
Influence of Forest Structural Complexity and Management Intensity on Woodpecker Communities in Mediterranean Chestnut (Castanea sativa) Forests


{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
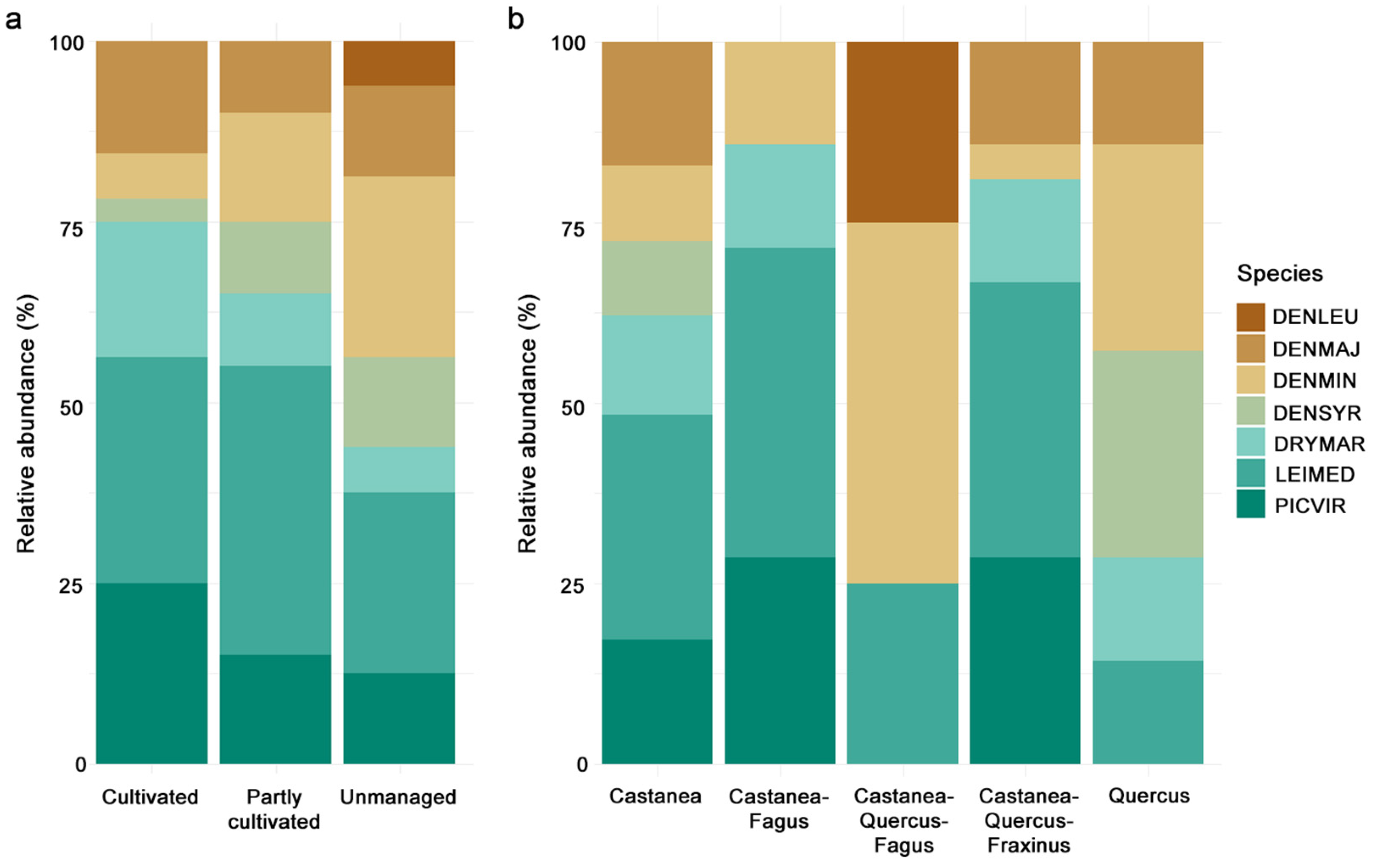
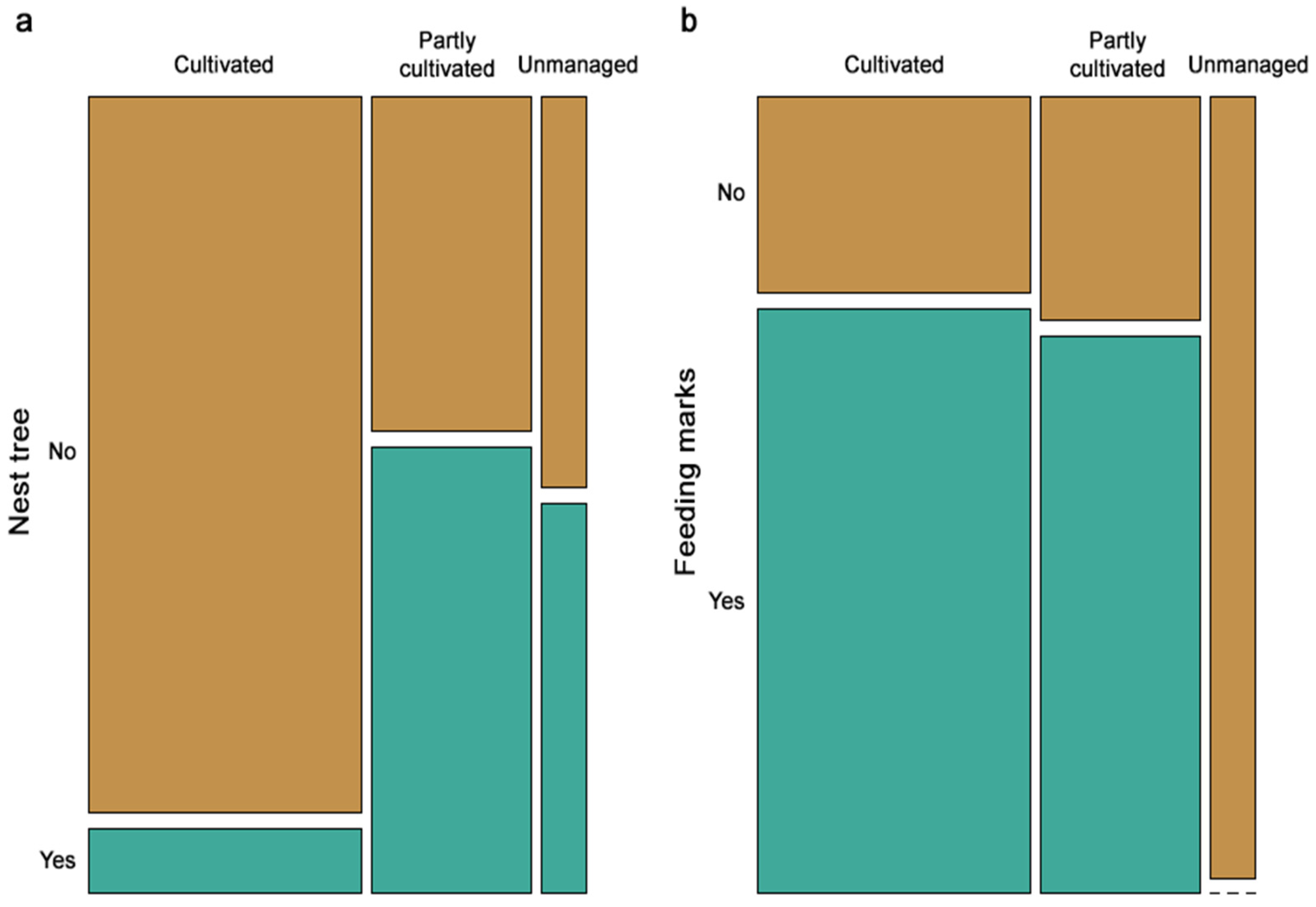
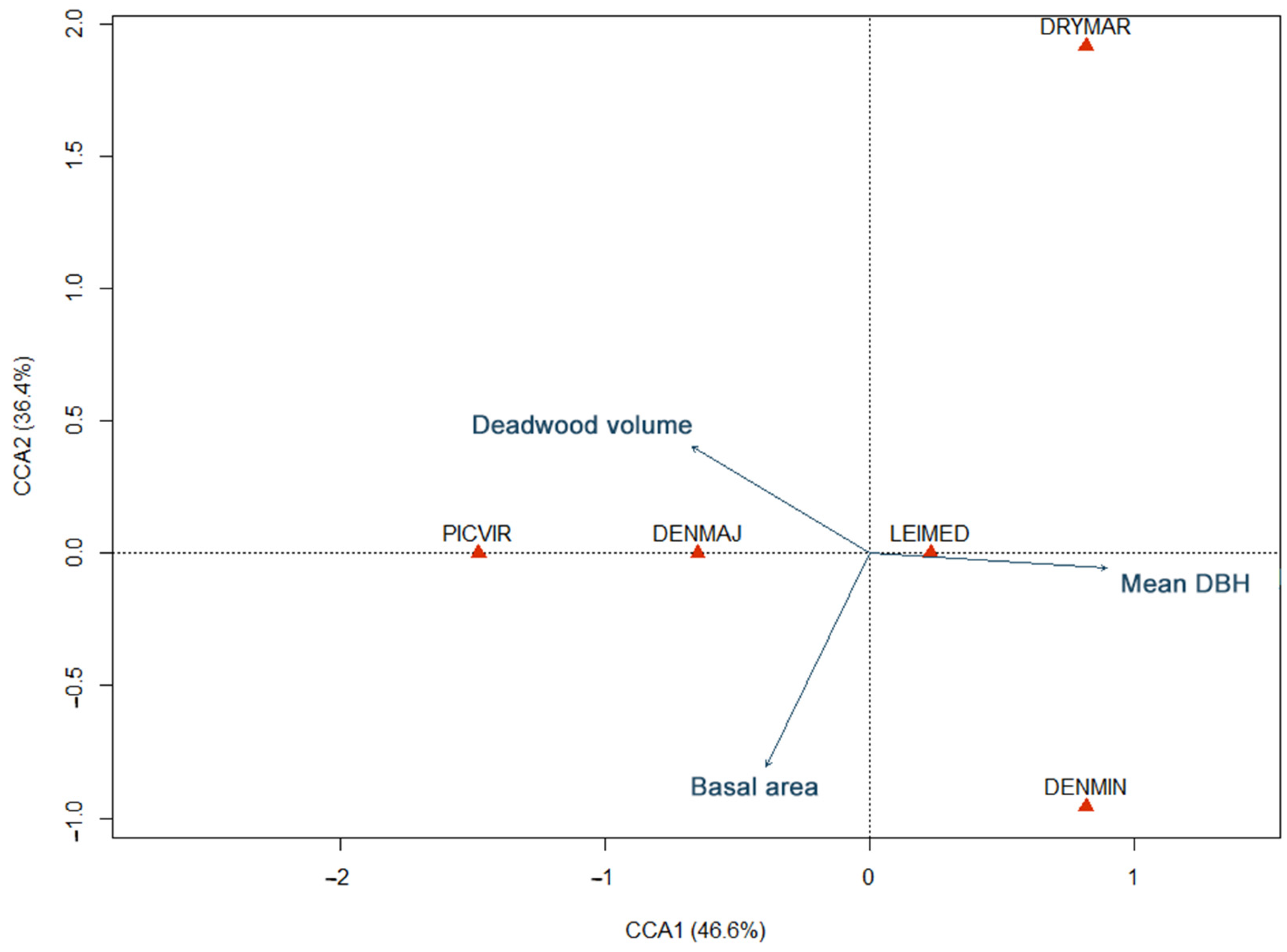
3. Results
4. Discussion
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| DBH | diameter at breast height |
References
- Jose, S. Agroforestry for ecosystem services and environmental benefits: An overview. Agrofor. Syst. 2009, 76, 1–10. [Google Scholar] [CrossRef]
- Baptista, P.; Martins, A.; Tavares, R.M.; Lino-Neto, T. Diversity and fruiting pattern of macrofungi associated with chestnut (Castanea sativa) in the Trás-os-Montes region (Northeast Portugal). Fungal Ecol. 2010, 3, 9–19. [Google Scholar] [CrossRef]
- Venanzi, R.; Picchio, R.; Piovesan, G. Silvicultural and logging impact on soil characteristics in Chestnut (Castanea sativa Mill.) Mediterranean coppice. Ecol. Eng. 2016, 92, 82–89. [Google Scholar] [CrossRef]
- Roces Diaz, J.V.; Varela, E.R.D.; Anta, M.B.; Álvarez, P.Á. Sweet chestnut agroforestry systems in North-western Spain: Classification, spatial distribution and an ecosystem services assessment. For. Syst. 2018, 27, 10. [Google Scholar] [CrossRef]
- Pezzi, G.; Maresi, G.; Conedera, M.; Ferrari, C. Woody species composition of chestnut stands in the Northern Apennines: The result of 200 years of changes in land use. Landsc. Ecol. 2011, 26, 1463–1476. [Google Scholar] [CrossRef]
- Bombelli, A.; Di Paola, A.; Chiriacò, M.V.; Perugini, L.; Castaldi, S.; Valentini, R. Climate change, sustainable agriculture and food systems: The world after the Paris agreement. In Achieving the Sustainable Development Goals Through Sustainable Food Systems; Springer: Berlin/Heidelberg, Germany, 2019; pp. 25–34. [Google Scholar]
- Conedera, M.; Tinner, W.; Krebs, P.; de Rigo, D.; Caudullo, G. Castanea sativa in Europe: Distribution, habitat, usage and threats. In European Atlas of Forest Tree Species; San-Miguel-Ayanz, J., de Rigo, D., Caudullo, G., Houston Durrant, T., Mauri, A., Eds.; Publications Office of the European Union: Luxembourg, 2016; pp. 78–79. [Google Scholar]
- EC (European Comission). Interpretation Manual of European Habitats–EUR27; DG Environment, Nature and Biodiversity: Brussels, Belgium, 2007. [Google Scholar]
- Conedera, M.; Krebs, P.; Tinner, W.; Pradella, M.; Torriani, D. The cultivation of Castanea sativa (Mill.) in Europe, from its origin to its diffusion on a continental scale. Veg. Hist. Archaeobotany 2004, 13, 161–179. [Google Scholar] [CrossRef]
- Pezzi, G.; Gambini, S.; Buldrini, F.; Ferretti, F.; Muzzi, E.; Maresi, G.; Nascimbene, J. Contrasting patterns of tree features, lichen, and plant diversity in managed and abandoned old-growth chestnut orchards of the northern Apennines (Italy). For. Ecol. Manag. 2020, 470, 118207. [Google Scholar] [CrossRef]
- FAO. Production / Crops and Livestock Products. 2023. Available online: https://www.fao.org/faostat/en/#data/QCL (accessed on 19 February 2025).
- Diamandis, S.; Perlerou, C. The mycoflora of the chestnut ecosystems in Greece. For. Snow Landsc. Res. 2001, 76, 499–504. [Google Scholar]
- Konstantinidis, P.; Tsiourlis, G.; Xofis, P.; Buckley, G.P. Taxonomy and ecology of Castanea sativa Mill. forests in Greece. Plant Ecol. 2008, 195, 235–256. [Google Scholar] [CrossRef]
- Remm, J.; Lõhmus, A. Tree cavities in forests–the broad distribution pattern of a keystone structure for biodiversity. For. Ecol. Manag. 2011, 262, 579–585. [Google Scholar] [CrossRef]
- Drever, M.C.; Aitken, K.E.; Norris, A.R.; Martin, K. Woodpeckers as reliable indicators of bird richness, forest health and harvest. Biol. Conserv. 2008, 141, 624–634. [Google Scholar] [CrossRef]
- Menon, T.; Shahabuddin, G. Assessing woodpeckers as indicators of bird diversity and habitat structure in managed forests. Biodivers. Conserv. 2021, 30, 1689–1704. [Google Scholar] [CrossRef]
- Wübbenhorst, J.; Südbeck, P. Woodpeckers as indicators for sustainable forestry. In First Results of a Study in the EU/LIFE–Demonstration Areas Lüneburger Heide und Solling. Demonstration of Methods to Monitor Sustainable Forestry 2001, 1998–2001; EU-LIFE: Barcelona, Spain, 2001. [Google Scholar]
- Ilsøe, S.K.; Kissling, W.D.; Fjeldså, J.; Sandel, B.; Svenning, J.C. Global variation in woodpecker species richness shaped by tree availability. J. Biogeogr. 2017, 44, 1824–1835. [Google Scholar] [CrossRef]
- Martin, K.; Eadie, J.M. Nest webs: A community-wide approach to the management and conservation of cavity-nesting forest birds. For. Ecol. Manag. 1999, 115, 243–257. [Google Scholar] [CrossRef]
- Trzcinski, M.K.; Cockle, K.L.; Norris, A.R.; Edworthy, M.; Wiebe, K.L.; Martin, K. Woodpeckers and other excavators maintain the diversity of cavity-nesting vertebrates. J. Anim. Ecol. 2022, 91, 1251–1265. [Google Scholar] [CrossRef]
- Jusino, M.A.; Hagemeyer, N.D.; Banik, M.T.; Palmer, J.M.; Lindner, D.L.; Smith, M.E.; Koenig, W.D.; Walters, E.L. Fungal communities associated with acorn woodpeckers and their excavations. Fungal Ecol. 2022, 59, 101154. [Google Scholar] [CrossRef]
- Villard, M.A.; Trzcinski, M.K.; Merriam, G. Fragmentation effects on forest birds: Relative influence of woodland cover and configuration on landscape occupancy. Conserv. Biol. 1999, 13, 774–783. [Google Scholar] [CrossRef]
- Carlson, A. The effect of habitat loss on a deciduous forest specialist species: The White-backed Woodpecker (Dendrocopos leucotos). For. Ecol. Manag. 2000, 131, 215–221. [Google Scholar] [CrossRef]
- Basile, M.; Krištín, A.; Mikusiński, G.; Thorn, S.; Żmihorski, M.; Pasinelli, G.; Brockerhoff, E.G. Salvage logging strongly affects woodpecker abundance and reproduction: A meta-analysis. Curr. For. Rep. 2023, 9, 1–14. [Google Scholar] [CrossRef]
- Mortelliti, A.; Fagiani, S.; Battisti, C.; Capizzi, D.; Boitani, L. Independent effects of habitat loss, habitat fragmentation and structural connectivity on forest-dependent birds. Divers. Distrib. 2010, 16, 941–951. [Google Scholar] [CrossRef]
- Porro, Z.; Chiatante, G.; Bogliani, G. Associations between forest specialist birds and composition of woodland habitats in a highly modified landscape. For. Ecol. Manag. 2020, 458, 117732. [Google Scholar] [CrossRef]
- Fernandez, C.; Azkona, P. Influence of forest structure on the density and distribution of the White-backed Woodpecker Dendrocopos leucotos and Black Woodpecker Dryocopus martius in Quinto Real (Spanish western Pyrenees). Bird Study 1996, 43, 305–313. [Google Scholar] [CrossRef]
- Drever, M.C.; Martin, K. Response of woodpeckers to changes in forest health and harvest: Implications for conservation of avian biodiversity. For. Ecol. Manag. 2010, 259, 958–966. [Google Scholar] [CrossRef]
- Kajtoch, L.; Figarski, T.; Pelka, J. The role of forest structural elements in determining the occurrence of two specialist woodpecker species in the Carpathians, Poland. Ornis Fenn. 2013, 90, 23–40. [Google Scholar] [CrossRef]
- Basile, M.; Asbeck, T.; Pacioni, C.; Mikusiński, G.; Storch, I. Woodpecker cavity establishment in managed forests: Relative rather than absolute tree size matters. Wildl. Biol. 2020, 2020, 1–9. [Google Scholar] [CrossRef]
- Laiolo, P.; Rolando, A.; Valsania, V. Avian community structure in sweet chestnut coppiced woods facing natural restoration. Rev. D’écologie 2004, 59, 453–463. [Google Scholar] [CrossRef]
- González-Varo, J.P.; López-Bao, J.V.; Guitián, J. Presence and abundance of the Eurasian nuthatch Sitta europaea in relation to the size, isolation and the intensity of management of chestnut woodlands in the NW Iberian Peninsula. Landsc. Ecol. 2008, 23, 79–89. [Google Scholar] [CrossRef]
- Morelli, F.; Python, A.; Pezzatti, G.B.; Moretti, M. Bird response to woody pastoral management of ancient chestnut orchards: A case study from the southern Alps. For. Ecol. Manag. 2019, 453, 117560. [Google Scholar] [CrossRef]
- Miskos, D. PGI Chestnut Paikou: Benefits and Prospects for the Wider Region. Master’s Thesis, Agricultural University of Athens, Athens, Greece, 2023. [Google Scholar]
- Block, W.M. Foraging ecology of Nuttall’s woodpecker. Auk 1991, 108, 303–318. [Google Scholar]
- Badis, M.; Benchana, I.; Hamdi, N. Nest-site selection by Levaillant’s Woodpecker Picus vaillantii in the Aurès Mountains of northeastern Algeria. Ostrich 2023, 94, 60–64. [Google Scholar] [CrossRef]
- Bibby, C.J.; Burgess, N.D.; Hill, D.A.; Mustoe, S.H. Bird Census Techniques, 2nd ed.; Academic Press: London, UK, 2000. [Google Scholar]
- Dudley, J.; Saab, V. A Field Protocol to Monitor Cavity-Nesting Birds; U.S.D.A. Forest Service, Rocky Mountain Research Station: Fort Collins, CO, USA, 2003.
- Ettwein, A.; Korner, P.; Lanz, M.; Lachat, T.; Kokko, H.; Pasinelli, G. Habitat selection of an old-growth forest specialist in managed forests. Anim. Conserv. 2020, 23, 547–560. [Google Scholar] [CrossRef]
- Fernández, J.M.; Lammertink, M. Resource selection and home range size variation of Atlantic Forest woodpecker species: Implications for selective logging and conservation. For. Ecol. Manag. 2023, 548, 121440. [Google Scholar] [CrossRef]
- Paillet, Y.; Pernot, C.; Boulanger, V.; Debaive, N.; Fuhr, M.; Gilg, O.; Gosselin, F. Quantifying the recovery of old-growth attributes in forest reserves: A first reference for France. For. Ecol. Manag. 2015, 346, 51–64. [Google Scholar] [CrossRef]
- Hunter, M. Wildlife, Forests, and Forestry-Principles of Managing Forests for Biological Diversity; CABI: Wallingford, UK, 1990. [Google Scholar]
- Bettinger, P.; Boston, K.; Siry, J.; Grebner, D. Valuing and characterizing forest conditions. In Forest Management and Planning, 2nd ed.; Academic Press: New York, NY, USA, 2017; pp. 21–64. [Google Scholar]
- R CoreTeam. R: A language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2022. [Google Scholar]
- Oksanen, J.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchin, P.R.; O’hara, R.; Simpson, G.L.; Solymos, P.; Stevens, M.H.H.; Wagner, H. vegan: Community Ecology Package. R Package Version 2.5-7. 2020. Available online: https://CRAN.R-project.org/package=vegan (accessed on 29 January 2025).
- Torchiano, M. Package ‘effsize’: Efficient Effect Size Computation. R Package Version 0.8.1. 2020. Available online: https://CRAN.R-project.org/package=effsize (accessed on 29 January 2025).
- Pedersen, E.J.; Miller, D.L.; Simpson, G.L.; Ross, N. Hierarchical generalized additive models in ecology: An introduction with mgcv. PeerJ 2019, 7, e6876. [Google Scholar] [CrossRef]
- Pasinelli, G. Oaks (Quercus sp.) and only oaks? Relations between habitat structure and home range size of the middle spotted woodpecker (Dendrocopos medius). Biol. Conserv. 2000, 93, 227–235. [Google Scholar] [CrossRef]
- Domokos, E.; Cristea, V. Effects of managed forests structure on woodpeckers (Picidae) in the Niraj valley (Romania): Woodpecker populations in managed forests. North-West. J. Zool. 2014, 10, 131606. [Google Scholar]
- Varasteh Moradi, H.; Sepehri Roshan, Z.; Chamanefar, S. Habitat assessment of Green Woodpecker (Picus viridis) in Golestan National Park using classification tree method. J. Anim. Res. (Iran. J. Biol.) 2018, 30, 495–507. [Google Scholar]
- Stański, T.; Stańska, M.; Goławski, A.; Czeszczewik, D. Foraging site selection of the middle spotted woodpecker (Leiopicus medius L.) in primeval oak-lime-hornbeam forest of the Białowieża National Park: Comparison of breeding and non-breeding seasons. Forests 2021, 12, 837. [Google Scholar] [CrossRef]
- Pasinelli, G.; Hegelbach, J. Characteristics of trees preferred by foraging middle spotted woodpecker Dendrocopos medius in northern Switzerland. Ardea 1997, 85, 203–209. [Google Scholar]
- Delahaye, L.; Monticelli, D.; Lehaire, F.; Rondeux, J.; Claessens, H. Fine-scale habitat selection by two specialist woodpeckers occurring in beech and oak-dominated forests in southern Belgium. Ardeola 2010, 57, 339–362. [Google Scholar]
- Czeszczewik, D.; Walankiewicz, W.; Mitrus, C.; Tumiel, T.; Stanski, T.; Sahel, M.; Bednarczyk, G. Importance of dead wood resources for woodpeckers in coniferous stands of the Białowieża Forest. Bird Conserv. Int. 2013, 23, 414–425. [Google Scholar] [CrossRef]
- Kajtoch, Ł. The importance of traditional orchards for breeding birds: The preliminary study on Central European example. Acta Oecologica 2017, 78, 53–60. [Google Scholar] [CrossRef]
- Michalczuk, J. The importance of non-forest tree stand features for protection of the Syrian Woodpecker Dendrocopos syriacus in agricultural landscape: A case study from South-Eastern Poland. Agrofor. Syst. 2020, 94, 1825–1835. [Google Scholar] [CrossRef]
- Garmendia, A.; Cárcamo, S.; Schwendtner, O. Forest management considerations for conservation of black woodpecker Dryocopus martius and white-backed woodpecker Dendrocopos leucotos populations in Quinto Real (Spanish Western Pyrenees). In Forest Diversity and Management; Springer: Berlin/Heidelberg, Germany, 2006; pp. 339–355. [Google Scholar]
- Pirovano, A.R.; Zecca, G. Black Woodpecker Dryocopus martius habitat selection in the Italian Alps: Implications for conservation in Natura 2000 network. Bird Conserv. Int. 2014, 24, 299–315. [Google Scholar] [CrossRef]
- Olano, M.; Aierbe, T.; Beñaran, H.; Hurtado, R.; Ugarte, J.; Urruzola, A.; Vázquez, J.; Ansorregi, F.; Galdos, A.; Gracianteparaluceta, A. Black woodpecker Dryocopus martius (L., 1758) distribution, abundance, habitat use and breeding performance in a recently colonized region in SW Europe. Munibe Cienc. Nat. 2015, 63, 49–71. [Google Scholar] [CrossRef]
- de Gasperis, S.R.; De Zan, L.R.; Battisti, C.; Reichegger, I.; Carpaneto, G.M. Distribution and abundance of hole-nesting birds in Mediterranean forests: Impact of past management patterns on habitat preference. Ornis Fenn. 2016, 93, 100–110. [Google Scholar] [CrossRef]
- Shaw, T.; Scherer-Lorenzen, M.; Müller, S. Forest structural heterogeneity positively affects bird richness and acoustic diversity in a temperate, central European forest. Front. Ecol. Evol. 2024, 12, 1387879. [Google Scholar] [CrossRef]
- Ciudad, C.; Robles, H.; Matthysen, E. Postfledging habitat selection of juvenile middle spotted woodpeckers: A multi-scale approach. Ecography 2009, 32, 676–682. [Google Scholar] [CrossRef]
- Stański, T.; Stańska, M.; Czeszczewik, D. Foraging behaviour of the Great Spotted Woodpecker (Dendrocopos major) in the Białowieża National Park: Comparison of breeding and non-breeding seasons. Ornis Fenn. 2023, 100, 38–50. [Google Scholar] [CrossRef]
- Kumar, R.; Shahabuddin, G.; Kumar, A. Foraging niche differentiation among sympatric woodpecker species in forests of north-western India. Acta Ornithol. 2020, 55, 88–100. [Google Scholar] [CrossRef]
- Damoc, I.; Sahlean, T.; Ion, R.; Ion, M.; Meşter, L.E. Nesting preferences for two woodpecker species (Dendrocopos major and Dendrocopos medius) in Comana Forest, Southern Romania. Trav. Mus. d’Hist. Nat. Grigore Antipa 2014, 57, 35–45. [Google Scholar] [CrossRef]
- Ónodi, G.; Winkler, D. Nest site characteristics of the Great-spotted Woodpecker in a bottomland riparian forest in the presence of invasive tree species. Ornis Hung. 2016, 24, 81–95. [Google Scholar] [CrossRef]
- Martin, M.; Tremblay, J.A.; Ibarzabal, J.; Morin, H. An indicator species highlights continuous deadwood supply is a key ecological attribute of boreal old-growth forests. Ecosphere 2021, 12, e03507. [Google Scholar] [CrossRef]
- Rolstad, J.; Løken, B.; Rolstad, E. Habitat selection as a hierarchical spatial process: The green woodpecker at the northern edge of its distribution range. Oecologia 2000, 124, 116–129. [Google Scholar] [CrossRef]
- Mattioli, W.; Mancini, L.D.; Portoghesi, L.; Corona, P. Biodiversity conservation and forest management: The case of the sweet chestnut coppice stands in Central Italy. Plant Biosyst.-Int. J. Deal. All Asp. Plant Biol. 2016, 150, 592–600. [Google Scholar] [CrossRef]
- Gondard, H.; Romane, F.; Regina, I.S.; Leonardi, S. Forest management and plant species diversity in chestnut stands of three Mediterranean areas. In Forest Diversity and Management; Springer: Berlin/Heidelberg, Germany, 2006; pp. 69–82. [Google Scholar]
- Robles, H.; Ciudad, C.; Vera, R.; Olea, P.P.; Purroy, F.J.; Matthysen, E. Sylvopastoral management and conservation of the middle spotted woodpecker at the south-western edge of its distribution range. For. Ecol. Manag. 2007, 242, 343–352. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Demertzi, A.; Gerontidou, C.; Gogolos, N.; Bounas, A. Influence of Forest Structural Complexity and Management Intensity on Woodpecker Communities in Mediterranean Chestnut (Castanea sativa) Forests. Ecologies 2025, 6, 37. https://doi.org/10.3390/ecologies6020037
Demertzi A, Gerontidou C, Gogolos N, Bounas A. Influence of Forest Structural Complexity and Management Intensity on Woodpecker Communities in Mediterranean Chestnut (Castanea sativa) Forests. Ecologies. 2025; 6(2):37. https://doi.org/10.3390/ecologies6020037
Chicago/Turabian StyleDemertzi, Alexandra, Christina Gerontidou, Nikolaos Gogolos, and Anastasios Bounas. 2025. "Influence of Forest Structural Complexity and Management Intensity on Woodpecker Communities in Mediterranean Chestnut (Castanea sativa) Forests" Ecologies 6, no. 2: 37. https://doi.org/10.3390/ecologies6020037
APA StyleDemertzi, A., Gerontidou, C., Gogolos, N., & Bounas, A. (2025). Influence of Forest Structural Complexity and Management Intensity on Woodpecker Communities in Mediterranean Chestnut (Castanea sativa) Forests. Ecologies, 6(2), 37. https://doi.org/10.3390/ecologies6020037