
Assessing Habitat Suitability and Overlap for South China Sika Deer and Sympatric Ungulates

 ,
,
Abstract
1. Introduction
2. Materials and Methods
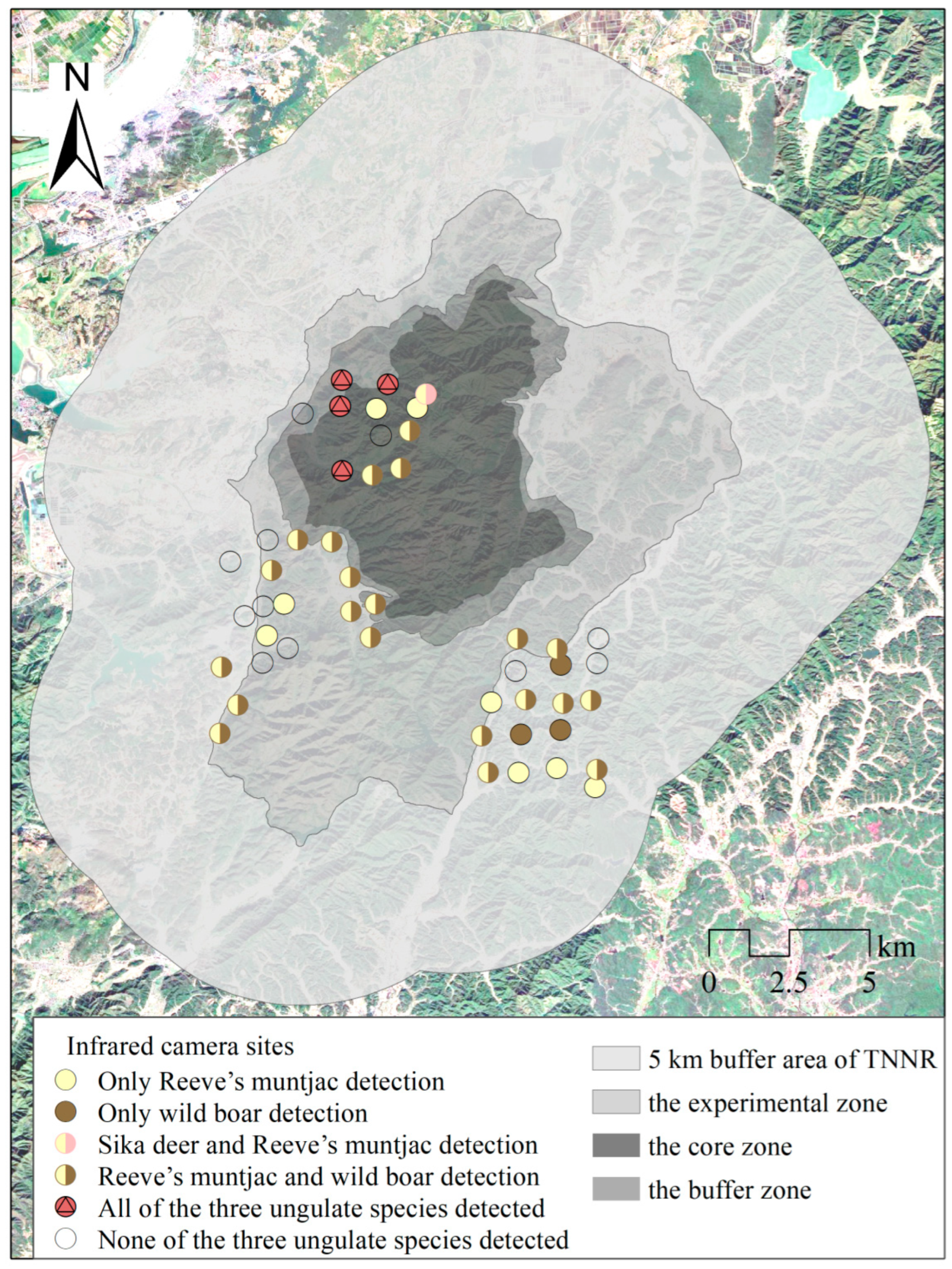
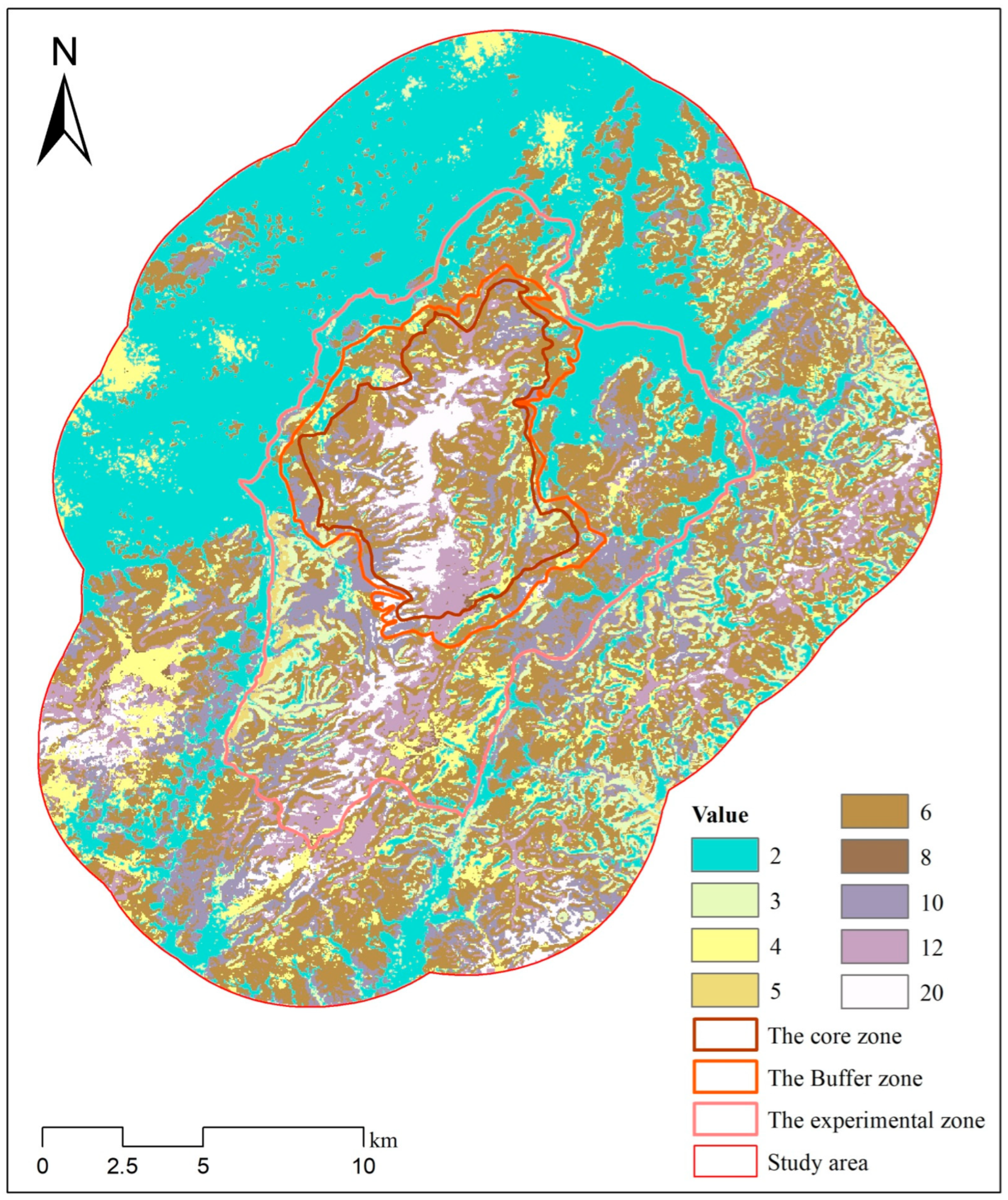
2.1. Study Area
2.2. Species Occurrences
2.3. Environmental Factors
2.4. Habitat Suitability
2.5. Spatial Niche Overlap Analysis
2.5.1. Pianka Index
2.5.2. Analysis of Overlapping Habitats with Different Suitability
3. Result
3.1. Model Performance
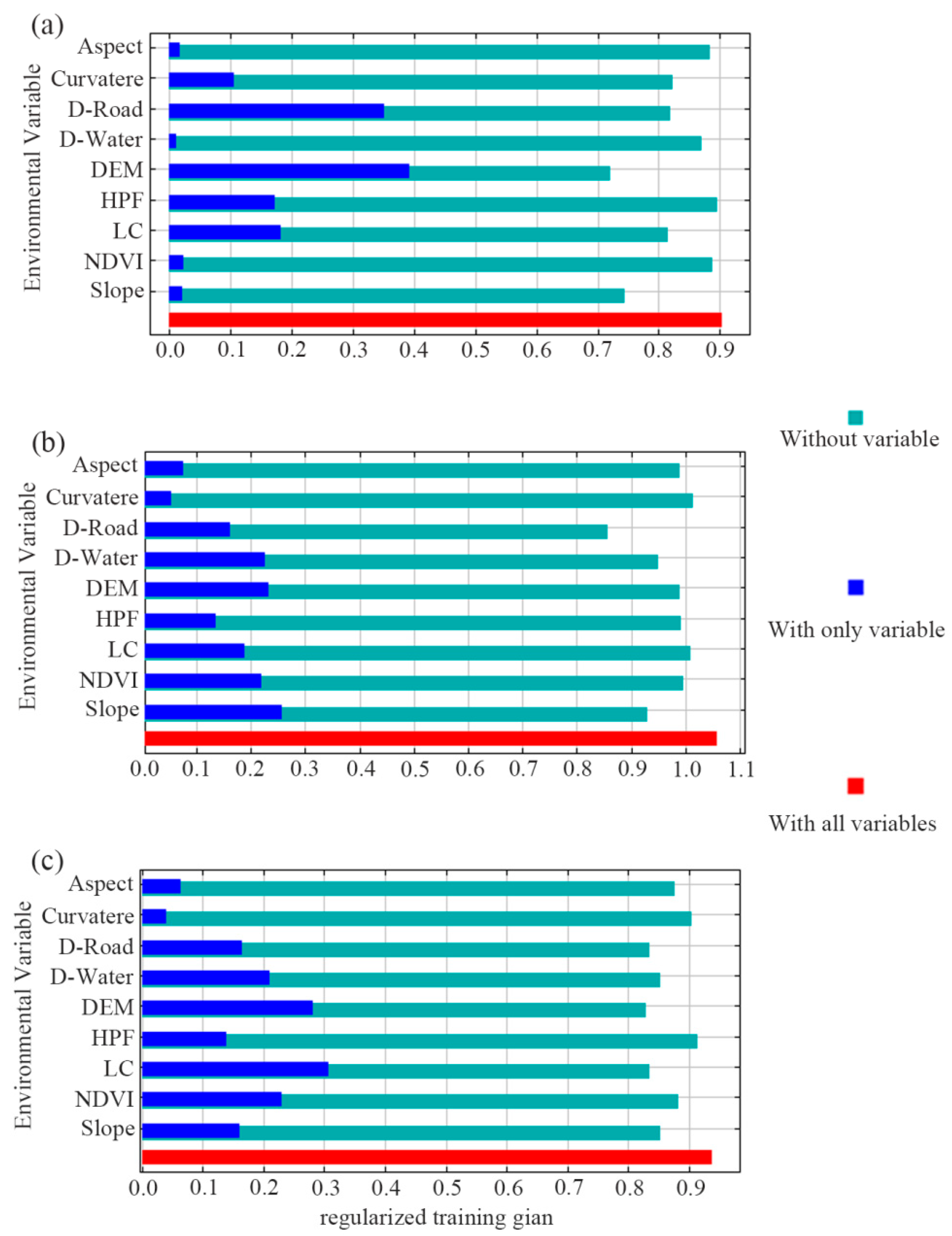
3.2. Effects of Environmental Variables
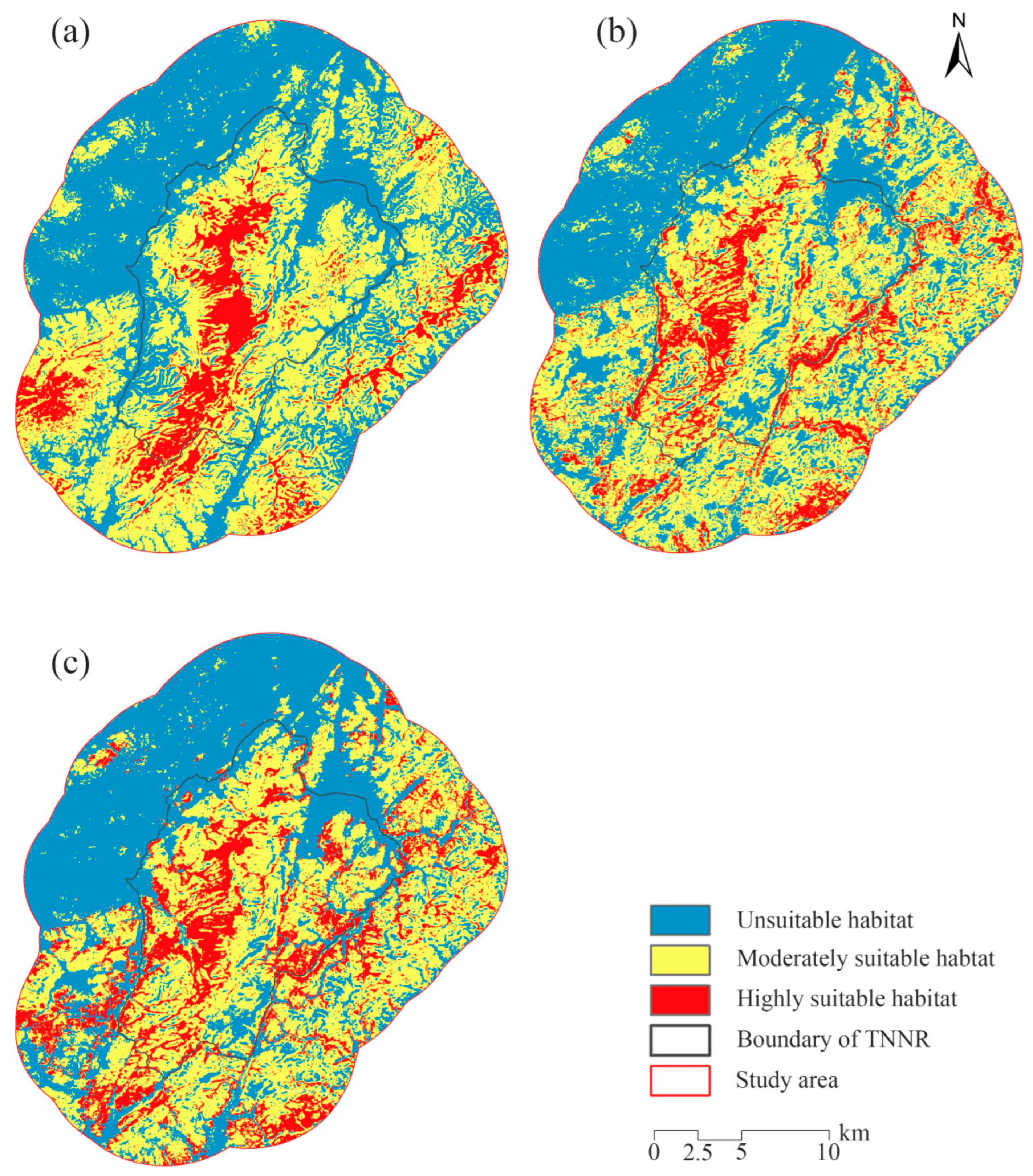
3.3. Habitat Suitability Assessment Results
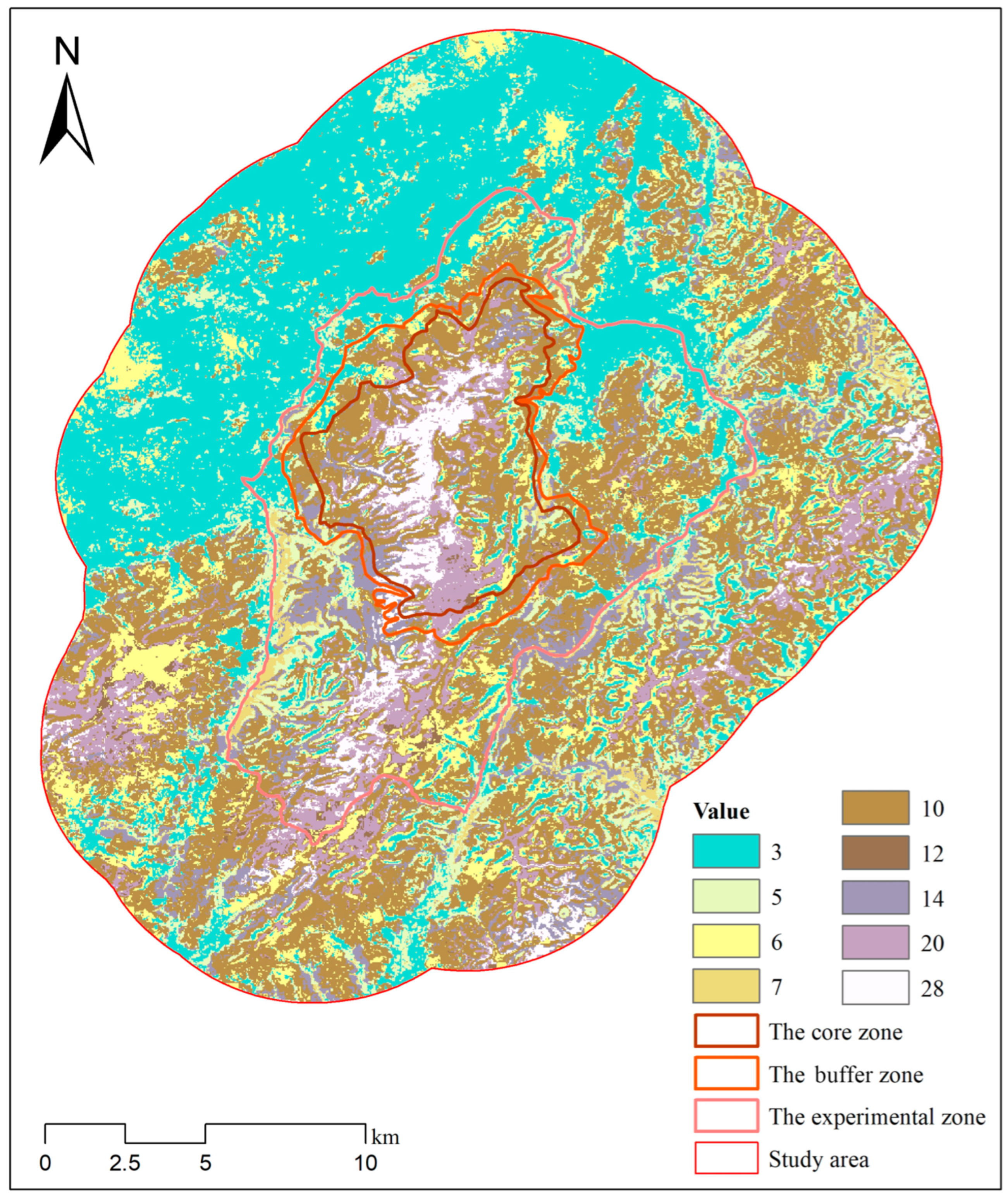
3.4. Habitat Overlap Among the Three Ungulates
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| TNNR | Jiangxi Taohongling Sika Deer National Nature Reserve |
| GIS | Geographic Information System |
| DEM | Digital Elevation Model |
| RFi | Relief |
| LC | Land cover |
| NDVI | Normalized Vegetation Index Values |
| HPF | Human Footprint Index |
| GHF | Global Human Footprint |
| UEMM | Urban Environmental Monitoring and Modeling |
| D-Road | Distance to nearest road |
| D-Water | Distance to nearest water |
| HSMs | Habitat suitability models |
| AUC | Area under the receiver operating characteristic curve |
| MTSS | Maximum training sensitivity plus specificity |
| BTPT | Balance training omission, predicted area, and threshold value |
| HSI | Habitat Suitability Index |
References
- Hofmann, M.M.; Zohner, C.M.; Renner, S.S. Narrow habitat breadth and late-summer emergence increases extinction vulnerability in Central European bees. Proc. R. Soc. B-Biol. Sci. 2019, 286, 20190316. [Google Scholar] [CrossRef] [PubMed]
- Block, W.M.; Brennan, L.A. The Habitat Concept in Ornithology. In Current Ornithology; Power, D.M., Ed.; Springer: Boston, MA, USA, 1993; Volume 11. [Google Scholar]
- Grinnell, J. Field Tests of Theories Concerning Distributional Control. Am. Nat. 1917, 51, 115–128. [Google Scholar] [CrossRef]
- Haddad, N.M.; Brudvig, L.A.; Clobert, J.; Davies, K.F.; Gonzalez, A.; Holt, R.D.; Lovejoy, T.E.; Sexton, J.O.; Austin, M.P.; Collins, C.D.; et al. Habitat fragmentation and its lasting impact on Earth’s ecosystems. Sci. Adv. 2015, 1, e1500052. [Google Scholar] [CrossRef]
- Podgórski, T.; Baś, G.; Jędrzejewska, B.; Sönnichsen, L.; Śnieżko, S.; Jędrzejewski, W.; Okarma, H. Spatiotemporal behavioral plasticity of wild boar (Sus scrofa) under contrasting conditions of human pressure: Primeval forest and metropolitan area. J. Mammal. 2013, 94, 109–119. [Google Scholar] [CrossRef]
- Sáyago, R.; Quesada, M.; Aguilar, R.; Ashworth, L.; Lopezaraiza-Mikel, M.; Martén-Rodríguez, S. Consequences of habitat fragmentation on the reproductive success of two Tillandsia species with contrasting life history strategies. AoB Plants 2018, 10, ply038. [Google Scholar] [CrossRef]
- Bennington, S.; Rayment, W.; Currey, R.; Oldridge, L.; Henderson, S.; Guerra, M.; Brough, T.; Johnston, D.; Corne, C.; Slooten, L.; et al. Long-term stability in core habitat of an endangered population of bottlenose dolphins (Tursiops truncatus): Implications for spatial management. Aquat. Conserv.-Mar. Freshw. Ecosyst. 2021, 31, 665–676. [Google Scholar] [CrossRef]
- Keinath, D.A.; Doak, D.F.; Hodges, K.E.; Prugh, L.R.; Fagan, W.; Sekercioglu, C.H.; Buchart, S.H.M.; Kauffman, M. A global analysis of traits predicting species sensitivity to habitat fragmentation. Glob. Ecol. Biogeogr. 2017, 26, 115–127. [Google Scholar] [CrossRef]
- Kazemi, E.; Kaboli, M.; Khosravi, R.; Khorasani, N. Evaluating the importance of environmental variables on spatial distribution of caspian cobra naja oxiana (Eichwald, 1831) in Iran. Asian Herpetol. Res. 2019, 10, 129–138. [Google Scholar] [CrossRef]
- Kong, F.; Shen, B.; Li, X.; Zhou, Y.X.; Li, Y.K.; Li, J.Q.; Wan, Y.Q.; Zhan, J.W.; Liu, W.H.; Hu, H.J.; et al. The difference of activity rhythm of synchro-distributed ungulates in Taohongling Reserve, Jiangxi Province. Chin. J. Wildl. 2024, 45, 242–250. (In Chinese) [Google Scholar] [CrossRef]
- Moll, R.J.; Killion, A.K.; Montgomery, R.A.; Tambling, C.J.; Hayward, M.W. Spatial patterns of African ungulate aggregation reveal complex but limited risk effects from reintroduced carnivores. Ecology 2016, 97, 1123–1134. [Google Scholar] [CrossRef]
- Bukombe, J.; Kittle, A.; Senzota, R.; Mduma, S.; Fryxell, J.; Sinclair, A.R. Resource selection, utilization and seasons influence spatial distribution of ungulates in the western Serengeti National Park. Afr. J. Ecol. 2018, 56, 3–11. [Google Scholar] [CrossRef]
- Mamo, Y.; Asefa, A.; Mengesha, G. Habitat use of ungulates in bale mountains national Park, Ethiopia. Afr. J. Ecol. 2015, 53, 512–520. [Google Scholar] [CrossRef]
- Phillips, S.J.; Dudík, M. Modeling of species distributions with Maxent: New extensions and a comprehensive evaluation. Ecography 2008, 31, 161–175. Available online: https://www.jstor.org/stable/30244563 (accessed on 20 June 2024). [CrossRef]
- Sun, X.; Long, Z.; Jia, J. A multi-scale Maxent approach to model habitat suitability for the giant pandas in the Qionglai mountain, China. Glob. Ecol. Conserv. 2021, 30, e01766. [Google Scholar] [CrossRef]
- Bukombe, J.; Kittle, A.; Senzota, R.B.; Kija, H.; Mduma, S.; Fryxell, J.M.; Magige, F.; Mligo, C.; Sinclair, A.R.E. The influence of food availability, quality and body size on patch selection of coexisting grazer ungulates in western Serengeti National Park. Wildl. Res. 2019, 46, 54–63. [Google Scholar] [CrossRef]
- Ebrahimi, A.; Farashi, A.; Rashki, A. Habitat suitability of Persian leopard (Panthera pardus saxicolor) in Iran in future. Environ. Earth Sci. 2017, 76, 697. [Google Scholar] [CrossRef]
- Zhong, L.; Zhang, W.; Yang, M.; Wu, S.; Zhi, X.; Zhang, M. Winter diet variation and overlap of sympatric red deer and sika deer in Northeast China. Pol. J. Ecol. 2020, 67, 354–366. [Google Scholar] [CrossRef]
- Ellerman, J.R.; Morrison-Scott, T.C.S. Checklist of Palaearctic and Indian Mammals—Amendments. J. Mammal. 1953, 34, 516–518. [Google Scholar] [CrossRef]
- Mccullough, D.R.; Jiang, Z.G.; Li, C.W. Sika Deer in Mainland China. In Sika Deer; McCullough, D.R., Takatsuki, S., Kaji, K., Eds.; Springer: Tokyo, Japan, 2009. [Google Scholar]
- Yan, L. Jiangxi Pengze discovered sika deer. Wildlife 1983, 3, 40. (In Chinese) [Google Scholar] [CrossRef]
- Zhou, Y. Diversity of Mammals and Birds, Ungulates Activity Rhythm and Habitat Selection by Camera Trapping in the Taohongling. Master’s Thesis, Jiangxi Normal University, Nanchang, China, 2019. [Google Scholar]
- Wu, W.; Li, Y.; Hu, Y. Simulation of potential habitat overlap between red deer (Cervus elaphus) and roe deer (Capreolus capreolus) in northeastern China. PeerJ 2016, 4, e1756. [Google Scholar] [CrossRef]
- Estevo, C.A.; Nagy-Reis, M.B.; Nichols, J.D. When habitat matters: Habitat preferences can modulate co-occurrence patterns of similar sympatric species. PLoS ONE 2017, 12, e0179489. [Google Scholar] [CrossRef] [PubMed]
- Petroelje, T.R.; Kautz, T.M.; Beyer, D.E., Jr.; Belant, J.L. Interference competition between wolves and coyotes during variable prey abundance. Ecol. Evol. 2021, 11, 1413–1431. [Google Scholar] [CrossRef] [PubMed]
- Pokharel, K.P.; Ludwig, T.; Storch, I. Spatial niche partitioning in sub-tropical solitary ungulates: Four-horned antelope and barking deer in Nepal. PLoS ONE 2015, 10, e0117917. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.; Li, Y.; Li, D.; Guan, J.; Xiong, Y.; Hu, J. Habitat suitability assessment of Sichuan sika deer in Tiebu Nature Reserve during periods of green and dry grass. Acta Ecol. Sin. 2014, 34, 135–140. [Google Scholar] [CrossRef]
- Bai, W.; Zhang, J.; He, K.; Zhao, S.; Gu, X.; Hu, J.; Songer, M.; Zhou, C.; Dong, X.; Huang, Q. Implications of habitat overlap between giant panda and sambar for sympatric multi-species conservation. Wildl. Res. 2022, 50, 820–826. [Google Scholar] [CrossRef]
- Liu, T.; Jiang, Z.; Wang, W.; Wang, G.; Song, X.; Xu, A.; Li, C. Changes in habitat suitability and population size of the endangered Przewalski’s gazelle. Glob. Ecol. Conserv. 2023, 43, e02465. [Google Scholar] [CrossRef]
- Morrison, M.L.; Marcot, B.; Mannan, W. Wildlife-habitat relationships: Concepts and applications. Condor Ornithol. Appl. 2012, 109, 980–981. [Google Scholar] [CrossRef]
- Endo, Y.; Takada, H.; Takatsuki, S. Comparison of the food habits of the sika deer (Cervus nippon), the Japanese serow (Capricornis crispus), and the wild boar (Sus. scrofa), sympatric herbivorous mammals from Mt. Asama, central Japan. Mammal Study 2017, 42, 131–140. [Google Scholar] [CrossRef]
- Huang, X.; Yang, J. The 30 m annual land cover datasets and its dynamics in China from 1985 to 2022 [dataset]. Earth Syst. Sci. Data 2023, 13, 3907–3925. [Google Scholar] [CrossRef]
- Chen, X.; Lei, Y. Effects of sample size on accuracy and stability of species distribution models: A comparison of GARP and MaxEnt. In Recent Advances in Computer Science and Information Engineering; Qian, Z., Cao, L., Su, W., Wang, T., Yang, H., Eds.; Springer: Berlin Heidelberg, Germany, 2012; Volume 2, pp. 601–609. [Google Scholar] [CrossRef]
- Qian, Z.; Cao, L.; Su, W.; Wang, T.; Yang, H. (Eds.) Recent Advances in Computer Science and Information Engineering; Lecture Notes in Electrical Engineering; Springer: Berlin/Heidelberg, Germany, 2012; Volume 125. [Google Scholar]
- Pianka, E.R. The structure of lizard communities. Annu. Rev. Ecol. Syst. 1973, 4, 53–74. Available online: http://www.jstor.org/stable/2096804 (accessed on 20 June 2024). [CrossRef]
- Pianka, E.R. Niche overlap and diffuse competition. Proc. Natl. Acad. Sci. USA 1974, 71, 2141–2145. [Google Scholar] [CrossRef] [PubMed]
- Yasuda, M. Monitoring diversity and abundance of mammals with camera traps: A case study on Mount Tsukuba, central Japan. Mammal Study 2004, 29, 37–46. [Google Scholar] [CrossRef]
- Xu, W.; Ouyang, Z.; Jiang, Z.; Zheng, H.; Liu, J. Assessment of giant panda habitat in the Daxiangling Mountain Range, Sichuan, China. Biodivers. Sci. 2006, 14, 223–231. (In Chinese) [Google Scholar] [CrossRef]
- Yan, W.; Wang, Q.; Wang, C. Evaluation of potential breeding habitat distribution with Maxent model for crested ibis in the Qinling-Bashan region. Chin. J. Zool. 2015, 50, 185–193. (In Chinese) [Google Scholar] [CrossRef]
- Duncan, N.P.; Kahl, S.S.; Gray, S.S.; Salice, C.J.; Stevens, R.D. Pronghorn habitat suitability in the Texas Panhandle. J. Wildl. Manag. 2016, 80, 1471–1478. [Google Scholar] [CrossRef]
- Bagchi, S.; Goyal, S.; Sankar, K. Niche relationships of an ungulate assemblage in a dry tropical forest. J. Mammal. 2003, 84, 981–988. [Google Scholar] [CrossRef]
- Newbold, T.; Hudson, L.N.; Hill, S.L.; Contu, S.; Lysenko, I.; Senior, R.A.; Börger, L.; Bennett, D.J.; Choimes, A.; Collen, B.; et al. Global effects of land use on local terrestrial biodiversity. Nature 2015, 520, 45–50. [Google Scholar] [CrossRef]
- Zhang, L.; Dong, L.; Lin, L.; Feng, L.; Yan, F.; Wang, L.; Guo, X.; Luo, A. Asian elephants in China: Estimating population size and evaluating habitat suitability. PLoS ONE 2015, 10, e0124834. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Category | Code | Description | Source |
|---|---|---|---|
| Terrain Factor | Aspect | — | http://www.gscloud.cn/ (accessed on 12 September 2024) |
| Curvature | — | http://www.gscloud.cn/ (accessed on 12 September 2024) | |
| DEM | Digital Elevation Model | http://www.gscloud.cn/ (accessed on 12 September 2024) | |
| Slope | — | http://www.gscloud.cn/ (accessed on 12 September 2024) | |
| Human Disturbance Factor | D-Road | Distance to nearest road | |
| D-Water | Distance to nearest water | http://www.webmap.cn/ (accessed on 13 September 2024) | |
| HPF | Human Pressure Footprint | https://wcshumanfootprint.org/ (accessed on 15 September 2024) | |
| Vegetation Factor | LC | Land cover | http://www.gscloud.cn/ (accessed on 12 September 2024) |
| NDVI | Normalized Difference Vegetation Index | http://www.gscloud.cn/ (accessed on 12 September 2024) |
| Environment Variable | Contribution Rate (%) | ||
|---|---|---|---|
| The Sika Deer | Reeve’s Muntjac | Wild Boar | |
| Aspect | 1.96 | 7.33 | 6.96 |
| Curvature | 7.57 | 3.34 | 2.39 |
| D-Road | 24.92 | 17.66 | 11.22 |
| D-Water | 3.35 | 13.59 | 9.96 |
| DEM | 24.41 | 9.55 | 13.29 |
| HPF | 0.84 | 6.03 | 3.47 |
| LC | 18.40 | 18.07 | 36.75 |
| NDVI | 1.69 | 5.59 | 5.08 |
| Slope | 16.86 | 18.84 | 9.89 |
| Sika Deer Suitability | Reeve’s Muntjac Suitability | Product Value | Area (km2) |
|---|---|---|---|
| 1 (Unsuitable) | 3 (Unsuitable) | 3 | 140.38 |
| 1 (Unsuitable) | 5 (Moderately suitable) | 5 | 52.55 |
| 1 (Unsuitable) | 7 (Highly suitable) | 7 | 8.18 |
| 2 (Moderately suitable) | 3 (Unsuitable) | 6 | 45.13 |
| 2 (Moderately suitable) | 5 (Moderately suitable) | 10 | 149.58 |
| 2 (Moderately suitable) | 7 (Highly suitable) | 14 | 34.20 |
| 4 (Highly suitable) | 3 (Unsuitable) | 12 | 4.55 |
| 4 (Highly suitable) | 5 (Moderately suitable) | 20 | 33.65 |
| 4 (Highly suitable) | 7 (Highly suitable) | 28 | 15.65 |
| Sika Deer Suitability | Wild Boar Suitability | Product Value | Area (km2) |
|---|---|---|---|
| 1 (Unsuitable) | 2 (Unsuitable) | 2 | 161.67 |
| 1 (Unsuitable) | 3 (Moderately suitable) | 3 | 34.43 |
| 1 (Unsuitable) | 5 (Highly suitable) | 5 | 5.01 |
| 2 (Moderately suitable) | 2 (Unsuitable) | 4 | 32.56 |
| 2 (Moderately suitable) | 3 (Moderately suitable) | 6 | 143.71 |
| 2 (Moderately suitable) | 5 (Highly suitable) | 10 | 52.64 |
| 4 (Highly suitable) | 2 (Unsuitable) | 8 | 1.45 |
| 4 (Highly suitable) | 3 (Moderately suitable) | 12 | 32.00 |
| 4 (Highly suitable) | 5 (Highly suitable) | 20 | 20.39 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, J.; Li, Y.; Wang, Z.; Wang, G.; He, S.; Zheng, Y.; Li, C. Assessing Habitat Suitability and Overlap for South China Sika Deer and Sympatric Ungulates. Ecologies 2025, 6, 41. https://doi.org/10.3390/ecologies6020041
Zhang J, Li Y, Wang Z, Wang G, He S, Zheng Y, Li C. Assessing Habitat Suitability and Overlap for South China Sika Deer and Sympatric Ungulates. Ecologies. 2025; 6(2):41. https://doi.org/10.3390/ecologies6020041
Chicago/Turabian StyleZhang, Jing, Yankuo Li, Zhaoyang Wang, Guangyao Wang, Shizhao He, Yu Zheng, and Chunlin Li. 2025. "Assessing Habitat Suitability and Overlap for South China Sika Deer and Sympatric Ungulates" Ecologies 6, no. 2: 41. https://doi.org/10.3390/ecologies6020041
APA StyleZhang, J., Li, Y., Wang, Z., Wang, G., He, S., Zheng, Y., & Li, C. (2025). Assessing Habitat Suitability and Overlap for South China Sika Deer and Sympatric Ungulates. Ecologies, 6(2), 41. https://doi.org/10.3390/ecologies6020041