
Link Between Umbilical Cord Blood Adipokines and Early Childhood Health



Abstract
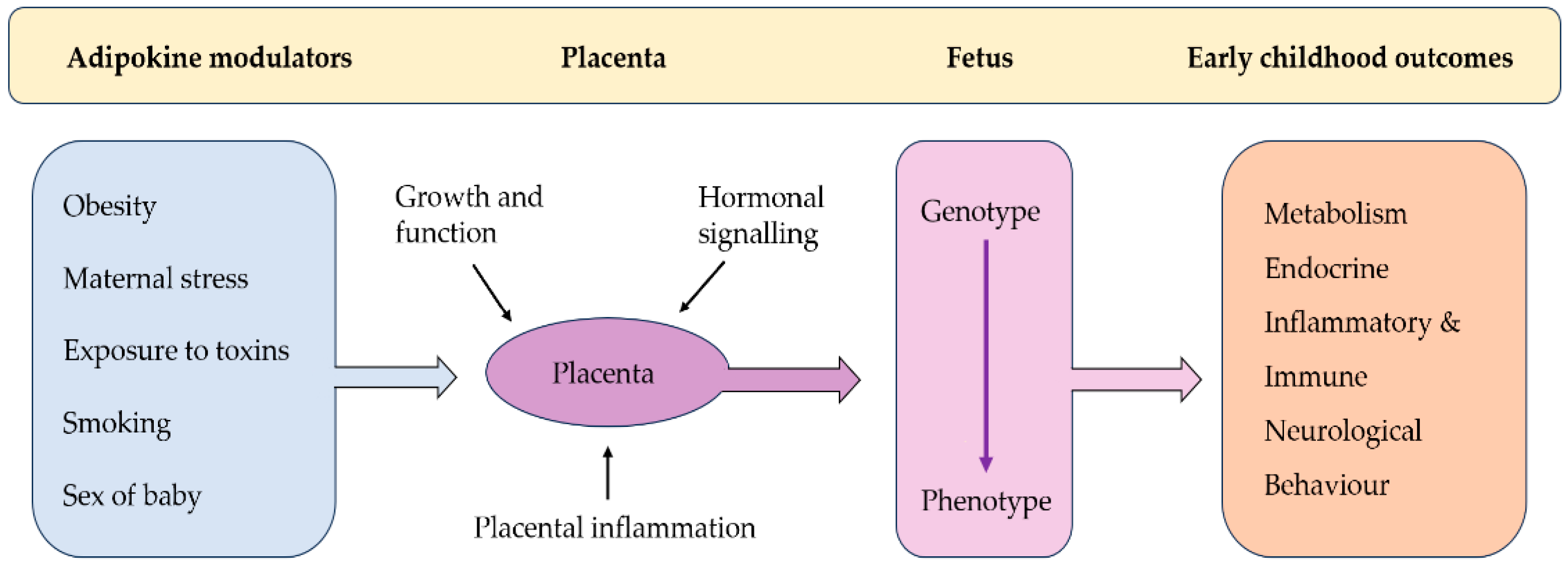
1. Introduction
2. Developmental Origins of Health and Disease
2.1. Role of the Placenta in DoHAD
2.2. Umbilical Cord Blood as a Child Health Indicator
3. Adipokines in Pregnancy
4. Role of Leptin and Adiponectin in Pregnancy
5. Role of Leptin and Adiponectin in Early Childhood Health
5.1. Child Inflammatory Disease
5.2. Early Childhood Neurological Disease
6. Conclusions and Future Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Australian Institute of Health and Welfare. Australia’s Children. 2022. Available online: https://www.aihw.gov.au/reports/children-youth/australias-children (accessed on 12 September 2024).
- Han, V.X.; Patel, S.; Jones, H.F.; Dale, R.C. Maternal immune activation and neuroinflammation in human neurodevelopmental disorders. Nat. Rev. Neurol. 2021, 17, 564–579. [Google Scholar] [CrossRef] [PubMed]
- Meier, U.; Gressner, A.M. Endocrine Regulation of Energy Metabolism: Review of Pathobiochemical and Clinical Chemical Aspects of Leptin, Ghrelin, Adiponectin, and Resistin. Clin. Chem. 2004, 50, 1511–1525. [Google Scholar] [CrossRef]
- Soma-Pillay, P.; Nelson-Piercy, C.; Tolppanen, H.; Mebazaa, A. Physiological changes in pregnancy. Cardiovasc. J. Afr. 2016, 27, 89–94. [Google Scholar] [CrossRef] [PubMed]
- Pardi, G.; Cetin, I. Human fetal growth and organ development: 50 years of discoveries. Am. J. Obstet. Gynecol. 2006, 194, 1088–1099. [Google Scholar] [CrossRef] [PubMed]
- Barker, D.J. The fetal and infant origins of adult disease. Bmj 1990, 301, 1111. [Google Scholar] [CrossRef]
- Rabadán-Diehl, C.; Nathanielsz, P. From Mice to Men: Research models of developmental programming. J. Dev. Orig. Health Dis. 2013, 4, 3–9. [Google Scholar] [CrossRef]
- Mandy, M.; Nyirenda, M. Developmental Origins of Health and Disease: The relevance to developing nations. Int. Health 2018, 10, 66–70. [Google Scholar] [CrossRef]
- Burton, G.J.; Fowden, A.L.; Thornburg, K.L. Placental Origins of Chronic Disease. Physiol. Rev. 2016, 96, 1509–1565. [Google Scholar] [CrossRef]
- Howell, K.R.; Powell, T.L. Effects of maternal obesity on placental function and fetal development. Reproduction 2017, 153, R97–R108. [Google Scholar] [CrossRef]
- Bhattacharya, A.; Freedman, A.N.; Avula, V.; Harris, R.; Liu, W.; Pan, C.; Lusis, A.J.; Joseph, R.M.; Smeester, L.; Hartwell, H.J.; et al. Placental genomics mediates genetic associations with complex health traits and disease. Nat. Commun. 2022, 13, 706. [Google Scholar] [CrossRef]
- Sullivan, E.L.; Molloy, K.R.; Dunn, G.A.; Balanzar, A.L.; Young, A.S.; Loftis, J.M.; Ablow, J.C.; Nigg, J.T.; Gustafsson, H.C. Adipokines measured during pregnancy and at birth are associated with infant negative affect. Brain Behav. Immun. 2024, 120, 34–43. [Google Scholar] [CrossRef] [PubMed]
- D’Ippolito, S.; Tersigni, C.; Scambia, G.; Di Simone, N. Adipokines, an adipose tissue and placental product with biological functions during pregnancy. Biofactors 2012, 38, 14–23. [Google Scholar] [CrossRef] [PubMed]
- Aye, I.L.; Powell, T.L.; Jansson, T. Review: Adiponectin—The missing link between maternal adiposity, placental transport and fetal growth? Placenta 2013, 34, S40–S45. [Google Scholar] [CrossRef]
- Hauguel-de Mouzon, S.; Lepercq, J.; Catalano, P. The known and unknown of leptin in pregnancy. Am. J. Obstet. Gynecol. 2006, 194, 1537–1545. [Google Scholar] [CrossRef]
- Henson, M.C.; Castracane, V.D. Leptin in Pregnancy. Biol. Reprod. 2000, 63, 1219–1228. [Google Scholar] [CrossRef]
- Perez-Perez, A.; Toro, A.; Vilarino-Garcia, T.; Maymo, J.; Guadix, P.; Duenas, J.L.; Fernandez-Sanchez, M.; Varone, C.; Sanchez-Margalet, V. Leptin action in normal and pathological pregnancies. J. Cell. Mol. Med. 2018, 22, 716–727. [Google Scholar] [CrossRef]
- Briffa, J.F.; McAinch, A.J.; Romano, T.; Wlodek, M.E.; Hryciw, D.H. Leptin in pregnancy and development: A contributor to adulthood disease? Am. J. Physiol. Endocrinol. Metab. 2015, 308, E335–E350. [Google Scholar] [CrossRef] [PubMed]
- Friedman, J.M. Leptin and the endocrine control of energy balance. Nat. Metab. 2019, 1, 754–764. [Google Scholar] [CrossRef]
- Challier, J.; Galtier, M.; Bintein, T.; Cortez, A.; Lepercq, J.; Hauguel-de Mouzon, S. Placental Leptin Receptor Isoforms in Normal and Pathological Pregnancies. Placenta 2003, 24, 92–99. [Google Scholar] [CrossRef]
- Schubring, C.; Englaro, P.; Siebler, T.; Blum, W.F.; Demirakca, T.; Kratzsch, J.; Kiess, W. Longitudinal Analysis of Maternal Serum Leptin Levels during Pregnancy, at Birth and Up To Six Weeks after Birth: Relation to Body Mass Index, Skinfolds, Sex Steroids and Umbilical Cord Blood Leptin Levels. Horm. Res. 1999, 50, 276–283. [Google Scholar] [CrossRef]
- Casado, M.E.; Collado-Pérez, R.; Frago, L.M.; Barrios, V. Recent Advances in the Knowledge of the Mechanisms of Leptin Physiology and Actions in Neurological and Metabolic Pathologies. Int. J. Mol. Sci. 2023, 24, 1422. [Google Scholar] [CrossRef] [PubMed]
- Emilsson, V.; Liu, Y.L.; Cawthorne, M.A.; Morton, N.M.; Davenport, M. Expression of the functional leptin receptor mRNA in pancreatic islets and direct inhibitory action of leptin on insulin secretion. Diabetes 1997, 46, 313–316. [Google Scholar] [CrossRef]
- Hoggard, N.; Hunter, L.; Duncan, J.S.; Williams, L.M.; Trayhurn, P.; Mercer, J.G. Leptin and leptin receptor mRNA and protein expression in the murine fetus and placenta. Proc. Natl. Acad. Sci. USA 1997, 94, 11073–11078. [Google Scholar] [CrossRef]
- Attig, L.; Larcher, T.; Gertler, A.; Abdennebi-Najar, L.; Djiane, J. Postnatal leptin is necessary for maturation of numerous organs in newborn rats. Organogenesis 2011, 7, 88–94. [Google Scholar] [CrossRef]
- Caminos, J.E.; Nogueiras, R.; Gallego, R.; Bravo, S.; Tovar, S.; García-Caballero, T.; Casanueva, F.F.; Diéguez, C. Expression and regulation of adiponectin and receptor in human and rat placenta. J. Clin. Endocrinol. Metab. 2005, 90, 4276–4286. [Google Scholar] [CrossRef]
- Chen, J.; Tan, B.; Karteris, E.; Zervou, S.; Digby, J.; Hillhouse, E.W.; Vatish, M.; Randeva, H.S. Secretion of adiponectin by human placenta: Differential modulation of adiponectin and its receptors by cytokines. Diabetologia 2006, 49, 1292–1302. [Google Scholar] [CrossRef] [PubMed]
- Moyce Gruber, B.L.; Dolinsky, V.W. The Role of Adiponectin during Pregnancy and Gestational Diabetes. Life 2023, 13, 301. [Google Scholar] [CrossRef] [PubMed]
- McDonald, E.A.; Wolfe, M.W. Adiponectin Attenuation of Endocrine Function within Human Term Trophoblast Cells. Endocrinology 2009, 150, 4358–4365. [Google Scholar] [CrossRef]
- Qiao, L.; Yoo H s Madon, A.; Kinney, B.; Hay, W.W., Jr.; Shao, J. Adiponectin Enhances Mouse Fetal Fat Deposition. Diabetes 2012, 61, 3199–3207. [Google Scholar] [CrossRef]
- Clemente-Suárez, V.J.; Redondo-Flórez, L.; Beltrán-Velasco, A.I.; Martín-Rodríguez, A.; Martínez-Guardado, I.; Navarro-Jiménez, E.; Laborde-Cárdenas, C.C.; Tornero-Aguilera, J.F. The Role of Adipokines in Health and Disease. Biomedicines 2023, 11, 1290. [Google Scholar] [CrossRef]
- Kelly, A.C.; Powell, T.L.; Jansson, T. Placental function in maternal obesity. Clin. Sci. 2020, 134, 961–984. [Google Scholar] [CrossRef] [PubMed]
- Parrettini, S.; Caroli, A.; Torlone, E. Nutrition and Metabolic Adaptations in Physiological and Complicated Pregnancy: Focus on Obesity and Gestational Diabetes. Front. Endocrinol. 2020, 11, 611929. [Google Scholar] [CrossRef]
- Christian, L.M.; Porter, K. Longitudinal changes in serum proinflammatory markers across pregnancy and postpartum: Effects of maternal body mass index. Cytokine 2014, 70, 134–140. [Google Scholar] [CrossRef] [PubMed]
- Sferruzzi-Perri, A.N.; Vaughan, O.R.; Haro, M.; Cooper, W.N.; Musial, B.; Charalambous, M.; Pestana, D.; Ayyar, S.; Ferguson-Smith, A.C.; Burton, G.J.; et al. An obesogenic diet during mouse pregnancy modifies maternal nutrient partitioning and the fetal growth trajectory. Faseb J. 2013, 27, 3928–3937. [Google Scholar] [CrossRef]
- Rosario, F.J.; Schumacher, M.A.; Jiang, J.; Kanai, Y.; Powell, T.L.; Jansson, T. Chronic maternal infusion of full-length adiponectin in pregnant mice down-regulates placental amino acid transporter activity and expression and decreases fetal growth. J. Physiol. 2012, 590, 1495–1509. [Google Scholar] [CrossRef] [PubMed]
- Jansson, T.; Aye, I.L.; Goberdhan, D.C. The emerging role of mTORC1 signaling in placental nutrient-sensing. Placenta 2012, 33, e23–e29. [Google Scholar] [CrossRef]
- Napso, T.; Yong, H.E.; Lopez-Tello, J.; Sferruzzi-Perri, A.N. The role of placental hormones in mediating maternal adaptations to support pregnancy and lactation. Front. Physiol. 2018, 9, 1091. [Google Scholar] [CrossRef]
- Aye, I.L.; Rosario, F.J.; Powell, T.L.; Jansson, T. Adiponectin supplementation in pregnant mice prevents the adverse effects of maternal obesity on placental function and fetal growth. Proc. Natl. Acad. Sci. USA 2015, 112, 12858–12863. [Google Scholar] [CrossRef]
- Nguyen, T.M.D. Adiponectin: Role in Physiology and Pathophysiology. Int. J. Prev. Med. 2020, 11, 136. [Google Scholar] [CrossRef]
- Bayol, S.A.; Simbi, B.H.; Bertrand, J.; Stickland, N.C. Offspring from mothers fed a ‘junk food’ diet in pregnancy and lactation exhibit exacerbated adiposity that is more pronounced in females. J. Physiol. 2008, 586, 3219–3230. [Google Scholar] [CrossRef]
- Kotani, Y.; Yokota, I.; Kitamura, S.; Matsuda, J.; Naito, E.; Kuroda, Y. Plasma adiponectin levels in newborns are higher than those in adults and positively correlated with birth weight. Clin. Endocrinol. 2004, 61, 418–423. [Google Scholar] [CrossRef] [PubMed]
- Rothenbacher, D.; Weyermann, M.; Fantuzzi, G.; Brenner, H. Adipokines in cord blood and risk of wheezing disorders within the first two years of life. Clin. Exp. Allergy 2007, 37, 1143–1149. [Google Scholar] [CrossRef]
- Kajantie, E.; Hytinantti, T.; Hovi, P.; Andersson, S. Cord plasma adiponectin: A 20-fold rise between 24 weeks gestation and term. J. Clin. Endocrinol. Metab. 2004, 89, 4031–4036. [Google Scholar] [CrossRef] [PubMed]
- Oken, E.; Morton-Eggleston, E.; Rifas-Shiman, S.L.; Switkowski, K.M.; Hivert, M.F.; Fleisch, A.F.; Mantzoros, C.; Gillman, M.W. Sex-Specific Associations of Maternal Gestational Glycemia with Hormones in Umbilical Cord Blood at Delivery. Am. J. Perinatol. 2016, 33, 1273–1281. [Google Scholar] [CrossRef] [PubMed]
- Ashley-Martin, J.; Karaceper, M.; Dodds, L.; Arbuckle, T.E.; Ettinger, A.S.; Fraser, W.D.; Muckle, G.; Monnier, P.; Fisher, M.; Kuhle, S. An examination of sex differences in associations between cord blood adipokines and childhood adiposity. Pediatr. Obes. 2020, 15, e12587. [Google Scholar] [CrossRef]
- Bjune, J.I.; Stromland, P.P.; Jersin, R.A.; Mellgren, G.; Dankel, S.N. Metabolic and Epigenetic Regulation by Estrogen in Adipocytes. Front. Endocrinol. 2022, 13, 828780. [Google Scholar] [CrossRef]
- Dearden, L.; Bouret, S.G.; Ozanne, S.E. Sex and gender differences in developmental programming of metabolism. Mol. Metab. 2018, 15, 8–19. [Google Scholar] [CrossRef]
- Howerton, C.L.; Bale, T.L. Targeted placental deletion of OGT recapitulates the prenatal stress phenotype including hypothalamic mitochondrial dysfunction. Proc. Natl. Acad. Sci. USA 2014, 111, 9639–9644. [Google Scholar] [CrossRef]
- Clegg, D.J.; Riedy, C.A.; Smith, K.A.; Benoit, S.C.; Woods, S.C. Differential sensitivity to central leptin and insulin in male and female rats. Diabetes 2003, 52, 682–687. [Google Scholar] [CrossRef]
- Laml, T.; Hartmann, B.W.; Preyer, O.; Ruecklinger, E.; Soeregi, G.; Wagenbichler, P. Serum leptin concentration in cord blood: Relationship to birth weight and gender in pregnancies complicated by pre-eclampsia. Gynecol. Endocrinol. 2000, 14, 442–447. [Google Scholar] [CrossRef]
- Tome, F.M.; Chateau, D.; Helbling-Leclerc, A.; Fardeau, M. Morphological changes in muscle fibers in oculopharyngeal muscular dystrophy. Neuromuscul. Disord. 1997, 7 (Suppl. 1), S63–S69. [Google Scholar] [CrossRef]
- Valleau, J.C.; Sullivan, E.L. The impact of leptin on perinatal development and psychopathology. J. Chem. Neuroanat. 2014, 61–62, 221–232. [Google Scholar] [CrossRef] [PubMed]
- Tilg, H.; Ianiro, G.; Gasbarrini, A.; Adolph, T.E. Adipokines: Masterminds of metabolic inflammation. Nat. Rev. Immunol. 2024. [Google Scholar] [CrossRef] [PubMed]
- Agrawal, S.; Gollapudi, S.; Su, H.; Gupta, S. Leptin Activates Human B Cells to Secrete TNF-α, IL-6, and IL-10 via JAK2/STAT3 and p38MAPK/ERK1/2 Signaling Pathway. J. Clin. Immunol. 2011, 31, 472–478. [Google Scholar] [CrossRef] [PubMed]
- Aleffi, S.; Petrai, I.; Bertolani, C.; Parola, M.; Colombatto, S.; Novo, E.; Vizzutti, F.; Anania, F.A.; Milani, S.; Rombouts, K. Upregulation of proinflammatory and proangiogenic cytokines by leptin in human hepatic stellate cells. Hepatology 2005, 42, 1339–1348. [Google Scholar] [CrossRef]
- Denizli, M.; Capitano, M.L.; Kua, K.L. Maternal obesity and the impact of associated early-life inflammation on long-term health of offspring. Front. Cell. Infect. Microbiol. 2022, 12, 940937. [Google Scholar] [CrossRef]
- Feijóo-Bandín, S.; Aragón-Herrera, A.; Moraña-Fernández, S.; Anido-Varela, L.; Tarazón, E.; Roselló-Lletí, E.; Portolés, M.; Moscoso, I.; Gualillo, O.; González-Juanatey, J.R.; et al. Adipokines and Inflammation: Focus on Cardiovascular Diseases. Int. J. Mol. Sci. 2020, 21, 7711. [Google Scholar] [CrossRef]
- Abu-Raya, B.; Michalski, C.; Sadarangani, M.; Lavoie, P.M. Maternal Immunological Adaptation During Normal Pregnancy. Front. Immunol. 2020, 11, 575197. [Google Scholar] [CrossRef]
- Han, V.X.; Patel, S.; Jones, H.F.; Nielsen, T.C.; Mohammad, S.S.; Hofer, M.J.; Gold, W.; Brilot, F.; Lain, S.J.; Nassar, N.; et al. Maternal acute and chronic inflammation in pregnancy is associated with common neurodevelopmental disorders: A systematic review. Transl. Psychiatry 2021, 11, 71. [Google Scholar] [CrossRef]
- Mancuso, P. The role of adipokines in chronic inflammation. Immunotargets Ther. 2016, 5, 47–56. [Google Scholar] [CrossRef]
- Sabic, D.; Koenig, J.M. A perfect storm: Fetal inflammation and the developing immune system. Pediatr. Res. 2020, 87, 319–326. [Google Scholar] [CrossRef] [PubMed]
- Kundakovic, M.; Jaric, I. The Epigenetic Link between Prenatal Adverse Environments and Neurodevelopmental Disorders. Genes 2017, 8, 104. [Google Scholar] [CrossRef] [PubMed]
- Park, S.; Vargas, Z.; Zhao, A.; Baltazar, P.I.; Friedman, J.F.; McDonald, E.A. Cord blood adiponectin and leptin are associated with a lower risk of stunting during infancy. Sci. Rep. 2022, 12, 15122. [Google Scholar] [CrossRef]
- Lee, S.; Lee, J.; Kang, G.M.; Kim, M.S. Leptin directly regulate intrinsic neuronal excitability in hypothalamic POMC neurons but not in AgRP neurons in food restricted mice. Neurosci. Lett. 2018, 681, 105–109. [Google Scholar] [CrossRef]
- Smith, P.M.; Brzezinska, P.; Hubert, F.; Mimee, A.; Maurice, D.H.; Ferguson, A.V. Leptin influences the excitability of area postrema neurons. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2016, 310, R440–R448. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Arbuckle, T.E.; Muckle, G.; Lanphear, B.P.; Boivin, M.; Chen, A.; Dodds, L.; Fraser, W.D.; Ouellet, E.; Seguin, J.R.; et al. Associations of cord blood leptin and adiponectin with children’s cognitive abilities. Psychoneuroendocrinology 2019, 99, 257–264. [Google Scholar] [CrossRef]
- Farr, S.A.; Banks, W.A.; Morley, J.E. Effects of leptin on memory processing. Peptides 2006, 27, 1420–1425. [Google Scholar] [CrossRef]
- Oomura, Y.; Hori, N.; Shiraishi, T.; Fukunaga, K.; Takeda, H.; Tsuji, M.; Matsumiya, T.; Ishibashi, M.; Aou, S.; Li, X.L.; et al. Leptin facilitates learning and memory performance and enhances hippocampal CA1 long-term potentiation and CaMK II phosphorylation in rats. Peptides 2006, 27, 2738–2749. [Google Scholar] [CrossRef]
- Fadel, J.R.; Jolivalt, C.G.; Reagan, L.P. Food for thought: The role of appetitive peptides in age-related cognitive decline. Ageing Res. Rev. 2013, 12, 764–776. [Google Scholar] [CrossRef]
- Morrison, C.D. Leptin signaling in brain: A link between nutrition and cognition? Biochim. Biophys. Acta 2009, 1792, 401–408. [Google Scholar] [CrossRef]
- Jeon, B.T.; Shin, H.J.; Kim, J.B.; Kim, Y.K.; Lee, D.H.; Kim, K.H.; Kim, H.J.; Kang, S.S.; Cho, G.J.; Choi, W.S.; et al. Adiponectin protects hippocampal neurons against kainic acid-induced excitotoxicity. Brain Res. Rev. 2009, 61, 81–88. [Google Scholar] [CrossRef] [PubMed]
- Nishimura, M.; Izumiya, Y.; Higuchi, A.; Shibata, R.; Qiu, J.; Kudo, C.; Shin, H.K.; Moskowitz, M.A.; Ouchi, N. Adiponectin prevents cerebral ischemic injury through endothelial nitric oxide synthase dependent mechanisms. Circulation 2008, 117, 216–223. [Google Scholar] [CrossRef] [PubMed]
- Arnoldussen, I.A.; Kiliaan, A.J.; Gustafson, D.R. Obesity and dementia: Adipokines interact with the brain. Eur. Neuropsychopharmacol. 2014, 24, 1982–1999. [Google Scholar] [CrossRef]
- Kasper, J.M.; Milton, A.J.; Smith, A.E.; Laezza, F.; Taglialatela, G.; Hommel, J.D.; Abate, N. Cognitive deficits associated with a high-fat diet and insulin resistance are potentiated by overexpression of ecto-nucleotide pyrophosphatase phosphodiesterase-1. Int. J. Dev. Neurosci. 2018, 64, 48–53. [Google Scholar] [CrossRef]
- Merabova, N.; Ugartemendia, L.; Edlow, A.G.; Ibarra, C.; Darbinian, N.; Tatevosian, G.; Goetzl, L. Maternal obesity: Sex-specific in utero changes in fetal brain autophagy and mTOR. Obesity 2024, 32, 1136–1143. [Google Scholar] [CrossRef]
- Lindberger, E.; Larsson, A.; Kunovac Kallak, T.; Sundstrom Poromaa, I.; Wikstrom, A.K.; Osterroos, A.; Ahlsson, F. Maternal early mid-pregnancy adiponectin in relation to infant birth weight and the likelihood of being born large-for-gestational-age. Sci. Rep. 2023, 13, 20919. [Google Scholar] [CrossRef]
- Castro-Rodriguez, J.A.; Forno, E.; Casanello, P.; Padilla, O.; Krause, B.J.; Uauy, R. Leptin in Cord Blood Associates with Asthma Risk at Age 3 in the Offspring of Women with Gestational Obesity. Ann. Am. Thorac. Soc. 2020, 17, 1583–1589. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Xiao, X.; Song, X.; Qi, Z.; Li, Y. Prediction of cord blood leptin on infant’s neurodevelopment: A birth cohort in rural Yunnan, China. Psychoneuroendocrinology 2023, 148, 105955. [Google Scholar] [CrossRef]
- Beltz, A.M.; Beery, A.K.; Becker, J.B. Analysis of sex differences in pre-clinical and clinical data sets. Neuropsychopharmacology 2019, 44, 2155–2158. [Google Scholar] [CrossRef]

{kind=link}
| Adipokine ↑ or ↓ (Adipokine Measurement) n Infant Age M/F Numbers % Maternal Overweight/Obese | Outcome (Assessment) | Sex Effect | Reference |
|---|---|---|---|
| Leptin ↑ (ELISA) n = 339 3 years NR Overweight: 23.59%/obese: 58.70% | ↑ 30% asthma risk being born from obese mothers (Asthma Prediction Index (API)) | NR | [78] |
| Leptin ↑ (ELISA) n = 740 2 years 383 M, 354 F Overweight: 20.7%/obese: 8.1% | ↔ wheezing disorder (physician-reported) | F > M | [43] |
| Leptin ↑ (ELISA) N = 612 3, 5, 8 years NR Overweight: 21.7%/obese: 18.4% | ↔ IQ (Wechsler Preschool and Primary Scales of Intelligence-III or Wechsler Intelligence Scale for Children-IV) | NR | [67] |
| Leptin ↓ (ELISA) n = 323 1 year NR Overweight OR obese: 20.1% | ↓ neurodevelopment score (Bayley Scales of Infant and Toddler Development, Third Edition (BSID-III)) | NR | [79] |
| Adiponectin ↑ (ELISA) n = 740 2 years 385M, 354 F Overweight: 20.7%/obese: 8.1% | ↑ asthma or obstructive bronchitis (physician-reported) | NR | [43] |
| Leptin ↑ (ELISA) n = 341 12 months 171M, 170F Overweight: 20.1% | ↔ neurodevelopmental outcomes (Bayley Scales of Infant and Toddler Development Test, third edition (BSID-III) | NR | [64] |
| Adiponectin ↑ (ELISA) n = 341 12 months 171M, 170F Overweight: 20.1% | ↓ fine motor skills (Bayley Scales of Infant and Toddler Development Test, third edition (BSID-III) | NR | [64] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Boyle, A.M.; Holland, O.J.; Hryciw, D.H. Link Between Umbilical Cord Blood Adipokines and Early Childhood Health. Endocrines 2025, 6, 13. https://doi.org/10.3390/endocrines6010013
Boyle AM, Holland OJ, Hryciw DH. Link Between Umbilical Cord Blood Adipokines and Early Childhood Health. Endocrines. 2025; 6(1):13. https://doi.org/10.3390/endocrines6010013
Chicago/Turabian StyleBoyle, Ava M., Olivia J. Holland, and Deanne H. Hryciw. 2025. "Link Between Umbilical Cord Blood Adipokines and Early Childhood Health" Endocrines 6, no. 1: 13. https://doi.org/10.3390/endocrines6010013
APA StyleBoyle, A. M., Holland, O. J., & Hryciw, D. H. (2025). Link Between Umbilical Cord Blood Adipokines and Early Childhood Health. Endocrines, 6(1), 13. https://doi.org/10.3390/endocrines6010013