
Are Dietary Sugars Potent Adipose Tissue and Immune Cell Modulators?


Abstract
1. Introduction
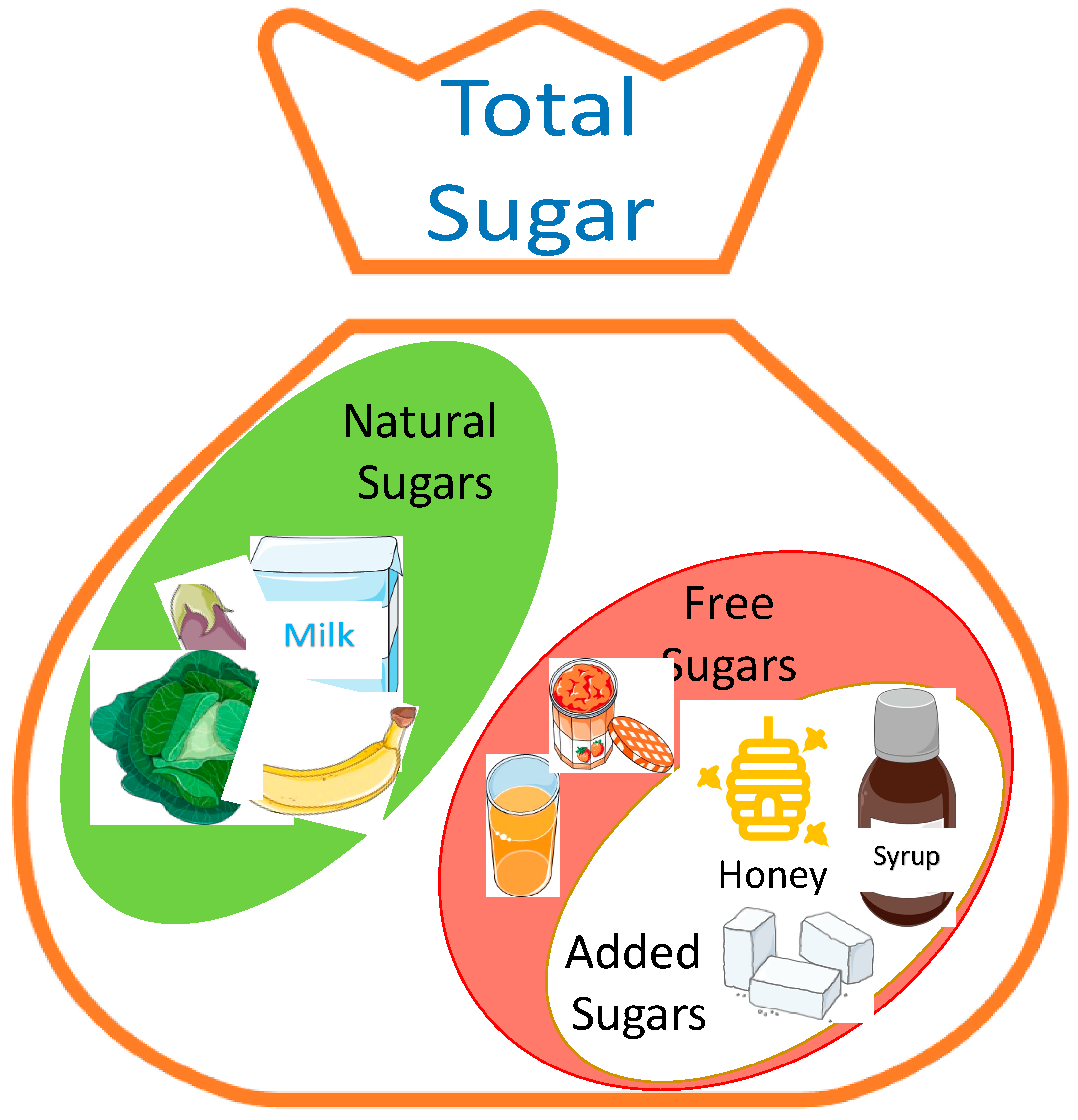
2. Dietary (Free) Sugars
3. Sugar Metabolism and Its Effect on Adipose Tissue
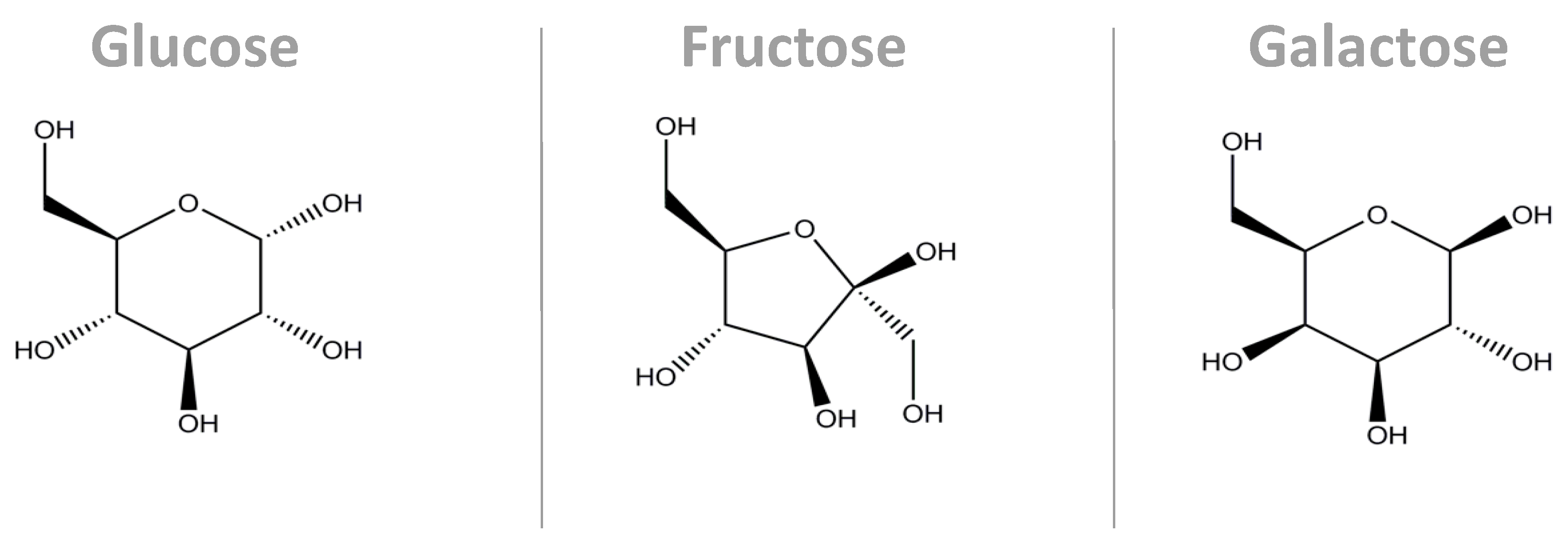
3.1. Sucrose and HFCS—Fructose and Glucose Metabolism
3.2. Lactose—Galactose and Glucose
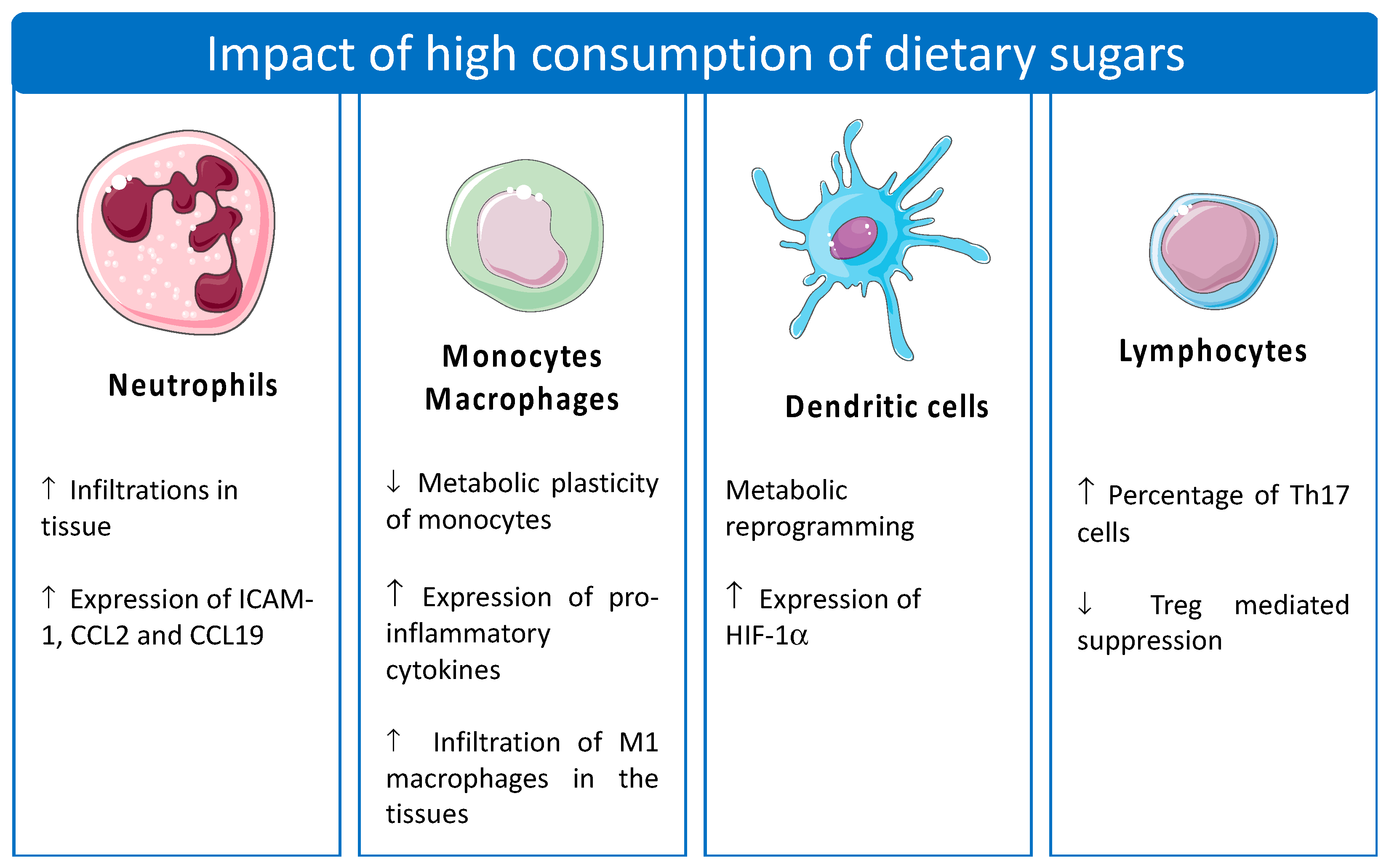
4. The Impact of Sugar on Immunometabolism

{kind=link}
{kind=link}
{kind=link}
| Dietary Sugar | Experimental Model | Concentrations of Sugar | Treatment Duration | Impact in Immune Populations | Reference |
|---|---|---|---|---|---|
| Fructose | in vivo—IDH2 KO and C57BL/6J mice | 34% fructose in H2O | 6 weeks | ↑ Neutrophil infiltration into the liver of IDH2 mice | [52] |
| in vivo—male C57BL/6J mice | 15% fructose in H2O | 10 days | ↑ Neutrophil trafficking to limbal region | [53] | |
| in vivo—C57BL/6J mice | 30% fructose in H2O | 8 weeks | ↑ Neutrophil infiltration into the liver ↑ Expression of ICAM-1, CCL2 and CCL19 | [44] | |
| in vivo—male Swiss mice | 20% fructose in H2O | 6–10 weeks | ↑ Infiltration of M1 macrophages into AT ↓ Infiltration of M2 macrophages into AT | [54] | |
| in vitro—primary human DCs | 5 to 25 mM fructose in culture medium | 24–72 h | ↑ Pro-inflammatory cytokine production and activation markers 🞄 Metabolic reprogramming in DCs | [34] | |
| in vivo—male Sprague Dawley rats | 60% fructose in chow | 5 weeks | ↓ Immune suppressive function of Treg cells without changes in the percentage of the population | [55] | |
| in vitro—primary human monocytes | 11.1 mM fructose in culture medium | Different time-points | ↓ Metabolic plasticity of monocytes ↑ Expression of IL-1β, IL-6, IL-10, and TNF-α | [47] | |
| in vivo—Dahl salt-sensitive and Dahl salt-resistant rats | 20% fructose in H2O | 4 weeks | ↑ % of Th17 cells | [56] | |
| Fructose and sucrose | in vivo—male BALB/c and LysM-eGFP mice | 20% fructose20% sucrose (control),both in the chow | 12–14 weeks | ↑ Leukocyte count in circulation and infiltrated neutrophils in the liver ↓ Neutrophil infiltration into AT | [26] |
| Galactose | in vitro—human monocytes | 11.1 mM galactose in culture medium | Different time-points | ↓ Metabolic plasticity of monocytes | [47] |
| in vitro—THP-1 cells | 2 g/L of RPMI | 5 days until experiments | ↓ TNF-α expression and preference for OXPHOS | [57] | |
| in vitro—bone marrow DCs from male C57BL/6J mice | 10 mM of galactose in culture medium | 24–72 h | 🞄 Induces the maintenance of activating markers on DCs for 72h | [58] | |
| Glucose | in vitro—human DCs | Range from 5 to 25 mM glucose in culture medium | 24–72 h | ↑ Expression of HIF-1α | [34] |
| in vitro—human and male C57BL/6J mice CD8+ T cells | 25 mM glucose | 3 days | ↑ Glycolysis and cytotoxic capacity of CTLs | [59] | |
| Lactose | in vitro—human T cells | 30 mM of lactose30 mM of sucrose in culture medium | 3 days | ↓ Treg immunosuppression capacity | [46] |
| in vivo—female BALB/c mice | 100 mg/kg of body weight | Different time-points | 🞄 Neutrophil and macrophage modulation in early acute pancreatitis 🞄 Possible interaction with lactose-Galectin 3 | [45] | |
| Sucrose | in vivo—male Wistar rats | 5% (w/v) in H2O | 12 weeks | ↑ Circulating neutrophils, lymphocytes, and basophiles | [11] |
| in vivo—male Sprague Dawley | 700 g/kg of chow | 5 weeks | ↑ CD68+ macrophage infiltration into tPVAT and aPVAT ↑ Expression of MCP-1 in both tissues | [57] |
4.1. Monocytes/Macrophages
4.2. Dendritic Cells
4.3. Lymphocytes
5. Concluding Remarks
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kopp, W. How Western Diet and Lifestyle Drive the Pandemic of Obesity and Civilization Diseases. Diabetes Metab. Syndr. Obes. 2019, 12, 2221–2236. [Google Scholar] [CrossRef] [PubMed]
- Amano, T.; Watanabe, D.; Otsuka, J.; Okamoto, Y.; Takada, S.; Fujii, N.; Kenny, G.P.; Enoki, Y.; Maejima, D. Comparison of hydration efficacy of carbohydrate-electrolytes beverages consisting of isomaltulose and sucrose in healthy young adults: A randomized crossover trial. Physiol. Behav. 2022, 249, 113770. [Google Scholar] [CrossRef] [PubMed]
- Arias-Chávez, D.J.; Mailloux-Salinas, P.; Altamirano, J.; Huang, F.; Gómez-Viquez, N.L.; Bravo, G. Consumption of combined fructose and sucrose diet exacerbates oxidative stress, hypertrophy and CaMKIIδ oxidation in hearts from rats with metabolic syndrome. Mol. Cell. Biochem. 2022, 477, 1309–1320. [Google Scholar] [CrossRef] [PubMed]
- Butterworth, P.J.; Bajka, B.H.; Edwards, C.H.; Warren, F.J.; Ellis, P.R. Enzyme kinetic approach for mechanistic insight and predictions of in vivo starch digestibility and the glycaemic index of foods. Trends Food Sci. Technol. 2022, 120, 254–264. [Google Scholar] [CrossRef]
- Omar, N.A.M.; Frank, J.; Kruger, J.; Bello, F.D.; Medana, C.; Collino, M.; Zamaratskaia, G.; Michaelsson, K.; Wolk, A.; Landberg, R. Effects of High Intakes of Fructose and Galactose, with or without Added Fructooligosaccharides, on Metabolic Factors, Inflammation, and Gut Integrity in a Rat Model. Mol. Nutr. Food Res. 2021, 65, e2001133. [Google Scholar] [CrossRef] [PubMed]
- Drake, I.; Sonestedt, E.; Ericson, U.; Wallström, P.; Orho-Melander, M. A Western dietary pattern is prospectively associated with cardio-metabolic traits and incidence of the metabolic syndrome. Br. J. Nutr. 2018, 119, 1168–1176. [Google Scholar] [CrossRef]
- Vasbinder, A.; Anderson, E.; Shadid, H.; Berlin, H.; Pan, M.; Azam, T.U.; Khaleel, I.; Padalia, K.; Meloche, C.; O’Hayer, P.; et al. Inflammation, Hyperglycemia, and Adverse Outcomes in Individuals With Diabetes Mellitus Hospitalized for COVID-19. Diabetes Care 2022, 45, 692–700. [Google Scholar] [CrossRef]
- Pepin, A.; Stanhope, K.L.; Imbeault, P. Are Fruit Juices Healthier Than Sugar-Sweetened Beverages? A Review. Nutrients 2019, 11, 1006. [Google Scholar] [CrossRef]
- Febbraio, M.A.; Karin, M. “Sweet death”: Fructose as a metabolic toxin that targets the gut-liver axis. Cell Metab. 2021, 33, 2316–2328. [Google Scholar] [CrossRef]
- McCain, H.; Kaliappan, S.; Drake, M. Invited review: Sugar reduction in dairy products. J. Dairy Sci. 2018, 101, 8619–8640. [Google Scholar] [CrossRef]
- Patkar, O.L.; Mohamed, A.Z.; Narayanan, A.; Mardon, K.; Cowin, G.; Bhalla, R.; Stimson, D.H.R.; Kassiou, M.; Beecher, K.; Belmer, A.; et al. A binge high sucrose diet provokes systemic and cerebral inflammation in rats without inducing obesity. Sci. Rep. 2021, 11, 11252. [Google Scholar] [CrossRef] [PubMed]
- Malik, V.S.; Popkin, B.M.; Bray, G.A.; Després, J.P.; Hu, F.B. Sugar-sweetened beverages, obesity, type 2 diabetes mellitus, and cardiovascular disease risk. Circulation 2010, 121, 1356–1364. [Google Scholar] [CrossRef] [PubMed]
- Shendurse, A.M.; Khedkar, C.D. Lactose. Encycl. Food Health 2016, 509–516. [Google Scholar] [CrossRef]
- Meyers, A.M.; Mourra, D.; Beeler, J.A. High fructose corn syrup induces metabolic dysregulation and altered dopamine signaling in the absence of obesity. PLoS ONE 2017, 12, e0190206. [Google Scholar] [CrossRef]
- Taskinen, M.-R.; Packard, C.J.; Borén, J. Dietary Fructose and the Metabolic Syndrome. Nutrients 2019, 11, 1987. [Google Scholar] [CrossRef]
- Krycer, J.R.; Quek, L.-E.; Francis, D.; Zadoorian, A.; Weiss, F.C.; Cooke, K.C.; Nelson, M.E.; Diaz-Vegas, A.; Humphrey, S.J.; Scalzo, R.; et al. Insulin signaling requires glucose to promote lipid anabolism in adipocytes. J. Biol. Chem. 2020, 295, 13250–13266. [Google Scholar] [CrossRef]
- Blanco, A.; Blanco, G. Carbohydrates. In Medical Biochemistry; Elsevier: Amsterdam, Netherlands, 2022; pp. 77–103. [Google Scholar] [CrossRef]
- Olofsson, C.; Eriksson, M.; Helin, A.-C.B.; Anderstam, B.; Orsini, N.; Stenvinkel, P.; Ekberg, N.R. Effects of Acute Fructose Loading on Markers of Inflammation—A Pilot Study. Nutrients 2021, 13, 3110. [Google Scholar] [CrossRef]
- Hannou, S.A.; Haslam, D.E.; McKeown, N.M.; Herman, M.A. Fructose metabolism and metabolic disease. J. Clin. Investig. 2018, 128, 545–555. [Google Scholar] [CrossRef]
- Legeza, B.; Marcolongo, P.; Gamberucci, A.; Varga, V.; Bánhegyi, G.; Benedetti, A.; Odermatt, A. Fructose, Glucocorticoids and Adipose Tissue: Implications for the Metabolic Syndrome. Nutrients 2017, 9, 426. [Google Scholar] [CrossRef]
- Stanhope, K.L.; Schwarz, J.M.; Keim, N.L.; Griffen, S.C.; Bremer, A.A.; Graham, J.L.; Hatcher, B.; Cox, C.L.; Dyachenko, A.; Zhang, W.; et al. Consuming fructose-sweetened, not glucose-sweetened, beverages increases visceral adiposity and lipids and decreases insulin sensitivity in overweight/obese humans. J. Clin. Investig. 2009, 119, 1322–1334. [Google Scholar] [CrossRef]
- Varma, V.; Boros, L.G.; Nolen, G.T.; Chang, C.-W.; Wabitsch, M.; Beger, R.D.; Kaput, J. Metabolic fate of fructose in human adipocytes: A targeted 13C tracer fate association study. Metabolomics 2015, 11, 529–544. [Google Scholar] [CrossRef] [PubMed]
- Varma, V.; Boros, L.G.; Nolen, G.T.; Chang, C.-W.; Wabitsch, M.; Beger, R.D.; Kaput, J. Fructose Alters Intermediary Metabolism of Glucose in Human Adipocytes and Diverts Glucose to Serine Oxidation in the One–Carbon Cycle Energy Producing Pathway. Metabolites 2015, 5, 364–385. [Google Scholar] [CrossRef] [PubMed]
- Tedeschi, P.M.; Markert, E.K.; Gounder, M.; Lin, H.; Dvorzhinski, D.; Dolfi, S.C.; Chan, L.L.Y.; Qiu, J.; DiPaola, R.S.; Hirshfield, K.M.; et al. Contribution of serine, folate and glycine metabolism to the ATP, NADPH and purine requirements of cancer cells. Cell Death Dis. 2013, 4, e877. [Google Scholar] [CrossRef] [PubMed]
- Pektas, M.B.; Koca, H.B.; Sadi, G.; Akar, F. Dietary Fructose Activates Insulin Signaling and Inflammation in Adipose Tissue: Modulatory Role of Resveratrol. BioMed Res. Int. 2016, 2016, 8014252. [Google Scholar] [CrossRef]
- Rodrigues, D.F.; Henriques, M.C.D.C.; Oliveira, M.C.; Menezes-Garcia, Z.; Marques, P.E.; Souza, D.D.G.; Menezes, G.B.; Teixeira, M.M.; Ferreira, A.V.M. Acute intake of a high-fructose diet alters the balance of adipokine concentrations and induces neutrophil influx in the liver. J. Nutr. Biochem. 2014, 25, 388–394. [Google Scholar] [CrossRef]
- Zhang, D.-M.; Jiao, R.-Q.; Kong, L.-D. High Dietary Fructose: Direct or Indirect Dangerous Factors Disturbing Tissue and Organ Functions. Nutrients 2017, 9, 335. [Google Scholar] [CrossRef]
- DiNicolantonio, J.J.; Mehta, V.; Onkaramurthy, N.; O’Keefe, J.H. Fructose-induced inflammation and increased cortisol: A new mechanism for how sugar induces visceral adiposity. Prog. Cardiovasc. Dis. 2018, 61, 3–9. [Google Scholar] [CrossRef]
- Marek, G.; Pannu, V.; Shanmugham, P.; Pancione, B.; Mascia, D.; Crosson, S.; Ishimoto, T.; Sautin, Y.Y. Adiponectin Resistance and Proinflammatory Changes in the Visceral Adipose Tissue Induced by Fructose Consumption via Ketohexokinase-Dependent Pathway. Diabetes 2015, 64, 508–518. [Google Scholar] [CrossRef]
- Kovačević, S.; Brkljačić, J.; Milutinović, D.V.; Gligorovska, L.; Bursać, B.; Elaković, I.; Djordjevic, A. Fructose Induces Visceral Adipose Tissue Inflammation and Insulin Resistance Even Without Development of Obesity in Adult Female but Not in Male Rats. Front. Nutr. 2021, 8, 749328. [Google Scholar] [CrossRef]
- Wang, Y.; Qi, W.; Song, G.; Pang, S.; Peng, Z.; Li, Y.; Wang, P. High-Fructose Diet Increases Inflammatory Cytokines and Alters Gut Microbiota Composition in Rats. Mediat. Inflamm. 2020, 2020, 6672636. [Google Scholar] [CrossRef]
- Alkhouri, N.; Dixon, L.J.; Feldstein, A.E. Lipotoxicity in nonalcoholic fatty liver disease: Not all lipids are created equal. Expert Rev. Gastroenterol. Hepatol. 2009, 3, 445–451. [Google Scholar] [CrossRef]
- Della Corte, K.W.; Perrar, I.; Penczynski, K.J.; Schwingshackl, L.; Herder, C.; Buyken, A.E. Effect of dietary sugar intake on biomarkers of subclinical inflammation: A systematic review and meta-analysis of intervention studies. Nutrients 2018, 10, 606. [Google Scholar] [CrossRef] [PubMed]
- Jaiswal, N.; Agrawal, S.; Agrawal, A. High fructose-induced metabolic changes enhance inflammation in human dendritic cells. Clin. Exp. Immunol. 2019, 197, 237–249. [Google Scholar] [CrossRef] [PubMed]
- Pimentel, G.; Burton, K.J.; Rosikiewicz, M.; Freiburghaus, C.; von Ah, U.; Münger, L.H.; Pralong, F.P.; Vionnet, N.; Greub, G.; Badertscher, R.; et al. Blood lactose after dairy product intake in healthy men. Br. J. Nutr. 2017, 118, 1070–1077. [Google Scholar] [CrossRef] [PubMed]
- Conte, F.; van Buuringen, N.; Voermans, N.C.; Lefeber, D.J. Galactose in human metabolism, glycosylation and congenital metabolic diseases: Time for a closer look. Biochim. et Biophys. Acta (BBA) Gen. Subj. 2021, 1865, 129898. [Google Scholar] [CrossRef] [PubMed]
- Krycer, J.R.; Elkington, S.D.; Diaz-Vegas, A.; Cooke, K.C.; Burchfield, J.G.; Fisher-Wellman, K.H.; Cooney, G.J.; Fazakerley, D.J.; James, D.E. Mitochondrial oxidants, but not respiration, are sensitive to glucose in adipocytes. J. Biol. Chem. 2020, 295, 99–110. [Google Scholar] [CrossRef] [PubMed]
- Krishna, M.S.; Revathy, V.M.; Jaleel, A. Adipocytes utilize sucrose as an energy source—Effect of different carbohydrates on adipocyte differentiation. J. Cell. Physiol. 2020, 235, 891–899. [Google Scholar] [CrossRef] [PubMed]
- Remigante, A.; Morabito, R.; Spinelli, S.; Trichilo, V.; Loddo, S.; Sarikas, A.; Dossena, S.; Marino, A. D-Galactose Decreases Anion Exchange Capability through Band 3 Protein in Human Erythrocytes. Antioxidants 2020, 9, 689. [Google Scholar] [CrossRef]
- Hajam, Y.A.; Rani, R.; Ganie, S.Y.; Sheikh, T.A.; Javaid, D.; Qadri, S.S.; Pramodh, S.; Alsulimani, A.; Alkhanani, M.F.; Harakeh, S.; et al. Oxidative Stress in Human Pathology and Aging: Molecular Mechanisms and Perspectives. Cells 2022, 11, 552. [Google Scholar] [CrossRef]
- Umbayev, B.; Askarova, S.; Almabayeva, A.; Saliev, T.; Masoud, A.-R.; Bulanin, D. Galactose-Induced Skin Aging: The Role of Oxidative Stress. Oxidative Med. Cell. Longev. 2020, 2020, 7145656. [Google Scholar] [CrossRef]
- Sasaoka, N.; Imamura, H.; Kakizuka, A. A Trace Amount of Galactose, a Major Component of Milk Sugar, Allows Maturation of Glycoproteins during Sugar Starvation. Iscience 2018, 10, 211–221. [Google Scholar] [CrossRef]
- Bouwman, L.M.S.; Fernández-Calleja, J.M.S.; van der Stelt, I.; Oosting, A.; Keijer, J.; van Schothorst, E.M. Replacing Part of Glucose with Galactose in the Postweaning Diet Protects Female But Not Male Mice from High-Fat Diet–Induced Adiposity in Later Life. J. Nutr. 2019, 149, 1140–1148. [Google Scholar] [CrossRef] [PubMed]
- Kanuri, G.; Spruss, A.; Wagnerberger, S.; Bischoff, S.C.; Bergheim, I. Role of tumor necrosis factor α (TNFα) in the onset of fructose-induced nonalcoholic fatty liver disease in mice. J. Nutr. Biochem. 2011, 22, 527–534. [Google Scholar] [CrossRef] [PubMed]
- Pan, L.-L.; Deng, Y.-Y.; Wang, R.; Wu, C.; Li, J.; Niu, W.; Yang, Q.; Bhatia, M.; Gudmundsson, G.H.; Agerberth, B.; et al. Lactose Induces Phenotypic and Functional Changes of Neutrophils and Macrophages to Alleviate Acute Pancreatitis in Mice. Front. Immunol. 2018, 9, 751. [Google Scholar] [CrossRef] [PubMed]
- Paasela, M.; Kolho, K.-L.; Vaarala, O.; Honkanen, J. Lactose inhibits regulatory T-cell-mediated suppression of effector T-cell interferon-γ and IL-17 production. Br. J. Nutr. 2014, 112, 1819–1825. [Google Scholar] [CrossRef]
- Jones, N.; Blagih, J.; Zani, F.; Rees, A.; Hill, D.G.; Jenkins, B.J.; Bull, C.J.; Moreira, D.; Bantan, A.I.M.; Cronin, J.G.; et al. Fructose reprogrammes glutamine-dependent oxidative metabolism to support LPS-induced inflammation. Nat. Commun. 2021, 12, 1209. [Google Scholar] [CrossRef]
- Braun, C.; Weichhart, T. mTOR-dependent immunometabolism as Achilles’ heel of anticancer therapy. Eur. J. Immunol. 2021, 51, 3161–3175. [Google Scholar] [CrossRef]
- Hortová-Kohoutková, M.; Lázničková, P.; Frič, J. How immune-cell fate and function are determined by metabolic pathway choice: The bioenergetics underlying the immune response. BioEssays 2021, 43, e2000067. [Google Scholar] [CrossRef]
- Menk, A.V.; Scharping, N.E.; Moreci, R.S.; Zeng, X.; Guy, C.; Salvatore, S.; Bae, H.; Xie, J.; Young, H.A.; Wendell, S.G.; et al. Early TCR Signaling Induces Rapid Aerobic Glycolysis Enabling Distinct Acute T Cell Effector Functions. Cell Rep. 2018, 22, 1509–1521. [Google Scholar] [CrossRef]
- Lee, M.K.S.; Al-Sharea, A.; Shihata, W.A.; Veiga, C.B.; Cooney, O.D.; Fleetwood, A.J.; Flynn, M.C.; Claeson, E.; Palmer, C.S.; Lancaster, G.I.; et al. Glycolysis Is Required for LPS-Induced Activation and Adhesion of Human CD14+CD16− Monocytes. Front. Immunol. 2019, 10, 2054. [Google Scholar] [CrossRef]
- Pan, J.H.; Kim, H.-S.; Beane, K.E.; Montalbano, A.M.; Lee, J.H.; Kim, Y.J.; Kim, J.H.; Kong, B.C.; Kim, S.; Park, J.-W.; et al. IDH2 Deficiency Aggravates Fructose-Induced NAFLD by Modulating Hepatic Fatty Acid Metabolism and Activating Inflammatory Signaling in Female Mice. Nutrients 2018, 10, 679. [Google Scholar] [CrossRef] [PubMed]
- He, J.; Jiao, X.; Sun, X.; Huang, Y.; Xu, P.; Xue, Y.; Fu, T.; Liu, J.; Li, Z. Short-Term High Fructose Intake Impairs Diurnal Oscillations in the Murine Cornea. Investig. Opthalmology Vis. Sci. 2021, 62, 22. [Google Scholar] [CrossRef] [PubMed]
- Gambaro, S.E.; Zubiría, M.G.; Portales, A.E.; Rey, M.A.; Rumbo, M.; Giovambattista, A. M1 macrophage subtypes activation and adipocyte dysfunction worsen during prolonged consumption of a fructose-rich diet. J. Nutr. Biochem. 2018, 61, 173–182. [Google Scholar] [CrossRef] [PubMed]
- Leibowitz, A.; Rehman, A.; Paradis, P.; Schiffrin, E.L. Role of T Regulatory Lymphocytes in the Pathogenesis of High-Fructose Diet–Induced Metabolic Syndrome. Hypertension 2013, 61, 1316–1321. [Google Scholar] [CrossRef] [PubMed]
- Lee, E.; Kim, N.; Kang, J.; Yoon, S.; Lee, H.-A.; Jung, H.; Kim, S.-H.; Kim, I. Activated pathogenic Th17 lymphocytes induce hypertension following high-fructose intake in Dahl salt-sensitive (SS) but not Dahl salt-resistant (SR) rats. Dis. Model. Mech. 2020, 13, dmm044107. [Google Scholar] [CrossRef] [PubMed]
- Sasoh, T.; Kugo, H.; Kondo, Y.; Miyamoto, K.; Minami, M.; Higashihara, M.; Kawamoto, H.; Takeshita, F.; Moriyama, T.; Zaima, N. Different effects of high-fat and high-sucrose diets on the physiology of perivascular adipose tissues of the thoracic and abdominal aorta. Adipocyte 2021, 10, 412–423. [Google Scholar] [CrossRef]
- Lawless, S.J.; Kedia-Mehta, N.; Walls, J.F.; McGarrigle, R.; Convery, O.; Sinclair, L.V.; Navarro, M.N.; Murray, J.; Finlay, D.K. Glucose represses dendritic cell-induced T cell responses. Nat. Commun. 2017, 8, 15620. [Google Scholar] [CrossRef]
- Zhu, J.; Yang, W.; Zhou, X.; Zöphel, D.; Soriano-Baguet, L.; Dolgener, D.; Carlein, C.; Hof, C.; Zhao, R.; Ye, S.; et al. High Glucose Enhances Cytotoxic T Lymphocyte-Mediated Cytotoxicity. Front. Immunol. 2021, 12, 689337. [Google Scholar] [CrossRef]
- Castro, M.C.; Villagarcía, H.; Nazar, A.; Arbeláez, L.G.; Massa, M.L.; Del Zotto, H.; Ríos, J.L.; Schinella, G.; Francini, F. Cacao extract enriched in polyphenols prevents endocrine-metabolic disturbances in a rat model of prediabetes triggered by a sucrose rich diet. J. Ethnopharmacol. 2020, 247, 112263. [Google Scholar] [CrossRef]
- Millet, P.; Vachharajani, V.; McPhail, L.; Yoza, B.; McCall, C.E. GAPDH Binding to TNF-α mRNA Contributes to Posttranscriptional Repression in Monocytes: A Novel Mechanism of Communication between Inflammation and Metabolism. J. Immunol. 2016, 196, 2541–2551. [Google Scholar] [CrossRef]
- Hasegawa, H.; Matsumoto, T. Mechanisms of Tolerance Induction by Dendritic Cells In Vivo. Front. Immunol. 2018, 9, 350. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharjee, J.; Kumar, J.M.; Arindkar, S.; Das, B.; Pramod, U.; Juyal, R.C.; Majumdar, S.S.; Nagarajan, P. Role of immunodeficient animal models in the development of fructose induced NAFLD. J. Nutr. Biochem. 2014, 25, 219–226. [Google Scholar] [CrossRef] [PubMed]
- Nishimura, S.; Manabe, I.; Nagasaki, M.; Eto, K.; Yamashita, H.; Ohsugi, M.; Otsu, M.; Hara, K.; Ueki, K.; Sugiura, S.; et al. CD8+ effector T cells contribute to macrophage recruitment and adipose tissue inflammation in obesity. Nat. Med. 2009, 15, 914–920. [Google Scholar] [CrossRef] [PubMed]
- Lynch, L.A.; O’Connell, J.M.; Kwasnik, A.K.; Cawood, T.J.; O’Farrelly, C.; O’Shea, D. Are Natural Killer Cells Protecting the Metabolically Healthy Obese Patient? Obesity 2009, 17, 601–605. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Jin, W.; Wu, R.; Li, J.; Park, S.-A.; Tu, E.; Zanvit, P.; Xu, J.; Liu, O.; Cain, A.; et al. High Glucose Intake Exacerbates Autoimmunity through Reactive-Oxygen-Species-Mediated TGF-β Cytokine Activation. Immunity 2019, 51, 671–681.e5. [Google Scholar] [CrossRef] [PubMed]
- Zeng, H.; Chi, H. The interplay between regulatory T cells and metabolism in immune regulation. Oncoimmunology 2013, 2, e26586. [Google Scholar] [CrossRef] [PubMed]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Barbosa, P.; Carvalho, E. Are Dietary Sugars Potent Adipose Tissue and Immune Cell Modulators? Diabetology 2023, 4, 30-45. https://doi.org/10.3390/diabetology4010005
Barbosa P, Carvalho E. Are Dietary Sugars Potent Adipose Tissue and Immune Cell Modulators? Diabetology. 2023; 4(1):30-45. https://doi.org/10.3390/diabetology4010005
Chicago/Turabian StyleBarbosa, Pedro, and Eugenia Carvalho. 2023. "Are Dietary Sugars Potent Adipose Tissue and Immune Cell Modulators?" Diabetology 4, no. 1: 30-45. https://doi.org/10.3390/diabetology4010005
APA StyleBarbosa, P., & Carvalho, E. (2023). Are Dietary Sugars Potent Adipose Tissue and Immune Cell Modulators? Diabetology, 4(1), 30-45. https://doi.org/10.3390/diabetology4010005