
Beyond Circumstantial Evidence on Wildlife–Vehicle Collisions During COVID-19 Lockdown: A Deterministic vs. Probabilistic Multi-Year Analysis from a Mediterranean Island



Abstract
1. Introduction
2. Materials and Methods
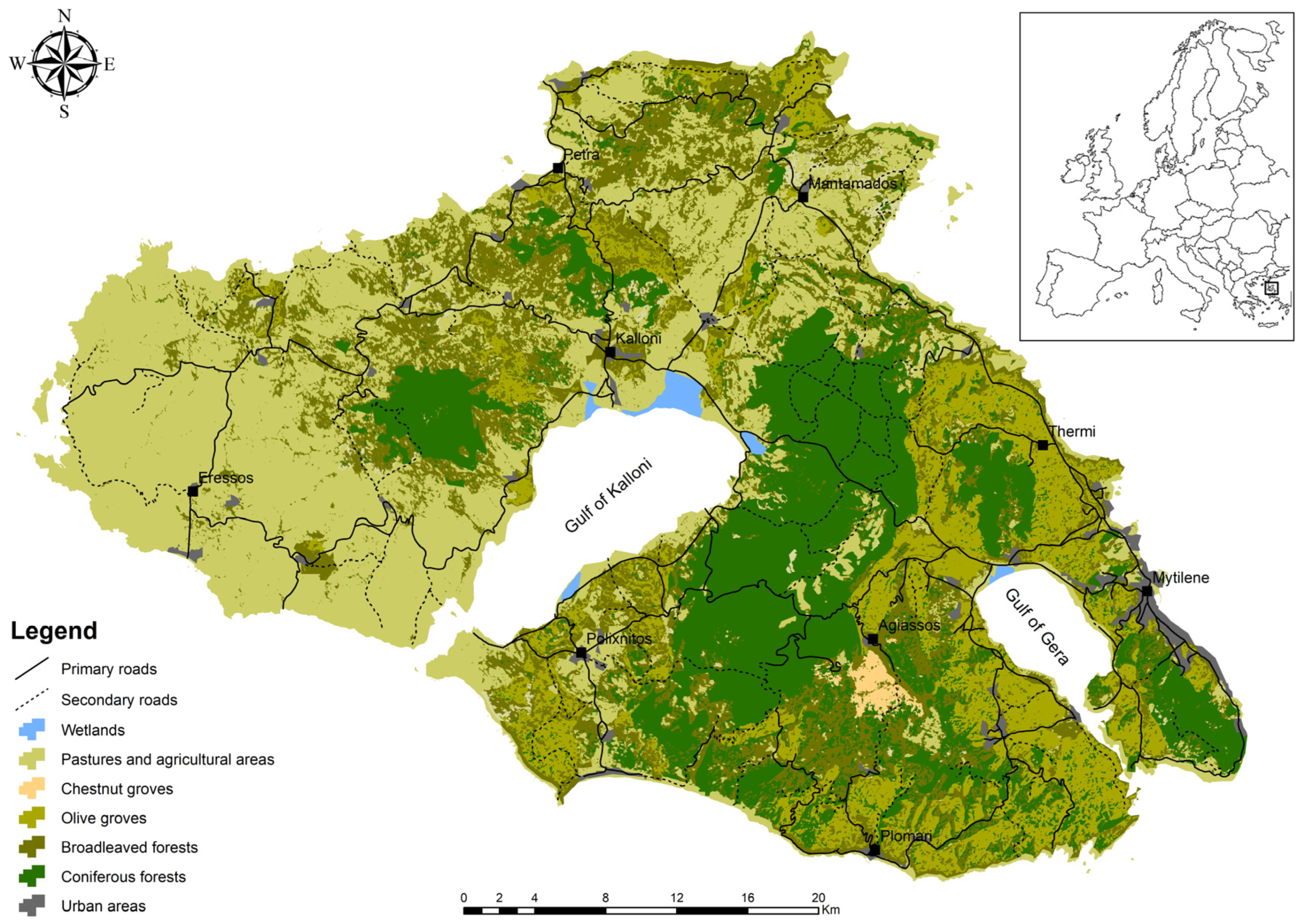
2.1. Study Area
2.2. Characterization and Mapping of the Road Network
2.3. Studied Taxa
2.4. Road Mortality Survey
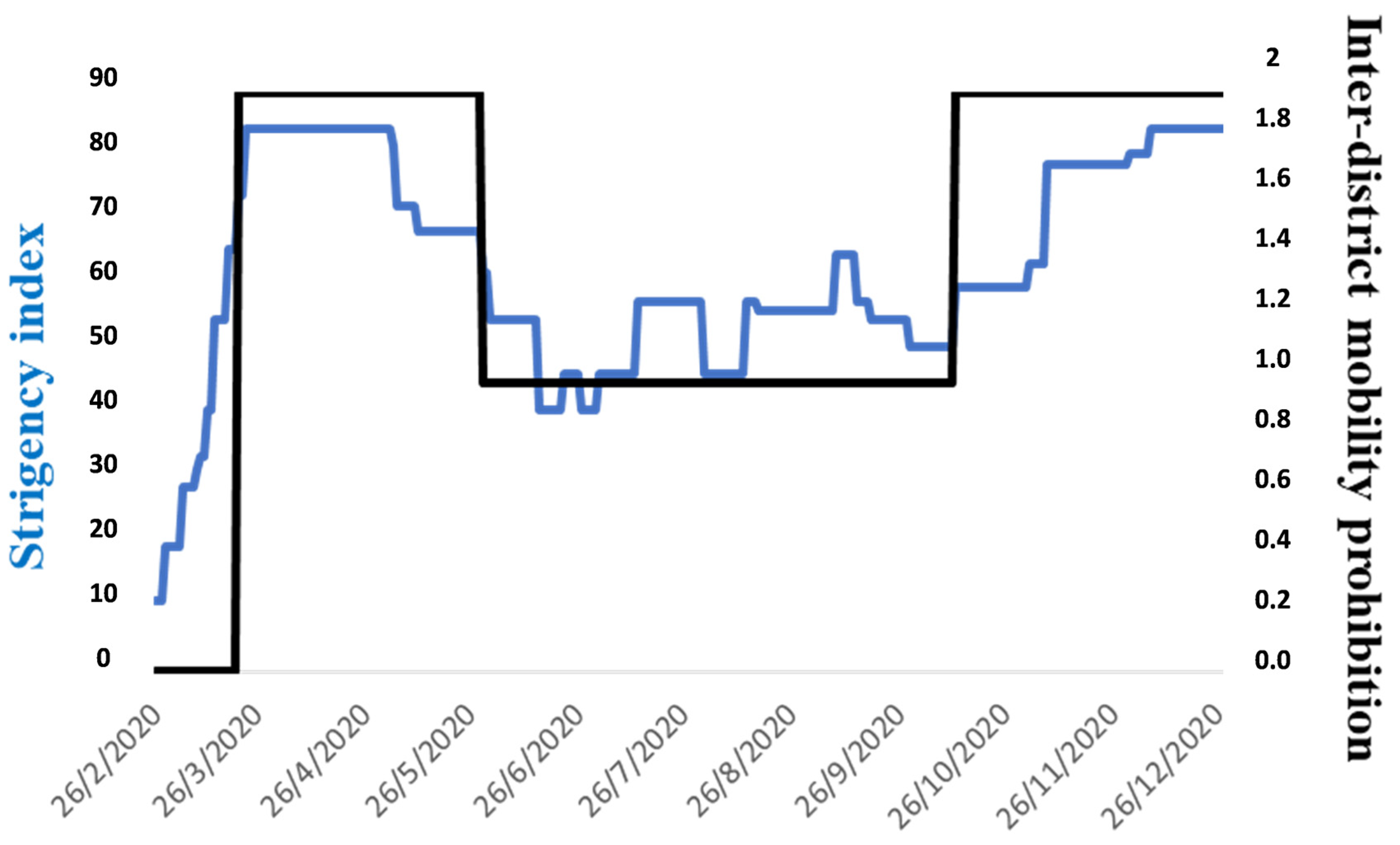
2.5. Calculation of Additional WVC (2018–2022) Against WVC During Lockdown Confinement
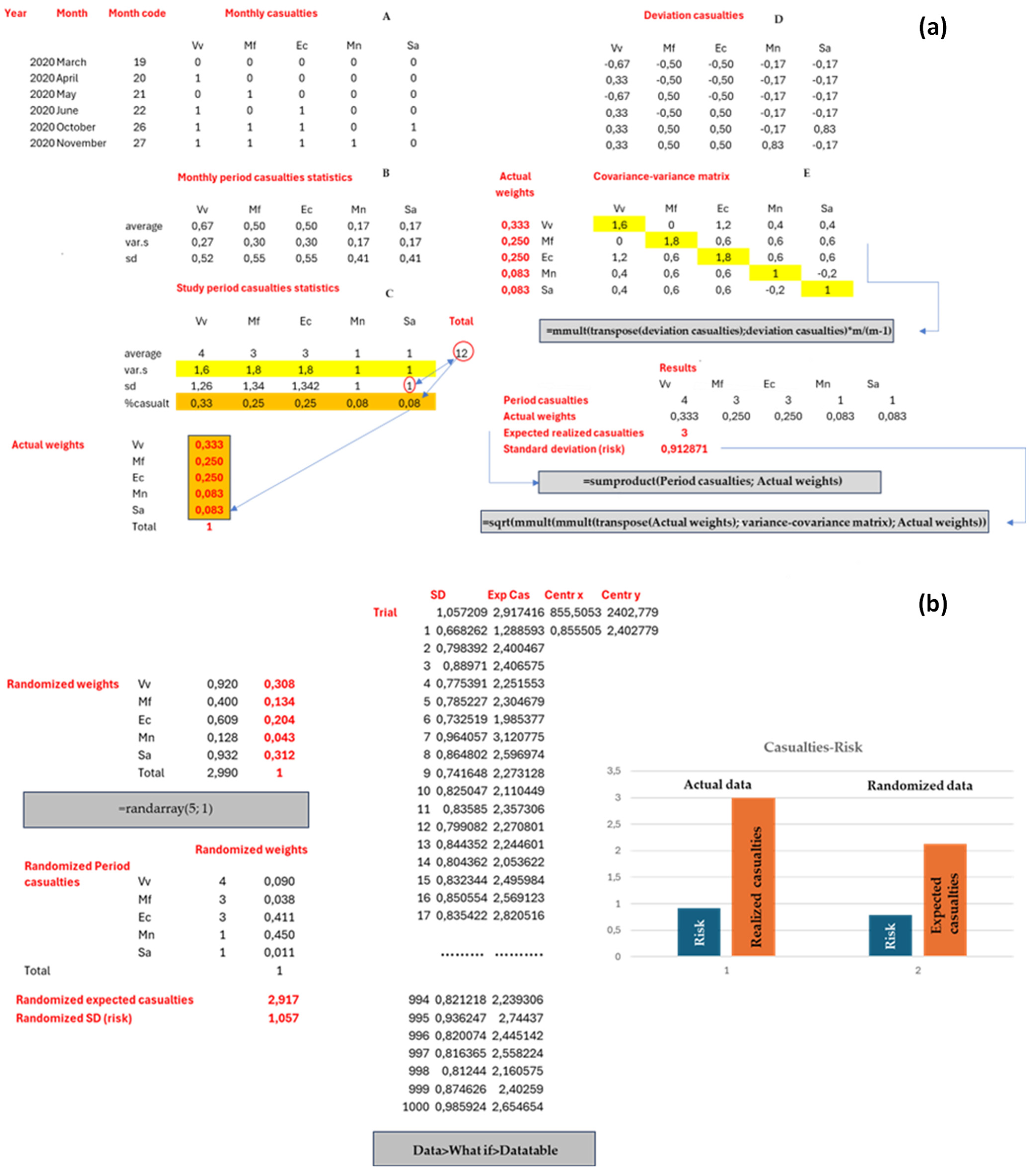
2.6. WVC as a Poisson Process Problem
3. Results
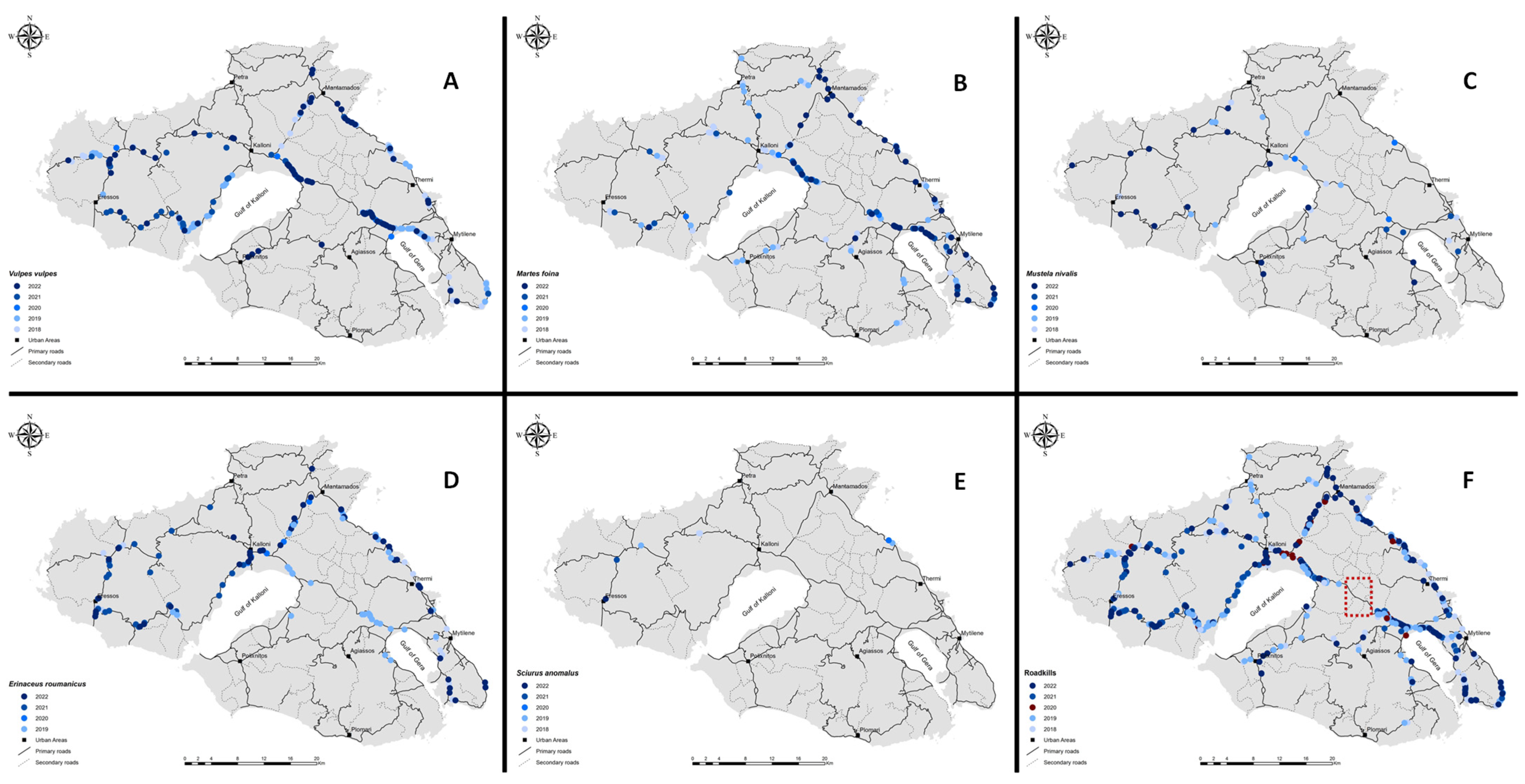
3.1. Spatiotemporal Distribution of WVC (2018–2022)
3.2. Comparative Assessment of WVC Under Mobility Restriction and Normal Conditions
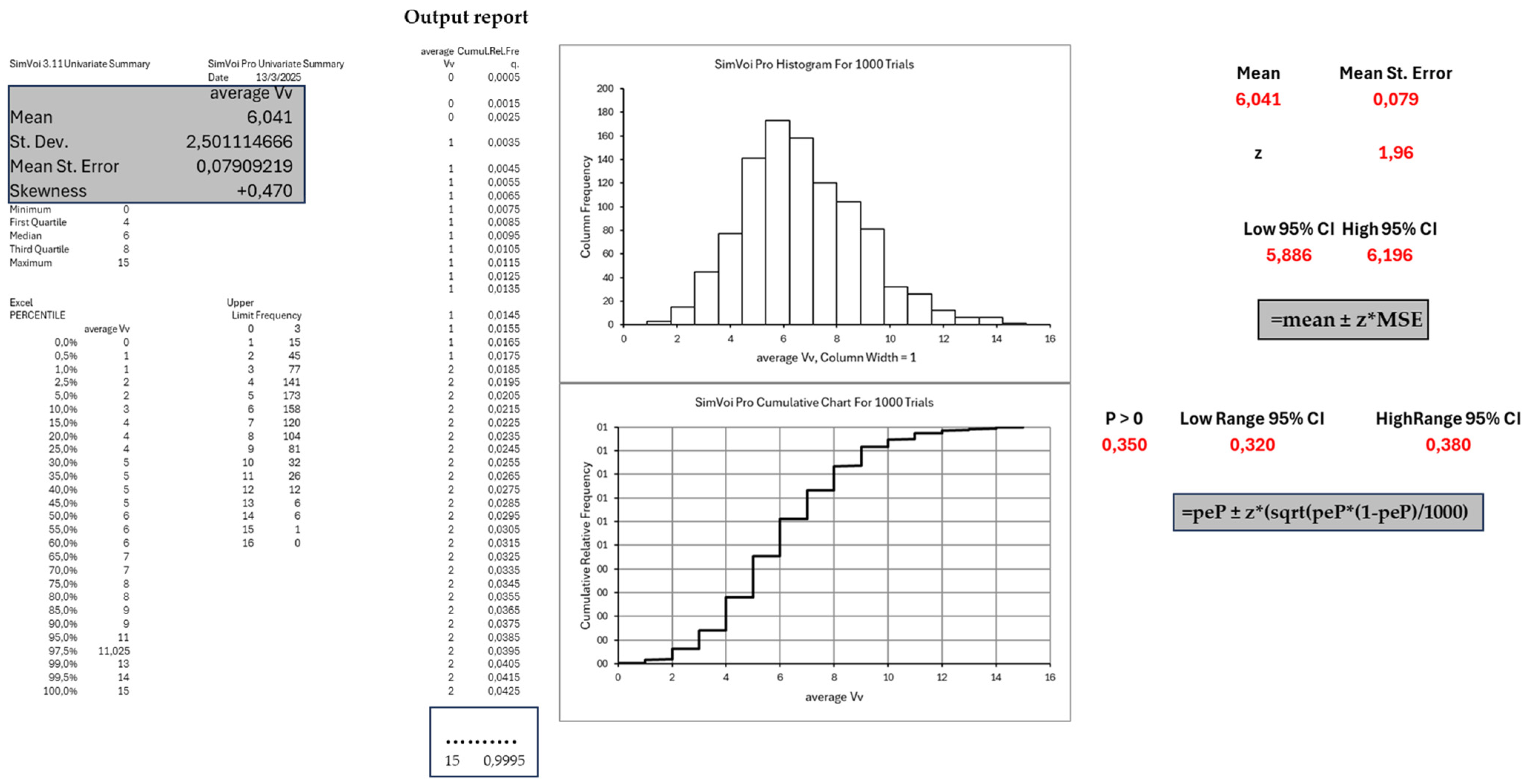
3.3. Probabilistic Modeling of Species-Level WVC Occurrence
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable/Factor | Expressions | Units | Typical Ranges |
|---|---|---|---|
| Vehicle | |||
| Vehicle speed | sv | km.h−1 | 50–90 |
| Distance traveled d in time t | km | - | |
| Vehicle length | λ | m (km × 10−3) | 3.8–4.5 |
| Average vehicle fleet length | m (km × 10−3) | 4.2 | |
| Human reaction time to trouble | τ | s (h/60) | 1–1.5 |
| The minimum separation between vehicles | km | 0.02–0.05 | |
| Number of vehicles per length of the lane | number | - | |
| Congestion factor | - | ||
| Number of vehicles passing the crossing point per unit of time | - | ||
| The average interval between vehicles | km | - | |
| Time to transport N vehicles over distance d | - | ||
| Trip speed of N vehicles over distance d | - | ||
| Vehicular travel capacity | - | ||
| Animal | |||
| Roaming animal mean daily speed | sw | km.h−1 | 1–25 (species-specific) |
| Daily home range | w | km2 | 0.01–13 |
| Population at risk | R = ind/(LA) | Ind/km2 | 0.5–15 |
| Daily roaming ratio | day−1 | 0.25–0.5 | |
| Motorway | |||
| Total length | L | km | 488.67 |
| Length in the animal hotspot area | d | km | 1–20 |
| Lanes/direction | x | number | 1–2 |
| Width | w | m | 5–7 |
| Lethal area | d × w | km2 | - |
| Critical distance | km | 0.1–0.5 | |
| WVC rate | WVCr | Ind.day−1 (or y−1) | - |
References
- Corlett, R.T.; Primack, R.B.; Devictor, V.; Maas, B.; Goswami, V.R.; Bates, A.E.; Koh, L.P.; Regan, T.J.; Loyola, R.; Pakeman, R.J.; et al. Impacts of the coronavirus pandemic on biodiversity conservation. Biol. Conserv. 2020, 246, 108571. [Google Scholar] [CrossRef]
- Schwartz, M.W.; Glikman, J.A.; Cook, C.N. The COVID-19 pandemic: A learnable moment for conservation. Conserv. Sci. Pract. 2020, 2, e255. [Google Scholar] [CrossRef]
- Troumbis, A.Y. Testing the socioeconomic determinants of COVID-19 pandemic hypothesis with aggregated Human Development Index. J. Epidemiol. Community Health 2021, 75, 414–415. [Google Scholar] [CrossRef]
- Bates, A.E.; Primack, R.B.; Moraga, P.; Duarte, C.M. COVID-19 pandemic and associated lockdown as a “Global Human Confinement Experiment” to investigate biodiversity conservation. Biol. Conserv. 2020, 248, 108665. [Google Scholar] [CrossRef]
- Perkins, S.E.; Shilling, F.; Collinson, W. Anthropause Opportunities: Experimental Perturbation of Road Traffic and the Potential Effects on Wildlife. Front. Ecol. Evol. 2022, 10, 833129. [Google Scholar] [CrossRef]
- Rutz, C.; Loretto, M.-C.; Bates, A.E.; Davidson, S.C.; Duarte, C.M.; Jetz, W.; Johnson, M.; Kato, A.; Kays, R.; Mueller, T.; et al. COVID-19 lockdown allows researchers to quantify the effects of human activity on wildlife. Nat. Ecol. Evol. 2020, 4, 1156–1159. [Google Scholar] [CrossRef] [PubMed]
- Hale, T.; Angrist, N.; Goldszmidt, R.; Kira, B.; Petherick, A.; Phillips, T.; Webster, S.; Cameron-Blake, E.; Hallas, L.; Majumdar, S.; et al. A global panel database of pandemic policies (Oxford COVID-19 Government Response Tracker). Nat. Hum. Behav. 2021, 5, 529–538. [Google Scholar] [CrossRef]
- Oxford COVID-19 Government Response Tracker. Available online: https://www.bsg.ox.ac.uk/research/covid-19-government-response-tracker (accessed on 19 July 2023).
- Primack, R.B.; Bates, A.E.; Duarte, C.M. The conservation and ecological impacts of the COVID-19 pandemic. Biol. Conserv. 2021, 260, 109204. [Google Scholar] [CrossRef] [PubMed]
- Bates, A.E.; Primack, R.B.; Biggar, B.S.; Bird, T.J.; Clinton, M.E.; Command, R.J.; Richards, C.; Shellard, M.; Geraldi, N.R.; Vergara, V.; et al. Global COVID-19 lockdown highlights humans as both threats and custodians of the environment. Biol. Conserv. 2021, 263, 109175. [Google Scholar] [CrossRef]
- Loh, H.C.; Looi, I.; Ch’ng, A.S.H.; Goh, K.W.; Ming, L.C.; Ang, K.H. Positive global environmental impacts of the COVID-19 pandemic lockdown: A review. GeoJournal 2022, 87, 4425–4437. [Google Scholar] [CrossRef]
- Derryberry, E.P.; Phillips, J.N.; Derryberry, G.E.; Blum, M.J.; Luther, D. Singing in a silent spring: Birds respond to a half-century soundscape reversion during the COVID-19 shutdown. Science 2020, 370, 575–579. [Google Scholar] [CrossRef] [PubMed]
- Souza, C.N.; Rodrigues, A.C.; Correia, R.A.; Normande, I.C.; Costa, H.C.M.; Guedes-Santos, J.; Malhado, A.C.M.; Carvalho, A.R.; Ladle, R.J. No visit, no interest: How COVID-19 has affected public interest in world’s national parks. Biol. Conserv. 2021, 256, 109015. [Google Scholar] [CrossRef]
- Sánchez-Clavijo, L.M.; Martínez-Callejas, S.J.; Acevedo-Charry, O.; Diaz-Pulido, A.; Gómez-Valencia, B.; Ocampo-Peñuela, N.; Ocampo, D.; Olaya-Rodríguez, M.H.; Rey-Velasco, J.C.; Soto-Vargas, C.; et al. Differential reporting of biodiversity in two citizen science platforms during COVID-19 lockdown in Colombia. Biol. Conserv. 2021, 256, 109077. [Google Scholar] [CrossRef] [PubMed]
- Hochachka, W.M.; Alonso, H.; Gutiérrez-Expósito, C.; Miller, E.; Johnston, A. Regional variation in the impacts of the COVID-19 pandemic on the quantity and quality of data collected by the project eBird. Biol. Conserv. 2021, 254, 108974. [Google Scholar] [CrossRef]
- Asari, Y. Decreased traffic volume during COVID-19 did not reduce roadkill on fenced highway network in Japan. Landsc. Ecol. Eng. 2022, 18, 121–124. [Google Scholar] [CrossRef]
- Basak, S.M.; O’Mahony, D.T.; Lesiak, M.; Basak, A.K.; Ziółkowska, E.; Kaim, D.; Hossain, M.S.; Wierzbowska, I.A. Animal-vehicle collisions during the COVID-19 lockdown in early 2020 in the Krakow metropolitan region, Poland. Sci. Rep. 2022, 12, 7572. [Google Scholar] [CrossRef]
- Raymond, S.; Spencer, M.; Chadwick, E.A.; Madden, J.R.; Perkins, S.E. The impact of the COVID-19 lockdowns on wildlife–vehicle collisions in the UK. J. Anim. Ecol. 2023, 92, 1244–1255. [Google Scholar] [CrossRef]
- García-Martínez-de-Albéniz, Í.; Ruiz-de-Villa, J.A.; Rodriguez-Hernandez, J. Impact of COVID-19 Lockdown on Wildlife-Vehicle Collisions in NW of Spain. Sustainability 2022, 14, 4849. [Google Scholar] [CrossRef]
- LeClair, G.; Chatfield, M.W.H.; Wood, Z.; Parmelee, J.; Frederick, C.A. Influence of the COVID-19 pandemic on amphibian road mortality. Conserv. Sci. Pract. 2021, 3, e535. [Google Scholar] [CrossRef]
- Stiles, J.; Kar, A.; Lee, J.; Miller, H.J. Lower Volumes, Higher Speeds: Changes to Crash Type, Timing, and Severity on Urban Roads from COVID-19 Stay-at-Home Policies. Transp. Res. Rec. J. Transp. Res. Board 2023, 2677, 15–27. [Google Scholar] [CrossRef]
- Łopucki, R.; Kitowski, I.; Perlińska-Teresiak, M.; Klich, D. How Is Wildlife Affected by the COVID-19 Pandemic? Lockdown Effect on the Road Mortality of Hedgehogs. Animals 2021, 11, 868. [Google Scholar] [CrossRef] [PubMed]
- Manenti, R.; Mori, E.; Di Canio, V.; Mercurio, S.; Picone, M.; Caffi, M.; Brambilla, M.; Ficetola, G.F.; Rubolini, D. The good, the bad and the ugly of COVID-19 lockdown effects on wildlife conservation: Insights from the first European locked down country. Biol. Conserv. 2020, 249, 108728. [Google Scholar] [CrossRef] [PubMed]
- Bíl, M.; Andrášik, R.; Cícha, V.; Arnon, A.; Kruuse, M.; Langbein, J.; Náhlik, A.; Niemi, M.; Pokorny, B.; Colino-Rabanal, V.J.; et al. COVID-19 related travel restrictions prevented numerous wildlife deaths on roads: A comparative analysis of results from 11 countries. Biol. Conserv. 2021, 256, 109076. [Google Scholar] [CrossRef] [PubMed]
- Driessen, M.M. COVID-19 restrictions provide a brief respite from the wildlife roadkill toll. Biol. Conserv. 2021, 256, 109012. [Google Scholar] [CrossRef]
- Abraham, J.O.; Mumma, M.A. Elevated wildlife-vehicle collision rates during the COVID-19 pandemic. Sci. Rep. 2021, 11, 20391. [Google Scholar] [CrossRef]
- Ibisch, P.L.; Hoffmann, M.T.; Kreft, S.; Pe’er, G.; Kati, V.; Biber-Freudenberger, L.; DellaSala, D.A.; Vale, M.M.; Hobson, P.R.; Selva, N. A global map of roadless areas and their conservation status. Science 2016, 354, 1423–1427. [Google Scholar] [CrossRef]
- Bíl, M.; Heigl, F.; Janoška, Z.; Vercayie, D.; Perkins, S.E. Benefits and challenges of collaborating with volunteers: Examples from National Wildlife Roadkill Reporting Systems in Europe. J. Nat. Conserv. 2020, 54, 125798. [Google Scholar] [CrossRef]
- Grilo, C.; Koroleva, E.; Andrášik, R.; Bíl, M.; González-Suárez, M. Roadkill risk and population vulnerability in European birds and mammals. Front. Ecol. Environ. 2020, 18, 323–328. [Google Scholar] [CrossRef]
- Pagany, R. Wildlife-vehicle collisions—Influencing factors, data collection and research methods. Biol. Conserv. 2020, 251, 108758. [Google Scholar] [CrossRef]
- Chowdhury, D. Statistical physics of vehicular traffic and some related systems. Phys. Rep. 2000, 329, 199–329. [Google Scholar] [CrossRef]
- Nagatani, T. Traffic dispersion and its mapping to one-sided ballistic deposition. Phys. A Stat. Mech. Its Appl. 2007, 376, 641–648. [Google Scholar] [CrossRef]
- Olson, D.D.; Bissonette, J.A.; Cramer, P.C.; Bunnell, K.D.; Coster, D.C.; Jackson, P.J. How does variation in winter weather affect deer—Vehicle collision rates? Wildl. Biol. 2015, 21, 80–87. [Google Scholar] [CrossRef]
- Santos, R.A.L.; Mota-Ferreira, M.; Aguiar, L.M.S.; Ascensão, F. Predicting wildlife road-crossing probability from roadkill data using occupancy-detection models. Sci. Total Environ. 2018, 642, 629–637. [Google Scholar] [CrossRef]
- Gunson, K.E.; Mountrakis, G.; Quackenbush, L.J. Spatial wildlife-vehicle collision models: A review of current work and its application to transportation mitigation projects. J. Environ. Manag. 2011, 92, 1074–1082. [Google Scholar] [CrossRef]
- Valerio, F.; Carvalho, F.; Barbosa, A.M.; Mira, A.; Santos, S.M. Accounting for Connectivity Uncertainties in Predicting Roadkills: A Comparative Approach between Path Selection Functions and Habitat Suitability Models. Environ. Manag. 2019, 64, 329–343. [Google Scholar] [CrossRef] [PubMed]
- Valerio, F.; Basile, M.; Balestrieri, R. The identification of wildlife-vehicle collision hotspots: Citizen science reveals spatial and temporal patterns. Ecol. Process. 2021, 10, 6. [Google Scholar] [CrossRef]
- Kopnina, H.; Washington, H.; Gray, J.; Taylor, B. The ‘future of conservation’ debate: Defending ecocentrism and the Nature Needs Half movement. Biol. Conserv. 2018, 217, 140–148. [Google Scholar] [CrossRef]
- Negi, C.S. Religion and biodiversity conservation: Not a mere analogy. Int. J. Biodivers. Sci. Manag. 2005, 1, 85–96. [Google Scholar] [CrossRef]
- Millennium. Ecosystem Assessment Ecosystems and Human Well-Being: Opportunities and Challenges for Business and Industry; Island Press: Washington, DC, USA, 2005. [Google Scholar]
- Troumbis, A.Y. The circularity entrapment of the ‘Global Human Confinement Experiment’ in conservation culturomics. Biol. Conserv. 2021, 260, 109244. [Google Scholar] [CrossRef]
- Louder, E.; Wyborn, C. Biodiversity narratives: Stories of the evolving conservation landscape. Environ. Conserv. 2020, 47, 251–259. [Google Scholar] [CrossRef]
- Balmford, A.; Knowlton, N. Why Earth Optimism? Science 2017, 356, 225. [Google Scholar] [CrossRef] [PubMed]
- Kitcher, P. Two Approaches to Explanation. J. Philos. 1985, 82, 632. [Google Scholar] [CrossRef]
- Shrader-Frechette, K.S.; McCoy, E.D. Method in Ecology; Cambridge University Press: Cambridge, UK, 1993; ISBN 9780521418614. [Google Scholar]
- Bostrom, N. Existential Risk Prevention as Global Priority. Glob. Policy 2013, 4, 15–31. [Google Scholar] [CrossRef]
- Moynihan, T. Existential risk and human extinction: An intellectual history. Futures 2020, 116, 102495. [Google Scholar] [CrossRef]
- Troumbis, A.Y. Did the COVID-19 Crisis Reframe Public Awareness of Environmental Topics as Humanity’s Existential Risks? A Case from the UK. World 2024, 5, 1194–1210. [Google Scholar] [CrossRef]
- Kesenheimer, J.S.; Greitemeyer, T. A “Lockdown” of Materialism Values and Pro-Environmental Behavior: Short-Term Effects of the COVID-19 Pandemic. Sustainability 2021, 13, 11774. [Google Scholar] [CrossRef]
- Rutz, C. Studying pauses and pulses in human mobility and their environmental impacts. Nat. Rev. Earth Environ. 2022, 3, 157–159. [Google Scholar] [CrossRef]
- Young, N.; Kadykalo, A.N.; Beaudoin, C.; Hackenburg, D.M.; Cooke, S.J. Is the Anthropause a useful symbol and metaphor for raising environmental awareness and promoting reform? Environ. Conserv. 2021, 48, 274–277. [Google Scholar] [CrossRef]
- Zevgolis, Y.G.; Kamatsos, E.; Akriotis, T.; Dimitrakopoulos, P.G.; Troumbis, A.Y. Estimating Productivity, Detecting Biotic Disturbances, and Assessing the Health State of Traditional Olive Groves, Using Nondestructive Phenotypic Techniques. Sustainability 2021, 14, 391. [Google Scholar] [CrossRef]
- Zevgolis, Y.G.; Sazeides, C.I.; Zannetos, S.P.; Grammenou, V.; Fyllas, N.M.; Akriotis, T.; Dimitrakopoulos, P.G.; Troumbis, A.Y. Investigating the effect of resin collection and detecting fungal infection in resin-tapped and non-tapped pine trees, using minimally invasive and non-invasive diagnostics. For. Ecol. Manag. 2022, 524, 120498. [Google Scholar] [CrossRef]
- Christopoulos, A.; Zevgolis, Y.G. A New Invasion of the Common Slider on a Mediterranean Island (Lesvos, Greece): A Potential Threat to Native Terrapin Populations? Diversity 2022, 14, 1018. [Google Scholar] [CrossRef]
- Zevgolis, Y.G.; ZannetoS, P.S.; Akriotis, T. Physiological response of a wild rodent to experimental manipulations in its natural environment using infrared thermography. Hystrix. Ital. J. Mammal. 2022, 33, 80–91. [Google Scholar] [CrossRef]
- Zevgolis, Y.G.; Zotou, S.; Iliou, A.; Christopoulos, A. Subterranean to submarine: Stress-induced locomotor repertoire expansion and aquatic escape in the Anatolian mole rat (Nannospalax xanthodon) under risk of predation. J. Ethol. 2024, 43, 39–43. [Google Scholar] [CrossRef]
- HNMS. Climatic Data for Selected Stations in Greece. Available online: https://www.emy.gr/en/climatic-data?tab=statistics-tab (accessed on 15 December 2024).
- Kosmas, C.; Danalatos, N.G.; Poesen, J.; Van Wesemael, B. The effect of water vapour adsorption on soil moisture content under Mediterranean climatic conditions. Agric. Water Manag. 1998, 36, 157–168. [Google Scholar] [CrossRef]
- Zevgolis, Y.G.; Kouris, A.; Christopoulos, A. Spatiotemporal Patterns and Road Mortality Hotspots of Herpetofauna on a Mediterranean Island. Diversity 2023, 15, 478. [Google Scholar] [CrossRef]
- Frafjord, K. Winter range of a red fox (Vulpes vulpes) group in a northern birch forest. Mamm. Biol. 2004, 69, 342–348. [Google Scholar] [CrossRef]
- Geiser, F. Seasonal Expression of Avian and Mammalian Daily Torpor and Hibernation: Not a Simple Summer-Winter Affair. Front. Physiol. 2020, 11, 434. [Google Scholar] [CrossRef]
- Rutovskaya, M.V.; Diatroptov, M.E.; Kuznetzova, E.V.; Anufriev, A.I.; Feoktistova, N.Y.; Surov, A.V. The Dynamics of Body Temperature of the Eastern European Hedgehog (Erinaceus roumanicus) during Winter Hibernation. Biol. Bull. 2019, 46, 1136–1145. [Google Scholar] [CrossRef]
- Enukidze, G.P. On the biology of the Transcaucasian squirrel. Soobshcheniia Akad. Nauk Gruz. 1960, 17, 101–123. [Google Scholar]
- Zevgolis, Y.G.; Kouris, A.D.; Christopoulos, A.; Leros, M.; Loupou, M.; Rammou, D.-L.; Youlatos, D.; Troumbis, A.Y. Where to Protect? Spatial Ecology and Conservation Prioritization of the Persian Squirrel at the Westernmost Edge of Its Distribution. Land 2025, 14, 876. [Google Scholar] [CrossRef]
- Herr, J.; Schley, L.; Engel, E.; Roper, T.J. Den preferences and denning behaviour in urban stone martens (Martes foina). Mamm. Biol. 2010, 75, 138–145. [Google Scholar] [CrossRef]
- Zub, K.; Sönnichsen, L.; Szafrańska, P.A. Habitat requirements of weasels Mustela nivalis constrain their impact on prey populations in complex ecosystems of the temperate zone. Oecologia 2008, 157, 571–582. [Google Scholar] [CrossRef]
- Kidawa, D.; Kowalczyk, R. The effects of sex, age, season and habitat on diet of the red fox Vulpes vulpes in northeastern Poland. Acta Theriol. 2011, 56, 209–218. [Google Scholar] [CrossRef]
- Zevgolis, Y.G.; Sazeides, C.I.; Bintsi-Frantzi, E.; Kouris, A.D.; Christopoulos, A. Ecological implications of deep pruning: A case report on Persian squirrel nesting in a centennial olive grove on the island of Lesvos, Greece. Hist. Nat. Bulg. 2024, 46, 89–97. [Google Scholar] [CrossRef]
- Markowitz, H.M. Portfolio Selection: Efficient Diversification of Investments, 2nd ed.; Basil Blackwell: Oxford, UK, 1959; ISBN 978-1-55786-108-5. [Google Scholar]
- Zannetos, S.P.; Zevgolis, Y.G.; Christopoulos, A.; Akriotis, T. Assessing habitat suitability of the Persian Squirrel in the island of Lesvos, Greece. Hystrix Ital. J. Mammal. 2022, 33, 193–197. [Google Scholar]
- Zevgolis, Y.G.; Christopoulos, A.; Kalargalis, I.I.; Zannetos, S.P.; Botetzagias, I.; Dimitrakopoulos, P.G. An (Un)Expected Threat for a Regionally Near-Threatened Species: A Predation Case of a Persian Squirrel on an Insular Ecosystem. Animals 2022, 13, 24. [Google Scholar] [CrossRef]
- Loumou, A.; Giourga, C.; Dimitrakopoulos, P.; Koukoulas, S. Tourism Contribution to Agro-Ecosystems Conservation: The Case of Lesbos Island, Greece. Environ. Manag. 2000, 26, 363–370. [Google Scholar] [CrossRef]
- Gaganis, C.M.; Troumbis, A.Y. Bayesian Inference of Human-Made Hazards in Networks of Island Wetlands: The Case of the Aegean Archipelago. Diversity 2025, 17, 54. [Google Scholar] [CrossRef]
- Mosse, C. Sappho of Lesbos. Histoire 1984, 63, 20–23. [Google Scholar]
- Zouros, N.C. The Miocene Petrified Forest of Lesvos, Greece: Research and Geoconservation Activities. Geoconservation Res. 2021, 4. [Google Scholar] [CrossRef]
- ELSTAT. Human Road Mortality Toll 2007–2022. Available online: https://www.statistics.gr/el/statistics/-/publication/SDT03/-A2102_STD04_TS_MM_00_2007_00-2023_F_GR (accessed on 8 February 2025).
- Lantremange, H. Functionalism in ecology and economics: Epistemological affinities and temptations. Biol. Conserv. 2024, 298, 110762. [Google Scholar] [CrossRef]
- Geldmann, J.; Coad, L.; Barnes, M.; Craigie, I.D.; Hockings, M.; Knights, K.; Leverington, F.; Cuadros, I.C.; Zamora, C.; Woodley, S.; et al. Changes in protected area management effectiveness over time: A global analysis. Biol. Conserv. 2015, 191, 692–699. [Google Scholar] [CrossRef]
- Langhammer, P.F.; Bull, J.W.; Bicknell, J.E.; Oakley, J.L.; Brown, M.H.; Bruford, M.W.; Butchart, S.H.M.; Carr, J.A.; Church, D.; Cooney, R.; et al. The positive impact of conservation action. Science 2024, 384, 453–458. [Google Scholar] [CrossRef] [PubMed]
- Grilo, C.; Borda-de-Água, L.; Beja, P.; Goolsby, E.; Soanes, K.; le Roux, A.; Koroleva, E.; Ferreira, F.Z.; Gagné, S.A.; Wang, Y.; et al. Conservation threats from roadkill in the global road network. Glob. Ecol. Biogeogr. 2021, 30, 2200–2210. [Google Scholar] [CrossRef]
- European Commission. Road Safety Statistics in the EU. Available online: https://ec.europa.eu/eurostat/statistics-explained/index.php?oldid=630784 (accessed on 8 February 2025).



| Scientific Name | Common Name | Average Habitat Range * (km2) | Distance of Roaming * (km) | Speed (km/h) | Population Density * (ind/km2) | References |
|---|---|---|---|---|---|---|
| Vulpes vulpes | Red fox | 5.5–13 (rich habitats) 21–52 (poorer habitats) | 10–15 | ~50 (bursts) | 5–6 3–5 | [60,67] |
| Martes foina | Stone marten | 1.3–5.2 (females) 2.6–13 (males) | Several km/night | ~24 (bursts) | 10–12 | [65] |
| Erinaceus roumanicus | Northern White-breasted Hedgehog | 0.1–0.2 | Several km/night | ~6 | 10–15 | [62] |
| Mustela nivalis | Weasel | 0.01–0.1 | Several km (diurnal) | ~25 | 0.5–1 | [66] |
| Sciurus anomalus | Persian squirrel | 0.01–0.02 | 0.5 (diurnal) | ~20 | 2–4 | [64,68] |
| Species | Average | MSE | Low 95% CI | High 95% CI |
|---|---|---|---|---|
| Vulpes vulpes | 6 | 0.0778 | 6.0715 | 6.224 |
| Martes foina | 4.32 | 0.0673 | 4.1881 | 4.320 |
| Erinaceus roumanicus | 3.294 | 0.0571 | 3.1821 | 3.294 |
| Mustela nivalis | 1 | 0.0325 | 0.9363 | 1.000 |
| Sciurus anomalus | 0.532 | 0.0240 | 0.4849 | 0.532 |
| Species | p > 1 | Low 95% PR | High 95% PR | p > Average | Low 95% PR | High 95% PR |
|---|---|---|---|---|---|---|
| Vulpes vulpes | 0.0035 | 6.2239 | 6.2241 | 0.5650 | 0.5644 | 0.5656 |
| Martes foina | 0.011 | 4.3198 | 4.3202 | 0.5690 | 0.5684 | 0.5696 |
| Erinaceus roumanicus | 0.0325 | 3.2937 | 3.2940 | 0.5930 | 0.5924 | 0.5936 |
| Mustela nivalis | 0.3795 | 0.9993 | 1.0000 | 0.3795 | 0.3788 | 0.3802 |
| Sciurus anomalus | 0.2985 | 0.5312 | 0.5320 | 0.2985 | 0.2977 | 0.2993 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Troumbis, A.Y.; Zevgolis, Y.G. Beyond Circumstantial Evidence on Wildlife–Vehicle Collisions During COVID-19 Lockdown: A Deterministic vs. Probabilistic Multi-Year Analysis from a Mediterranean Island. Ecologies 2025, 6, 42. https://doi.org/10.3390/ecologies6020042
Troumbis AY, Zevgolis YG. Beyond Circumstantial Evidence on Wildlife–Vehicle Collisions During COVID-19 Lockdown: A Deterministic vs. Probabilistic Multi-Year Analysis from a Mediterranean Island. Ecologies. 2025; 6(2):42. https://doi.org/10.3390/ecologies6020042
Chicago/Turabian StyleTroumbis, Andreas Y., and Yiannis G. Zevgolis. 2025. "Beyond Circumstantial Evidence on Wildlife–Vehicle Collisions During COVID-19 Lockdown: A Deterministic vs. Probabilistic Multi-Year Analysis from a Mediterranean Island" Ecologies 6, no. 2: 42. https://doi.org/10.3390/ecologies6020042
APA StyleTroumbis, A. Y., & Zevgolis, Y. G. (2025). Beyond Circumstantial Evidence on Wildlife–Vehicle Collisions During COVID-19 Lockdown: A Deterministic vs. Probabilistic Multi-Year Analysis from a Mediterranean Island. Ecologies, 6(2), 42. https://doi.org/10.3390/ecologies6020042