
A Critical Review on In Vitro and Ex Vivo Models of the Intestinal Epithelium of Humans and Monogastric Animals


 ,
,  and
and
Abstract
1. Introduction
2. Materials and Methods
Exclusion and Inclusion Criteria
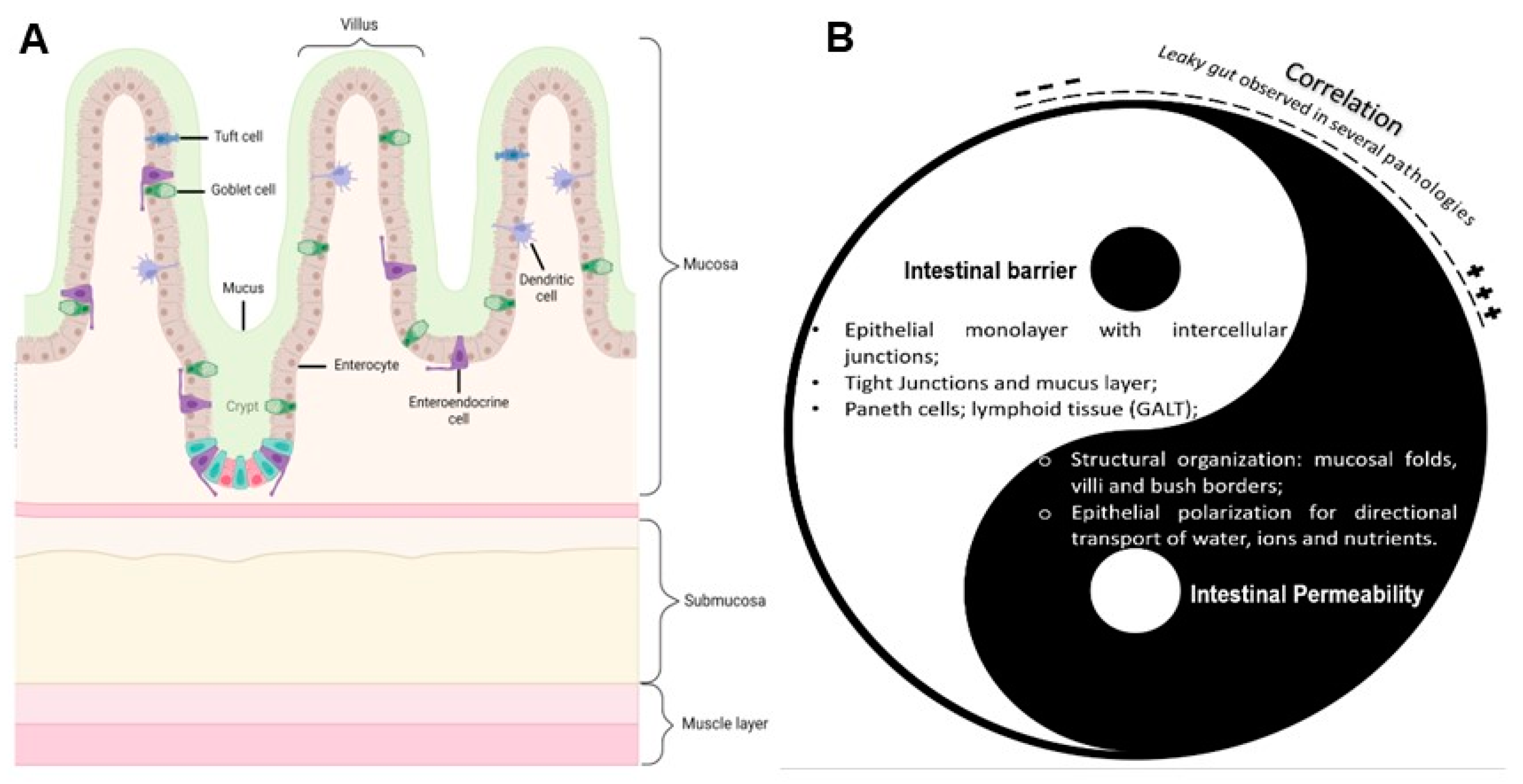
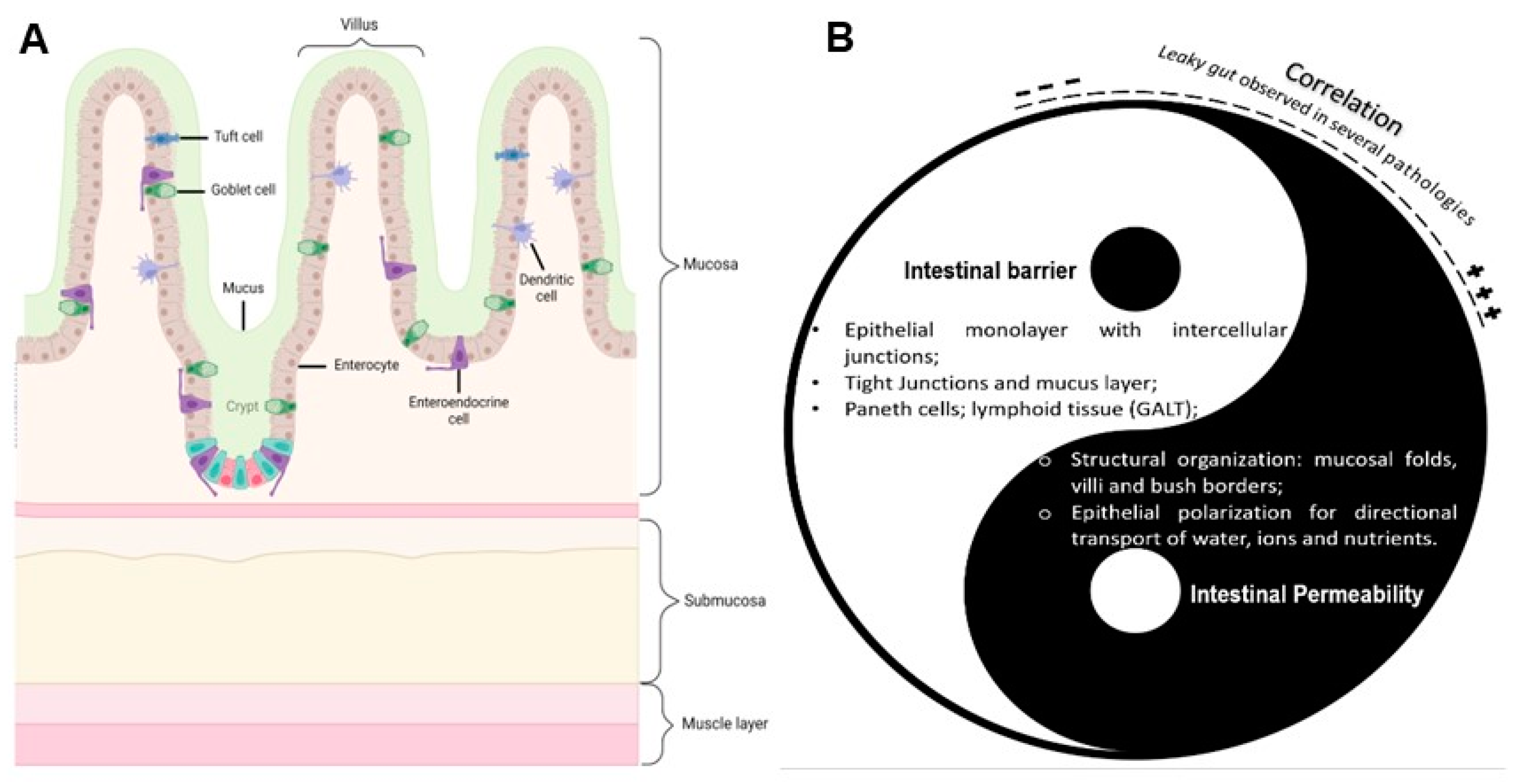
3. Overview of the Absorptive Epithelium—Structure and Physiology
3.1. Human Intestinal Epithelium
3.2. Pig and Chicken Intestinal Epitheliums
3.3. Nutrients’ Transportation Routes across Epithelium
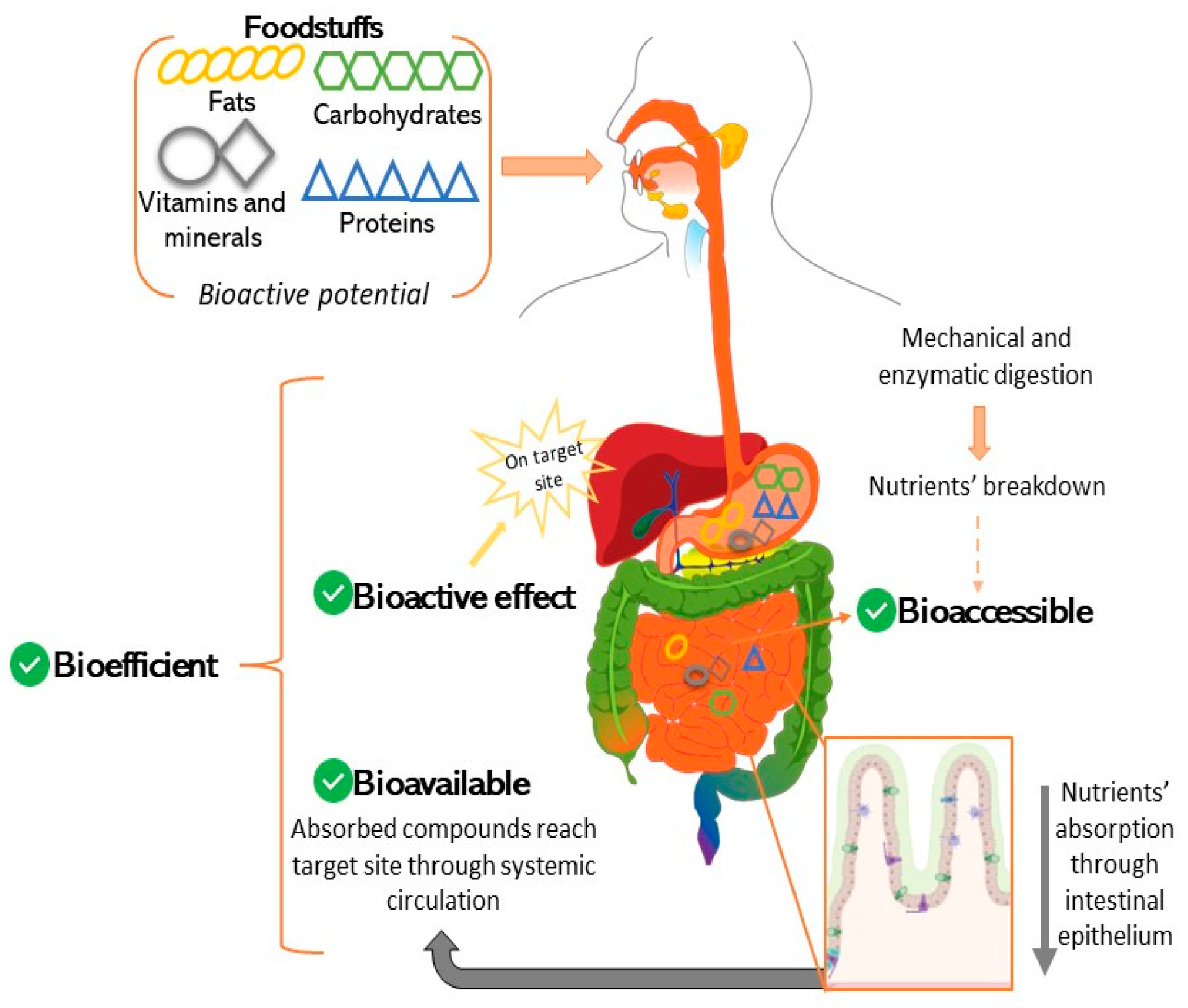
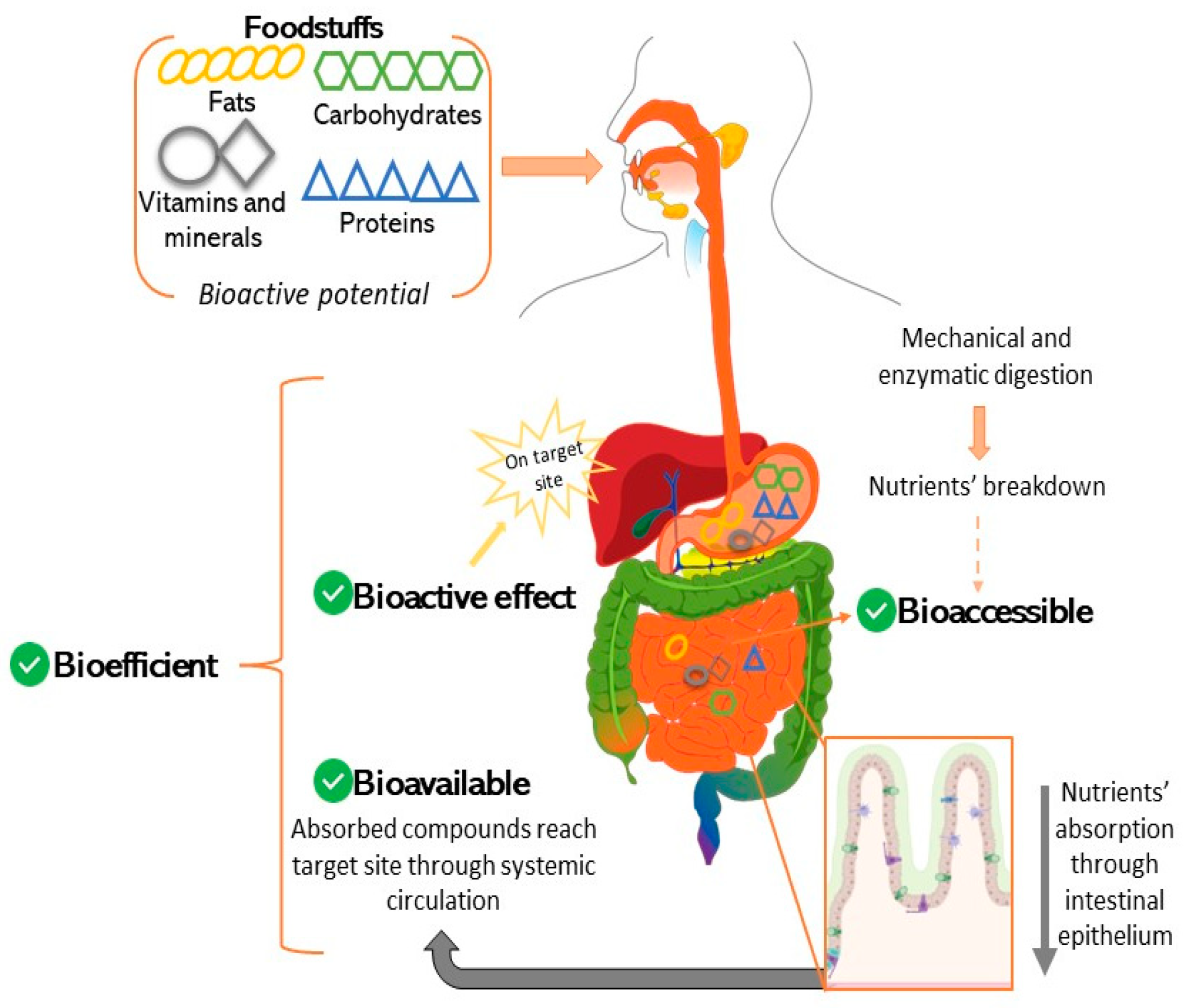
4. In Vitro Models for Predicting Bioaccessibility
5. Intestinal Permeability Models and Applications
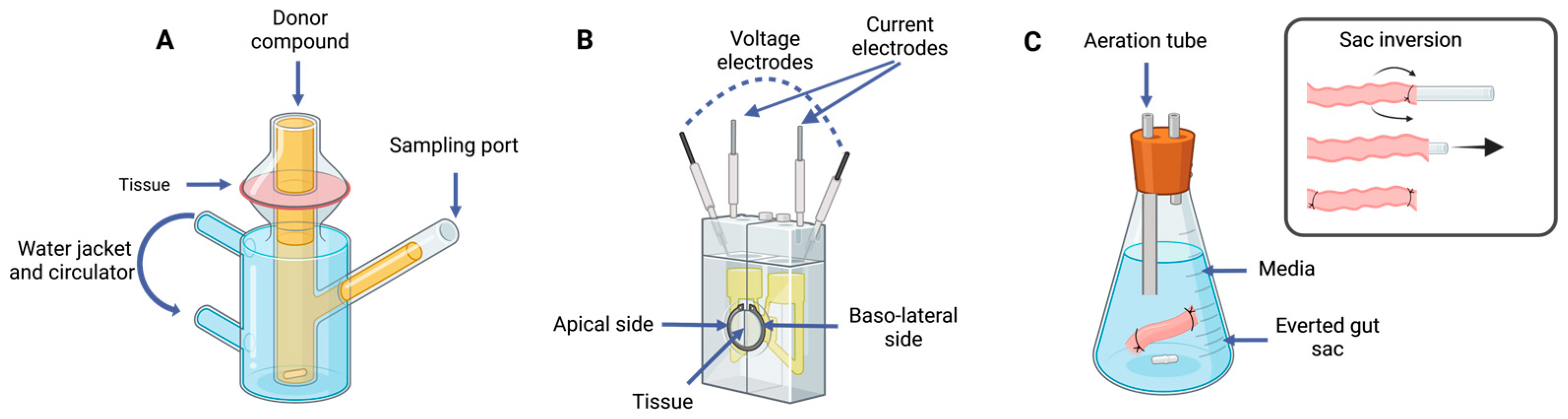
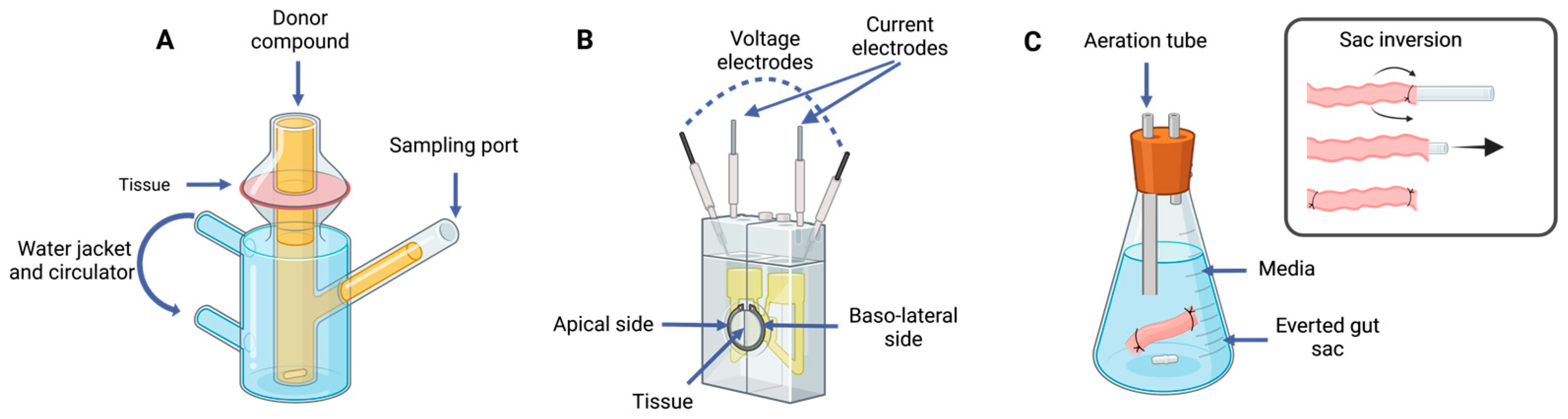
5.1. Tissue-Based Methods
5.1.1. Ussing Chamber
5.1.2. Franz Diffusion Cells
5.1.3. Intestinal Rings and Segments
5.1.4. Everted Gut Sac
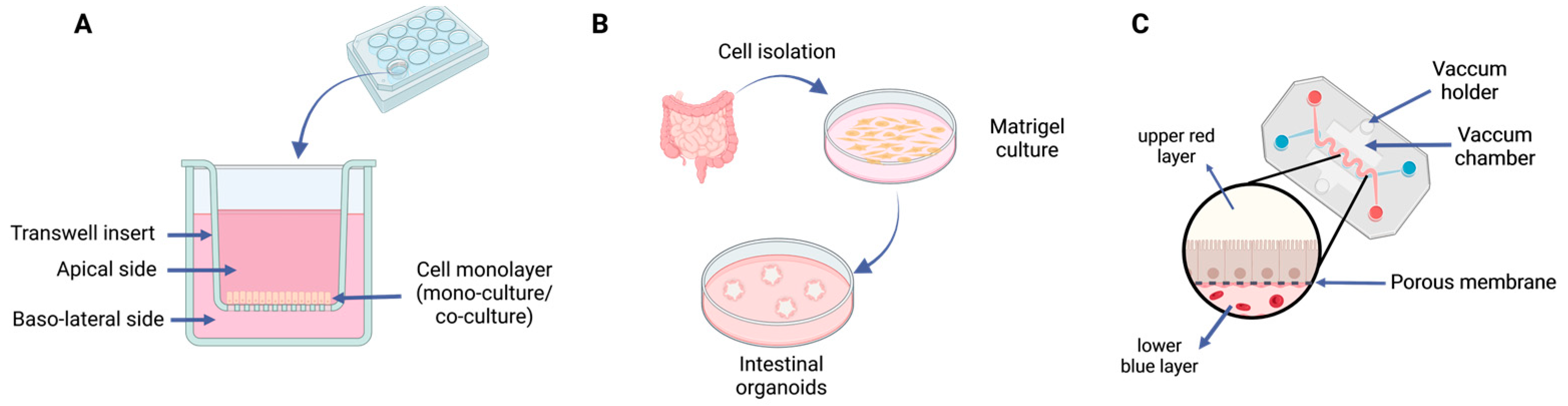
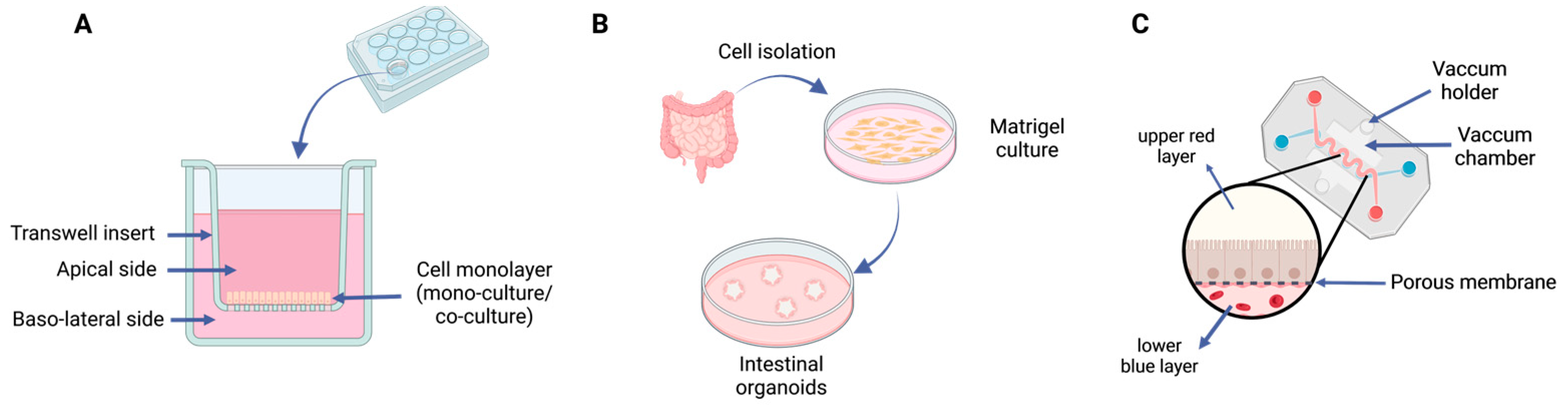
5.2. Cell-Based Methods
5.3. Nonbiological Methods

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Technique | Food/Ingredient Tested | Model | Description | Reference |
|---|---|---|---|---|
| Ussing chamber | Cinnamon bark oil and coconut oil emulsions | Laying hen | No previous simulation of the gastrointestinal tract | [103] |
| Apple polyphenols | Pig | Direct application of polyphenols to a Ussing chamber; no previous digestion of the polyphenols was performed | [104] | |
| Organic acid supplementation in feed | Chicken | In vivo administration of the supplement was performed; a Ussing chamber was used to assess intestinal permeability changes | [105] | |
| Oligopeptides from whey protein hydrolysate | Pig | In vitro digestion and absorption simulation through Ussing chamber; | [106] | |
| Franz cells | Tetrahydrocurcumin-hyaluronic acid conjugate (metabolite of curcumin) | Pig | TNO dynamic gastrointestinal model-1 (TIM-1) followed by Franz cell assay | [107] |
| Monosaccharides, amino acids and a corn oil-in-water emulsion | Semi-permeable cellulose membrane | No gastrointestinal simulation was performed; no biological tissue was used | [108] | |
| Everted intestinal sac | Fructose uptake | Chicken | No previous digestive process was simulated | [109] |
| Garra fish meal | Chicken | In vivo studies carried out in chickens, followed by an evaluation of diet effects on intestinal permeability | [110] | |
| Encapsulation of β-carotene in zein protein | Chicken | Study focused on human health; human GIT digestion simulation was simulated, followed by absorption experiments with chicken intestines | [111] | |
| Phenolic compounds from non-extruded and extruded Mango Bagasse-added confections | Pig | Human in vitro GIT digestion, followed by permeability assessment | [112] | |
| Encapsulated curcumin and resveratrol | Pig | Human in vitro GIT digestion was carried out, followed by an everted gut sac for BA assessment | [113] | |
| Mono-cultures | Angiotensin I-converting enzyme inhibitory peptides, from cooked chicken breast/thighs | Caco-2 | Peptides identified after in vitro digestion, followed by PET inserts with a Caco-2 monoculture | [114] |
| Curcumin alone or with polyvinylpyrrolidone | Caco-2 | This study focused on chickens’ health; despite an in vitro simulation of the chicken GIT, a Transwell permeability assay was performed with the initial samples, not the digested ones | [115] | |
| Co-cultures | Sardine protein hydrolysate | Caco-2 + HT29-MTX | After human in vitro digestion, permeability was assessed using PET inserts (like the TranswellTM system) | [116] |
| Encapsulated rosemary extract | Caco-2 + HT29-MTX | In vitro human digestion followed by co-culture in a TranswellTM system | [117] | |
| Salmosan (derived from Mannan oligosaccharide) | Caco-2 + THP-1 (macrophages) | Salmosan and Salmosan with L. plantarum were tested for the effects on the intestinal permeability and barrier, as potential feed additives; nevertheless, no GIT digestion was simulated | [118] | |
| Dialysis membrane | Gelatinized starch dispersions | Hollow fiber membrane (synthetic) | Study on starch digestion and the consequent absorption of hydrolytic products generated in the human small intestine, using an in vitro intestinal digestion system (i-IDS) | [95] |
| Phenolics, flavonoids, rutin, β-carotene and lutein in six edible greens | Cellulose membrane (12,000 Da) | Bioaccessibility and BA was evaluated; GIT simulation was carried out followed by dialysis, as a simplified model of intestinal permeation | [119] | |
| PAMPA | Crude plant extracts (Angelica archangelica, Waltheria indica, Pueraria montana var. lobata) | Polycarbonate filter plate (5–20% porosity with a 0.45 µm pore size and 9–10 µm thickness) impregnated with a hexadecane/hexane (5/95 % (v/v)) solution | Prediction of the passive intestinal absorption of a representative set of frequently occurring natural products from Angelica archangelica, Waltheria indica and Pueraria montana var. lobata; no GIT digestion simulation was performed before PAMPA assay | [120] |
| Saponins and sapogenins from seed extracts from red quinoa and seeds of fenugreek. | Lipid mixture containing L-α-phosphatidylcholine and cholesterol in 1,7-octadiene solution added to the PVDF filter of each well; after membrane coating, the donor solutions were added | Evaluation and comparison of the permeability of saponins and sapogenins from fenugreek and quinoa extracts with and without previous in vitro digestion simulation, through the previous development of a GIT digestion protocol attached to PAMPA | [121] |
6. Conclusions and Opportunities
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Ozen, A.E.; Pons, A.; Tur, J.A. Worldwide Consumption of Functional Foods: A Systematic Review. Nutr. Rev. 2012, 70, 472–481. [Google Scholar] [CrossRef]
- Pandey, A.K.; Kumar, P.; Saxena, M.J. Feed Additives in Animal Health. In Nutraceuticals in Veterinary Medicine; Springer: Berlin/Heidelberg, Germany, 2019; pp. 345–362. [Google Scholar] [CrossRef]
- Dima, C.; Assadpour, E.; Dima, S.; Jafari, S.M. Bioavailability and Bioaccessibility of Food Bioactive Compounds; Overview and Assessment by in vitro Methods. Compr. Rev. Food Sci. Food Saf. 2020, 19, 2862–2884. [Google Scholar] [CrossRef]
- Santos, D.I.; Saraiva, J.M.A.; Vicente, A.A.; Moldão-Martins, M. Innovative Thermal and Non-Thermal Processing, Bioaccessibility and Bioavailability of Nutrients and Bioactive Compounds; Woodhead Publishing: Sawston, UK, 2019; Chapter 2; pp. 23–54. [Google Scholar]
- Sensoy, I. A Review on the Relationship Between Food Structure, Processing, and Bioavailability. Crit. Rev. Food Sci. Nutr. 2014, 54, 902–909. [Google Scholar] [CrossRef]
- Brodkorb, A.; Egger, L.; Alminger, M.; Alvito, P.; Assunção, R.; Ballance, S.; Bohn, T.; Bourlieu-Lacanal, C.; Boutrou, R.; Carrière, F.; et al. INFOGEST Static in vitro Simulation of Gastrointestinal Food Digestion. Nat. Protoc. 2019, 14, 991–1014. [Google Scholar] [CrossRef]
- Rodrigues, D.B.; Chitchumroonchokchai, C.; Mariutti, L.R.B.; Mercadante, A.Z.; Failla, M.L. Comparison of Two Static in vitro Digestion Methods for Screening the Bioaccessibility of Carotenoids in Fruits, Vegetables, and Animal Products. J. Agric. Food Chem. 2017, 65, 11220–11228. [Google Scholar] [CrossRef]
- Bryan, D.D.S.L.; Abbott, D.A.; Classen, H.L. Development of an in vitro Protein Digestibility Assay Mimicking the Chicken Digestive Tract. Anim. Nutr. 2018, 4, 401–409. [Google Scholar] [CrossRef]
- Bean, T.G.; Arnold, K.E.; Lane, J.; Pietravalle, S.; Boxall, A.B.A. An in vitro Method for Determining the Bioaccessibility of Pharmaceuticals in Wildlife. Environ. Toxicol. Chem. 2016, 35, 2349–2357. [Google Scholar] [CrossRef]
- Ng, J.C.; Juhasz, A.; Smith, E.; Naidu, R. Assessing the Bioavailability and Bioaccessibility of Metals and Metalloids. Environ. Sci. Pollut. Res. 2015, 22, 8802–8825. [Google Scholar] [CrossRef] [PubMed]
- Ting, Y.; Zhao, Q.; Xia, C.; Huang, Q. Using in vitro and in vivo Models To Evaluate the Oral Bioavailability of Nutraceuticals. J. Agric. Food Chem. 2015, 63, 1332–1338. [Google Scholar] [CrossRef] [PubMed]
- Verhoeckx, K.; Cotter, P.; López-Expósito, I.; Kleiveland, C.; Lea, T.; Mackie, A.; Requena, T.; Swiatecka, D.; Wichers, H. The Impact of Food Bioactives on Health: In Vitro and Ex Vivo Models; Springer International Publishing: London, UK, 2015. [Google Scholar]
- Nunes, R.; Silva, C.; Chaves, L. Tissue-Based in vitro and Ex vivo Models for Intestinal Permeability Studies. In Concepts and Models for Drug Permeability Studies; Elsevier: Amsterdam, The Netherlands, 2016; pp. 203–236. [Google Scholar]
- Gleeson, J.P.; McCartney, F. Striving towards the Perfect In vitro Oral Drug Absorption Model. Trends Pharmacol. Sci. 2019, 40, 720–724. [Google Scholar] [CrossRef] [PubMed]
- Amirabadi, H.E.; Donkers, J.M.; Wierenga, E.; Ingenhut, B.; Pieters, L.; Stevens, L.; Donkers, T.; Westerhout, J.; Masereeuw, R.; Bobeldijk-Pastorova, I.; et al. Intestinal Explant Barrier Chip: Long-Term Intestinal Absorption Screening in a Novel Microphysiological System Using Tissue Explants. Lab Chip 2022, 22, 326–342. [Google Scholar] [CrossRef]
- Yin, L.; Yang, H.; Li, J.; Li, Y.; Ding, X.; Wu, G.; Yin, Y. Pig Models on Intestinal Development and Therapeutics. Amino Acids 2017, 49, 2099–2106. [Google Scholar] [CrossRef]
- Zhang, Q.; Widmer, G.; Tzipori, S. A Pig Model of the Human Gastrointestinal Tract. Gut Microbes 2013, 4, 193–200. [Google Scholar] [CrossRef]
- Franz-Montan, M.; Serpe, L.; Martinelli, C.C.M.; da Silva, C.B.; dos Santos, C.P.; Novaes, P.D.; Volpato, M.C.; de Paula, E.; Lopez, R.F.V.; Groppo, F.C. Evaluation of Different Pig Oral Mucosa Sites as Permeability Barrier Models for Drug Permeation Studies. Eur. J. Pharm. Sci. 2016, 81, 52–59. [Google Scholar] [CrossRef] [PubMed]
- Pastorelli, G.; Faustini, M.; Attard, E. In vitro Fermentation of Feed Ingredients by Fresh or Frozen Pig Fecal Inocula. Anim. Sci. J. 2014, 85, 690–697. [Google Scholar] [CrossRef] [PubMed]
- Ringø, E.; Jutfelt, F.; Kanapathippillai, P.; Bakken, Y.; Sundell, K.; Glette, J.; Mayhew, T.M.; Myklebust, R.; Olsen, R.E. Damaging Effect of the Fish Pathogen Aeromonas salmonicida ssp. salmonicida on Intestinal Enterocytes of Atlantic Salmon (Salmo salar L.). Cell Tissue Res. 2004, 318, 305–311. [Google Scholar] [CrossRef] [PubMed]
- Kent-Dennis, C.; Penner, G.B. Effects of Lipopolysaccharide Exposure on the Inflammatory Response, Butyrate Flux, and Metabolic Function of the Ruminal Epithelium Using an Ex vivo Model. J. Dairy Sci. 2021, 104, 2334–2345. [Google Scholar] [CrossRef] [PubMed]
- Mota de Carvalho, N.; Oliveira, D.L.; Saleh, M.A.D.; Pintado, M.E.; Madureira, A.R. Importance of Gastrointestinal in vitro Models for the Poultry Industry and Feed Formulations. Anim. Feed Sci. Technol. 2021, 271, 114730. [Google Scholar] [CrossRef]
- Lickwar, C.R.; Camp, J.G.; Weiser, M.; Cocchiaro, J.L.; Kingsley, D.M.; Furey, T.S.; Sheikh, S.Z.; Rawls, J.F. Genomic Dissection of Conserved Transcriptional Regulation in Intestinal Epithelial Cells. PLoS Biol. 2017, 15, e2002054. [Google Scholar] [CrossRef] [PubMed]
- Zachos, N.C. Viral Gastroenteritis; Elsevier Inc.: Amsterdam, The Netherlands, 2016; pp. 1–21. [Google Scholar]
- Garcia, M.A.; Nelson, W.J.; Chavez, N. Cell–Cell Junctions Organize Structural and Signaling Networks. Cold Spring Harb. Perspect. Biol. 2018, 10, a029181. [Google Scholar] [CrossRef] [PubMed]
- Odenwald, M.A.; Turner, J.R. The Intestinal Epithelial Barrier: A Therapeutic Target? Nat. Rev. Gastroenterol. Hepatol. 2017, 14, 9–21. [Google Scholar] [CrossRef]
- Edelblum, K.L.; Turner, J.R. Epithelial Cells: Structure, Transport, and Barrier Function. Structure, Transport, and Barrier Function. In Mucosal Immunology, 4th ed.; Elsevier: Amsterdam, The Netherlands, 2015; Volume 1–2, pp. 187–210. [Google Scholar] [CrossRef]
- Bischoff, S.C.; Barbara, G.; Buurman, W.; Ockhuizen, T.; Schulzke, J.D.; Serino, M.; Tilg, H.; Watson, A.; Wells, J.M. Intestinal Permeability—A New Target for Disease Prevention and Therapy. BMC Gastroenterol. 2014, 14, 189. [Google Scholar] [CrossRef]
- Lee, B.; Moon, K.M.; Kim, C.Y. Tight Junction in the Intestinal Epithelium: Its Association with Diseases and Regulation by Phytochemicals. J. Immunol. Res. 2018, 2018, 2645465. [Google Scholar] [CrossRef]
- Landy, J.; Ronde, E.; English, N.; Clark, S.K.; Hart, A.L.; Knight, S.C.; Ciclitira, P.J.; Al-Hassi, H.O. Tight Junctions in Inflammatory Bowel Diseases and Inflammatory Bowel Disease Associated Colorectal Cancer. World J. Gastroenterol. 2016, 22, 3117. [Google Scholar] [CrossRef]
- Modina, S.C.; Polito, U.; Rossi, R.; Corino, C.; di Giancamillo, A. Nutritional Regulation of Gut Barrier Integrity in Weaning Piglets. Animals 2019, 9, 1045. [Google Scholar] [CrossRef]
- Modina, S.C.; Aidos, L.; Rossi, R.; Pocar, P.; Corino, C.; di Giancamillo, A. Stages of Gut Development as a Useful Tool to Prevent Gut Alterations in Piglets. Animals 2021, 11, 1412. [Google Scholar] [CrossRef]
- Tako, E.; Bar, H.; Glahn, R.P. The Combined Application of the Caco-2 Cell Bioassay Coupled with In vivo (Gallus gallus) Feeding Trial Represents an Effective Approach to Predicting Fe Bioavailability in Humans. Nutrients 2016, 8, 732. [Google Scholar] [CrossRef]
- Denbow, D.M. Gastrointestinal Anatomy and Physiology. In Sturkie’s Avian Physiology, 6th ed.; Elsevier: Amsterdam, The Netherlands, 2015; pp. 337–366. [Google Scholar] [CrossRef]
- Alshamy, Z.; Richardson, K.C.; Hünigen, H.; Hafez, H.M.; Plendl, J.; al Masri, S. Comparison of the Gastrointestinal Tract of a Dual-Purpose to a Broiler Chicken Line: A Qualitative and Quantitative Macroscopic and Microscopic Study. PLoS ONE 2018, 13, e0204921. [Google Scholar] [CrossRef] [PubMed]
- Wijtten, P.J.A.; Langhout, D.J.; Verstegen, M.W.A. Small Intestine Development in Chicks after Hatch and in Pigs around the Time of Weaning and Its Relation with Nutrition: A Review. Acta Agric. Scand. Sect. A—Anim. Sci. 2012, 62, 1–12. [Google Scholar] [CrossRef]
- Hollemans, M.S.; van Baal, J.; de Vries Reilingh, G.; Kemp, B.; Lammers, A.; de Vries, S. Intestinal Epithelium Integrity after Delayed Onset of Nutrition in Broiler Chickens. Poult. Sci. 2020, 99, 6818–6827. [Google Scholar] [CrossRef] [PubMed]
- Günzel, D. Claudins: Vital Partners in Transcellular and Paracellular Transport Coupling. Pflugers Arch. 2017, 469, 35–44. [Google Scholar] [CrossRef]
- Yu, M.; Yang, Y.; Zhu, C.; Guo, S.; Gan, Y. Advances in the Transepithelial Transport of Nanoparticles. Drug Discov. Today 2016, 21, 1155–1161. [Google Scholar] [CrossRef] [PubMed]
- Michielan, A.; D’Incà, R. Intestinal Permeability in Inflammatory Bowel Disease: Pathogenesis, Clinical Evaluation, and Therapy of Leaky Gut. Mediators Inflamm. 2015, 2015, 628157. [Google Scholar] [CrossRef]
- Ma, T.Y.; Nighot, P.; Al-Sadi, R. Tight Junctions and the Intestinal Barrier. In Physiology of the Gastrointestinal Tract, 6th ed.; Elsevier: Amsterdam, The Netherlands, 2018; Volume 1–2, pp. 587–639. [Google Scholar] [CrossRef]
- Kiela, P.R.; Ghishan, F.K. Physiology of Intestinal Absorption and Secretion. Best Pract. Res. Clin. Gastroenterol. 2016, 30, 145–159. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.Q.; Cheung, L.S.; Feng, L.; Tanner, W.; Frommer, W.B. Transport of Sugars. Annu. Rev. Biochem. 2015, 84, 865–894. [Google Scholar] [CrossRef] [PubMed]
- Shen, W.; Matsui, T. Current Knowledge of Intestinal Absorption of Bioactive Peptides. Food Funct. 2017, 8, 4306–4314. [Google Scholar] [CrossRef]
- Ko, C.W.; Qu, J.; Black, D.D.; Tso, P. Regulation of Intestinal Lipid Metabolism: Current Concepts and Relevance to Disease. Nat. Rev. Gastroenterol. Hepatol. 2020, 17, 169–183. [Google Scholar] [CrossRef]
- Goncalves, A.; Roi, S.; Nowicki, M.; Dhaussy, A.; Huertas, A.; Amiot, M.J.; Reboul, E. Fat-Soluble Vitamin Intestinal Absorption: Absorption Sites in the Intestine and Interactions for Absorption. Food Chem. 2015, 172, 155–160. [Google Scholar] [CrossRef]
- Mace, O.J.; Marshall, F. Digestive Physiology of the Pig Symposium: Gut Chemosensing and the Regulation of Nutrient Absorption and Energy Supply. J. Anim. Sci. 2013, 91, 1932–1945. [Google Scholar] [CrossRef]
- Wijtten, P.J.A.; van der Meulen, J.; Verstegen, M.W.A. Intestinal Barrier Function and Absorption in Pigs after Weaning: A Review. Br. J. Nutr. 2011, 105, 967–981. [Google Scholar] [CrossRef]
- van der Wielen, N.; Moughan, P.J.; Mensink, M. Amino Acid Absorption in the Large Intestine of Humans and Porcine Models. J. Nutr. 2017, 147, 1493–1498. [Google Scholar] [CrossRef]
- Sun, X.; Zhang, H.; Sheikhahmadi, A.; Wang, Y.; Jiao, H.; Lin, H.; Song, Z. Effects of Heat Stress on the Gene Expression of Nutrient Transporters in the Jejunum of Broiler Chickens (Gallus gallus domesticus). Int. J. Biometeorol. 2015, 59, 127–135. [Google Scholar] [CrossRef] [PubMed]
- Karasov, W.H. Integrative Physiology of Transcellular and Paracellular Intestinal Absorption. J. Exp. Biol. 2017, 220, 2495–2501. [Google Scholar] [CrossRef]
- Berben, P.; Bauer-Brandl, A.; Brandl, M.; Faller, B.; Flaten, G.E.; Jacobsen, A.C.; Brouwers, J.; Augustijns, P. Drug Permeability Profiling Using Cell-Free Permeation Tools: Overview and Applications. Eur. J. Pharm. Sci. 2018, 119, 219–233. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.; Lu, L.; Wang, S.; Wu, J.; Shi, J.; Yan, T.; Xie, C.; Li, Q.; Hu, M.; Liu, Z. Oral Absorption Basics: Pathways and Physicochemical and Biological Factors Affecting Absorption. In Developing Solid Oral Dosage Forms: Pharmaceutical Theory and Practice, 2nd ed.; Elsevier: Amsterdam, The Netherlands, 2017; pp. 297–329. [Google Scholar] [CrossRef]
- Capuano, E.; Janssen, A.E.M. Food Matrix and Macronutrient Digestion. Annu. Rev. Food Sci. Technol. 2021, 12, 193–212. [Google Scholar] [CrossRef]
- Zhou, H.; Tan, Y.; McClements, D.J. Applications of the INFOGEST In vitro Digestion Model to Foods: A Review. Annu. Rev. Food Sci. Technol. 2023, 14, 135–156. [Google Scholar] [CrossRef] [PubMed]
- Minekus, M.; Marteau, P.; Havenaar, R.; Veld, J.H.J.H. A Multicompartmental Dynamic Computer-Controlled Model Simulating the Stomach and Small Intestine. Altern. Lab. Anim. 1995, 23, 197–209. [Google Scholar] [CrossRef]
- Molly, K.; Woestyne, M.V.; Smet, I.D.; Verstraete, W. Validation of the Simulator of the Human Intestinal Microbial Ecosystem (SHIME) Reactor Using Microorganism-Associated Activities. Microb. Ecol. Health Dis. 1994, 7, 191–200. [Google Scholar] [CrossRef]
- Egger, L.; Schlegel, P.; Baumann, C.; Stoffers, H.; Guggisberg, D.; Brügger, C.; Dürr, D.; Stoll, P.; Vergères, G.; Portmann, R. Physiological Comparability of the Harmonized INFOGEST in vitro Digestion Method to in vivo Pig Digestion. Food Res. Int. 2017, 102, 567–574. [Google Scholar] [CrossRef]
- Grundy, M.M.L.; Tang, J.; van Milgen, J.; Renaudeau, D. Cell Wall of Feeds and Their Impact on Protein Digestibility: An in vitro Method Applied for Pig Nutrition. Anim. Feed Sci. Technol. 2022, 293, 115467. [Google Scholar] [CrossRef]
- Boisen, S.; Fernández, J.A. Prediction of the Total Tract Digestibility of Energy in Feedstuffs and Pig Diets by in vitro Analyses. Anim. Feed Sci. Technol. 1997, 68, 277–286. [Google Scholar] [CrossRef]
- Święch, E. Alternative Prediction Methods of Protein and Energy Evaluation of Pig Feeds. J. Anim. Sci. Biotechnol. 2017, 8, 39. [Google Scholar] [CrossRef]
- Nash, T.; Vervelde, L. Advances, Challenges and Future Applications of Avian Intestinal in vitro Models. Avian Pathol. 2022, 51, 317–329. [Google Scholar] [CrossRef]
- Martinez-Haro, M.; Taggart, M.A.; Green, A.J.; Mateo, R. Avian Digestive Tract Simulation To Study the Effect of Grit Geochemistry and Food on Pb Shot Bioaccessibility. Environ. Sci. Technol. 2009, 43, 9480–9486. [Google Scholar] [CrossRef]
- Meimandipour, A.; Shuhaimi, M.; Hair-Bejo, M.; Azhar, K.; Kabeir, B.M.; Rasti, B.; Yazid, A.M. In vitro Fermentation of Broiler Cecal Content: The Role of Lactobacilli and PH Value on the Composition of Microbiota and End Products Fermentation. Lett. Appl. Microbiol. 2009, 49, 415–420. [Google Scholar] [CrossRef]
- de Carvalho, N.M.; Costa, C.M.; Castro, C.; Saleh, M.A.D.; Pintado, M.E.; Oliveira, D.L.; Madureira, A.R. Development of a Chicken Gastrointestinal Tract (GIT) Simulation Model: Impact of Cecal Inoculum Storage Preservation Conditions. Appl. Microbiol. 2023, 3, 968–992. [Google Scholar] [CrossRef]
- Xu, Y.; Shrestha, N.; Préat, V.; Beloqui, A. An Overview of in vitro, Ex vivo and in vivo Models for Studying the Transport of Drugs across Intestinal Barriers. Adv. Drug Deliv. Rev. 2021, 175, 113795. [Google Scholar] [CrossRef] [PubMed]
- Luo, Z.; Liu, Y.; Zhao, B.; Tang, M.; Dong, H.; Zhang, L.; Lv, B.; Wei, L. Ex vivo and in Situ Approaches Used to Study Intestinal Absorption. J. Pharmacol. Toxicol. Methods 2013, 68, 208–216. [Google Scholar] [CrossRef] [PubMed]
- Ussing, H.H.; Zerahn, K. Active Transport of Sodium as the Source of Electric Current in the Short-Circuited Isolated Frog Skin. Acta Physiol. Scand. 1951, 23, 110–127. [Google Scholar] [CrossRef] [PubMed]
- Thomson, A.; Smart, K.; Somerville, M.S.; Lauder, S.N.; Appanna, G.; Horwood, J.; Sunder Raj, L.; Srivastava, B.; Durai, D.; Scurr, M.J.; et al. The Ussing Chamber System for Measuring Intestinal Permeability in Health and Disease. BMC Gastroenterol. 2019, 19, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Clarke, L.L. A Guide to Ussing Chamber Studies of Mouse Intestine. Am. J. Physiol. Gastrointest. Liver Physiol. 2009, 296, 1151–1166. [Google Scholar] [CrossRef]
- Sjögren, E.; Eriksson, J.; Vedin, C.; Breitholtz, K.; Hilgendorf, C. Excised Segments of Rat Small Intestine in Ussing Chamber Studies: A Comparison of Native and Stripped Tissue Viability and Permeability to Drugs. Int. J. Pharm. 2016, 505, 361–368. [Google Scholar] [CrossRef] [PubMed]
- Wuyts, B.; Riethorst, D.; Brouwers, J.; Tack, J.; Annaert, P.; Augustijns, P. Evaluation of Fasted and Fed State Simulated and Human Intestinal Fluids as Solvent System in the Ussing Chambers Model to Explore Food Effects on Intestinal Permeability. Int. J. Pharm. 2015, 478, 736–744. [Google Scholar] [CrossRef] [PubMed]
- Westerhout, J.; van de Steeg, E.; Grossouw, D.; Zeijdner, E.E.; Krul, C.A.M.; Verwei, M.; Wortelboer, H.M. A New Approach to Predict Human Intestinal Absorption Using Porcine Intestinal Tissue and Biorelevant Matrices. Eur. J. Pharm. Sci. 2014, 63, 167–177. [Google Scholar] [CrossRef] [PubMed]
- Supe, S.; Takudage, P. Methods for Evaluating Penetration of Drug into the Skin: A Review. Ski. Res. Technol. 2021, 27, 299–308. [Google Scholar] [CrossRef] [PubMed]
- Dezani, A.B.; Pereira, T.M.; Caffaro, A.M.; Reis, J.M.; Serra, C.H. dos R. Determination of Lamivudine and Zidovudine Permeability Using a Different Ex vivo Method in Franz Cells. J. Pharmacol. Toxicol. Methods 2013, 67, 194–202. [Google Scholar] [CrossRef] [PubMed]
- Wilson, T.H.; Wiseman, G. The Use of Sacs of Everted Small Intestine for the Study of the Transference of Substances from the Mucosal to the Serosal Surface. J. Physiol. 1954, 123, 116. [Google Scholar] [CrossRef] [PubMed]
- Alam, M.A.; Al-Jenoobi, F.I.; Al-Mohizea, A.M. Everted Gut Sac Model as a Tool in Pharmaceutical Research: Limitations and Applications. J. Pharm. Pharmacol. 2012, 64, 326–336. [Google Scholar] [CrossRef]
- Noben, M.; Vanhove, W.; Arnauts, K.; Ramalho, A.S.; van Assche, G.; Vermeire, S.; Verfaillie, C.; Ferrante, M. Human Intestinal Epithelium in a Dish: Current Models for Research into Gastrointestinal Pathophysiology. United Eur. Gastroenterol. J. 2017, 5, 1073–1081. [Google Scholar] [CrossRef]
- Lechanteur, A.; Almeida, A.; Sarmento, B. Elucidation of the Impact of Cell Culture Conditions of Caco-2 Cell Monolayer on Barrier Integrity and Intestinal Permeability. Eur. J. Pharm. Biopharm. 2017, 119, 137–141. [Google Scholar] [CrossRef]
- Pereira, C.; Costa, J.; Sarmento, B.; Araújo, F. Cell-Based in vitro Models for Intestinal Permeability Studies. In Concepts and Models for Drug Permeability Studies: Cell and Tissue Based In Vitro Culture Models; Elsevier: Amsterdam, The Netherlands, 2016; pp. 57–81. [Google Scholar] [CrossRef]
- Volpe, D.A. Advances in Cell-Based Permeability Assays to Screen Drugs for Intestinal Absorption. Expert Opin. Drug Discov. 2020, 15, 539–549. [Google Scholar] [CrossRef]
- Pan, F.; Han, L.; Zhang, Y.; Yu, Y.; Liu, J. Optimization of Caco-2 and HT29 Co-Culture in vitro Cell Models for Permeability Studies. Int. J. Food Sci. Nutr. 2015, 66, 680–685. [Google Scholar] [CrossRef]
- Lozoya-Agullo, I.; Araújo, F.; González-Álvarez, I.; Merino-Sanjuán, M.; González-Álvarez, M.; Bermejo, M.; Sarmento, B. Usefulness of Caco-2/HT29-MTX and Caco-2/HT29-MTX/Raji B Coculture Models to Predict Intestinal and Colonic Permeability Compared to Caco-2 Monoculture. Mol. Pharm. 2017, 14, 1264–1270. [Google Scholar] [CrossRef] [PubMed]
- Silva-Almeida, C.; Ewart, M.A.; Wilde, C. 3D Gastrointestinal Models and Organoids to Study Metabolism in Human Colon Cancer. Semin. Cell Dev. Biol. 2020, 98, 98–104. [Google Scholar] [CrossRef]
- Meneses, A.M.C.; Schneeberger, K.; Kruitwagen, H.S.; Penning, L.C.; van Steenbeek, F.G.; Burgener, I.A.; Spee, B. Intestinal Organoids—Current and Future Applications. Vet. Sci. 2016, 3, 31. [Google Scholar] [CrossRef] [PubMed]
- Dotti, I.; Mayorgas, A.; Salas, A. Generation of Human Colon Organoids from Healthy and Inflammatory Bowel Disease Mucosa. PLoS ONE 2022, 17, e0276195. [Google Scholar] [CrossRef]
- Fois, C.A.M.; Le, T.Y.L.; Schindeler, A.; Naficy, S.; McClure, D.D.; Read, M.N.; Valtchev, P.; Khademhosseini, A.; Dehghani, F. Models of the Gut for Analyzing the Impact of Food and Drugs. Adv. Healthc. Mater. 2019, 8, 1900968. [Google Scholar] [CrossRef]
- Signore, M.A.; de Pascali, C.; Giampetruzzi, L.; Siciliano, P.A.; Francioso, L. Gut-on-Chip Microphysiological Systems: Latest Advances in the Integration of Sensing Strategies and Adoption of Mature Detection Mechanisms. Sens. Biosensing Res. 2021, 33, 100443. [Google Scholar] [CrossRef]
- Jalili-Firoozinezhad, S.; Gazzaniga, F.S.; Calamari, E.L.; Camacho, D.M.; Fadel, C.W.; Bein, A.; Swenor, B.; Nestor, B.; Cronce, M.J.; Tovaglieri, A.; et al. A Complex Human Gut Microbiome Cultured in an Anaerobic Intestine-on-a-Chip. Nat. Biomed. Eng. 2019, 3, 520–531. [Google Scholar] [CrossRef]
- Xiang, Y.; Wen, H.; Yu, Y.; Li, M.; Fu, X.; Huang, S. Gut-on-Chip: Recreating Human Intestine in vitro. J. Tissue Eng. 2020, 11, 2041731420965318. [Google Scholar] [CrossRef]
- Ingber, D.E. Human Organs-on-Chips for Disease Modelling, Drug Development and Personalized Medicine. Nat. Rev. Genet. 2022, 23, 467–491. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Pan, H.; Zhang, C.; Zhao, L.; Zhao, R.; Zhu, Y.; Pan, W. Developments in Methods for Measuring the Intestinal Absorption of Nanoparticle-Bound Drugs. Int. J. Mol. Sci. 2016, 17, 1171. [Google Scholar] [CrossRef]
- Moreno-Montoro, M.; Jauregi, P.; Navarro-Alarcón, M.; Olalla-Herrera, M.; Giménez-Martínez, R.; Amigo, L.; Miralles, B. Bioaccessible Peptides Released by in vitro Gastrointestinal Digestion of Fermented Goat Milks. Anal. Bioanal. Chem. 2018, 410, 3597–3606. [Google Scholar] [CrossRef] [PubMed]
- Blancas-Benitez, F.J.; Pérez-Jiménez, J.; Montalvo-González, E.; González-Aguilar, G.A.; Sáyago-Ayerdi, S.G. In vitro Evaluation of the Kinetics of the Release of Phenolic Compounds from Guava (Psidium guajava L.) Fruit. J. Funct. Foods 2018, 43, 139–145. [Google Scholar] [CrossRef]
- González, C.; González, D.; Zúñiga, R.N.; Estay, H.; Troncoso, E. Simulation of Human Small Intestinal Digestion of Starch Using an In vitro System Based on a Dialysis Membrane Process. Foods 2020, 9, 913. [Google Scholar] [CrossRef]
- Gayoso, L.; Claerbout, A.S.; Calvo, M.I.; Cavero, R.Y.; Astiasarán, I.; Ansorena, D. Bioaccessibility of Rutin, Caffeic Acid and Rosmarinic Acid: Influence of the in vitro Gastrointestinal Digestion Models. J. Funct. Foods 2016, 26, 428–438. [Google Scholar] [CrossRef]
- Guerra, A.; Denis, S.; le Goff, O.; Sicardi, V.; François, O.; Yao, A.F.; Garrait, G.; Manzi, A.P.; Beyssac, E.; Alric, M.; et al. Development and Validation of a New Dynamic Computer-Controlled Model of the Human Stomach and Small Intestine. Biotechnol. Bioeng. 2016, 113, 1325–1335. [Google Scholar] [CrossRef]
- Carrasco-Correa, E.J.; Ruiz-Allica, J.; Rodríguez-Fernández, J.F.; Miró, M. Human Artificial Membranes in (Bio)Analytical Science: Potential for in vitro Prediction of Intestinal Absorption-A Review. TrAC Trends Anal. Chem. 2021, 145, 116446. [Google Scholar] [CrossRef]
- di Cagno, M.; Bibi, H.A.; Bauer-Brandl, A. New Biomimetic Barrier PermeapadTM for Efficient Investigation of Passive Permeability of Drugs. Eur. J. Pharm. Sci. 2015, 73, 29–34. [Google Scholar] [CrossRef]
- B Sánchez, A.; C Calpena, A.; Mallandrich, M.; Clares, B. Validation of an Ex vivo Permeation Method for the Intestinal Permeability of Different BCS Drugs and Its Correlation with Caco-2 in vitro Experiments. Pharmaceutics 2019, 11, 638. [Google Scholar] [CrossRef]
- Chen, P.; Zhao, M.; Chen, Q.; Fan, L.; Gao, F.; Zhao, L. Absorption Characteristics of Chitobiose and Chitopentaose in the Human Intestinal Cell Line Caco-2 and Everted Gut Sacs. J. Agric. Food Chem. 2019, 67, 4513–4523. [Google Scholar] [CrossRef]
- Pierzchalska, M.; Panek, M.; Czyrnek, M.; Grabacka, M. The Three-Dimensional Culture of Epithelial Organoids Derived from Embryonic Chicken Intestine. Methods Mol. Biol. 2016, 1576, 135–144. [Google Scholar] [CrossRef]
- Baskara, A.P.; Sharma, S.; Sener-Aydemir, A.; Koger, S.; Ariyadi, B.; Dono, N.D.; Zuprizal, Z.; Metzler-Zebeli, B.U. Cinnamon Bark Oil and Coconut Oil Emulsions Modified Small Intestinal Motility and Barrier Function in Laying Hens in an Ex vivo Experiment. Br. Poult. Sci. 2021, 62, 435–442. [Google Scholar] [CrossRef] [PubMed]
- Deußer, H.; Rogoll, D.; Scheppach, W.; Volk, A.; Melcher, R.; Richling, E. Gastrointestinal Absorption and Metabolism of Apple Polyphenols Ex vivo by the Pig Intestinal Mucosa in the Ussing Chamber. Biotechnol. J. 2013, 8, 363–370. [Google Scholar] [CrossRef] [PubMed]
- Ruhnke, I.; Röhe, I.; Goodarzi Boroojeni, F.; Knorr, F.; Mader, A.; Hafeez, A.; Zentek, J. Feed Supplemented with Organic Acids Does Not Affect Starch Digestibility, nor Intestinal Absorptive or Secretory Function in Broiler Chickens. J. Anim. Physiol. Anim. Nutr. 2015, 99, 29–35. [Google Scholar] [CrossRef] [PubMed]
- Ozorio, L.; Mellinger-Silva, C.; Cabral, L.M.C.; Jardin, J.; Boudry, G.; Dupont, D. The Influence of Peptidases in Intestinal Brush Border Membranes on the Absorption of Oligopeptides from Whey Protein Hydrolysate: An Ex vivo Study Using an Ussing Chamber. Foods 2020, 9, 1415. [Google Scholar] [CrossRef]
- Tang, X.; Zhang, M.; Zhang, H.; Pan, Y.; Dong, Q.; Xin, Y.; Ho, C.T.; Huang, Q. Evaluation of the Bioaccessibility of Tetrahydrocurcumin-Hyaluronic Acid Conjugate Using in vitro and Ex vivo Models. Int. J. Biol. Macromol. 2021, 182, 1322–1330. [Google Scholar] [CrossRef] [PubMed]
- Espinal-Ruiz, M.; Restrepo-Sánchez, L.P.; Narváez-Cuenca, C.E. Effect of Pectins on the Mass Transfer Kinetics of Monosaccharides, Amino Acids, and a Corn Oil-in-Water Emulsion in a Franz Diffusion Cell. Food Chem. 2016, 209, 144–153. [Google Scholar] [CrossRef] [PubMed]
- Shimizu, K.; Komaki, Y.; Fukano, N.; Bungo, T. Transporter Gene Expression and Transference of Fructose in Broiler Chick Intestine. J. Poult. Sci. 2018, 55, 0170095. [Google Scholar] [CrossRef]
- Mebratu, A.T.; Asfaw, Y.T.; Paul, G.; Janssens, J. Exploring the Functional and Metabolic Effects of Feeding Garra Fish Meal to Broiler Chickens. Prepint 2021. [Google Scholar] [CrossRef]
- Mahalakshmi, L.; Leena, M.M.; Moses, J.A.; Anandharamakrishnan, C. Micro- and Nano-Encapsulation of β-Carotene in Zein Protein: Size-Dependent Release and Absorption Behavior. Food Funct. 2020, 11, 1647–1660. [Google Scholar] [CrossRef]
- Herrera-Cazares, L.A.; Luzardo-Ocampo, I.; Ramírez-Jiménez, A.K.; Gutiérrez-Uribe, J.A.; Campos-Vega, R.; Gaytán-Martínez, M. Influence of Extrusion Process on the Release of Phenolic Compounds from Mango (Mangifera indica L.) Bagasse-Added Confections and Evaluation of Their Bioaccessibility, Intestinal Permeability, and Antioxidant Capacity. Food Res. Int. 2021, 148, 110591. [Google Scholar] [CrossRef]
- Leena, M.M.; Anukiruthika, T.; Moses, J.A.; Anandharamakrishnan, C. Co-Delivery of Curcumin and Resveratrol through Electrosprayed Core-Shell Nanoparticles in 3D Printed Hydrogel. Food Hydrocoll. 2022, 124, 107200. [Google Scholar] [CrossRef]
- Sangsawad, P.; Roytrakul, S.; Choowongkomon, K.; Kitts, D.D.; Chen, X.M.; Meng, G.; Li-Chan, E.C.Y.; Yongsawatdigul, J. Transepithelial Transport across Caco-2 Cell Monolayers of Angiotensin Converting Enzyme (ACE) Inhibitory Peptides Derived from Simulated in vitro Gastrointestinal Digestion of Cooked Chicken Muscles. Food Chem. 2018, 251, 77–85. [Google Scholar] [CrossRef] [PubMed]
- Hernandez-Patlan, D.; Solis-Cruz, B.; Pontin, K.P.; Latorre, J.D.; Baxter, M.F.A.; Hernandez-Velasco, X.; Merino-Guzman, R.; Méndez-Albores, A.; Hargis, B.M.; Lopez-Arellano, R.; et al. Evaluation of a Solid Dispersion of Curcumin with Polyvinylpyrrolidone and Boric Acid against Salmonella Enteritidis Infection and Intestinal Permeability in Broiler Chickens: A Pilot Study. Front. Microbiol. 2018, 9, 1289. [Google Scholar] [CrossRef]
- Vieira, E.F.; das Neves, J.; Ferreira, I.M.P.L.V.O. Bioactive Protein Hydrolysate Obtained from Canned Sardine and Brewing By-Products: Impact of Gastrointestinal Digestion and Transepithelial Absorption. Waste Biomass Valorization 2021, 12, 1281–1292. [Google Scholar] [CrossRef]
- Arranz, E.; Guri, A.; Fornari, T.; Mendiola, J.A.; Reglero, G.; Corredig, M. In vitro Uptake and Immune Functionality of Digested Rosemary Extract Delivered through Food Grade Vehicles. Food Res. Int. 2017, 97, 71–77. [Google Scholar] [CrossRef]
- Brufau, M.T.; Campo-Sabariz, J.; Carné, S.; Ferrer, R.; Martín-Venegas, R. Salmosan, a β-Galactomannan-Rich Product, in Combination with Lactobacillus Plantarum Contributes to Restore Intestinal Epithelial Barrier Function by Modulation of Cytokine Production. J. Nutr. Biochem. 2017, 41, 20–24. [Google Scholar] [CrossRef]
- Gunathilake, K.D.P.P.; Ranaweera, K.K.D.S.; Rupasinghe, H.P.V. Change of Phenolics, Carotenoids, and Antioxidant Capacity Following Simulated Gastrointestinal Digestion and Dialysis of Selected Edible Green Leaves. Food Chem. 2018, 245, 371–379. [Google Scholar] [CrossRef] [PubMed]
- Petit, C.; Bujard, A.; Skalicka-Woźniak, K.; Cretton, S.; Houriet, J.; Christen, P.; Carrupt, P.A.; Wolfender, J.L. Prediction of the Passive Intestinal Absorption of Medicinal Plant Extract Constituents with the Parallel Artificial Membrane Permeability Assay (PAMPA). Planta Med. 2016, 82, 424–431. [Google Scholar] [CrossRef] [PubMed]
- Navarro Del Hierro, J.; Piazzini, V.; Reglero, G.; Martin, D.; Bergonzi, M.C. In vitro Permeability of Saponins and Sapogenins from Seed Extracts by the Parallel Artificial Membrane Permeability Assay: Effect of in vitro Gastrointestinal Digestion. J. Agric. Food Chem. 2020, 68, 1297–1305. [Google Scholar] [CrossRef] [PubMed]
- Charalabidis, A.; Sfouni, M.; Bergström, C.; Macheras, P. The Biopharmaceutics Classification System (BCS) and the Biopharmaceutics Drug Disposition Classification System (BDDCS): Beyond Guidelines. Int. J. Pharm. 2019, 566, 264–281. [Google Scholar] [CrossRef] [PubMed]
| Technique | Description | Advantages | Disadvantages | |
|---|---|---|---|---|
| Tissue-based methods | Ussing chamber | Two-chamber system split up by a tissue segment in a horizontal orientation; two electrodes, one that provides current and another that measures TEER; continuous temperature and gas supply to media. | - Technique is well established and validated; - Allows for permeation evaluation across distinct intestinal regions; - Continuous system upgrade; - Allows the assessment of carrier-mediated transport. | - Not adequate for all animal tissues; - Mandatory muscle layer removal; - High costs of system implementation and requires technical skills; - Low to medium throughput. |
| Franz diffusion cells | System with two chambers (donor and receptor) displayed in a vertical orientation, divided by a tissue segment; controlled agitation and temperature. | - Less technically demanding than Ussing chamber; - Low-cost implementation system; - Versatile by allowing the use of distinct intestinal regions; - Medium throughput, as it depends on the number of Franz cells acquired. | - Not robustly validated for intestinal permeation studies, as it is for skin permeation assays. | |
| Intestinal rings and segments | Isolated intestinal segment or ring submerged in a buffer solution with the compound of interest dissolved to assess its uptake through enterocytes. | - Practical and simple; - Agitation available (optional); - High throughput of samples; - Versatility that allows the evaluation of distinct intestinal sections; - More valuable for radiolabeled compounds. | - Overly simplified system; - Does not allow for the determination of the direction of the transepithelial transport; - Decreasing relevance of this technique since less publications report it. | |
| Everted gut sac | Intestinal section reverted and filled with a buffer; both ends are tied, before incubation, with the compound in study; controlled temperature, aeration and stirring. | - Simple and inexpensive implementation; - High throughput; - Useful for determining drug absorption mechanisms and their metabolization and the roles of enzymes and transporters; - More useful if radiolabeled compounds are used. | - Tissue reversion process may induce structural damages; - Leads to unrealistic absorption times. | |
| Cell-based methods | 2D (monocultures/co-cultures of Caco-2 and/or HT29-MTX) | Cultured cells against a flat surface, such as an insert. This can be conducted as a monoculture or as a co-culture (Caco-2 plus HT29-MTX). | - Most traditional approach with several implemented protocols, with widely accepted results among regulatory institutions; - Highly proliferative cells that are relatively easy and cheap to culture; - Co-cultures of Caco-2 and HT29-MTX represent a more realistic approach than monocultures. | - Reduced complexity and physiological relevance; - Tumor-like nature may introduce bias in the results obtained; - Possibility of flat surface interference in cells’ polarization. - Relative incompatibility with certain food components; - Variability in transporters’ expression may impair comparisons of permeability rates. |
| 3D (organoids) | Primary multicellular system, isolated from crypt cells, that can grow indefinitely and is allowed to grow in a 3D manner. | - Allows cell changes in their shape; - Promotion of more complex cell-to-cell connections than other rigid systems; - Comprises all cell types and the complexity of the in vivo epithelium; - Can be cultured indefinitely; - Can be cultured from different species. | - Demands a cell culture laboratory, with skilled personnel; - The established protocols are mainly focused on regenerative medicine purposes; - Expensive and demanding of complex technical skills. | |
| Gut-on-chip | The human Gut Chip is a microfluidic culture device composed of a transparent silicone polymer that is composed of two channels separated by a porous membrane. On one side, human intestinal epithelial cells are cultured, and human microvascular endothelial cells are on the opposite side. | - In comparison with organoids, gut-on-chip offers greater experimental control through multiple connected microfluidic channels; - Allows for the mimicking of the interactions of the intestine with the microbiota; - Allows for the customization of a disease phenotype. | - Further optimization required for size—TEER electrodes; - Lack of standardized design and materials for the polymer device. | |
| Non-biological methods | Synthetic membranes for dialysis purposes | Semi-permeable membrane with a determined pore size that separates two solutions; it evaluates molecule movement across the membrane, depending on a concentration gradient as a driving force and can be subdivided into positive or negative dialysis. | - Wide range of systems from simple to high throughputs; - Alternative for biological tissue shortages. | - Does not mimic the epithelium physiology; - More sophisticated or automated systems require skills and initial investment; - Different pore sizes reported in the literature. |
| Cell-free systems (PAMPA, PVPA, Permeapad® and AMI-system) | Cell-free permeation systems that are usually placed between a donor and acceptor chamber and differ on the type of barrier, which can be biomimetic or non-biomimetic. These systems can be applied in 96-well plates, Franz cells and plate inserts. | - Simple and ready to use, not requiring lengthy and expensive preparation steps, as in cell-based methods; - Distinct types of membranes can be applied according to the physicochemical properties of the compound in study. | - Only applicable for predicting passive transcellular drug transport, since paracellular and active transports cannot be mimicked. | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Costa, C.M.; de Carvalho, N.M.; de Oliveira, D.L.; Madureira, A.R. A Critical Review on In Vitro and Ex Vivo Models of the Intestinal Epithelium of Humans and Monogastric Animals. Gastrointest. Disord. 2024, 6, 337-358. https://doi.org/10.3390/gidisord6010023
Costa CM, de Carvalho NM, de Oliveira DL, Madureira AR. A Critical Review on In Vitro and Ex Vivo Models of the Intestinal Epithelium of Humans and Monogastric Animals. Gastrointestinal Disorders. 2024; 6(1):337-358. https://doi.org/10.3390/gidisord6010023
Chicago/Turabian StyleCosta, Célia Maria, Nelson Mota de Carvalho, Diana Luazi de Oliveira, and Ana Raquel Madureira. 2024. "A Critical Review on In Vitro and Ex Vivo Models of the Intestinal Epithelium of Humans and Monogastric Animals" Gastrointestinal Disorders 6, no. 1: 337-358. https://doi.org/10.3390/gidisord6010023
APA StyleCosta, C. M., de Carvalho, N. M., de Oliveira, D. L., & Madureira, A. R. (2024). A Critical Review on In Vitro and Ex Vivo Models of the Intestinal Epithelium of Humans and Monogastric Animals. Gastrointestinal Disorders, 6(1), 337-358. https://doi.org/10.3390/gidisord6010023