
Dynamics of Beddington–DeAngelis Type Eco-Epidemiological Model with Prey Refuge and Prey Harvesting †

 , and
, and
Abstract
:1. Introduction
2. Model Formation
3. Mathematical Results
3.1. Positive Invariance
3.2. Positivity of Solutions
3.3. Boundedness of Solution
4. Equilibrium Points
- represents the essence of trivial equilibrium.
- is the free of infection and predator-free equilibrium that exists for .
- is the predator-free equilibrium, where , .
- The positive equilibrium is , where, , and exist unique positive roots of the below polynomial equations, , where, , , .
5. Local Stability Analysis
6. Global Stability Analysis
7. Hopf Bifurcation Analysis
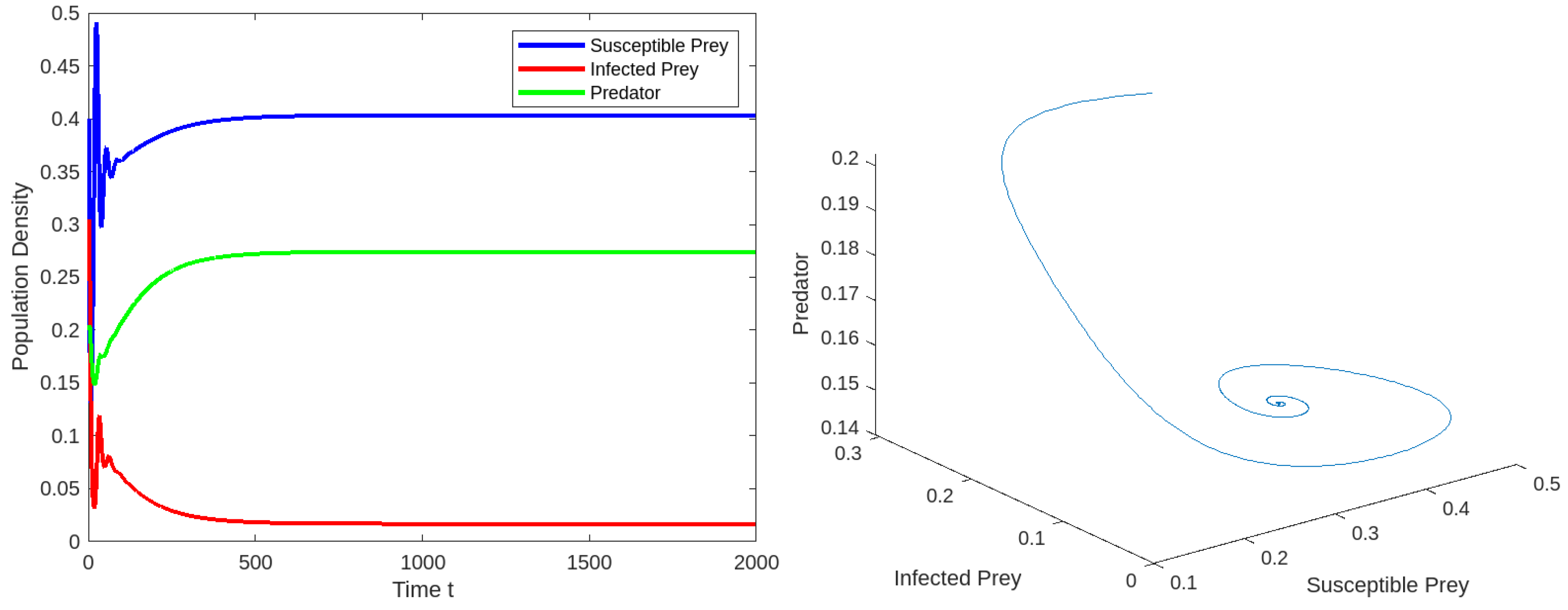
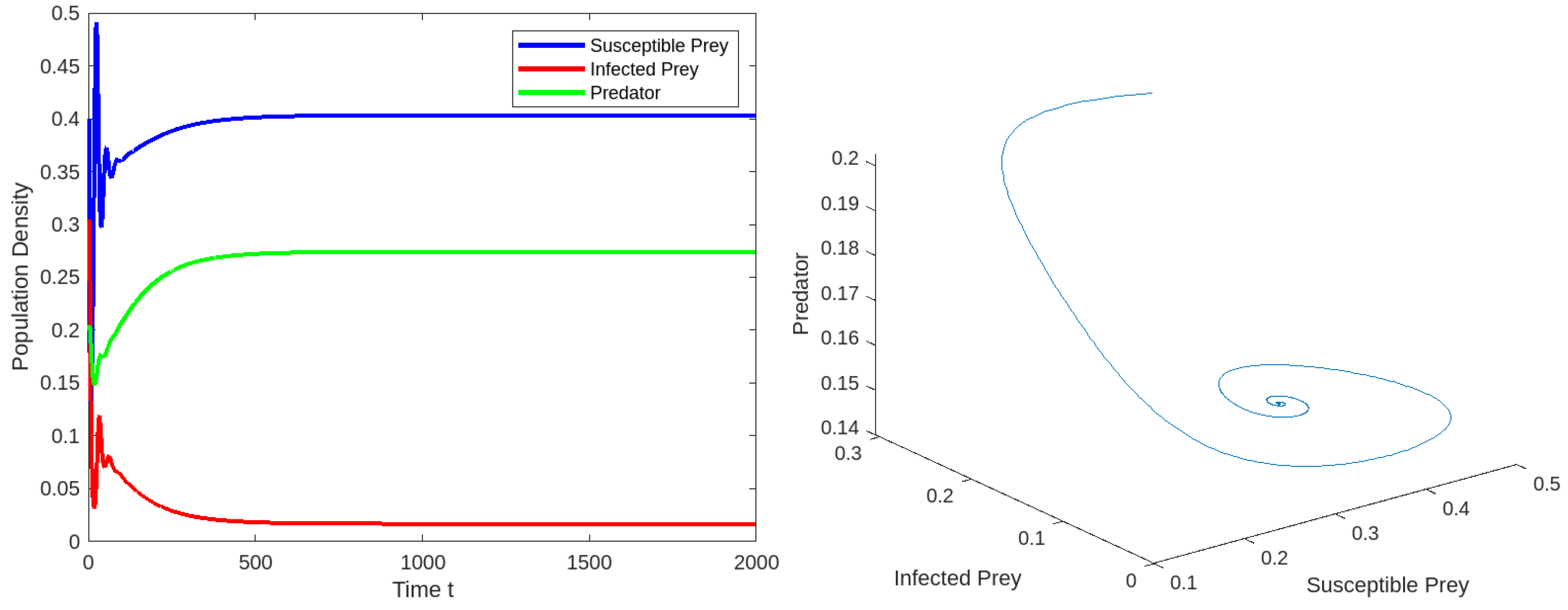
8. Numerical Simulations
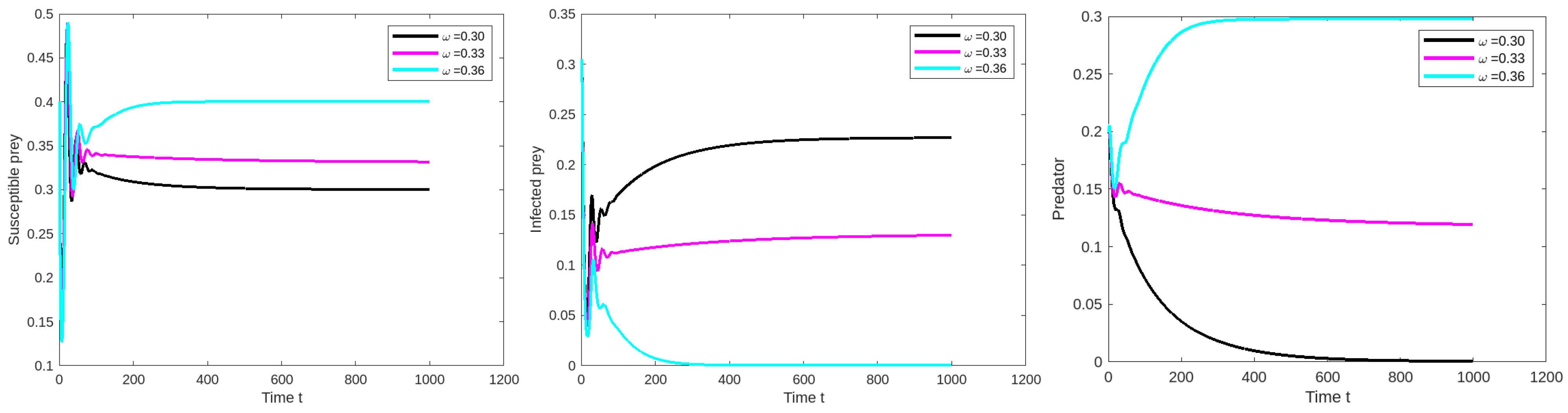
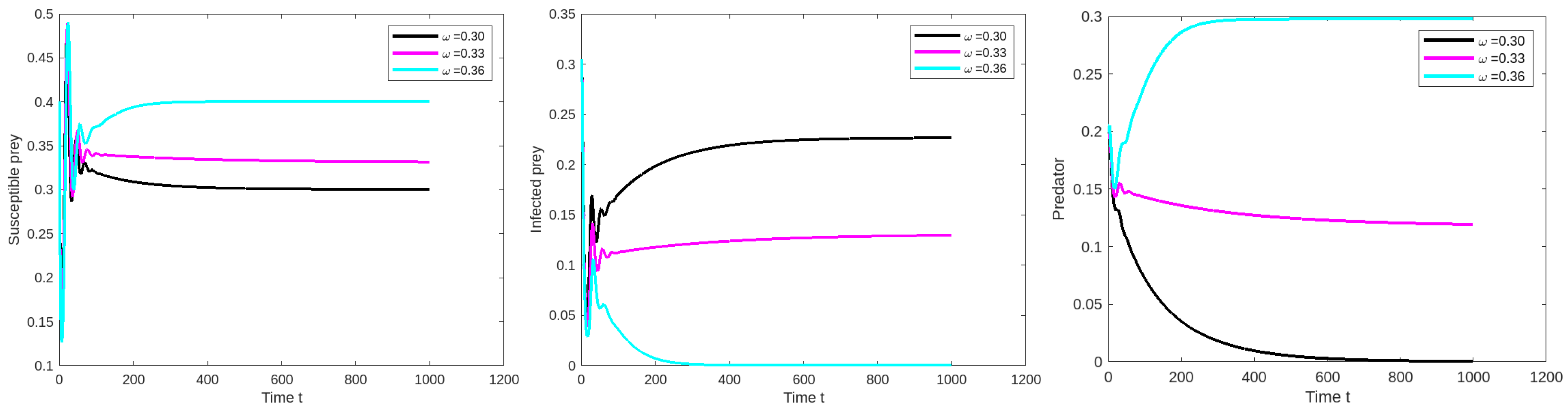
8.1. Effect of Varying the Susceptible Prey Predator Rate
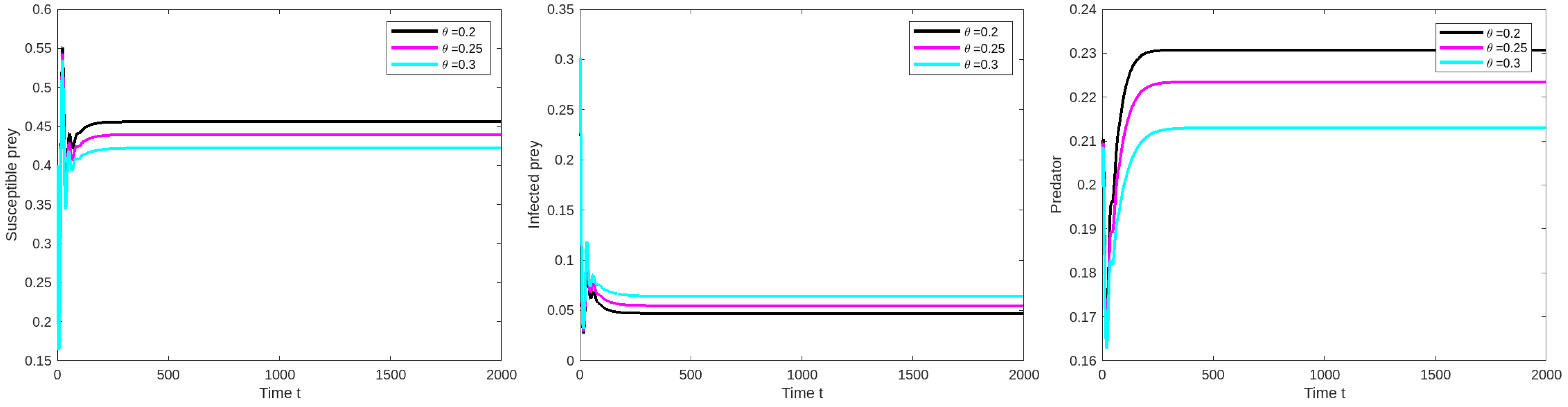
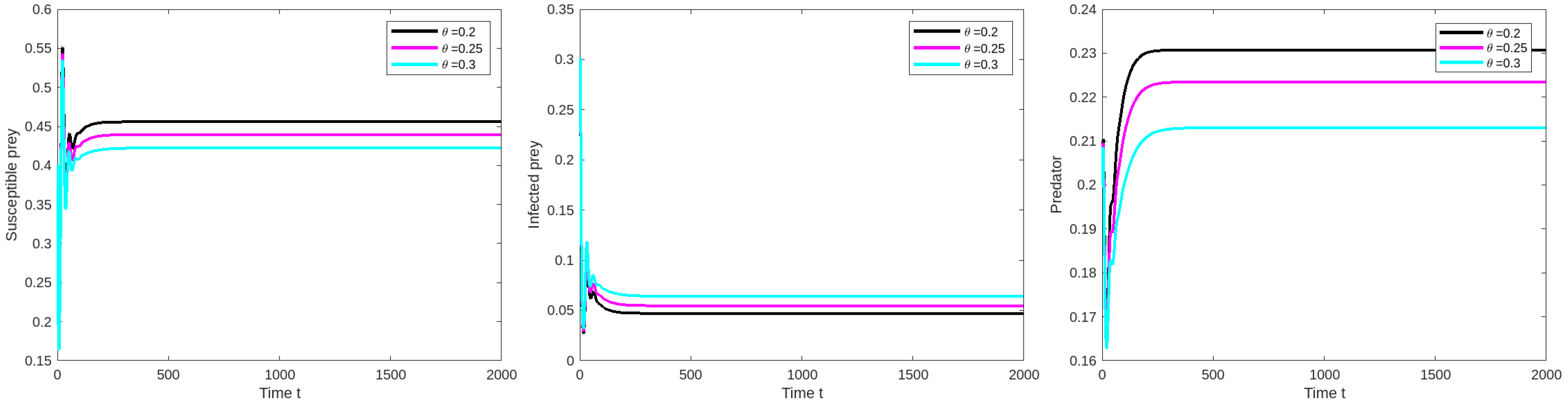
8.2. Effect of Varying the Prey Refuge
9. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Dimitrov, D.T.; Kojouharov, H.V. Complete mathematical analysis of predator–prey models with linear prey growth and Beddington–DeAngelis functional response. Appl. Math. Comput. 2005, 162, 523–538. [Google Scholar] [CrossRef]
- Tripathi, J.P.; Abbas, S.; Thakur, M. A density dependent delayed predator–prey model with Beddington–DeAngelis type function response incorporating a prey refuge. Commun. Nonlinear Sci. Numer. Simul. 2015, 22, 427–450. [Google Scholar] [CrossRef]
- Divya, A.; Sivabalan, M.; Ashwin, A.; Siva Pradeep, M. Dynamics of ratio dependent eco epidemiological model with prey refuge and prey harvesting. In Proceedings of the 1st International Online Conference on Mathematics and Applications, Online, 1–15 May 2023. [Google Scholar]
- Cantrell, R.S.; Cosner, C. On the dynamics of predator–prey models with the Beddington–DeAngelis functional response. J. Math. Anal. Appl. 2001, 257, 206–222. [Google Scholar] [CrossRef]
- Fan, M.; Kuang, Y. Dynamics of a nonautonomous predator–prey system with the Beddington–DeAngelis functional response. J. Math. Anal. Appl. 2004, 295, 15–39. [Google Scholar] [CrossRef]
- Ashwin, A.; Sivabalan, M.; Divya, A.; Siva Pradeep, M. Dynamics of Holling type II eco-epidemiological model with fear effect, prey refuge, and prey harvesting. In Proceedings of the 1st International Online Conference on Mathematics and Applications, Online, 1–15 May 2023. [Google Scholar]
- Melese, D.; Muhye, O.; Sahu, S.K. Dynamical behavior of an eco-epidemiological model incorporating prey refuge and prey harvesting. Appl. Appl. Math. Int. J. (AAM) 2020, 15, 28. [Google Scholar]
- Tripathi, J.P.; Jana, D.; Tiwari, V. A Beddington–DeAngelis type one-predator two-prey competitive system with help. Nonlinear Dyn. 2018, 94, 553–573. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
| Parameters | Physiological Representation | Units |
|---|---|---|
| Magnitude of interference of predator | m | |
| Effect of handing time for predator | m | |
| The harvesting effort of predator | No. per unit area (tons) | |
| Prey growth rate | per day (t−1) | |
| L | Environment carrying capacity | No. per unit area (tons) |
| and | Catchability coefficient of predator | per day (t−1) |
| Constant of half-saturation | m | |
| Susceptible prey rate of predation | per day (t−1) | |
| c | Conversion rate of prey to predator | |
| and | Death rate of infected prey and predator | per day (t−1) |
| Infected prey predation rate | per day (t−1) | |
| The incidence of contamination for prey | per day (t−1) | |
| Refuge of prey | m−1 | |
| Predator, susceptible and infected prey | No. per unit area (tons) |
| Parameters | Numeric Value |
|---|---|
| r | 0.5 |
| 0.2 | |
| d | 0.1 |
| c | 0.5 |
| 0.1 | |
| 0.2 | |
| 0.3 | |
| 0.2 | |
| 0.12 | |
| 0.01 | |
| 0.1 | |
| variable | |
| variable |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ashwin, A.R.; Sivabalan, M.; Divya, A.; Siva Pradeep, M. Dynamics of Beddington–DeAngelis Type Eco-Epidemiological Model with Prey Refuge and Prey Harvesting. Eng. Proc. 2023, 56, 306. https://doi.org/10.3390/ASEC2023-15691
Ashwin AR, Sivabalan M, Divya A, Siva Pradeep M. Dynamics of Beddington–DeAngelis Type Eco-Epidemiological Model with Prey Refuge and Prey Harvesting. Engineering Proceedings. 2023; 56(1):306. https://doi.org/10.3390/ASEC2023-15691
Chicago/Turabian StyleAshwin, Anbulinga Raja, Muthuradhinam Sivabalan, Arumugam Divya, and Manickasundaram Siva Pradeep. 2023. "Dynamics of Beddington–DeAngelis Type Eco-Epidemiological Model with Prey Refuge and Prey Harvesting" Engineering Proceedings 56, no. 1: 306. https://doi.org/10.3390/ASEC2023-15691
APA StyleAshwin, A. R., Sivabalan, M., Divya, A., & Siva Pradeep, M. (2023). Dynamics of Beddington–DeAngelis Type Eco-Epidemiological Model with Prey Refuge and Prey Harvesting. Engineering Proceedings, 56(1), 306. https://doi.org/10.3390/ASEC2023-15691