
Climatic Effects on Grey-Faced Petrel (Pterodroma gouldi) Chick Growth and Survival

 , ,
, ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Species
2.2. Study Site
2.3. Field Methods
2.4. Statistical Methods
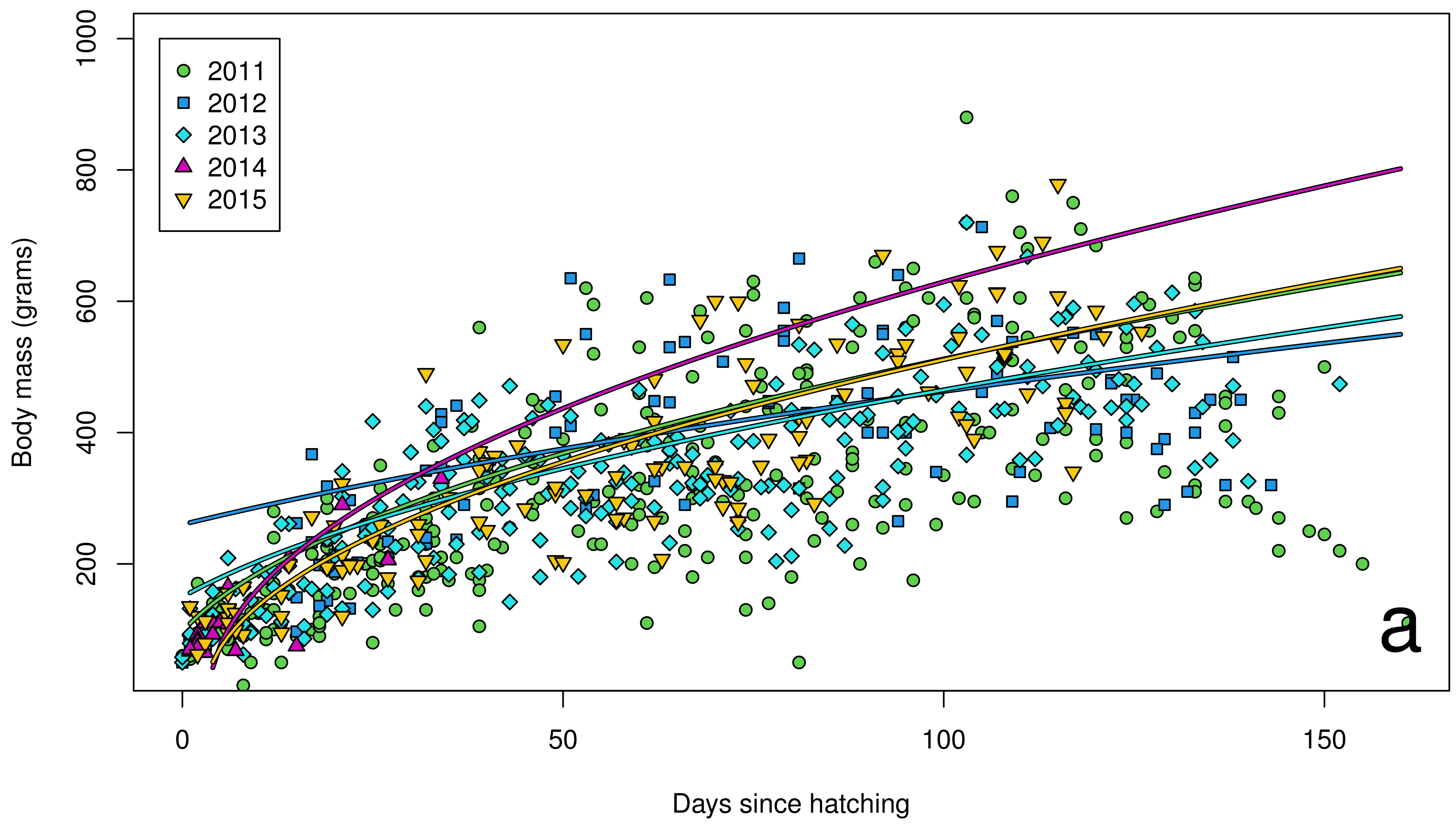
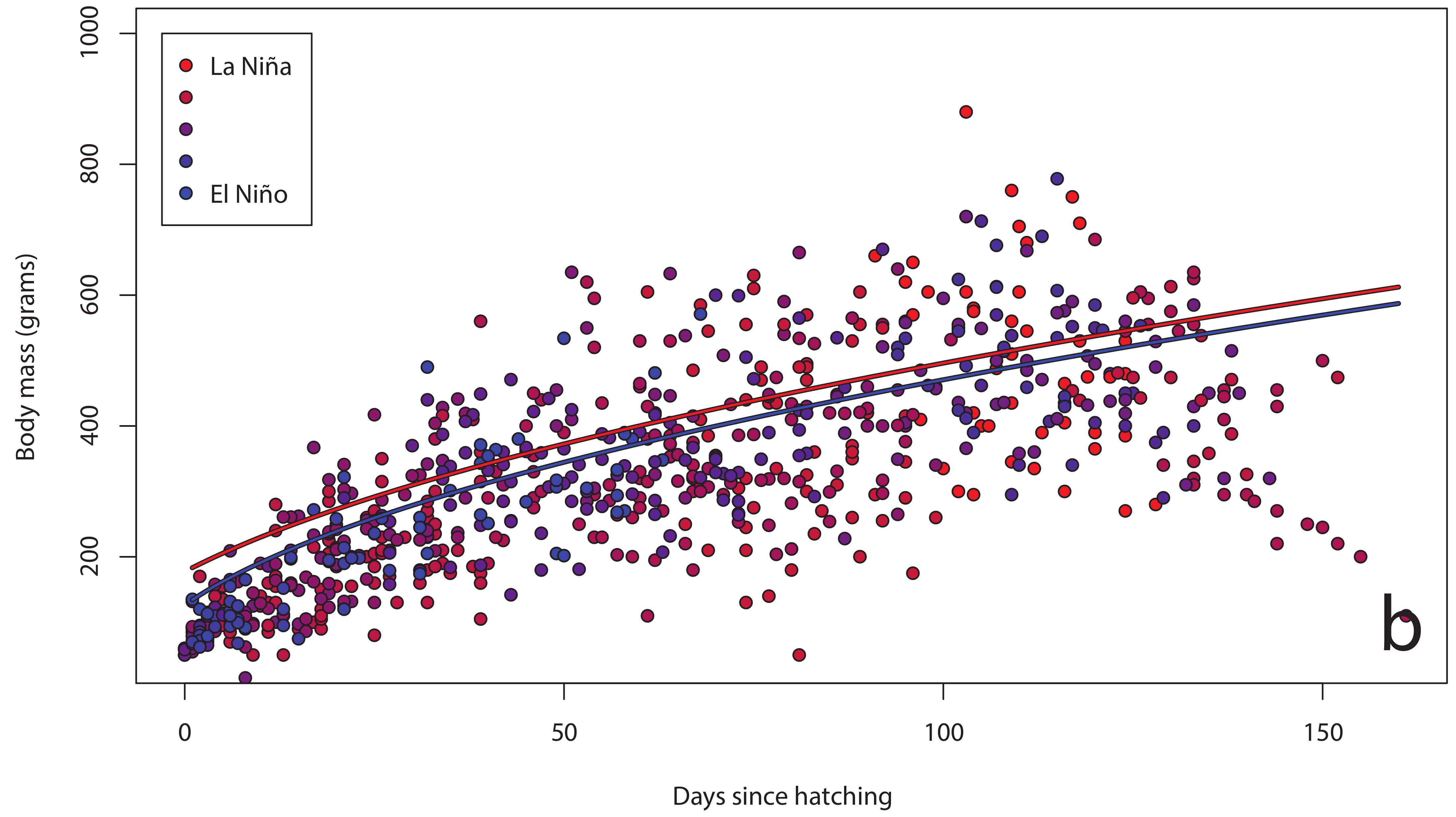
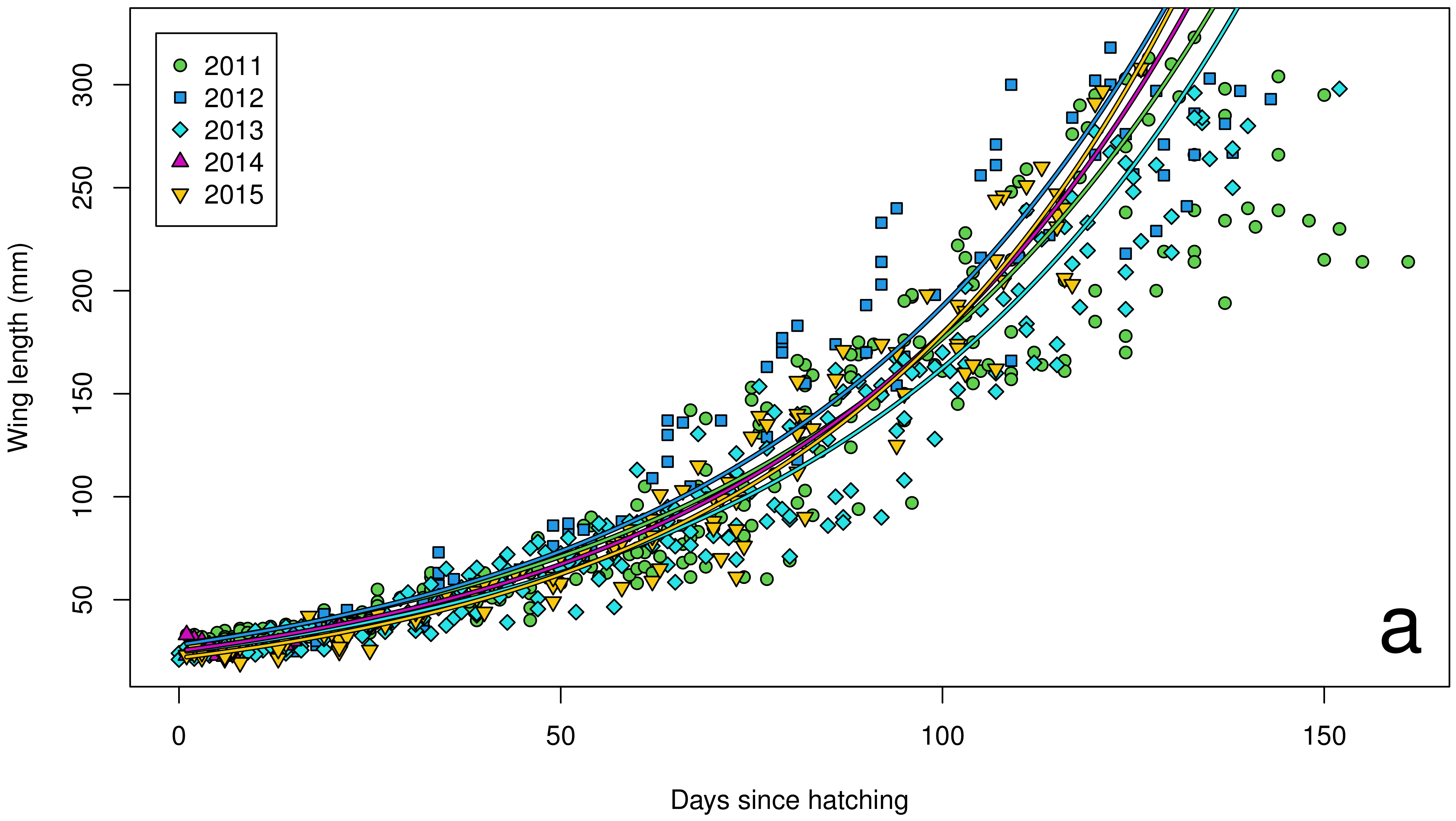
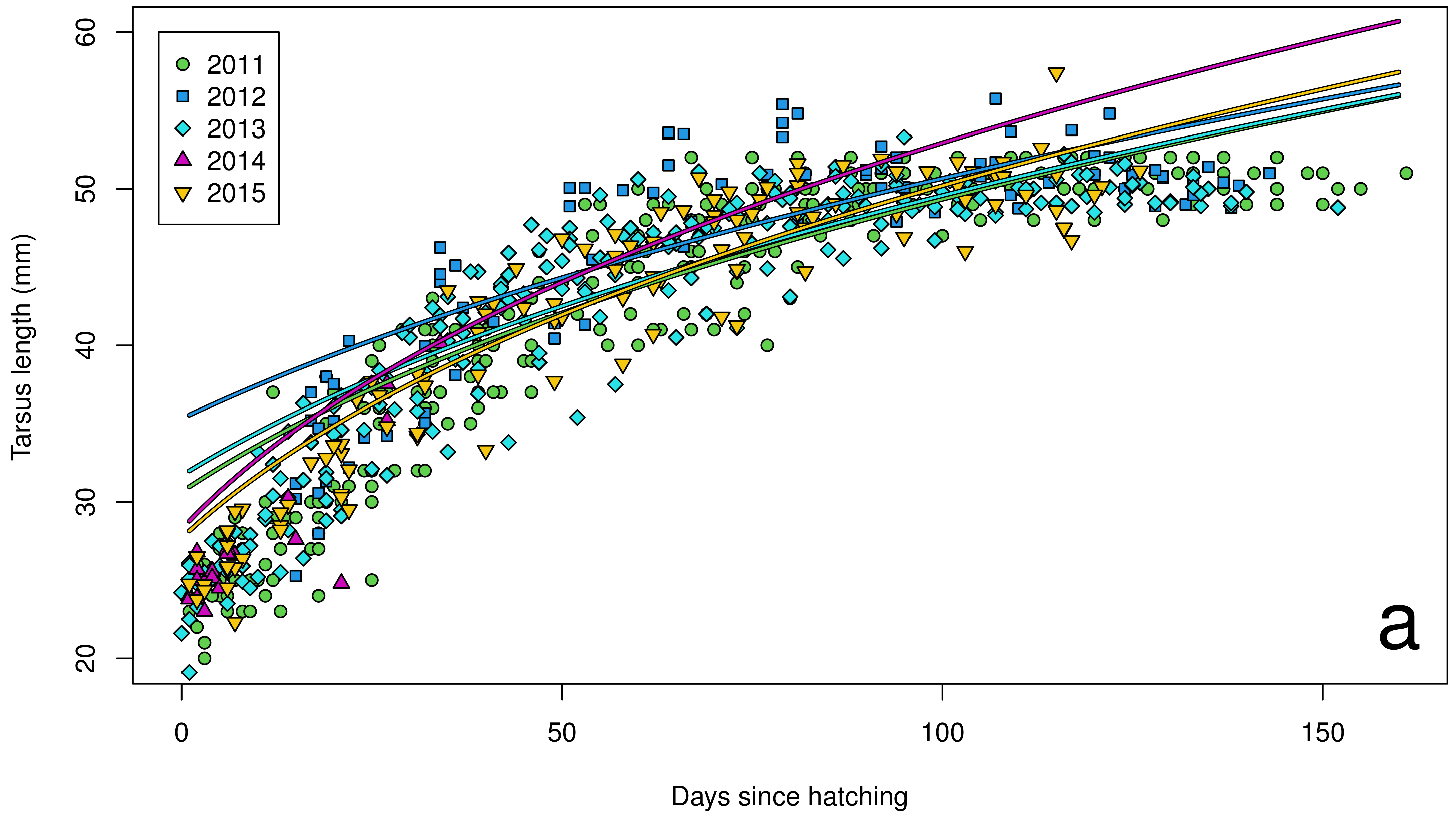
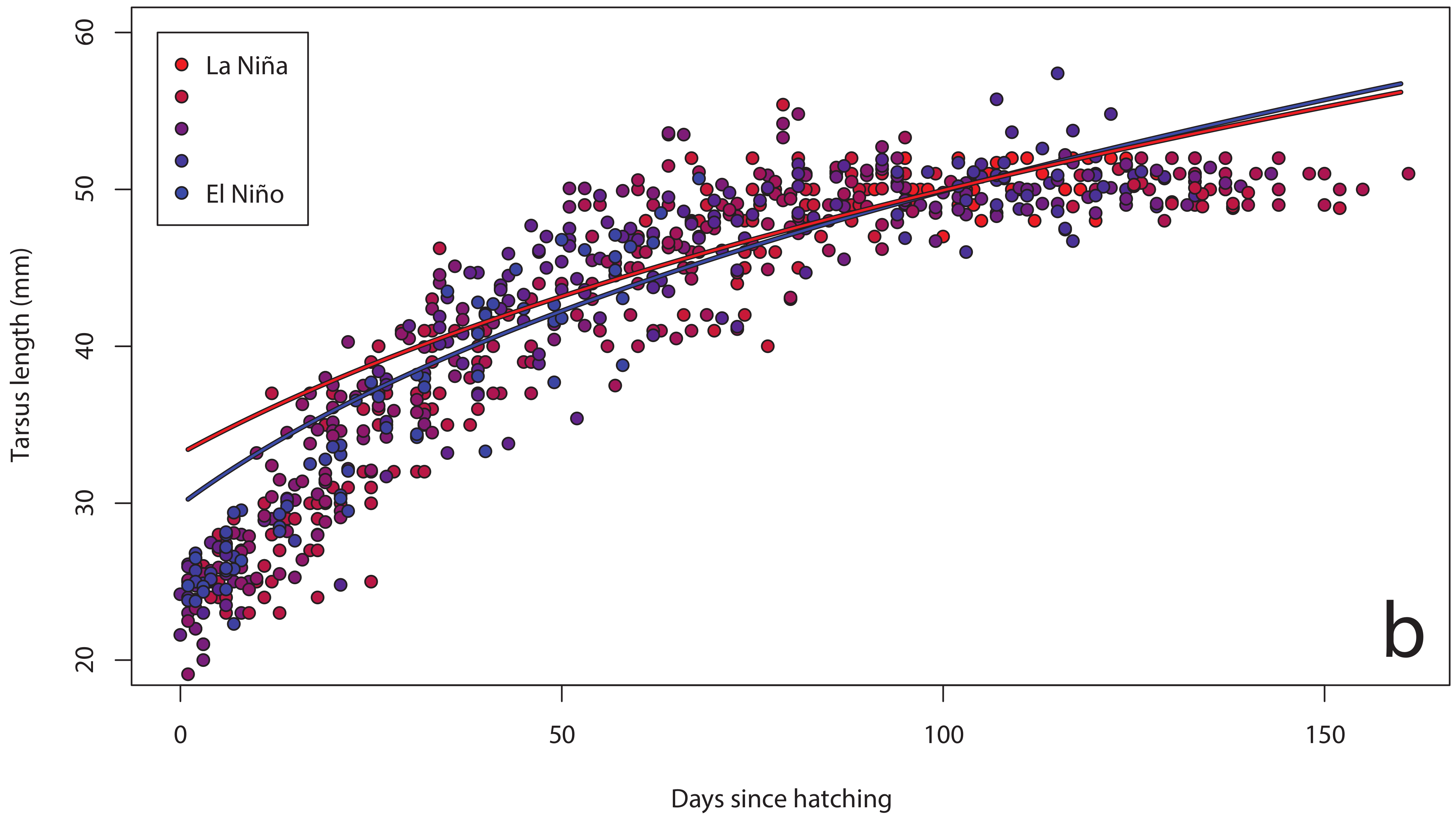
3. Results

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Year | Island | Visits | Burrows | Hatched | Date | Fledged | Date | SOI |
|---|---|---|---|---|---|---|---|---|
| 2011 | Motuora | fortnightly | 24 | 0.88 | NA | 0.54 | NA | +1.15 |
| 2011 | Tiritiri | weekly | 15 | 0.93 | 30 August | 0.47 | 13 January | +1.15 |
| 2011 | Te Hāwere | weekly | 22 | 0.86 | 26 August | 0.36 | 5 January | +1.15 |
| 2012 | Te Hāwere | fortnightly | 22 | 0.77 | 25 August | 0.41 | 31 December | −0.15 |
| 2013 | Te Hāwere | weekly | 22 | 0.68 | 28 August | 0.45 | 7 January | +0.29 |
| 2014 | Te Hāwere | weekly | 22 | 0.82 | 25 August | 0 1 | ˗ | −0.91 |
| 2015 | Te Hāwere | fortnightly | 22 | 0.73 | 30 August | 0.59 | 22 December | −1.58 |
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Grosbois, V.; Gimenez, O.; Gaillard, J.M.; Pradel, R.; Barbraud, C.; Clobert, J.; Weimerskirch, H. Assessing the impact of climate variation on survival in vertebrate populations. Biol. Rev. 2008, 83, 357–399. [Google Scholar] [CrossRef]
- Bronson, F.H. Climate change and seasonal reproduction in mammals. Phil. Trans. Roy. Soc. B 2009, 364, 3331–3340. [Google Scholar] [CrossRef] [Green Version]
- White, T.C.R. The role of food, weather and climate in limiting the abundance of animals. Biol. Rev. 2008, 83, 227–248. [Google Scholar] [CrossRef]
- Frederiksen, M.; Harris, M.P.; Daunt, F.; Rothery, P.; Wanless, S. Scale-dependent climate signals drive breeding phenology of three seabird species. Glob. Chang. Biol. 2004, 10, 1214–1221. [Google Scholar] [CrossRef]
- Bellard, C.; Bertelsmeier, C.; Leadley, P.; Thuiller, W.; Courchamp, F. Impacts of climate change on the future of biodiversity. Ecol. Lett. 2012, 15, 365–377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beal, M.; Dias, M.P.; Phillips, R.A.; Oppel, S.; Hazin, C.; Pearmain, E.J.; Catry, P. Global political responsibility for the conservation of albatrosses and large petrels. Sci. Adv. 2021, 27, eabd7225. [Google Scholar] [CrossRef]
- Watanuki, Y.; Yamamoto, M.; Okado, J.; Ito, M.; Sydeman, W. Seabird reproductive responses to changing climate and prey communities are mediated by prey packaging. Mar. Ecol. Prog. Ser. 2022, 683, 179–194. [Google Scholar] [CrossRef]
- Anderson, D.J. Differential responses of boobies and other seabirds in the Galápagos to the 1986–1987 El Nino-Southern Oscillation event. Mar. Ecol. Prog. Ser. 1989, 52, 209–216. [Google Scholar] [CrossRef]
- Schreiber, R.W.; Schreiber, E.A. Central Pacific seabirds and the El Niño southern oscillation: 1982 to 1983 perspectives. Science 1984, 225, 713–716. [Google Scholar] [CrossRef]
- Gjerdrum, C.; Vallée, A.M.; St Clair, C.C.; Bertram, D.F.; Ryder, J.L.; Blackburn, G.S. Tufted puffin reproduction reveals ocean climate variability. Proc. Natl. Acad. Sci. USA 2003, 100, 9377–9382. [Google Scholar] [CrossRef] [Green Version]
- Sauve, D.; Charmantier, A.; Hatch, S.A.; Friesen, V.L. Environmental conditions variably affect growth across the breeding season in a subarctic seabird. Oecologia 2021, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Greene, B.S.; Taylor, G.A.; Earl, R. Distribution, population status and trends of grey-faced petrel (Pterodroma macroptera gouldi) in the northern North Island, New Zealand. Notornis 2015, 62, 143–161. [Google Scholar]
- Miskelly, C.M.; Gilad, D.; Taylor, G.A.; Tennyson, A.J.; Waugh, S.M. A review of the distribution and size of gadfly petrel (Pterodroma spp.) colonies throughout New Zealand. Tuhinga 2019, 30, 93–173. [Google Scholar]
- Russell, J.C.; Welch, J.R.; Dromzée, S.; Bourgeois, K.; Thoresen, J.; Earl, R.; McNutt, K. Developing a National Framework for Monitoring the Grey-Faced Petrel (Pterodroma gouldi) as an Indicator Species; DOC Research and Development Series; Department of Conservation: Wellington, New Zealand, 2017; Volume 350, pp. 1–19.
- Stolpmann, L.M.; Landers, T.J.; Russell, J.C. Camera trapping of Grey-faced Petrel (Pterodroma gouldi) breeding burrows reveals interactions with introduced mammals throughout the breeding season. Emu Austral Ornith. 2019, 119, 391–396. [Google Scholar] [CrossRef]
- Ross, E.L.; Brunton, D. Seasonal trends and nightly variation in colony attendance of grey-faced petrels (Pterodroma macroptera gouldi). Notornis 2002, 49, 153–157. [Google Scholar]
- Mullan, A.B. On the linearity and stability of Southern Oscillation-climate relationships for New Zealand. Int. J. Climat. 1995, 15, 1365–1386. [Google Scholar] [CrossRef]
- Bourgeois, K.; Welch, J.R.; Dromzée, S.; Taylor, G.A.; Russell, J.C. Flexible foraging strategies in a highly pelagic seabird revealed by seasonal isotopic niche variation. Mar. Biol. 2022, 169, 28. [Google Scholar] [CrossRef]
- Wood, J.R.; Lawrence, H.A.; Scofield, R.P.; Taylor, G.A.; Lyver, P.O.; Gleeson, D.M. Morphological, behavioural, and genetic evidence supports reinstatement of full species status for the grey-faced petrel, Pterodroma macroptera gouldi (Procellariiformes: Procellariidae). Zool. J. Linn. Soc. 2017, 179, 201–216. [Google Scholar] [CrossRef] [Green Version]
- Jones, C.J.; Clifford, H.; Fletcher, D.; Cuming, P.; Lyver, P.O.B. Survival and age-at-first-return estimates for grey-faced petrels (Pterodroma macroptera gouldi) breeding on Mauao and Motuotau Island in the Bay of Plenty, New Zealand. Notornis 2011, 58, 71–80. [Google Scholar]
- Imber, M.J. Breeding biology of the grey-faced petrel Pterodroma macroptera gouldi. Ibis 1976, 118, 51–64. [Google Scholar] [CrossRef]
- Russell, J.C.; Mackay, J.W.; Abdelkrim, J. Insular pest control within a metapopulation context. Biol. Cons. 2009, 142, 1404–1410. [Google Scholar] [CrossRef]
- Pichlmueller, F.; Russell, J.C. Survivors or reinvaders? Intraspecific priority effect masks reinvasion potential. Biol. Cons. 2018, 227, 213–218. [Google Scholar] [CrossRef]
- Clout, M.N.; Russell, J.C. The eradication of mammals from New Zealand islands. In Assessment and Control of Biological Invasion Risks; Koike, F., Clout, M.N., Kawamichi, M., De Poorter, M., Iwatsuki, K., Eds.; IUCN: Gland, Switzerland, 2006; pp. 127–141. [Google Scholar]
- Bourgeois, K.; Dromzée, S.; Welch, J.R.; Russell, J.C. Sex and geographic variation in grey-faced petrel (Pterodroma gouldi) morphometrics. Waterbirds 2017, 40, 144–153. [Google Scholar] [CrossRef]
- Hart, D.R.; Chute, A.S. Estimating von bertalanffy growth parameters from growth increment data using a linear mixed-effects model, with an application to the sea scallop Placopecten magellanicus. ICES J. Mar. Sci. 2009, 66, 2165–2175. [Google Scholar] [CrossRef]
- Ropelewski, C.F.; Jones, P.D. An extension of the Tahiti–Darwin southern oscillation index. Mon. Weather Rev. 1987, 115, 2161–2165. [Google Scholar] [CrossRef] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021; Available online: https://www.R-project.org/ (accessed on 7 February 2022).
- Pinheiro, J.; Bates, D.; DebRoy, S.; Sarkar, D.; R Core Team. nlme: Linear and Nonlinear Mixed Effects Models. R Package Version 3.1-152. 2021. Available online: https://CRAN.R-project.org/package=nlme (accessed on 7 February 2022).
- Cruz, J.B.; Cruz, F. Effect of El Niño-Southern Oscillation conditions on nestling growth rate in the Dark-rumped Petrel. Condor 1990, 92, 160–165. [Google Scholar] [CrossRef]
- Rodríguez, A.; Arcos, J.M.; Bretagnolle, V.; Dias, M.P.; Holmes, N.D.; Louzao, M.; Chiaradia, A. Future directions in conservation research on petrels and shearwaters. Front. Mar. Sci. 2019, 6, 94. [Google Scholar] [CrossRef]
- Miskelly, C.M.; Tennyson, A.J.; Stahl, J.C.; Smart, A.F.; Edmonds, H.K.; McMurtrie, P.G. Breeding petrels of Dusky Sound, Fiordland–survivors from a century of stoat invasions. Notornis 2017, 64, 136–153. [Google Scholar]
- McKechnie, S.; Fletcher, D.; Newman, J.; Bragg, C.; Dillingham, P.W.; Clucas, R.; Moller, H. Separating the effects of climate, bycatch, predation and harvesting on tītī (Ardenna grisea) population dynamics in New Zealand: A model-based assessment. PLoS ONE 2020, 15, e0243794. [Google Scholar] [CrossRef]
- Fletcher, D.; Newman, J.; McKechnie, S.; Bragg, C.; Dillingham, P.; Clucas, R.; Moller, H. Projected impacts of climate change, bycatch, harvesting, and predation on the Aotearoa New Zealand tītī Ardenna grisea population. Mar. Ecol. Prog. Ser. 2021, 670, 223–238. [Google Scholar] [CrossRef]
- Gronwald, M.; Russell, J.C. Behaviour of invasive ship rats, Rattus rattus, around Goodnature A24 self-resetting traps. Manag. Biol. Inv. 2022, 13, 1–15. [Google Scholar] [CrossRef]
- Russell, J.C.; Lecomte, V.; Dumont, Y.; Le Corre, M. Intraguild predation and mesopredator release effect on long-lived prey. Ecol. Mod. 2009, 220, 1098–1104. [Google Scholar] [CrossRef]
- Brothers, N.; Bone, C. The response of burrow-nesting petrels and other vulnerable bird species to vertebrate pest management and climate change on sub-Antarctic Macquarie Island. Pap. Proc. Roy. Soc. Tasmania 2008, 142, 123–148. [Google Scholar] [CrossRef] [Green Version]
- Shears, N.T.; Bowen, M.M. Half a century of coastal temperature records reveal complex warming trends in western boundary currents. Sci. Rep. 2017, 7, 14527. [Google Scholar] [CrossRef]
- Votier, S.C.; Birkhead, T.R.; Oro, D.; Trinder, M.; Grantham, M.J.; Clark, J.A.; Hatchwell, B.J. Recruitment and survival of immature seabirds in relation to oil spills and climate variability. J. Anim. Ecol. 2008, 77, 974–983. [Google Scholar] [CrossRef]
- Carey, M.J. The effects of investigator disturbance on procellariiform seabirds: A review. N. Z. J. Zool. 2009, 36, 367–377. [Google Scholar] [CrossRef] [Green Version]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Russell, J.C.; Welch, J.R.; Bourgeois, K.; Dromzée, S.; Dunn, R.; Friesen, M.R.; Rayner, M.J. Climatic Effects on Grey-Faced Petrel (Pterodroma gouldi) Chick Growth and Survival. Birds 2022, 3, 138-148. https://doi.org/10.3390/birds3010010
Russell JC, Welch JR, Bourgeois K, Dromzée S, Dunn R, Friesen MR, Rayner MJ. Climatic Effects on Grey-Faced Petrel (Pterodroma gouldi) Chick Growth and Survival. Birds. 2022; 3(1):138-148. https://doi.org/10.3390/birds3010010
Chicago/Turabian StyleRussell, James C., Jemma R. Welch, Karen Bourgeois, Sylvain Dromzée, Rob Dunn, Megan R. Friesen, and Matt J. Rayner. 2022. "Climatic Effects on Grey-Faced Petrel (Pterodroma gouldi) Chick Growth and Survival" Birds 3, no. 1: 138-148. https://doi.org/10.3390/birds3010010
APA StyleRussell, J. C., Welch, J. R., Bourgeois, K., Dromzée, S., Dunn, R., Friesen, M. R., & Rayner, M. J. (2022). Climatic Effects on Grey-Faced Petrel (Pterodroma gouldi) Chick Growth and Survival. Birds, 3(1), 138-148. https://doi.org/10.3390/birds3010010