.; Tindle, L.E. The biology of an isolated population of the American flamingo Phoenicopterus ruber in the Galapagos Islands. Galapagos Res. 2016, 68, 15–27. [Google Scholar]
- Bildstein, K.L.; Frederick, P.C.; Spalding, M.G. Feeding patterns and aggressive behavior in juvenile and adult American flamingos. Condor 1991, 93, 916–925. [Google Scholar] [CrossRef]
- Robinson, V.J. The ecology of East African soda lakes: Implications for lesser flamingo (Phoeniconaias minor) feeding behaviours. In Department of Biology; University of Leicester: Leicester, UK, 2015. [Google Scholar]
- Rose, P.E.; Soole, L. What influences aggression and foraging activity in social birds? Measuring individual, group and environmental characteristics. Ethology 2020, 126, 900–913. [Google Scholar]
- Kalejta-Summers, B.; McCarthy, M.; Underhill, L.G. Long-term trends, seasonal abundance and energy consumption of waterbirds at Strandfontein, Western Cape, South Africa, 1953–1993. Ostrich 2001, 72, 80–95. [Google Scholar] [CrossRef]
- Jiang, X.; Liu, J.; Zhang, C.; Liang, W. Face masks matter: Eurasian tree sparrows show reduced fear responses to people wearing face masks during the COVID-19 pandemic. Glob. Ecol. Conserv. 2020, 24, e01277. [Google Scholar] [CrossRef]
- Mikula, P.; Jokimäki, J.; Kaisanlahti-Jokimäki, M.-L.; Markó, G.; Morelli, F.; Møller, A.P.; Szakony, S.; Yosef, R.; Albrecht, T.; Tryjanowski, P. Face mask-wear did not affect large-scale patterns in escape and alertness of urban and rural birds during the COVID-19 pandemic. Sci. Total Environ. 2021, 793, 148672. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Study Enclosure Site (Size) | Study Species (N) | Period of Data Collection | ||
|---|---|---|---|---|
| Zoo Closed | Zoo Open | |||
| Dates | Dates | Gate Admissions (Zoo Open Dates Only) | ||
| Africa Alive, Suffolk, UK (2868 m2) | Greater Flamingo (N = 36) | 25/06/2020 | 29/06/2020 | 200 |
| 26/06/2020 | 01/07/2020 | 563 | ||
| 27/06/2020 | 02/07/2020 | 384 | ||
| 28/06/2020 | 03/07/2020 | 544 | ||
| 30/06/2020 | 04/07/2020 | 1032 | ||
| 06/07/2020 | 693 | |||
| 07/07/2020 | 746 | |||
| 08/07/2020 | 558 | |||
| Banham Zoo, Norfolk, UK (2118 m2) | Chilean Flamingo (N = 37) | 24/06/2020 | 28/06/2020 | 283 |
| 25/06/2020 | 01/07/2020 | 904 | ||
| 26/06/2020 | 02/07/2020 | 593 | ||
| 27/06/2020 | 03/07/2020 | 1032 | ||
| 29/06/2020 | 04/07/2020 | 1754 | ||
| 30/06/2020 | 06/07/2020 | 1296 | ||
| 07/07/2020 | 1097 | |||
| 08/07/2020 | 868 | |||
| Behaviour | Definition |
|---|---|
| Preening | Cleaning feathers with bill or water by scooping water over the body with wings and or bill, either sitting or standing. Flamingo uses feet or head to move plumage behind its head. |
| Foraging | Trough feeding (Consumption of food from feeders provided by zoo: head is either inside trough or directly above trough); Natural feeding (Moving the bill through the water from side to side or picking food off the ground with bill. Can occur either whilst still or moving. Flamingo is extending its neck down to the floor, either whilst standing, walking, or sitting) |
| Inactive | Rest (Motionless with head tucked under wing, standing, or sitting, with eye(s) open or closed); Standing (motionless, not vigilant, head is held in front of body. General inactivity, with neck in a relaxed ‘S’ shape. Bill is pointed at the ground); Sitting (Flamingo is sitting motionless, not vigilant. Eyes are open, legs are tucked under body and neck is not erect). |
| Aggression | Spreading the scapular feathers to look more threatening, either whilst sitting or standing; Hooking and/or jousting (extension of the neck either fully straight or in an extended S-bend, and pointing of the bill at a nearby bird with the head swayed side to side and engaging in direct contact, spreading of scapular feathers; Fighting (birds push and shove one-another, using wings and beaks, either sitting or standing). |
| Courtship | Head flagging (neck straight and erect, head is held above a 90 degree ankle, jerked from side to side quickly); Marching displays (birds pack closely together and move quickly in an exaggerated fashion with straight heads and necks); Wing salute (wings are opened quickly and then snapped shut, whilst standing.); Wing-leg stretch (one wing is outstretched along the leg, which is also being stretched on that side of the body); Twist preen (wing is opened up and outwards and to the side, but not fully extended and the head and bill are placed behind the opened wing as if preening its black primary feathers.); Mating attempt (copulation between male and female birds). |
| Movement | Running/walking (bipedal movement along the ground at a hurried /gentle pace. Identifiable as the ‘peeling’ of one foot off the ground, or the foot in mid-air extended forward. Includes wading through water. If running, wings may also be open); Swimming (movement across the water similar to a duck); Social following (positive social interaction where one flamingo follows another directly behind across the enclosure). |
| Vigilance | Neck held in an erect ‘S’ shape or fully extended, whilst visually scanning surroundings, either sitting or standing. Bill is held up slightly compared to relaxed standing posture. |
| Unknown | Bird is out of sight or performing a behaviour not described in this ethogram. |
| Correlates | Greater Flamingo | Chilean Flamingo | ||
|---|---|---|---|---|
| No. Visitors | No. Visitors | |||
| rho | p | rho | p | |
| Temp | 0.123 | <0.001 | 0.129 | <0.001 |
| Wind | 0.032 | 0.167 | 0.084 | <0.001 |
| Precipitation | 0.100 | <0.001 | −0.167 | <0.001 |
| Humidity | 0.129 | <0.001 | 0.230 | <0.001 |
| Sunshine | −0.084 | <0.001 | −0.206 | <0.001 |
| Flock | Did Zoo Reopening Affect Behaviour? | Did Zoo Reopening Affect Enclosure Usage? | Did Increasing Visitor Numbers Show a Relationship with Behaviour? | Did Increasing Visitor Numbers Show a Relationship with Enclosure Usage? | Did Increasing Visitor Numbers Show a Relationship with Weather? | Did Flamingo Behaviour Show Any Relationship with Weather? | Did Flamingo Enclosure Usage Show Any Relationship with Weather? |
|---|---|---|---|---|---|---|---|
| Greater Flamingo | No. | No. | No. | No. | Yes, with temperature, precipitation, and humidity. | Yes, for feeding, preening, inactivity, movement, and vigilance. | Yes, for all zones analysed. |
| Chilean Flamingo | Yes, for inactivity, movement, and feeding. Inactivity increased, while movement and feeding decreased when zoo reopened. | No. | Yes, weak negative association with vigilance. Relationship maintained when controlling for weather. | No, weak negative association with occupancy in zone 6 disappeared when controlling for weather. | Yes, with temperature, precipitation, humidity, and sunshine. | Yes, for feeding, preening, inactivity, movement, and vigilance. | Yes, for all zones analysed. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kidd, P.; Ford, S.; Rose, P.E. Exploring the Effect of the COVID-19 Zoo Closure Period on Flamingo Behaviour and Enclosure Use at Two Institutions. Birds 2022, 3, 117-137. https://doi.org/10.3390/birds3010009
Kidd P, Ford S, Rose PE. Exploring the Effect of the COVID-19 Zoo Closure Period on Flamingo Behaviour and Enclosure Use at Two Institutions. Birds. 2022; 3(1):117-137. https://doi.org/10.3390/birds3010009
Chicago/Turabian StyleKidd, Peter, Steph Ford, and Paul E. Rose. 2022. "Exploring the Effect of the COVID-19 Zoo Closure Period on Flamingo Behaviour and Enclosure Use at Two Institutions" Birds 3, no. 1: 117-137. https://doi.org/10.3390/birds3010009
APA StyleKidd, P., Ford, S., & Rose, P. E. (2022). Exploring the Effect of the COVID-19 Zoo Closure Period on Flamingo Behaviour and Enclosure Use at Two Institutions. Birds, 3(1), 117-137. https://doi.org/10.3390/birds3010009