Cellular Effects of Rhynchophylline and Relevance to Sleep Regulation

 and
and 
Abstract
1. Introduction
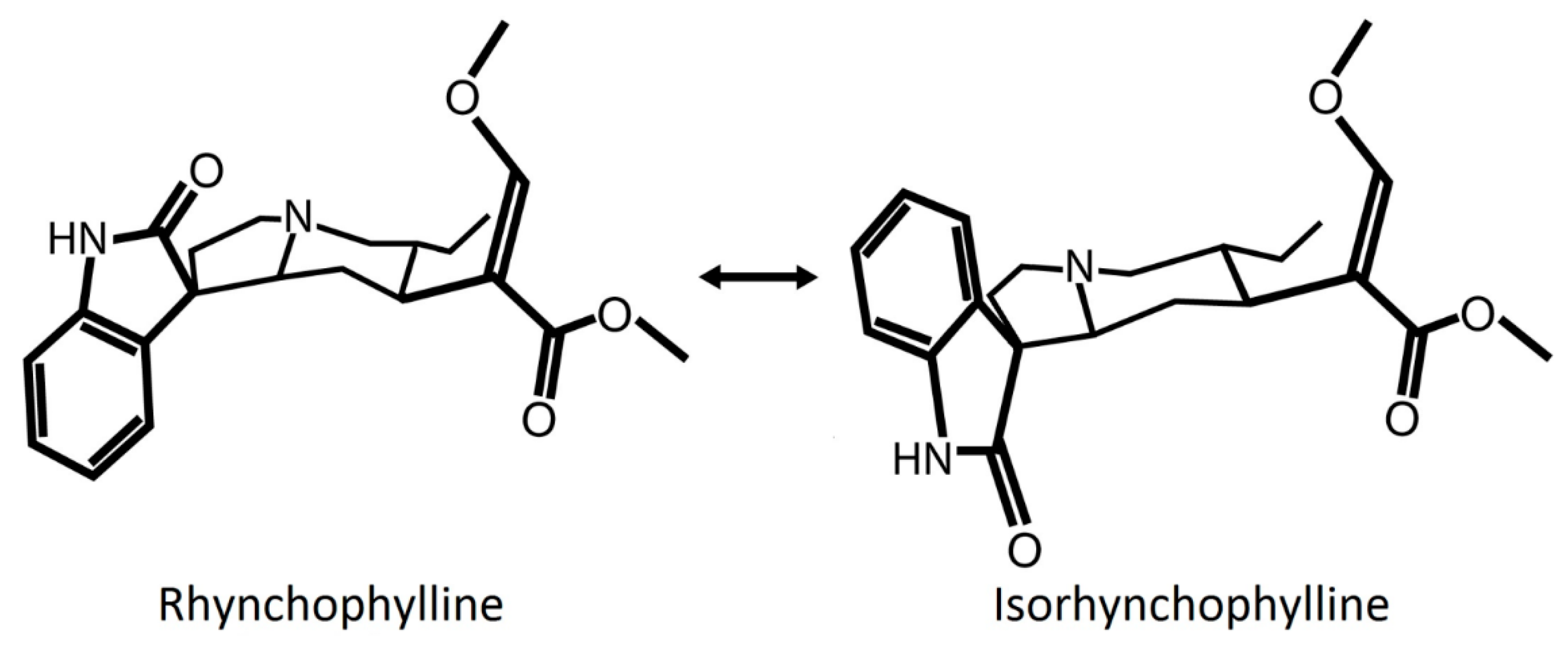
1.1. Rhynchophylline Pharmacology
1.2. Sleep and Its Regulation
1.3. Rhynchophylline and Sleep
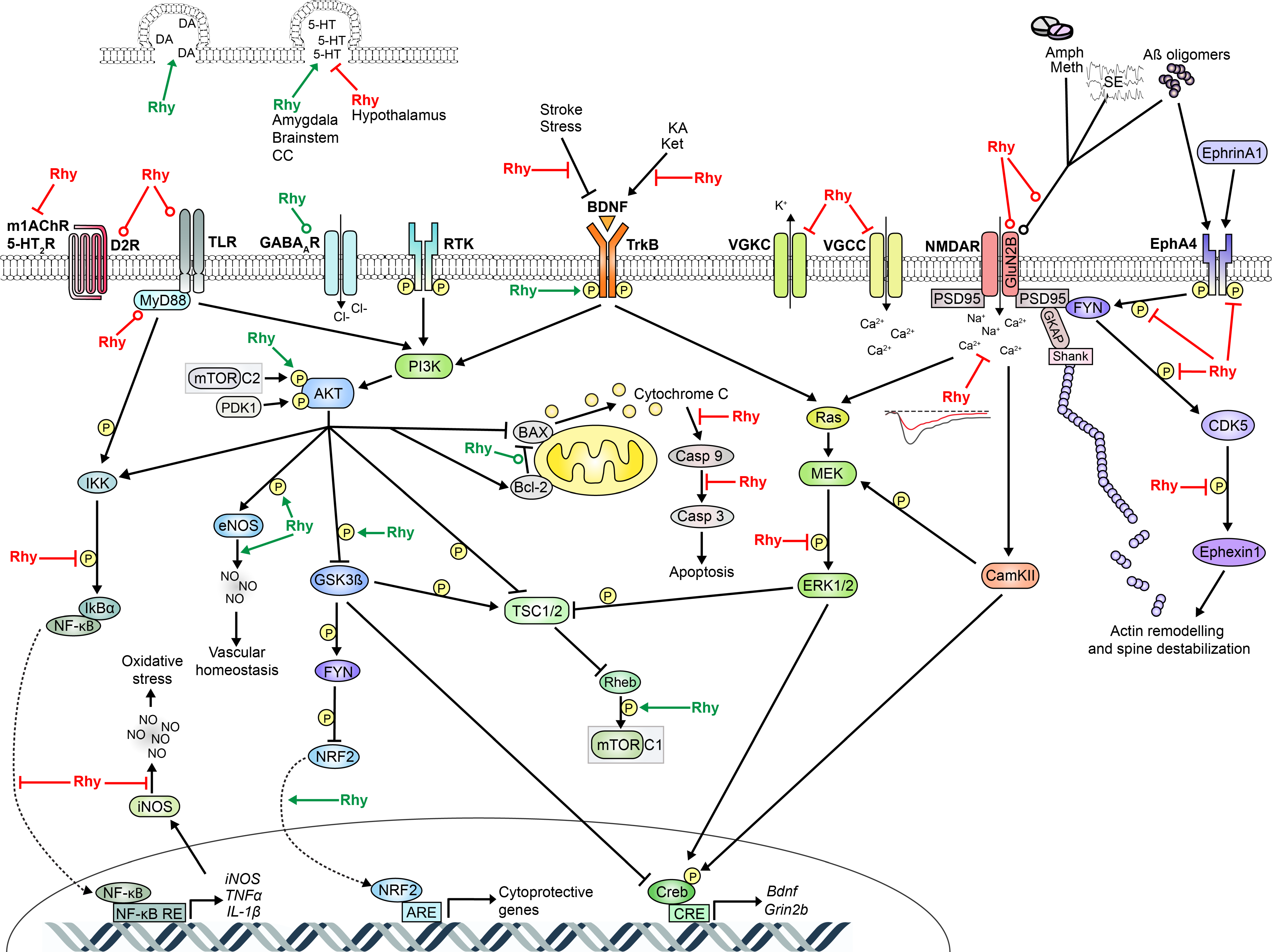
2. Rhy Targets and Links to Sleep Regulation
2.1. Ion Channels
2.1.1. Voltage-Gated Calcium Channels
2.1.2. Potassium Channels
2.2. NMDA Receptors
2.3. EphA4 and Downstream Pathways
2.4. BDNF/TrkB Signaling
2.5. ERK/MAPK Pathway
2.6. PI3K/AKT Signaling Network
2.7. NF-κB and Neuroinflammation
2.8. Neurotransmitters Signaling
3. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sarris, J.; Panossian, A.; Schweitzer, I.; Stough, C.; Scholey, A. Herbal medicine for depression, anxiety and insomnia: A review of psychopharmacology and clinical evidence. Eur. Neuropsychopharmacol. 2011, 21, 841–860. [Google Scholar] [CrossRef]
- Singh, A.; Zhao, K. Treatment of insomnia with traditional chinese herbal medicine. Int. Rev. Neurobiol. 2017, 135, 97–115. [Google Scholar] [CrossRef]
- Yang, W.; Ip, S.P.; Liu, L.; Xian, Y.F.; Lin, Z.X. Uncaria rhynchophylla and its major constituents on central nervous system: A review on their pharmacological actions. Curr. Vasc. Pharmacol. 2019. [Google Scholar] [CrossRef]
- Ndagijimana, A.; Wang, X.; Pan, G.; Zhang, F.; Feng, H.; Olaleye, O. A review on indole alkaloids isolated from Uncaria rhynchophylla and their pharmacological studies. Fitoterapia 2013, 86, 35–47. [Google Scholar] [CrossRef]
- Shi, J.S.; Yu, J.X.; Chen, X.P.; Xu, R.X. Pharmacological actions of Uncaria alkaloids, rhynchophylline and isorhynchophylline. Acta Pharmacol. Sin. 2003, 24, 97–101. [Google Scholar]
- Shinno, H.; Inami, Y.; Inagaki, T.; Nakamura, Y.; Horiguchi, J. Effect of Yi-Gan San on psychiatric symptoms and sleep structure at patients with behavioral and psychological symptoms of dementia. Prog. Neuropsychopharmacol. Biol. Psychiatry 2008, 32, 881–885. [Google Scholar] [CrossRef] [PubMed]
- Ozone, M.; Yagi, T.; Chiba, S.; Aoki, K.; Kuroda, A.; Mitsui, K.; Itoh, H.; Sasaki, M. Effect of yokukansan on psychophysiological insomnia evaluated using cyclic alternating pattern as an objective marker of sleep instability. Sleep Biol. Rhythm 2012, 10, 157–160. [Google Scholar] [CrossRef]
- Nakamura, Y.; Tajima, K.; Kawagoe, I.; Kanai, M.; Mitsuhata, H. Efficacy of traditional herbal medicine, Yokukansan on patients with neuropathic pain. Masui 2009, 58, 1248–1255. [Google Scholar]
- Yamanaka, E.; Kimizuka, Y.; Aimi, N.; Sakai, S.; Haginiwa, J. Studies of plants containing indole alkaloids. IX. Quantitative analysis of tertiary alkaloids in various parts of Uncaria rhynchophylla MIQ. Yakugaku Zasshi 1983, 103, 1028–1033. [Google Scholar] [CrossRef]
- Zhou, J.; Zhou, S. Antihypertensive and neuroprotective activities of rhynchophylline: The role of rhynchophylline in neurotransmission and ion channel activity. J. Ethnopharmacol. 2010, 132, 15–27. [Google Scholar] [CrossRef]
- Laus, G.; Teppner, H. The alkaloids of an Uncaria rhynchophylla (Rubiaceae-Coptosapelteae). Phyton (Horn Austria) 1996, 36, 185–196. [Google Scholar]
- Laus, G.; Brössner, D.; Keplinger, K. Alkaloids of peruvian Uncaria tomentosa. Phytochemistry 1997, 45, 855–860. [Google Scholar] [CrossRef]
- Wang, X.; Zheng, M.; Liu, J.; Huang, Z.; Bai, Y.; Ren, Z.; Wang, Z.; Tian, Y.; Qiao, Z.; Liu, W.; et al. Differences of first-pass effect in the liver and intestine contribute to the stereoselective pharmacokinetics of rhynchophylline and isorhynchophylline epimers in rats. J. Ethnopharmacol. 2017, 209, 175–183. [Google Scholar] [CrossRef]
- Wu, Z.F.; Wang, Y.Q.; Wan, N.; Ke, G.; Yue, P.F.; Chen, H.; Zhan, J.J.; Yang, M. Structural stabilities and transformation mechanism of Rhynchophylline and Isorhynchophylline by ultra performance liquid chromatography/time-of-flight mass spectrometry (UPLC/Q-TOF-MS). Molecules 2015, 20, 14849–14859. [Google Scholar] [CrossRef]
- Lee, C.J.; Hsueh, T.Y.; Lin, L.C.; Tsai, T.H. Determination of protein-unbound rhynchophylline brain distribution by microdialysis and ultra-performance liquid chromatography with tandem mass spectrometry. Biomed. Chromatogr. 2014, 28, 901–906. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.N.; Yang, Y.F.; Xu, W.; Yang, X.W. The blood-brain barrier permeability of six indole alkaloids from Uncariae Ramulus cum Uncis in the MDCK-pHaMDR cell monolayer model. Molecules 2017, 22, 1944. [Google Scholar] [CrossRef]
- Zhang, J.C.; Yao, W.; Qu, Y.; Nakamura, M.; Dong, C.; Yang, C.; Ren, Q.; Ma, M.; Han, M.; Shirayama, Y.; et al. Increased EphA4-ephexin1 signaling in the medial prefrontal cortex plays a role in depression-like phenotype. Sci. Rep. 2017, 7, 7133. [Google Scholar] [CrossRef]
- Fu, A.K.; Hung, K.W.; Huang, H.; Gu, S.; Shen, Y.; Cheng, E.Y.; Ip, F.C.; Huang, X.; Fu, W.Y.; Ip, N.Y. Blockade of EphA4 signaling ameliorates hippocampal synaptic dysfunctions in mouse models of Alzheimer's disease. Proc. Natl. Acad. Sci. USA 2014, 111, 9959–9964. [Google Scholar] [CrossRef]
- Shao, H.; Yang, Y.; Mi, Z.; Zhu, G.X.; Qi, A.P.; Ji, W.G.; Zhu, Z.R. Anticonvulsant effect of Rhynchophylline involved in the inhibition of persistent sodium current and NMDA receptor current in the pilocarpine rat model of temporal lobe epilepsy. Neuroscience 2016, 337, 355–369. [Google Scholar] [CrossRef]
- Zhou, J.Y.; Mo, Z.X.; Zhou, S.W. Rhynchophylline down-regulates NR2B expression in cortex and hippocampal CA1 area of amphetamine-induced conditioned place preference rat. Arch. Pharm. Res. 2010, 33, 557–565. [Google Scholar] [CrossRef]
- Guo, Y.; Luo, C.; Tu, G.; Li, C.; Liu, Y.; Liu, W.; Lam Yung, K.K.; Mo, Z. Rhynchophylline downregulates phosphorylated cAMP response element binding protein, nuclear receptor-related-1, and brain-derived neurotrophic factor expression in the hippocampus of ketamine-induced conditioned place preference rats. Pharmacogn. Mag. 2018, 14, 81–86. [Google Scholar] [CrossRef]
- Yoo, J.H.; Ha, T.W.; Hong, J.T.; Oh, K.W. Rhynchophylline, one of major constituents of Uncariae Ramulus et Uncus enhances pentobarbital-induced sleep behaviors and Rapid Eye Movement Sleep in rodents. Nat. Prod. Sci. 2016, 22, 263–269. [Google Scholar] [CrossRef]
- Frank, M.G.; Heller, H.C. The function(s) of sleep. Handb. Exp. Pharmacol. 2019, 253, 3–34. [Google Scholar] [CrossRef] [PubMed]
- Kempf, A.; Song, S.M.; Talbot, C.B.; Miesenbock, G. A potassium channel beta-subunit couples mitochondrial electron transport to sleep. Nature 2019, 568, 230–234. [Google Scholar] [CrossRef]
- Vyazovskiy, V.V.; Walton, M.E.; Peirson, S.N.; Bannerman, D.M. Sleep homeostasis, habits and habituation. Curr. Opin. Neurobiol. 2017, 44, 202–211. [Google Scholar] [CrossRef]
- Boyce, R.; Williams, S.; Adamantidis, A. REM sleep and memory. Curr. Opin. Neurobiol. 2017, 44, 167–177. [Google Scholar] [CrossRef]
- Timofeev, I.; Chauvette, S. Sleep slow oscillation and plasticity. Curr. Opin. Neurobiol. 2017, 44, 116–126. [Google Scholar] [CrossRef]
- Irwin, M.R. Sleep and inflammation: Partners in sickness and in health. Nat. Rev. Immunol. 2019, 19, 702–715. [Google Scholar] [CrossRef]
- Abel, T.; Havekes, R.; Saletin, J.M.; Walker, M.P. Sleep, plasticity and memory from molecules to whole-brain networks. Curr. Biol. 2013, 23, R774–R788. [Google Scholar] [CrossRef]
- Headley, D.B.; Pare, D. Common oscillatory mechanisms across multiple memory systems. NPJ Sci. Learn. 2017, 2. [Google Scholar] [CrossRef] [PubMed]
- Franken, P.; Dijk, D.J.; Tobler, I.; Borbely, A.A. Sleep deprivation in rats: Effects on EEG power spectra, vigilance states, and cortical temperature. Am. J. Physiol. 1991, 261, R198–R208. [Google Scholar] [CrossRef] [PubMed]
- Hubbard, J.; Gent, T.C.; Hoekstra, M.M.B.; Emmenegger, Y.; Mongrain, V.; Landolt, H.P.; Adamantidis, A.R.; Franken, P. Rapid fast-delta decay following prolonged wakefulness marks a phase of wake-inertia in NREM sleep. Nat. Commun. 2020, 11, 3130. [Google Scholar] [CrossRef] [PubMed]
- Montgomery, S.M.; Sirota, A.; Buzsaki, G. Theta and gamma coordination of hippocampal networks during waking and rapid eye movement sleep. J. Neurosci. 2008, 28, 6731–6741. [Google Scholar] [CrossRef]
- Steriade, M.; McCormick, D.A.; Sejnowski, T.J. Thalamocortical oscillations in the sleeping and aroused brain. Science 1993, 262, 679–685. [Google Scholar] [CrossRef]
- Steriade, M.; Timofeev, I.; Grenier, F. Natural waking and sleep states: A view from inside neocortical neurons. J. Neurophysiol. 2001, 85, 1969–1985. [Google Scholar] [CrossRef]
- Borbely, A.A. A two process model of sleep regulation. Hum. Neurobiol. 1982, 1, 195–204. [Google Scholar] [PubMed]
- Daan, S.; Beersma, D.G.; Borbely, A.A. Timing of human sleep: Recovery process gated by a circadian pacemaker. Am. J. Physiol. 1984, 246, R161–R183. [Google Scholar] [CrossRef] [PubMed]
- Dijk, D.J.; Czeisler, C.A. Contribution of the circadian pacemaker and the sleep homeostat to sleep propensity, sleep structure, electroencephalographic slow waves, and sleep spindle activity in humans. J. Neurosci. 1995, 15, 3526–3538. [Google Scholar] [CrossRef]
- Jones, B.E. Arousal and sleep circuits. Neuropsychopharmacology 2020, 45, 6–20. [Google Scholar] [CrossRef]
- Saper, C.B.; Scammell, T.E.; Lu, J. Hypothalamic regulation of sleep and circadian rhythms. Nature 2005, 437, 1257–1263. [Google Scholar] [CrossRef]
- Adamantidis, A.R.; Zhang, F.; Aravanis, A.M.; Deisseroth, K.; de Lecea, L. Neural substrates of awakening probed with optogenetic control of hypocretin neurons. Nature 2007, 450, 420–424. [Google Scholar] [CrossRef]
- Carter, M.E.; Yizhar, O.; Chikahisa, S.; Nguyen, H.; Adamantidis, A.; Nishino, S.; Deisseroth, K.; de Lecea, L. Tuning arousal with optogenetic modulation of locus coeruleus neurons. Nat. Neurosci. 2010, 13, 1526–1533. [Google Scholar] [CrossRef]
- Han, Y.; Shi, Y.F.; Xi, W.; Zhou, R.; Tan, Z.B.; Wang, H.; Li, X.M.; Chen, Z.; Feng, G.; Luo, M.; et al. Selective activation of cholinergic basal forebrain neurons induces immediate sleep-wake transitions. Curr. Biol. 2014, 24, 693–698. [Google Scholar] [CrossRef]
- Kroeger, D.; Ferrari, L.L.; Petit, G.; Mahoney, C.E.; Fuller, P.M.; Arrigoni, E.; Scammell, T.E. Cholinergic, glutamatergic, and GABAergic neurons of the pedunculopontine tegmental nucleus have distinct effects on sleep/wake behavior in mice. J. Neurosci. 2017, 37, 1352–1366. [Google Scholar] [CrossRef] [PubMed]
- Smith, H.R.; Leibold, N.K.; Rappoport, D.A.; Ginapp, C.M.; Purnell, B.S.; Bode, N.M.; Alberico, S.L.; Kim, Y.C.; Audero, E.; Gross, C.T.; et al. Dorsal raphe serotonin neurons mediate CO2-induced arousal from sleep. J. Neurosci. 2018, 38, 1915–1925. [Google Scholar] [CrossRef] [PubMed]
- Boucetta, S.; Cisse, Y.; Mainville, L.; Morales, M.; Jones, B.E. Discharge profiles across the sleep-waking cycle of identified cholinergic, GABAergic, and glutamatergic neurons in the pontomesencephalic tegmentum of the rat. J. Neurosci. 2014, 34, 4708–4727. [Google Scholar] [CrossRef]
- Chung, S.; Weber, F.; Zhong, P.; Tan, C.L.; Nguyen, T.N.; Beier, K.T.; Hormann, N.; Chang, W.C.; Zhang, Z.; Do, J.P.; et al. Identification of preoptic sleep neurons using retrograde labelling and gene profiling. Nature 2017, 545, 477–481. [Google Scholar] [CrossRef]
- Van Dort, C.J.; Zachs, D.P.; Kenny, J.D.; Zheng, S.; Goldblum, R.R.; Gelwan, N.A.; Ramos, D.M.; Nolan, M.A.; Wang, K.; Weng, F.J.; et al. Optogenetic activation of cholinergic neurons in the PPT or LDT induces REM sleep. Proc. Natl. Acad. Sci. USA 2015, 112, 584–589. [Google Scholar] [CrossRef] [PubMed]
- Luppi, P.H.; Billwiller, F.; Fort, P. Selective activation of a few limbic structures during paradoxical (REM) sleep by the claustrum and the supramammillary nucleus: Evidence and function. Curr. Opin. Neurobiol. 2017, 44, 59–64. [Google Scholar] [CrossRef] [PubMed]
- Shinno, H.; Kamei, M.; Nakamura, Y.; Inami, Y.; Horiguchi, J. Successful treatment with Yi-Gan San for rapid eye movement sleep behavior disorder. Prog. Neuropsychopharmacol. Biol. Psychiatry 2008, 32, 1749–1751. [Google Scholar] [CrossRef] [PubMed]
- Matsui, K.; Sasai-Sakuma, T.; Ishigooka, J.; Nishimura, K.; Inoue, Y. Effect of Yokukansan for the treatment of idiopathic rapid eye movement sleep behavior disorder: A retrospective analysis of consecutive patients. J. Clin. Sleep Med. 2019, 15, 1173–1178. [Google Scholar] [CrossRef]
- Ozone, M.; Shimazaki, H.; Ichikawa, H.; Shigeta, M. Efficacy of yokukansan compared with clonazepam for rapid eye movement sleep behaviour disorder: A preliminary retrospective study. Psychogeriatrics 2020. [Google Scholar] [CrossRef] [PubMed]
- Ohtomo, Y.; Umino, D.; Nijama, S.; Fujinaga, S.; Shimizu, T. Yokukansan: A treatment option for nocturnal enuresis in children by improving sleep quality. Juntendo Med. J. 2014, 60, 536–542. [Google Scholar] [CrossRef]
- Aizawa, R.; Kanbayashi, T.; Saito, Y.; Ogawa, Y.; Sugiyama, T.; Kitajima, T.; Kaneko, Y.; Abe, M.; Shimizu, T. Effects of Yoku-kan-san-ka-chimpi-hange on the sleep of normal healthy adult subjects. Psychiatry Clin. Neurosci. 2002, 56, 303–304. [Google Scholar] [CrossRef]
- Pan, W.; Kwak, S.; Li, G.; Chen, Y.; Cai, D. Therapeutic effect of Yang-Xue-Qing-Nao granules on sleep dysfunction in Parkinson’s disease. Chin. Med. 2013, 8, 14. [Google Scholar] [CrossRef]
- Sun, Y.Z.; Liu, R. Therapeutic evaluation on needling method of regulating the conception vessel and calming the mind for perimenopausal sleep disorder. J. Acupunct. Tuina Sci. 2013, 11, 142–146. [Google Scholar] [CrossRef]
- Egashira, N.; Nogami, A.; Iwasaki, K.; Ishibashi, A.; Uchida, N.; Takasaki, K.; Mishima, K.; Nishimura, R.; Oishi, R.; Fujiwara, M. Yokukansan enhances pentobarbital-induced sleep in socially isolated mice: Possible involvement of GABA(A)-benzodiazepine receptor complex. J. Pharmacol. Sci. 2011, 116, 316–320. [Google Scholar] [CrossRef]
- Murata, K.; Li, F.; Shinguchi, K.; Ogata, M.; Fujita, N.; Takahashi, R. Yokukansankachimpihange improves the social isolation-induced sleep disruption and allopregnanolone reduction in mice. Front. Nutr. 2020, 7, 8. [Google Scholar] [CrossRef]
- Nagao, M.; Takasaki, K.; Nogami, A.; Hirai, Y.; Moriyama, H.; Uchida, N.; Kubota, K.; Katsurabayashi, S.; Mishima, K.; Nishimura, R.; et al. Effect of Yokukansan on sleep disturbance in a rat model of cerebrovascular dementia. Tradit. Kampo Med. 2014, 1, 19–26. [Google Scholar] [CrossRef]
- Jeenapongsa, R.; Tohda, M. Effects of Choto-san and Chotoko on thiopental-induced sleeping time. J. Tradit. Med. 2003, 20, 165–167. [Google Scholar]
- Shi, J.S.; Huang, B.; Wu, Q.; Ren, R.X.; Xie, X.L. Effects of rhynchophylline on motor activity of mice and serotonin and dopamine in rat brain. Zhongguo Yao Li Xue Bao 1993, 14, 114–117. [Google Scholar]
- Sakakibara, I.; Terabayashi, S.; Kubo, M.; Higuchi, M.; Komatsu, Y.; Okada, M.; Taki, K.; Kamei, J. Effect on locomotion of indole alkaloids from the hooks of uncaria plants. Phytomedicine 1999, 6, 163–168. [Google Scholar] [CrossRef]
- Quílez, A.; Saenz, M.T.; García, M.D. Uncaria tomentosa (Willd. ex. Roem. & Schult.) DC. and Eucalyptus globulus Labill. interactions when administered with diazepam. Phytother. Res. 2012, 26, 458–461. [Google Scholar]
- Chou, C.H.; Gong, C.L.; Chao, C.C.; Lin, C.H.; Kwan, C.Y.; Hsieh, C.L.; Leung, Y.M. Rhynchophylline from Uncaria rhynchophylla functionally turns delayed rectifiers into A-Type K+ channels. J. Nat. Prod. 2009, 72, 830–834. [Google Scholar] [CrossRef]
- Kang, T.H.; Murakami, Y.; Takayama, H.; Kitajima, M.; Aimi, N.; Watanabe, H.; Matsumoto, K. Protective effect of rhynchophylline and isorhynchophylline on in vitro ischemia-induced neuronal damage in the hippocampus: Putative neurotransmitter receptors involved in their action. Life Sci. 2004, 76, 331–343. [Google Scholar] [CrossRef] [PubMed]
- Kai, L.; Wang, Z.F.; Xue, C.H. Effects of Rhynchophylline on L-type calcium channels in isolated rat cortical neurons during acute hypoxia. J. Chin. Pharm. Sci. 1998, 7, 205–208. [Google Scholar]
- Wang, X.L.; Zhang, L.M.; Hua, Z. Blocking effect of rhynchophylline on calcium channels in isolated rat ventricular myocytes. Zhongguo Yao Li Xue Bao 1994, 15, 115–118. [Google Scholar] [PubMed]
- Kang, T.H.; Murakami, Y.; Matsumoto, K.; Takayama, H.; Kitajima, M.; Aimi, N.; Watanabe, H. Rhynchophylline and isorhynchophylline inhibit NMDA receptors expressed in Xenopus oocytes. Eur. J. Pharmacol. 2002, 455, 27–34. [Google Scholar] [CrossRef]
- Li, P.Y.; Zeng, X.R.; Cheng, J.; Wen, J.; Inoue, I.; Yang, Y. Rhynchophylline-induced vasodilation in human mesenteric artery is mainly due to blockage of L-type calcium channels in vascular smooth muscle cells. Naunyn Schmiedebergs Arch. Pharmacol. 2013, 386, 973–982. [Google Scholar] [CrossRef] [PubMed]
- Qin, Q.J.; Cui, L.Q.; Li, P.; Wang, Y.B.; Zhang, X.Z.; Guo, M.L. Rhynchophylline ameliorates myocardial ischemia/reperfusion injury through the modulation of mitochondrial mechanisms to mediate myocardial apoptosis. Mol. Med. Rep. 2019, 19, 2581–2590. [Google Scholar] [CrossRef]
- Song, Y.; Qu, R.; Zhu, S.; Zhang, R.; Ma, S. Rhynchophylline attenuates LPS-induced pro-inflammatory responses through down-regulation of MAPK/NF-kappaB signaling pathways in primary microglia. Phytother. Res. 2012, 26, 1528–1533. [Google Scholar] [CrossRef]
- He, Y.; Zeng, S.Y.; Zhou, S.W.; Qian, G.S.; Peng, K.; Mo, Z.X.; Zhou, J.Y. Effects of rhynchophylline on GluN1 and GluN2B expressions in primary cultured hippocampal neurons. Fitoterapia 2014, 98, 166–173. [Google Scholar] [CrossRef] [PubMed]
- Hao, H.F.; Liu, L.M.; Pan, C.S.; Wang, C.S.; Gao, Y.S.; Fan, J.Y.; Han, J.Y. Rhynchophylline ameliorates endothelial dysfunction via Src-PI3K/Akt-eNOS cascade in the cultured intrarenal arteries of spontaneous hypertensive rats. Front. Physiol. 2017, 8, 928. [Google Scholar] [CrossRef]
- Li, Q.; Niu, C.; Zhang, X.; Dong, M. Gastrodin and Isorhynchophylline synergistically inhibit MPP(+)-induced oxidative stress in SH-SY5Y cells by targeting ERK1/2 and GSK-3beta pathways: Involvement of Nrf2 nuclear translocation. ACS Chem. Neurosci. 2018, 9, 482–493. [Google Scholar] [CrossRef] [PubMed]
- Hu, S.; Mak, S.; Zuo, X.; Li, H.; Wang, Y.; Han, Y. Neuroprotection against MPP(+)-induced cytotoxicity through the activation of PI3-K/Akt/GSK3beta/MEF2D signaling pathway by Rhynchophylline, the major tetracyclic oxindole alkaloid isolated from Uncaria rhynchophylla. Front. Pharmacol. 2018, 9, 768. [Google Scholar] [CrossRef] [PubMed]
- Yuan, D.; Ma, B.; Yang, J.Y.; Xie, Y.Y.; Wang, L.; Zhang, L.J.; Kano, Y.; Wu, C.F. Anti-inflammatory effects of rhynchophylline and isorhynchophylline in mouse N9 microglial cells and the molecular mechanism. Int. Immunopharmacol. 2009, 9, 1549–1554. [Google Scholar] [CrossRef]
- Zhou, J.Y.; Chen, J.; Zhou, S.W.; Mo, Z.X. Individual and combined effects of rhynchophylline and ketamine on proliferation, NMDAR1 and GluA2/3 protein expression in PC12 cells. Fitoterapia 2013, 85, 125–129. [Google Scholar] [CrossRef] [PubMed]
- Kaneko, Y.; Coats, A.B.; Tuazon, J.P.; Jo, M.; Borlongan, C.V. Rhynchophylline promotes stem cell autonomous metabolic homeostasis. Cytotherapy 2020, 22, 106–113. [Google Scholar] [CrossRef] [PubMed]
- Zheng, M.; Chen, M.; Wang, W.; Zhou, M.; Liu, C.; Fan, Y.; Shi, D. Protection by rhynchophylline against MPTP/MPP(+)-induced neurotoxicity via regulating PI3K/Akt pathway. J. Ethnopharmacol. 2021, 268, 113568. [Google Scholar] [CrossRef]
- Shao, H.; Mi, Z.; Ji, W.G.; Zhang, C.H.; Zhang, T.; Ren, S.C.; Zhu, Z.R. Rhynchophylline protects against the amyloid beta-induced increase of spontaneous discharges in the hippocampal CA1 region of rats. Neurochem. Res. 2015, 40, 2365–2373. [Google Scholar] [CrossRef]
- Lu, Y.F.; Xie, X.L.; Wu, Q.; Wen, G.R.; Yang, S.F.; Shi, J.S. Effects of rhynchophylline on monoamine transmitter contents of striatum and hippocampus in cerebral ischemic rats. Chin. J. Pharmacol. Toxicol. 2004, 18, 253–258. [Google Scholar]
- Lai, T.; Chen, L.; Chen, X.; He, J.; Lv, P.; Ge, H. Rhynchophylline attenuates migraine in trigeminal nucleus caudalis in nitroglycerin-induced rat model by inhibiting MAPK/NF-kB signaling. Mol. Cell. Biochem. 2019, 461, 205–212. [Google Scholar] [CrossRef]
- Yang, Y.; Ji, W.G.; Zhu, Z.R.; Wu, Y.L.; Zhang, Z.Y.; Qu, S.C. Rhynchophylline suppresses soluble Abeta1-42-induced impairment of spatial cognition function via inhibiting excessive activation of extrasynaptic NR2B-containing NMDA receptors. Neuropharmacology 2018, 135, 100–112. [Google Scholar] [CrossRef] [PubMed]
- Cao, W.; Wang, Y.; Lv, X.; Yu, X.; Li, X.; Li, H.; Wang, Y.; Lu, D.; Qi, R.; Wang, H. Rhynchophylline prevents cardiac dysfunction and improves survival in lipopolysaccharide-challenged mice via suppressing macrophage I-kappaBalpha phosphorylation. Int. Immunopharmacol. 2012, 14, 243–251. [Google Scholar] [CrossRef] [PubMed]
- Ho, T.Y.; Tang, N.Y.; Hsiang, C.Y.; Hsieh, C.L. Uncaria rhynchophylla and rhynchophylline improved kainic acid-induced epileptic seizures via IL-1beta and brain-derived neurotrophic factor. Phytomedicine 2014, 21, 893–900. [Google Scholar] [CrossRef]
- Hsieh, C.L.; Ho, T.Y.; Su, S.Y.; Lo, W.Y.; Liu, C.H.; Tang, N.Y. Uncaria rhynchophylla and Rhynchophylline inhibit c-Jun N-terminal kinase phosphorylation and nuclear factor-kappaB activity in kainic acid-treated rats. Am. J. Chin. Med. 2009, 37, 351–360. [Google Scholar] [CrossRef] [PubMed]
- Hsu, H.C.; Tang, N.Y.; Liu, C.H.; Hsieh, C.L. Antiepileptic effect of Uncaria rhynchophylla and Rhynchophylline involved in the initiation of c-Jun N-terminal kinase phosphorylation of MAPK signal pathways in acute seizures of kainic acid-treated rats. Evid. Based Complement. Altern. Med. 2013, 2013, 961289. [Google Scholar] [CrossRef]
- Jiang, M.; Chen, Y.; Li, C.; Peng, Q.; Fang, M.; Liu, W.; Kang, Q.; Lin, Y.; Yung, K.K.; Mo, Z. Inhibiting effects of rhynchophylline on zebrafish methamphetamine dependence are associated with amelioration of neurotransmitters content and down-regulation of TH and NR2B expression. Prog. Neuropsychopharmacol. Biol. Psychiatry 2016, 68, 31–43. [Google Scholar] [CrossRef]
- Zhu, C.; Liu, W.; Li, J.; Chen, Z.J.; Li, C.; Zhou, Y.T.; Mo, Z.X. Rhynchophylline reverses methamphetamine-induced CPP by regulating GluR1 expression in zebrafish. Chin. Pharmacol. Bull. 2019, 35, 620–623. [Google Scholar]
- Liu, W.; Peng, Q.X.; Lin, X.L.; Luo, C.H.; Jiang, M.J.; Mo, Z.X.; Yung, K.K. Effect of rhynchophylline on the expression of p-CREB and sc-Fos in striatum and hippocampal CA1 area of methamphetamine-induced conditioned place preference rats. Fitoterapia 2014, 92, 16–22. [Google Scholar] [CrossRef]
- Zhou, J.Y.; Mo, Z.X.; Zhou, S.W. Effect of rhynchophylline on central neurotransmitter levels in amphetamine-induced conditioned place preference rat brain. Fitoterapia 2010, 81, 844–848. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Tu, G.; Luo, C.; Guo, Y.; Fang, M.; Zhu, C.; Li, H.; Ou, J.; Zhou, Y.; Liu, W.; et al. Effects of rhynchophylline on the hippocampal miRNA expression profile in ketamine-addicted rats. Prog. Neuropsychopharmacol. Biol. Psychiatry 2018, 86, 379–389. [Google Scholar] [CrossRef]
- Li, J.; Liu, W.; Peng, Q.; Jiang, M.; Luo, C.; Guo, Y.; Liu, Y.; Fang, M.; Mo, Z. Effect of rhynchophylline on conditioned place preference on expression of NR2B in methamphetamine-dependent mice. Biochem. Biophys Res. Commun. 2014, 452, 695–700. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.; Zhong, R.; Xia, Z.; Song, J.; Feng, L. Neuroprotective effects of rhynchophylline against ischemic brain injury via regulation of the Akt/mTOR and TLRs signaling pathways. Molecules 2014, 19, 11196–11210. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.W.; Hsieh, C.L. Oral Uncaria rhynchophylla (UR) reduces kainic acid-induced epileptic seizures and neuronal death accompanied by attenuating glial cell proliferation and S100B proteins in rats. J. Ethnopharmacol. 2011, 135, 313–320. [Google Scholar] [CrossRef]
- Long, H.; Ruan, J.; Zhang, M.; Wang, C.; Huang, Y. Rhynchophylline attenuates Tourette Syndrome via BDNF/NF-kappaB pathway in vivo and in vitro. Neurotox Res. 2019, 36, 756–763. [Google Scholar] [CrossRef]
- Long, H.; Zhang, M.; Wang, C.; Hang, Y. Rhynchophylline attenuates neurotoxicity in Tourette Syndrome rats. Neurotox. Res. 2019, 36, 679–687. [Google Scholar] [CrossRef]
- Xian, Y.F.; Mao, Q.Q.; Wu, J.C.; Su, Z.R.; Chen, J.N.; Lai, X.P.; Ip, S.P.; Lin, Z.X. Isorhynchophylline treatment improves the amyloid-beta-induced cognitive impairment in rats via inhibition of neuronal apoptosis and tau protein hyperphosphorylation. J. Alzheimers Dis. 2014, 39, 331–346. [Google Scholar] [CrossRef]
- Xian, Y.F.; Ip, S.P.; Li, H.Q.; Qu, C.; Su, Z.R.; Chen, J.N.; Lin, Z.X. Isorhynchophylline exerts antidepressant-like effects in mice via modulating neuroinflammation and neurotrophins: Involvement of the PI3K/Akt/GSK-3beta signaling pathway. FASEB J. 2019, 33, 10393–10408. [Google Scholar] [CrossRef]
- Wang, M.; Li, H.; Zhao, Y.; Lv, C.; Zhou, G. Rhynchophylline attenuates allergic bronchial asthma by inhibiting transforming growth factor-beta1-mediated Smad and mitogen-activated protein kinase signaling transductions in vivo and in vitro. Exp. Ther. Med. 2019, 17, 251–259. [Google Scholar] [CrossRef]
- Zhang, Y.; Cui, Y.; Dai, S.; Deng, W.; Wang, H.; Qin, W.; Yang, H.; Liu, H.; Yue, J.; Wu, D.; et al. Isorhynchophylline enhances Nrf2 and inhibits MAPK pathway in cardiac hypertrophy. Naunyn Schmiedebergs Arch. Pharmacol. 2020, 393, 203–212. [Google Scholar] [CrossRef]
- Lipscombe, D.; Helton, T.D.; Xu, W. L-type calcium channels: The low down. J. Neurophysiol. 2004, 92, 2633–2641. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.B.; Chen, C.X.; Sim, S.M.; Kwan, C.Y. In vitro vasodilator mechanisms of the indole alkaloids rhynchophylline and isorhynchophylline, isolated from the hook of Uncaria rhynchophylla (Miquel). Naunyn Schmiedebergs Arch. Pharmacol. 2004, 369, 232–238. [Google Scholar] [CrossRef]
- Zhang, W.; Liu, G.X.; Huang, X.N. Effect of rhynchophylline on the contraction of rabbit aorta. Zhongguo Yao Li Xue Bao 1987, 8, 425–429. [Google Scholar] [PubMed]
- Wiera, G.; Nowak, D.; van Hove, I.; Dziegiel, P.; Moons, L.; Mozrzymas, J.W. Mechanisms of NMDA receptor- and voltage-gated L-Type calcium channel-dependent hippocampal LTP critically rely on proteolysis that is mediated by distinct metalloproteinases. J. Neurosci. 2017, 37, 1240–1256. [Google Scholar] [CrossRef] [PubMed]
- Kumar, M.; John, M.; Madhavan, M.; James, J.; Omkumar, R.V. Alteration in the phosphorylation status of NMDA receptor GluN2B subunit by activation of both NMDA receptor and L-type voltage gated calcium channel. Neurosci. Lett. 2019, 709, 134343. [Google Scholar] [CrossRef] [PubMed]
- Zamponi, G.W. Targeting voltage-gated calcium channels in neurological and psychiatric diseases. Nat. Rev. Drug Discov. 2016, 15, 19–34. [Google Scholar] [CrossRef]
- Lacinova, L.; Moosmang, S.; Langwieser, N.; Hofmann, F.; Kleppisch, T. Cav1.2 calcium channels modulate the spiking pattern of hippocampal pyramidal cells. Life Sci. 2008, 82, 41–49. [Google Scholar] [CrossRef] [PubMed]
- Hansen, A.K.; Nedergaard, S.; Andreasen, M. Intrinsic Ca2+-dependent theta oscillations in apical dendrites of hippocampal CA1 pyramidal cells in vitro. J. Neurophysiol. 2014, 112, 631–643. [Google Scholar] [CrossRef] [PubMed]
- Plumbly, W.; Brandon, N.; Deeb, T.Z.; Hall, J.; Harwood, A.J. L-type voltage-gated calcium channel regulation of in vitro human cortical neuronal networks. Sci. Rep. 2019, 9, 13810. [Google Scholar] [CrossRef] [PubMed]
- Kabir, Z.D.; Che, A.; Fischer, D.K.; Rice, R.C.; Rizzo, B.K.; Byrne, M.; Glass, M.J.; De Marco Garcia, N.V.; Rajadhyaksha, A.M. Rescue of impaired sociability and anxiety-like behavior in adult cacna1c-deficient mice by pharmacologically targeting eIF2alpha. Mol. Psychiatry 2017, 22, 1096–1109. [Google Scholar] [CrossRef]
- Ode, K.L.; Katsumata, T.; Tone, D.; Ueda, H.R. Fast and slow Ca(2+)-dependent hyperpolarization mechanisms connect membrane potential and sleep homeostasis. Curr. Opin. Neurobiol. 2017, 44, 212–221. [Google Scholar] [CrossRef] [PubMed]
- Bazhenov, M.; Timofeev, I.; Steriade, M.; Sejnowski, T.J. Model of thalamocortical slow-wave sleep oscillations and transitions to activated States. J. Neurosci. 2002, 22, 8691–8704. [Google Scholar] [CrossRef] [PubMed]
- Sinnegger-Brauns, M.J.; Huber, I.G.; Koschak, A.; Wild, C.; Obermair, G.J.; Einzinger, U.; Hoda, J.C.; Sartori, S.B.; Striessnig, J. Expression and 1,4-dihydropyridine-binding properties of brain L-type calcium channel isoforms. Mol. Pharmacol. 2009, 75, 407–414. [Google Scholar] [CrossRef] [PubMed]
- Kumar, D.; Dedic, N.; Flachskamm, C.; Voule, S.; Deussing, J.M.; Kimura, M. Cacna1c (Cav1.2) modulates electroencephalographic rhythm and rapid eye movement sleep recovery. Sleep 2015, 38, 1371–1380. [Google Scholar] [CrossRef]
- Kantojarvi, K.; Liuhanen, J.; Saarenpaa-Heikkila, O.; Satomaa, A.L.; Kylliainen, A.; Polkki, P.; Jaatela, J.; Toivola, A.; Milani, L.; Himanen, S.L.; et al. Variants in calcium voltage-gated channel subunit Alpha1 C-gene (CACNA1C) are associated with sleep latency in infants. PLoS ONE 2017, 12, e0180652. [Google Scholar] [CrossRef]
- Schmutz, I.; Chavan, R.; Ripperger, J.A.; Maywood, E.S.; Langwieser, N.; Jurik, A.; Stauffer, A.; Delorme, J.E.; Moosmang, S.; Hastings, M.H.; et al. A specific role for the REV-ERBalpha-controlled L-Type Voltage-Gated Calcium Channel CaV1.2 in resetting the circadian clock in the late night. J. Biol. Rhythm 2014, 29, 288–298. [Google Scholar] [CrossRef] [PubMed]
- Manis, P.B. Delayed rectifier and A-Type potassium channels. In Encyclopedia of Computational Neuroscience; Jaeger, D., Jung, R., Eds.; Springer: New York, NY, USA, 2015; pp. 971–985. [Google Scholar]
- Sheng, M.; Tsaur, M.L.; Jan, Y.N.; Jan, L.Y. Contrasting subcellular localization of the Kv1.2 K+ channel subunit in different neurons of rat brain. J. Neurosci. 1994, 14, 2408–2417. [Google Scholar] [CrossRef]
- Tsaur, M.L.; Sheng, M.; Lowenstein, D.H.; Jan, Y.N.; Jan, L.Y. Differential expression of K+ channel mRNAs in the rat brain and down-regulation in the hippocampus following seizures. Neuron 1992, 8, 1055–1067. [Google Scholar] [CrossRef]
- Espinosa, F.; Marks, G.; Heintz, N.; Joho, R.H. Increased motor drive and sleep loss in mice lacking Kv3-type potassium channels. Genes Brain Behav. 2004, 3, 90–100. [Google Scholar] [CrossRef]
- Douglas, C.L.; Vyazovskiy, V.; Southard, T.; Chiu, S.Y.; Messing, A.; Tononi, G.; Cirelli, C. Sleep in Kcna2 knockout mice. BMC Biol. 2007, 5, 42. [Google Scholar] [CrossRef] [PubMed]
- Vyazovskiy, V.V.; Deboer, T.; Rudy, B.; Lau, D.; Borbely, A.A.; Tobler, I. Sleep EEG in mice that are deficient in the potassium channel subunit K.v.3.2. Brain Res. 2002, 947, 204–211. [Google Scholar] [CrossRef]
- Douglas, C.L.; Vyazovskiy, V.; Southard, T.; Faraguna, U.; Cirelli, C.; Tononi, G. Voltage-dependent potassium channel Kv1.2: Effects on sleep and EEG power spectrum of intracortical injections of an anti-Kv1.2 antibody. Sleep 2006, 29, A36. [Google Scholar]
- Bushey, D.; Huber, R.; Tononi, G.; Cirelli, C. Drosophila Hyperkinetic mutants have reduced sleep and impaired memory. J. Neurosci. 2007, 27, 5384–5393. [Google Scholar] [CrossRef]
- Tatsuki, F.; Sunagawa, G.A.; Shi, S.; Susaki, E.A.; Yukinaga, H.; Perrin, D.; Sumiyama, K.; Ukai-Tadenuma, M.; Fujishima, H.; Ohno, R.; et al. Involvement of Ca(2+)-dependent hyperpolarization in sleep duration in mammals. Neuron 2016, 90, 70–85. [Google Scholar] [CrossRef] [PubMed]
- Malinow, R.; Malenka, R.C. AMPA receptor trafficking and synaptic plasticity. Annu. Rev. Neurosci. 2002, 25, 103–126. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Wong, T.P.; Aarts, M.; Rooyakkers, A.; Liu, L.; Lai, T.W.; Wu, D.C.; Lu, J.; Tymianski, M.; Craig, A.M.; et al. NMDA receptor subunits have differential roles in mediating excitotoxic neuronal death both in vitro and in vivo. J. Neurosci. 2007, 27, 2846–2857. [Google Scholar] [CrossRef]
- Xu, J.; Kurup, P.; Zhang, Y.; Goebel-Goody, S.M.; Wu, P.H.; Hawasli, A.H.; Baum, M.L.; Bibb, J.A.; Lombroso, P.J. Extrasynaptic NMDA receptors couple preferentially to excitotoxicity via calpain-mediated cleavage of STEP. J. Neurosci. 2009, 29, 9330–9343. [Google Scholar] [CrossRef]
- Yin, D.; Dong, H.; Wang, T.X.; Hu, Z.Z.; Cheng, N.N.; Qu, W.M.; Huang, Z.L. Glutamate activates the histaminergic tuberomammillary nucleus and increases wakefulness in rats. Neuroscience 2019, 413, 86–98. [Google Scholar] [CrossRef] [PubMed]
- Manfridi, A.; Brambilla, D.; Mancia, M. Stimulation of NMDA and AMPA receptors in the rat nucleus basalis of Meynert affects sleep. Am. J. Physiol. 1999, 277, R1488–R1492. [Google Scholar] [CrossRef]
- Datta, S.; Siwek, D.F. Excitation of the brain stem pedunculopontine tegmentum cholinergic cells induces wakefulness and REM sleep. J. Neurophysiol. 1997, 77, 2975–2988. [Google Scholar] [CrossRef] [PubMed]
- Datta, S.; Spoley, E.E.; Patterson, E.H. Microinjection of glutamate into the pedunculopontine tegmentum induces REM sleep and wakefulness in the rat. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2001, 280, R752–R759. [Google Scholar] [CrossRef] [PubMed]
- Campbell, I.G.; Feinberg, I. NREM delta stimulation following MK-801 is a response of sleep systems. J. Neurophysiol. 1996, 76, 3714–3720. [Google Scholar] [CrossRef]
- Campbell, I.G.; Feinberg, I. Comparison of MK-801 and sleep deprivation effects on NREM, REM, and waking spectra in the rat. Sleep 1999, 22, 423–432. [Google Scholar] [CrossRef] [PubMed]
- Ishikawa, I.; Shinno, H.; Ando, N.; Mori, T.; Nakamura, Y. The effect of memantine on sleep architecture and psychiatric symptoms in patients with Alzheimer’s disease. Acta Neuropsychiatr. 2016, 28, 157–164. [Google Scholar] [CrossRef]
- Kaushik, M.K.; Kumar, V.M.; Mallick, H.N. Glutamate microinjection at the medial preoptic area enhances slow wave sleep in rats. Behav. Brain Res. 2011, 217, 240–243. [Google Scholar] [CrossRef]
- Mukherjee, D.; Kaushik, M.K.; Jaryal, A.K.; Kumar, V.M.; Mallick, H.N. Glutamate microinjection in the medial septum of rats decreases paradoxical sleep and increases slow wave sleep. Neuroreport 2012, 23, 451–456. [Google Scholar] [CrossRef]
- Kocsis, B. State-dependent increase of cortical gamma activity during REM sleep after selective blockade of NR2B subunit containing NMDA receptors. Sleep 2012, 35, 1011–1016. [Google Scholar] [CrossRef]
- El Gaamouch, F.; Buisson, A.; Moustie, O.; Lemieux, M.; Labrecque, S.; Bontempi, B.; De Koninck, P.; Nicole, O. Interaction between alphaCaMKII and GluN2B controls ERK-dependent plasticity. J. Neurosci. 2012, 32, 10767–10779. [Google Scholar] [CrossRef]
- Krapivinsky, G.; Krapivinsky, L.; Manasian, Y.; Ivanov, A.; Tyzio, R.; Pellegrino, C.; Ben-Ari, Y.; Clapham, D.E.; Medina, I. The NMDA receptor is coupled to the ERK pathway by a direct interaction between NR2B and RasGRF1. Neuron 2003, 40, 775–784. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, W.; Li, D.; Li, M.; Wang, P.; Wen, J.; Liang, M.; Su, B.; Yin, Y. IGF-1 alleviates NMDA-induced excitotoxicity in cultured hippocampal neurons against autophagy via the NR2B/PI3K-AKT-mTOR pathway. J. Cell. Physiol. 2014, 229, 1618–1629. [Google Scholar] [CrossRef]
- Yoshii, A.; Constantine-Paton, M. BDNF induces transport of PSD-95 to dendrites through PI3K-AKT signaling after NMDA receptor activation. Nat. Neurosci. 2007, 10, 702–711. [Google Scholar] [CrossRef] [PubMed]
- Dolmetsch, R.E.; Pajvani, U.; Fife, K.; Spotts, J.M.; Greenberg, M.E. Signaling to the nucleus by an L-type calcium channel-calmodulin complex through the MAP kinase pathway. Science 2001, 294, 333–339. [Google Scholar] [CrossRef] [PubMed]
- Wheeler, D.G.; Groth, R.D.; Ma, H.; Barrett, C.F.; Owen, S.F.; Safa, P.; Tsien, R.W. Ca(V)1 and Ca(V)2 channels engage distinct modes of Ca(2+) signaling to control CREB-dependent gene expression. Cell 2012, 149, 1112–1124. [Google Scholar] [CrossRef]
- Gale, N.W.; Baluk, P.; Pan, L.; Kwan, M.; Holash, J.; DeChiara, T.M.; McDonald, D.M.; Yancopoulos, G.D. Ephrin-B2 selectively marks arterial vessels and neovascularization sites in the adult, with expression in both endothelial and smooth-muscle cells. Dev. Biol. 2001, 230, 151–160. [Google Scholar] [CrossRef]
- Matsuo, K.; Otaki, N. Bone cell interactions through Eph/ephrin: Bone modeling, remodeling and associated diseases. Cell Adh. Migr. 2012, 6, 148–156. [Google Scholar] [CrossRef]
- Stark, D.A.; Karvas, R.M.; Siegel, A.L.; Cornelison, D.D. Eph/ephrin interactions modulate muscle satellite cell motility and patterning. Development 2011, 138, 5279–5289. [Google Scholar] [CrossRef]
- Murai, K.K.; Pasquale, E.B. ‘Eph’ective signaling: Forward, reverse and crosstalk. J. Cell Sci. 2003, 116, 2823–2832. [Google Scholar] [CrossRef]
- Murai, K.K.; Pasquale, E.B. Eph receptors and ephrins in neuron-astrocyte communication at synapses. Glia 2011, 59, 1567–1578. [Google Scholar] [CrossRef] [PubMed]
- Carmona, M.A.; Murai, K.K.; Wang, L.; Roberts, A.J.; Pasquale, E.B. Glial ephrin-A3 regulates hippocampal dendritic spine morphology and glutamate transport. Proc. Natl. Acad. Sci. USA 2009, 106, 12524–12529. [Google Scholar] [CrossRef]
- Fu, A.K.; Hung, K.W.; Fu, W.Y.; Shen, C.; Chen, Y.; Xia, J.; Lai, K.O.; Ip, N.Y. APC(Cdh1) mediates EphA4-dependent downregulation of AMPA receptors in homeostatic plasticity. Nat. Neurosci. 2011, 14, 181–189. [Google Scholar] [CrossRef] [PubMed]
- Fu, W.Y.; Chen, Y.; Sahin, M.; Zhao, X.S.; Shi, L.; Bikoff, J.B.; Lai, K.O.; Yung, W.H.; Fu, A.K.; Greenberg, M.E.; et al. Cdk5 regulates EphA4-mediated dendritic spine retraction through an ephexin1-dependent mechanism. Nat. Neurosci. 2007, 10, 67–76. [Google Scholar] [CrossRef] [PubMed]
- Freyburger, M.; Pierre, A.; Paquette, G.; Belanger-Nelson, E.; Bedont, J.; Gaudreault, P.O.; Drolet, G.; Laforest, S.; Blackshaw, S.; Cermakian, N.; et al. EphA4 is involved in sleep regulation but not in the electrophysiological response to sleep deprivation. Sleep 2016, 39, 613–624. [Google Scholar] [CrossRef]
- Freyburger, M.; Poirier, G.; Carrier, J.; Mongrain, V. Shorter duration of non-rapid eye movement sleep slow waves in EphA4 knockout mice. J. Sleep Res. 2017, 26, 539–546. [Google Scholar] [CrossRef] [PubMed]
- Kiessling, S.; O’Callaghan, E.K.; Freyburger, M.; Cermakian, N.; Mongrain, V. The cell adhesion molecule EphA4 is involved in circadian clock functions. Genes Brain Behav. 2018, 17, 82–92. [Google Scholar] [CrossRef]
- Korte, M.; Carroll, P.; Wolf, E.; Brem, G.; Thoenen, H.; Bonhoeffer, T. Hippocampal long-term potentiation is impaired in mice lacking brain-derived neurotrophic factor. Proc. Natl. Acad. Sci. USA 1995, 92, 8856–8860. [Google Scholar] [CrossRef]
- Kang, H.; Schuman, E.M. Long-lasting neurotrophin-induced enhancement of synaptic transmission in the adult hippocampus. Science 1995, 267, 1658–1662. [Google Scholar] [CrossRef]
- Lu, B. BDNF and activity-dependent synaptic modulation. Learn. Mem. 2003, 10, 86–98. [Google Scholar] [CrossRef]
- Vigers, A.J.; Amin, D.S.; Talley-Farnham, T.; Gorski, J.A.; Xu, B.; Jones, K.R. Sustained expression of brain-derived neurotrophic factor is required for maintenance of dendritic spines and normal behavior. Neuroscience 2012, 212, 1–18. [Google Scholar] [CrossRef]
- Ghosh, A.; Carnahan, J.; Greenberg, M.E. Requirement for BDNF in activity-dependent survival of cortical neurons. Science 1994, 263, 1618–1623. [Google Scholar] [CrossRef]
- Kowianski, P.; Lietzau, G.; Czuba, E.; Waskow, M.; Steliga, A.; Morys, J. BDNF: A key factor with multipotent impact on brain signaling and synaptic plasticity. Cell. Mol. Neurobiol. 2018, 38, 579–593. [Google Scholar] [CrossRef]
- Li, W.; Keifer, J. BDNF-induced synaptic delivery of AMPAR subunits is differentially dependent on NMDA receptors and requires ERK. Neurobiol. Learn. Mem. 2009, 91, 243–249. [Google Scholar] [CrossRef][Green Version]
- Ying, S.W.; Futter, M.; Rosenblum, K.; Webber, M.J.; Hunt, S.P.; Bliss, T.V.; Bramham, C.R. Brain-derived neurotrophic factor induces long-term potentiation in intact adult hippocampus: Requirement for ERK activation coupled to CREB and upregulation of Arc synthesis. J. Neurosci. 2002, 22, 1532–1540. [Google Scholar] [CrossRef]
- Yasuda, M.; Fukuchi, M.; Tabuchi, A.; Kawahara, M.; Tsuneki, H.; Azuma, Y.; Chiba, Y.; Tsuda, M. Robust stimulation of TrkB induces delayed increases in BDNF and Arc mRNA expressions in cultured rat cortical neurons via distinct mechanisms. J. Neurochem. 2007, 103, 626–636. [Google Scholar] [CrossRef]
- Blum, R.; Konnerth, A. Neurotrophin-mediated rapid signaling in the central nervous system: Mechanisms and functions. Physiology (Bethesda) 2005, 20, 70–78. [Google Scholar] [CrossRef] [PubMed]
- Rahmani, M.; Rahmani, F.; Rezaei, N. The brain-derived neurotrophic factor: Missing link between sleep deprivation, insomnia, and depression. Neurochem. Res. 2020, 45, 221–231. [Google Scholar] [CrossRef] [PubMed]
- Monteiro, B.C.; Monteiro, S.; Candida, M.; Adler, N.; Paes, F.; Rocha, N.; Nardi, A.E.; Murillo-Rodriguez, E.; Machado, S. Relationship between brain-derived neurotrofic factor (Bdnf) and sleep on depression: A critical review. Clin. Pract. Epidemiol. Ment. Health 2017, 13, 213–219. [Google Scholar] [CrossRef]
- Schmitt, K.; Holsboer-Trachsler, E.; Eckert, A. BDNF in sleep, insomnia, and sleep deprivation. Ann. Med. 2016, 48, 42–51. [Google Scholar] [CrossRef]
- Kushikata, T.; Fang, J.; Krueger, J.M. Brain-derived neurotrophic factor enhances spontaneous sleep in rats and rabbits. Am. J. Physiol. 1999, 276, R1334–R1338. [Google Scholar] [CrossRef]
- Deuschle, M.; Schredl, M.; Wisch, C.; Schilling, C.; Gilles, M.; Geisel, O.; Hellweg, R. Serum brain-derived neurotrophic factor (BDNF) in sleep-disordered patients: Relation to sleep stage N3 and rapid eye movement (REM) sleep across diagnostic entities. J. Sleep Res. 2018, 27, 73–77. [Google Scholar] [CrossRef] [PubMed]
- Fan, T.T.; Chen, W.H.; Shi, L.; Lin, X.; Tabarak, S.; Chen, S.J.; Que, J.Y.; Bao, Y.P.; Tang, X.D.; Shi, J.; et al. Objective sleep duration is associated with cognitive deficits in primary insomnia: BDNF may play a role. Sleep 2019, 42. [Google Scholar] [CrossRef]
- Watson, A.J.; Henson, K.; Dorsey, S.G.; Frank, M.G. The truncated TrkB receptor influences mammalian sleep. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2015, 308, R199–R207. [Google Scholar] [CrossRef][Green Version]
- Faraguna, U.; Vyazovskiy, V.V.; Nelson, A.B.; Tononi, G.; Cirelli, C. A causal role for brain-derived neurotrophic factor in the homeostatic regulation of sleep. J. Neurosci. 2008, 28, 4088–4095. [Google Scholar] [CrossRef]
- Bachmann, V.; Klein, C.; Bodenmann, S.; Schafer, N.; Berger, W.; Brugger, P.; Landolt, H.P. The BDNF Val66Met polymorphism modulates sleep intensity: EEG frequency- and state-specificity. Sleep 2012, 35, 335–344. [Google Scholar] [CrossRef]
- Guindalini, C.; Mazzotti, D.R.; Castro, L.S.; D’Aurea, C.V.; Andersen, M.L.; Poyares, D.; Bittencourt, L.R.; Tufik, S. Brain-derived neurotrophic factor gene polymorphism predicts interindividual variation in the sleep electroencephalogram. J. Neurosci. Res. 2014, 92, 1018–1023. [Google Scholar] [CrossRef] [PubMed]
- Gosselin, N.; De Beaumont, L.; Gagnon, K.; Baril, A.A.; Mongrain, V.; Blais, H.; Montplaisir, J.; Gagnon, J.F.; Pelleieux, S.; Poirier, J.; et al. BDNF Val66Met polymorphism interacts with sleep consolidation to predict ability to create new declarative memories. J. Neurosci. 2016, 36, 8390–8398. [Google Scholar] [CrossRef] [PubMed]
- Ma, T.; Zhang, H.; Xu, Z.P.; Lu, Y.; Fu, Q.; Wang, W.; Li, G.H.; Wang, Y.Y.; Yang, Y.T.; Mi, W.D. Activation of brain-derived neurotrophic factor signaling in the basal forebrain reverses acute sleep deprivation-induced fear memory impairments. Brain Behav. 2020, 10, e01592. [Google Scholar] [CrossRef] [PubMed]
- Xue, J.; Li, H.; Xu, Z.; Ma, D.; Guo, R.; Yang, K.; Wang, Y. Paradoxical sleep deprivation aggravates and prolongs incision-induced pain hypersensitivity via BDNF signaling-mediated descending facilitation in rats. Neurochem. Res. 2018, 43, 2353–2361. [Google Scholar] [CrossRef] [PubMed]
- Barnes, A.K.; Koul-Tiwari, R.; Garner, J.M.; Geist, P.A.; Datta, S. Activation of brain-derived neurotrophic factor-tropomyosin receptor kinase B signaling in the pedunculopontine tegmental nucleus: A novel mechanism for the homeostatic regulation of rapid eye movement sleep. J. Neurochem. 2017, 141, 111–123. [Google Scholar] [CrossRef]
- Datta, S.; Knapp, C.M.; Koul-Tiwari, R.; Barnes, A. The homeostatic regulation of REM sleep: A role for localized expression of brain-derived neurotrophic factor in the brainstem. Behav. Brain Res. 2015, 292, 381–392. [Google Scholar] [CrossRef]
- Giese, M.; Beck, J.; Brand, S.; Muheim, F.; Hemmeter, U.; Hatzinger, M.; Holsboer-Trachsler, E.; Eckert, A. Fast BDNF serum level increase and diurnal BDNF oscillations are associated with therapeutic response after partial sleep deprivation. J. Psychiatr. Res. 2014, 59, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Giese, M.; Unternahrer, E.; Huttig, H.; Beck, J.; Brand, S.; Calabrese, P.; Holsboer-Trachsler, E.; Eckert, A. BDNF: An indicator of insomnia? Mol. Psychiatry 2014, 19, 151–152. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Bae, C.H.; Park, S.Y.; Lee, S.J.; Kim, Y. Uncaria rhynchophylla inhibits the production of nitric oxide and interleukin-1beta through blocking nuclear factor kappaB, Akt, and mitogen-activated protein kinase activation in macrophages. J. Med. Food 2010, 13, 1133–1140. [Google Scholar] [CrossRef]
- Cao, H.; Ren, W.H.; Zhu, M.Y.; Zhao, Z.Q.; Zhang, Y.Q. Activation of glycine site and GluN2B subunit of NMDA receptors is necessary for ERK/CREB signaling cascade in rostral anterior cingulate cortex in rats: Implications for affective pain. Neurosci. Bull. 2012, 28, 77–87. [Google Scholar] [CrossRef][Green Version]
- Mikhail, C.; Vaucher, A.; Jimenez, S.; Tafti, M. ERK signaling pathway regulates sleep duration through activity-induced gene expression during wakefulness. Sci. Signal. 2017, 10. [Google Scholar] [CrossRef] [PubMed]
- Tian, X.; Gotoh, T.; Tsuji, K.; Lo, E.H.; Huang, S.; Feig, L.A. Developmentally regulated role for Ras-GRFs in coupling NMDA glutamate receptors to Ras, Erk and CREB. EMBO J. 2004, 23, 1567–1575. [Google Scholar] [CrossRef]
- Hu, X.; Paik, P.K.; Chen, J.; Yarilina, A.; Kockeritz, L.; Lu, T.T.; Woodgett, J.R.; Ivashkiv, L.B. IFN-gamma suppresses IL-10 production and synergizes with TLR2 by regulating GSK3 and CREB/AP-1 proteins. Immunity 2006, 24, 563–574. [Google Scholar] [CrossRef]
- Su, X.; Wang, C.; Wang, X.; Han, F.; Lv, C.; Zhang, X. Sweet dream liquid chinese medicine ameliorates learning and memory deficit in a rat model of paradoxical sleep deprivation through the ERK/CREB signaling pathway. J. Med. Food 2016, 19, 472–480. [Google Scholar] [CrossRef]
- Foltenyi, K.; Greenspan, R.J.; Newport, J.W. Activation of EGFR and ERK by rhomboid signaling regulates the consolidation and maintenance of sleep in Drosophila. Nat. Neurosci. 2007, 10, 1160–1167. [Google Scholar] [CrossRef]
- Dumoulin Bridi, M.C.; Aton, S.J.; Seibt, J.; Renouard, L.; Coleman, T.; Frank, M.G. Rapid eye movement sleep promotes cortical plasticity in the developing brain. Sci. Adv. 2015, 1, e1500105. [Google Scholar] [CrossRef] [PubMed]
- Graves, L.A.; Hellman, K.; Veasey, S.; Blendy, J.A.; Pack, A.I.; Abel, T. Genetic evidence for a role of CREB in sustained cortical arousal. J. Neurophysiol. 2003, 90, 1152–1159. [Google Scholar] [CrossRef]
- Wimmer, M.E.; Cui, R.; Blackwell, J.M.; Abel, T. Cyclic AMP response element-binding protein is required in excitatory neurons in the forebrain to sustain wakefulness. Sleep 2020, zsaa267. [Google Scholar] [CrossRef]
- Cirelli, C.; Tononi, G. Differential expression of plasticity-related genes in waking and sleep and their regulation by the noradrenergic system. J. Neurosci. 2000, 20, 9187–9194. [Google Scholar] [CrossRef] [PubMed]
- Cirelli, C.; Pompeiano, M.; Tononi, G. Neuronal gene expression in the waking state: A role for the locus coeruleus. Science 1996, 274, 1211–1215. [Google Scholar] [CrossRef]
- Hendricks, J.C.; Williams, J.A.; Panckeri, K.; Kirk, D.; Tello, M.; Yin, J.C.; Sehgal, A. A non-circadian role for cAMP signaling and CREB activity in Drosophila rest homeostasis. Nat. Neurosci. 2001, 4, 1108–1115. [Google Scholar] [CrossRef]
- Fruman, D.A.; Chiu, H.; Hopkins, B.D.; Bagrodia, S.; Cantley, L.C.; Abraham, R.T. The PI3K pathway in human disease. Cell 2017, 170, 605–635. [Google Scholar] [CrossRef]
- Alessi, D.R.; James, S.R.; Downes, C.P.; Holmes, A.B.; Gaffney, P.R.; Reese, C.B.; Cohen, P. Characterization of a 3-phosphoinositide-dependent protein kinase which phosphorylates and activates protein kinase Balpha. Curr. Biol. 1997, 7, 261–269. [Google Scholar] [CrossRef]
- Sarbassov, D.D.; Guertin, D.A.; Ali, S.M.; Sabatini, D.M. Phosphorylation and regulation of Akt/PKB by the rictor-mTOR complex. Science 2005, 307, 1098–1101. [Google Scholar] [CrossRef]
- Cross, D.A.; Alessi, D.R.; Cohen, P.; Andjelkovich, M.; Hemmings, B.A. Inhibition of glycogen synthase kinase-3 by insulin mediated by protein kinase B. Nature 1995, 378, 785–789. [Google Scholar] [CrossRef] [PubMed]
- Inoki, K.; Li, Y.; Zhu, T.; Wu, J.; Guan, K.L. TSC2 is phosphorylated and inhibited by Akt and suppresses mTOR signalling. Nat. Cell Biol. 2002, 4, 648–657. [Google Scholar] [CrossRef] [PubMed]
- Manning, B.D.; Toker, A. AKT/PKB signaling: Navigating the network. Cell 2017, 169, 381–405. [Google Scholar] [CrossRef] [PubMed]
- Brunet, A.; Bonni, A.; Zigmond, M.J.; Lin, M.Z.; Juo, P.; Hu, L.S.; Anderson, M.J.; Arden, K.C.; Blenis, J.; Greenberg, M.E. Akt promotes cell survival by phosphorylating and inhibiting a Forkhead transcription factor. Cell 1999, 96, 857–868. [Google Scholar] [CrossRef]
- Coloff, J.L.; Mason, E.F.; Altman, B.J.; Gerriets, V.A.; Liu, T.; Nichols, A.N.; Zhao, Y.; Wofford, J.A.; Jacobs, S.R.; Ilkayeva, O.; et al. Akt requires glucose metabolism to suppress puma expression and prevent apoptosis of leukemic T cells. J. Biol. Chem. 2011, 286, 5921–5933. [Google Scholar] [CrossRef] [PubMed]
- del Peso, L.; Gonzalez-Garcia, M.; Page, C.; Herrera, R.; Nunez, G. Interleukin-3-induced phosphorylation of BAD through the protein kinase Akt. Science 1997, 278, 687–689. [Google Scholar] [CrossRef] [PubMed]
- Pugazhenthi, S.; Nesterova, A.; Sable, C.; Heidenreich, K.A.; Boxer, L.M.; Heasley, L.E.; Reusch, J.E. Akt/protein kinase B up-regulates Bcl-2 expression through cAMP-response element-binding protein. J. Biol. Chem. 2000, 275, 10761–10766. [Google Scholar] [CrossRef]
- Lan, Y.L.; Zhou, J.J.; Liu, J.; Huo, X.K.; Wang, Y.L.; Liang, J.H.; Zhao, J.C.; Sun, C.P.; Yu, Z.L.; Fang, L.L.; et al. Uncaria rhynchophylla ameliorates Parkinson's Disease by inhibiting HSP90 expression: Insights from quantitative proteomics. Cell. Physiol. Biochem. 2018, 47, 1453–1464. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Sun, J.; Zhu, S.; Xu, T.; Lu, J.; Han, H.; Zhou, C.; Yan, J. The role of rhynchophylline in alleviating early brain injury following subarachnoid hemorrhage in rats. Brain Res. 2016, 1631, 92–100. [Google Scholar] [CrossRef] [PubMed]
- Julien, O.; Wells, J.A. Caspases and their substrates. Cell Death Differ. 2017, 24, 1380–1389. [Google Scholar] [CrossRef] [PubMed]
- Beurel, E.; Mines, M.A.; Song, L.; Jope, R.S. Glycogen synthase kinase-3 levels and phosphorylation undergo large fluctuations in mouse brain during development. Bipolar Disord. 2012, 14, 822–830. [Google Scholar] [CrossRef]
- Leroy, K.; Brion, J.P. Developmental expression and localization of glycogen synthase kinase-3beta in rat brain. J. Chem. Neuroanat. 1999, 16, 279–293. [Google Scholar] [CrossRef]
- Woodgett, J.R. Molecular cloning and expression of glycogen synthase kinase-3/factor A. EMBO J. 1990, 9, 2431–2438. [Google Scholar] [CrossRef] [PubMed]
- Beaulieu, J.M.; Gainetdinov, R.R.; Caron, M.G. Akt/GSK3 signaling in the action of psychotropic drugs. Annu. Rev. Pharmacol. Toxicol. 2009, 49, 327–347. [Google Scholar] [CrossRef]
- Beaulieu, J.M.; Sotnikova, T.D.; Yao, W.D.; Kockeritz, L.; Woodgett, J.R.; Gainetdinov, R.R.; Caron, M.G. Lithium antagonizes dopamine-dependent behaviors mediated by an AKT/glycogen synthase kinase 3 signaling cascade. Proc. Natl. Acad. Sci. USA 2004, 101, 5099–5104. [Google Scholar] [CrossRef]
- Jain, A.K.; Jaiswal, A.K. GSK-3beta acts upstream of Fyn kinase in regulation of nuclear export and degradation of NF-E2 related factor 2. J. Biol. Chem. 2007, 282, 16502–16510. [Google Scholar] [CrossRef] [PubMed]
- Wild, A.C.; Moinova, H.R.; Mulcahy, R.T. Regulation of gamma-glutamylcysteine synthetase subunit gene expression by the transcription factor Nrf2. J. Biol. Chem. 1999, 274, 33627–33636. [Google Scholar] [CrossRef]
- Nguyen, T.; Huang, H.C.; Pickett, C.B. Transcriptional regulation of the antioxidant response element. Activation by Nrf2 and repression by MafK. J. Biol. Chem. 2000, 275, 15466–15473. [Google Scholar] [CrossRef] [PubMed]
- Xue, R.; Wan, Y.; Sun, X.; Zhang, X.; Gao, W.; Wu, W. Nicotinic mitigation of neuroinflammation and oxidative stress after chronic sleep deprivation. Front. Immunol. 2019, 10, 2546. [Google Scholar] [CrossRef] [PubMed]
- Ahnaou, A.; Drinkenburg, W.H. Disruption of glycogen synthase kinase-3-beta activity leads to abnormalities in physiological measures in mice. Behav. Brain Res. 2011, 221, 246–252. [Google Scholar] [CrossRef]
- Benedetti, F.; Dallaspezia, S.; Lorenzi, C.; Pirovano, A.; Radaelli, D.; Locatelli, C.; Poletti, S.; Colombo, C.; Smeraldi, E. Gene-gene interaction of glycogen synthase kinase 3-beta and serotonin transporter on human antidepressant response to sleep deprivation. J. Affect. Disord. 2012, 136, 514–519. [Google Scholar] [CrossRef]
- Benedetti, F.; Serretti, A.; Colombo, C.; Lorenzi, C.; Tubazio, V.; Smeraldi, E. A glycogen synthase kinase 3-beta promoter gene single nucleotide polymorphism is associated with age at onset and response to total sleep deprivation in bipolar depression. Neurosci. Lett. 2004, 368, 123–126. [Google Scholar] [CrossRef]
- Vyazovskiy, V.V.; Cirelli, C.; Pfister-Genskow, M.; Faraguna, U.; Tononi, G. Molecular and electrophysiological evidence for net synaptic potentiation in wake and depression in sleep. Nat. Neurosci. 2008, 11, 200–208. [Google Scholar] [CrossRef]
- Bruning, F.; Noya, S.B.; Bange, T.; Koutsouli, S.; Rudolph, J.D.; Tyagarajan, S.K.; Cox, J.; Mann, M.; Brown, S.A.; Robles, M.S. Sleep-wake cycles drive daily dynamics of synaptic phosphorylation. Science 2019, 366. [Google Scholar] [CrossRef] [PubMed]
- Khlghatyan, J.; Evstratova, A.; Bozoyan, L.; Chamberland, S.; Chatterjee, D.; Marakhovskaia, A.; Soares Silva, T.; Toth, K.; Mongrain, V.; Beaulieu, J.M. Fxr1 regulates sleep and synaptic homeostasis. EMBO J. 2020, 39, e103864. [Google Scholar] [CrossRef] [PubMed]
- Yatham, L.N.; Kennedy, S.H.; Parikh, S.V.; Schaffer, A.; Bond, D.J.; Frey, B.N.; Sharma, V.; Goldstein, B.I.; Rej, S.; Beaulieu, S.; et al. Canadian Network for Mood and Anxiety Treatments (CANMAT) and International Society for Bipolar Disorders (ISBD) 2018 guidelines for the management of patients with bipolar disorder. Bipolar Disord. 2018, 20, 97–170. [Google Scholar] [CrossRef]
- Geoffroy, P.A.; Samalin, L.; Llorca, P.M.; Curis, E.; Bellivier, F. Influence of lithium on sleep and chronotypes in remitted patients with bipolar disorder. J. Affect. Disord. 2016, 204, 32–39. [Google Scholar] [CrossRef] [PubMed]
- Friston, K.J.; Sharpley, A.L.; Solomon, R.A.; Cowen, P.J. Lithium increases slow wave sleep: Possible mediation by brain 5-HT2 receptors? Psychopharmacology 1989, 98, 139–140. [Google Scholar] [CrossRef] [PubMed]
- Jones, C.A.; Perez, E.; Amici, R.; Luppi, M.; Baracchi, F.; Cerri, M.; Dentico, D.; Zamboni, G. Lithium affects REM sleep occurrence, autonomic activity and brain second messengers in the rat. Behav. Brain Res. 2008, 187, 254–261. [Google Scholar] [CrossRef]
- Rubinsztein, D.C.; Gestwicki, J.E.; Murphy, L.O.; Klionsky, D.J. Potential therapeutic applications of autophagy. Nat. Rev. Drug Discov. 2007, 6, 304–312. [Google Scholar] [CrossRef]
- Tudor, J.C.; Davis, E.J.; Peixoto, L.; Wimmer, M.E.; van Tilborg, E.; Park, A.J.; Poplawski, S.G.; Chung, C.W.; Havekes, R.; Huang, J.; et al. Sleep deprivation impairs memory by attenuating mTORC1-dependent protein synthesis. Sci. Signal. 2016, 9, ra41. [Google Scholar] [CrossRef]
- Shabab, T.; Khanabdali, R.; Moghadamtousi, S.Z.; Kadir, H.A.; Mohan, G. Neuroinflammation pathways: A general review. Int. J. Neurosci. 2017, 127, 624–633. [Google Scholar] [CrossRef] [PubMed]
- Kielian, T. Overview of toll-like receptors in the CNS. Curr. Top. Microbiol. Immunol. 2009, 336, 1–14. [Google Scholar] [CrossRef]
- Cespuglio, R.; Amrouni, D.; Meiller, A.; Buguet, A.; Gautier-Sauvigne, S. Nitric oxide in the regulation of the sleep-wake states. Sleep Med. Rev. 2012, 16, 265–279. [Google Scholar] [CrossRef]
- Jewett, K.A.; Krueger, J.M. Humoral sleep regulation; interleukin-1 and tumor necrosis factor. Vitam. Horm. 2012, 89, 241–257. [Google Scholar] [CrossRef] [PubMed]
- Hars, B. Endogenous nitric oxide in the rat pons promotes sleep. Brain Res. 1999, 816, 209–219. [Google Scholar] [CrossRef]
- Fang, J.; Wang, Y.; Krueger, J.M. Effects of interleukin-1 beta on sleep are mediated by the type I receptor. Am. J. Physiol. 1998, 274, R655–R660. [Google Scholar] [CrossRef] [PubMed]
- Datta, S.; Patterson, E.H.; Siwek, D.F. Endogenous and exogenous nitric oxide in the pedunculopontine tegmentum induces sleep. Synapse 1997, 27, 69–78. [Google Scholar] [CrossRef]
- Opp, M.R.; Obal, F., Jr.; Krueger, J.M. Interleukin 1 alters rat sleep: Temporal and dose-related effects. Am. J. Physiol. 1991, 260, R52–R58. [Google Scholar] [CrossRef]
- Opp, M.R.; Krueger, J.M. Interleukin 1-receptor antagonist blocks interleukin 1-induced sleep and fever. Am. J. Physiol. 1991, 260, R453–R457. [Google Scholar] [CrossRef] [PubMed]
- Dickstein, J.B.; Moldofsky, H.; Lue, F.A.; Hay, J.B. Intracerebroventricular injection of TNF-alpha promotes sleep and is recovered in cervical lymph. Am. J. Physiol. 1999, 276, R1018–R1022. [Google Scholar] [CrossRef]
- Kapas, L.; Shibata, M.; Kimura, M.; Krueger, J.M. Inhibition of nitric oxide synthesis suppresses sleep in rabbits. Am. J. Physiol. 1994, 266, R151–R157. [Google Scholar] [CrossRef]
- Opp, M.R.; Krueger, J.M. Interleukin-1 is involved in responses to sleep deprivation in the rabbit. Brain Res. 1994, 639, 57–65. [Google Scholar] [CrossRef]
- Opp, M.R.; Krueger, J.M. Anti-interleukin-1 beta reduces sleep and sleep rebound after sleep deprivation in rats. Am. J. Physiol. 1994, 266, R688–R695. [Google Scholar] [CrossRef]
- Kapas, L.; Fang, J.; Krueger, J.M. Inhibition of nitric oxide synthesis inhibits rat sleep. Brain Res. 1994, 664, 189–196. [Google Scholar] [CrossRef]
- Dzoljic, M.R.; de Vries, R.; van Leeuwen, R. Sleep and nitric oxide: Effects of 7-nitro indazole, inhibitor of brain nitric oxide synthase. Brain Res. 1996, 718, 145–150. [Google Scholar] [CrossRef]
- Takahashi, S.; Kapas, L.; Seyer, J.M.; Wang, Y.; Krueger, J.M. Inhibition of tumor necrosis factor attenuates physiological sleep in rabbits. Neuroreport 1996, 7, 642–646. [Google Scholar] [CrossRef]
- Takahashi, S.; Tooley, D.D.; Kapas, L.; Fang, J.; Seyer, J.M.; Krueger, J.M. Inhibition of tumor necrosis factor in the brain suppresses rabbit sleep. Pflug. Arch. 1995, 431, 155–160. [Google Scholar] [CrossRef]
- Takahashi, S.; Fang, J.; Kapas, L.; Wang, Y.; Krueger, J.M. Inhibition of brain interleukin-1 attenuates sleep rebound after sleep deprivation in rabbits. Am. J. Physiol. 1997, 273, R677–R682. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, A.C.; Kapas, L. Day- and night time injection of a nitric oxide synthase inhibitor elicits opposite sleep responses in rats. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2005, 289, R521–R531. [Google Scholar] [CrossRef] [PubMed]
- Monti, J.M.; Jantos, H. Microinjection of the nitric oxide synthase inhibitor L-NAME into the lateral basal forebrain alters the sleep/wake cycle of the rat. Prog. Neuropsychopharmacol. Biol. Psychiatry 2004, 28, 239–247. [Google Scholar] [CrossRef]
- Kubota, T.; Kushikata, T.; Fang, J.; Krueger, J.M. Nuclear factor-kappaB inhibitor peptide inhibits spontaneous and interleukin-1beta-induced sleep. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2000, 279, R404–R413. [Google Scholar] [CrossRef]
- Zielinski, M.R.; Kim, Y.; Karpova, S.A.; McCarley, R.W.; Strecker, R.E.; Gerashchenko, D. Chronic sleep restriction elevates brain interleukin-1 beta and tumor necrosis factor-alpha and attenuates brain-derived neurotrophic factor expression. Neurosci. Lett. 2014, 580, 27–31. [Google Scholar] [CrossRef]
- Kalinchuk, A.V.; McCarley, R.W.; Porkka-Heiskanen, T.; Basheer, R. Sleep deprivation triggers inducible nitric oxide-dependent nitric oxide production in wake-active basal forebrain neurons. J. Neurosci. 2010, 30, 13254–13264. [Google Scholar] [CrossRef]
- Mackiewicz, M.; Sollars, P.J.; Ogilvie, M.D.; Pack, A.I. Modulation of IL-1 beta gene expression in the rat CNS during sleep deprivation. Neuroreport 1996, 7, 529–533. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Gardi, J.; Kushikata, T.; Fang, J.; Krueger, J.M. Nuclear factor-kappaB-like activity increases in murine cerebral cortex after sleep deprivation. Am. J. Physiol. 1999, 276, R1812–R1818. [Google Scholar] [CrossRef]
- Ribeiro, A.C.; Gilligan, J.G.; Kapas, L. Systemic injection of a nitric oxide synthase inhibitor suppresses sleep responses to sleep deprivation in rats. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2000, 278, R1048–R1056. [Google Scholar] [CrossRef]
- Shoham, S.; Davenne, D.; Cady, A.B.; Dinarello, C.A.; Krueger, J.M. Recombinant tumor necrosis factor and interleukin 1 enhance slow-wave sleep. Am. J. Physiol. 1987, 253, R142–R149. [Google Scholar] [CrossRef] [PubMed]
- Surmeier, D.J.; Bargas, J.; Hemmings, H.C., Jr.; Nairn, A.C.; Greengard, P. Modulation of calcium currents by a D1 dopaminergic protein kinase/phosphatase cascade in rat neostriatal neurons. Neuron 1995, 14, 385–397. [Google Scholar] [CrossRef]
- Huang, X.Y.; Morielli, A.D.; Peralta, E.G. Tyrosine kinase-dependent suppression of a potassium channel by the G protein-coupled m1 muscarinic acetylcholine receptor. Cell 1993, 75, 1145–1156. [Google Scholar] [CrossRef]
- Baghdoyan, H.A.; Rodrigo-Angulo, M.L.; McCarley, R.W.; Hobson, J.A. Site-specific enhancement and suppression of desynchronized sleep signs following cholinergic stimulation of three brainstem regions. Brain Res. 1984, 306, 39–52. [Google Scholar] [CrossRef]
- Niwa, Y.; Kanda, G.N.; Yamada, R.G.; Shi, S.; Sunagawa, G.A.; Ukai-Tadenuma, M.; Fujishima, H.; Matsumoto, N.; Masumoto, K.H.; Nagano, M.; et al. Muscarinic acetylcholine receptors Chrm1 and Chrm3 are essential for REM sleep. Cell Rep. 2018, 24, 2231–2247. [Google Scholar] [CrossRef]
- Gillin, J.C.; Sutton, L.; Ruiz, C.; Golshan, S.; Hirsch, S.; Warmann, C.; Shiromani, P. Dose dependent inhibition of REM sleep in normal volunteers by biperiden, a muscarinic antagonist. Biol. Psychiatry 1991, 30, 151–156. [Google Scholar] [CrossRef]
- Kurimoto, E.; Nakashima, M.; Kimura, H.; Suzuki, M. TAK-071, a muscarinic M1 receptor positive allosteric modulator, attenuates scopolamine-induced quantitative electroencephalogram power spectral changes in cynomolgus monkeys. PLoS ONE 2019, 14, e0207969. [Google Scholar] [CrossRef]
- Ma, X.; Zhang, Y.; Wang, L.; Li, N.; Barkai, E.; Zhang, X.; Lin, L.; Xu, J. The firing of theta state-related septal cholinergic neurons disrupt hippocampal ripple oscillations via muscarinic receptors. J. Neurosci. 2020, 40, 3591–3603. [Google Scholar] [CrossRef] [PubMed]
- Shirey, J.K.; Brady, A.E.; Jones, P.J.; Davis, A.A.; Bridges, T.M.; Kennedy, J.P.; Jadhav, S.B.; Menon, U.N.; Xiang, Z.; Watson, M.L.; et al. A selective allosteric potentiator of the M1 muscarinic acetylcholine receptor increases activity of medial prefrontal cortical neurons and restores impairments in reversal learning. J. Neurosci. 2009, 29, 14271–14286. [Google Scholar] [CrossRef] [PubMed]
- Williams, J.H.; Kauer, J.A. Properties of carbachol-induced oscillatory activity in rat hippocampus. J. Neurophysiol. 1997, 78, 2631–2640. [Google Scholar] [CrossRef]
- Cea-del Rio, C.A.; Lawrence, J.J.; Tricoire, L.; Erdelyi, F.; Szabo, G.; McBain, C.J. M3 muscarinic acetylcholine receptor expression confers differential cholinergic modulation to neurochemically distinct hippocampal basket cell subtypes. J. Neurosci. 2010, 30, 6011–6024. [Google Scholar] [CrossRef] [PubMed]
- Fisahn, A.; Yamada, M.; Duttaroy, A.; Gan, J.W.; Deng, C.X.; McBain, C.J.; Wess, J. Muscarinic induction of hippocampal gamma oscillations requires coupling of the M1 receptor to two mixed cation currents. Neuron 2002, 33, 615–624. [Google Scholar] [CrossRef]
- Langmead, C.J.; Austin, N.E.; Branch, C.L.; Brown, J.T.; Buchanan, K.A.; Davies, C.H.; Forbes, I.T.; Fry, V.A.; Hagan, J.J.; Herdon, H.J.; et al. Characterization of a CNS penetrant, selective M1 muscarinic receptor agonist, 77-LH-28-1. Br. J. Pharmacol. 2008, 154, 1104–1115. [Google Scholar] [CrossRef] [PubMed]
- Ursin, R. Serotonin and sleep. Sleep Med. Rev. 2002, 6, 55–69. [Google Scholar] [CrossRef]
- Ito, H.; Yanase, M.; Yamashita, A.; Kitabatake, C.; Hamada, A.; Suhara, Y.; Narita, M.; Ikegami, D.; Sakai, H.; Yamazaki, M.; et al. Analysis of sleep disorders under pain using an optogenetic tool: Possible involvement of the activation of dorsal raphe nucleus-serotonergic neurons. Mol. Brain 2013, 6, 59. [Google Scholar] [CrossRef]
- Cape, E.G.; Jones, B.E. Differential modulation of high-frequency gamma-electroencephalogram activity and sleep-wake state by noradrenaline and serotonin microinjections into the region of cholinergic basalis neurons. J. Neurosci. 1998, 18, 2653–2666. [Google Scholar] [CrossRef] [PubMed]
- Horner, R.L.; Sanford, L.D.; Annis, D.; Pack, A.I.; Morrison, A.R. Serotonin at the laterodorsal tegmental nucleus suppresses rapid-eye-movement sleep in freely behaving rats. J. Neurosci. 1997, 17, 7541–7552. [Google Scholar] [CrossRef]
- Chowdhury, S.; Yamanaka, A. Optogenetic activation of serotonergic terminals facilitates GABAergic inhibitory input to orexin/hypocretin neurons. Sci. Rep. 2016, 6, 36039. [Google Scholar] [CrossRef] [PubMed]
- Saito, Y.C.; Maejima, T.; Nishitani, M.; Hasegawa, E.; Yanagawa, Y.; Mieda, M.; Sakurai, T. Monoamines inhibit GABAergic neurons in ventrolateral preoptic area that make direct synaptic connections to hypothalamic arousal neurons. J. Neurosci. 2018, 38, 6366–6378. [Google Scholar] [CrossRef] [PubMed]
- Linley, S.B.; Vertes, R.P. Serotonergic systems in sleep and waking. In Handbook of Behavioral Neuroscience; Dringenberg, H.C., Ed.; Elsevier: Amsterdam, The Netherlands, 2019; Volume 30, pp. 101–123. [Google Scholar]
- Bjorvatn, B.; Fagerland, S.; Eid, T.; Ursin, R. Sleep/waking effects of a selective 5-HT1A receptor agonist given systemically as well as perfused in the dorsal raphe nucleus in rats. Brain Res. 1997, 770, 81–88. [Google Scholar] [CrossRef]
- Monti, J.M.; Jantos, H.; Monti, D. Increased REM sleep after intra-dorsal raphe nucleus injection of flesinoxan or 8-OHDPAT: Prevention with WAY 100635. Eur. Neuropsychopharmacol. 2002, 12, 47–55. [Google Scholar] [CrossRef]
- Portas, C.M.; Thakkar, M.; Rainnie, D.; McCarley, R.W. Microdialysis perfusion of 8-hydroxy-2-(di-n-propylamino)tetralin (8-OH-DPAT) in the dorsal raphe nucleus decreases serotonin release and increases rapid eye movement sleep in the freely moving cat. J. Neurosci. 1996, 16, 2820–2828. [Google Scholar] [CrossRef]
- Vertes, R.P.; Kinney, G.G.; Kocsis, B.; Fortin, W.J. Pharmacological suppression of the median raphe nucleus with serotonin1A agonists, 8-OH-DPAT and buspirone, produces hippocampal theta rhythm in the rat. Neuroscience 1994, 60, 441–451. [Google Scholar] [CrossRef]
- Monti, J.M.; Jantos, H. Effects of activation and blockade of 5-HT2A/2C receptors in the dorsal raphe nucleus on sleep and waking in the rat. Prog. Neuropsychopharmacol. Biol. Psychiatry 2006, 30, 1189–1195. [Google Scholar] [CrossRef]
- Monti, J.M.; Leopoldo, M.; Jantos, H. The serotonin 5-HT7 receptor agonist LP-44 microinjected into the dorsal raphe nucleus suppresses REM sleep in the rat. Behav. Brain Res. 2008, 191, 184–189. [Google Scholar] [CrossRef]
- Monti, J.M.; Jantos, H.; Lagos, P. Activation of serotonin 5-HT(1B) receptor in the dorsal raphe nucleus affects REM sleep in the rat. Behav. Brain Res. 2010, 206, 8–16. [Google Scholar] [CrossRef]
- Monti, J.M.; Jantos, H. Effects of the serotonin 5-HT2A/2C receptor agonist DOI and of the selective 5-HT2A or 5-HT2C receptor antagonists EMD 281014 and SB-243213, respectively, on sleep and waking in the rat. Eur. J. Pharmacol. 2006, 553, 163–170. [Google Scholar] [CrossRef]
- Popa, D.; Lena, C.; Fabre, V.; Prenat, C.; Gingrich, J.; Escourrou, P.; Hamon, M.; Adrien, J. Contribution of 5-HT2 receptor subtypes to sleep-wakefulness and respiratory control, and functional adaptations in knock-out mice lacking 5-HT2A receptors. J. Neurosci. 2005, 25, 11231–11238. [Google Scholar] [CrossRef] [PubMed]
- Dahan, L.; Astier, B.; Vautrelle, N.; Urbain, N.; Kocsis, B.; Chouvet, G. Prominent burst firing of dopaminergic neurons in the ventral tegmental area during paradoxical sleep. Neuropsychopharmacology 2007, 32, 1232–1241. [Google Scholar] [CrossRef]
- Eban-Rothschild, A.; Rothschild, G.; Giardino, W.J.; Jones, J.R.; de Lecea, L. VTA dopaminergic neurons regulate ethologically relevant sleep-wake behaviors. Nat. Neurosci. 2016, 19, 1356–1366. [Google Scholar] [CrossRef]
- Lancel, M. Role of GABAA receptors in the regulation of sleep: Initial sleep responses to peripherally administered modulators and agonists. Sleep 1999, 22, 33–42. [Google Scholar] [CrossRef] [PubMed]
- Winsky-Sommerer, R. Role of GABAA receptors in the physiology and pharmacology of sleep. Eur. J. Neurosci. 2009, 29, 1779–1794. [Google Scholar] [CrossRef] [PubMed]
- Hannou, L.; Roy, P.G.; Ballester Roig, M.N.; Mongrain, V. Transcriptional control of synaptic components by the clock machinery. Eur. J. Neurosci. 2020, 51, 241–267. [Google Scholar] [CrossRef] [PubMed]
- Biello, S.M.; Bonsall, D.R.; Atkinson, L.A.; Molyneux, P.C.; Harrington, M.E.; Lall, G.S. Alterations in glutamatergic signaling contribute to the decline of circadian photoentrainment in aged mice. Neurobiol. Aging 2018, 66, 75–84. [Google Scholar] [CrossRef]
- Wang, L.M.; Schroeder, A.; Loh, D.; Smith, D.; Lin, K.; Han, J.H.; Michel, S.; Hummer, D.L.; Ehlen, J.C.; Albers, H.E.; et al. Role for the NR2B subunit of the N-methyl-D-aspartate receptor in mediating light input to the circadian system. Eur. J. Neurosci. 2008, 27, 1771–1779. [Google Scholar] [CrossRef]
- Bendova, Z.; Sladek, M.; Svobodova, I. The expression of NR2B subunit of NMDA receptor in the suprachiasmatic nucleus of Wistar rats and its role in glutamate-induced CREB and ERK1/2 phosphorylation. Neurochem. Int. 2012, 61, 43–47. [Google Scholar] [CrossRef]
- Bendova, Z.; Sumova, A.; Mikkelsen, J.D. Circadian and developmental regulation of N-methyl-d-aspartate-receptor 1 mRNA splice variants and N-methyl-d-aspartate-receptor 3 subunit expression within the rat suprachiasmatic nucleus. Neuroscience 2009, 159, 599–609. [Google Scholar] [CrossRef] [PubMed]
- Coria-Lucero, C.D.; Golini, R.S.; Ponce, I.T.; Deyurka, N.; Anzulovich, A.C.; Delgado, S.M.; Navigatore-Fonzo, L.S. Rhythmic Bdnf and TrkB expression patterns in the prefrontal cortex are lost in aged rats. Brain Res. 2016, 1653, 51–58. [Google Scholar] [CrossRef]
- Cai, Y.; Ding, H.; Li, N.; Chai, Y.; Zhang, Y.; Chan, P. Oscillation development for neurotransmitter-related genes in the mouse striatum. Neuroreport 2010, 21, 79–83. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Bi, Q.; Cui, H.; Lv, C.; Wang, M. Suppression of autophagy through JAK2/STAT3 contributes to the therapeutic action of rhynchophylline on asthma. BMC Complement. Med. Ther. 2021, 21, 21. [Google Scholar] [CrossRef]
- Boudreau, P.; Yeh, W.H.; Dumont, G.A.; Boivin, D.B. Circadian variation of heart rate variability across sleep stages. Sleep 2013, 36, 1919–1928. [Google Scholar] [CrossRef]
- Trinder, J.; Waloszek, J.; Woods, M.J.; Jordan, A.S. Sleep and cardiovascular regulation. Pflug. Arch. 2012, 463, 161–168. [Google Scholar] [CrossRef]
- Song, M.F.; Guan, Y.H.; Li, H.T.; Wei, S.G.; Zhang, L.X.; Zhang, Z.L.; Ma, X.J. The effects of genetic variation and environmental factors on rhynchophylline and isorhynchophylline in Uncaria macrophylla Wall. from different populations in China. PLoS ONE 2018, 13, e0199259. [Google Scholar] [CrossRef]
- Williamson, E.M. Synergy and other interactions in phytomedicines. Phytomedicine 2001, 8, 401–409. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Rhy Application | Timing of Measurement | Rhy Effect | Model | Reference |
|---|---|---|---|---|
| INCUBATIONS | ||||
| 20 s | Immediate | Attenuates epilepsy-induced ↑ in NMDAR current in EC slices | Rat brain slices | [19] |
| 80 s | Immediate | Accelerates activation and inactivation of VGKC Accelerates activation and inactivation of Kv1.2 | N2A cells HEK293 | [64] |
| 3–8 min | Immediate | ↓ mAChR1- and 5-HT2-mediated currents (effect disappears after 1 min) | Xenopus oocytes | [65] |
| Attenuates epilepsy-induced ↑ of EC neuron discharge frequency | Rats | [19] | ||
| ↓ open time and ↑ close time of L-VGCCs | Rat cortical neurons | [66] | ||
| ↓ Ca2+ influx via L-VGCCs | Rat cardiomyocytes | [67] | ||
| Non-competitive inhibition of NMDAR current | Xenopus oocytes | [68] | ||
| 15–30 min | Immediate | ↓ EfnA1-dependent EphA4 phosphorylation and EphA4 clusters | Rat cortical neurons | [18] |
| Attenuates ischemia-induced ↓ in population spike amplitude | Rat hipp. slices | [65] | ||
| ↓ Ca2+ intracellular increase via L-VGCC, promotes vasodilation | Human artery smooth muscle cells | [69] | ||
| 1 h | Immediate | Attenuates ischemia-induced ↑ in ROS, MDA, LDH, mPTP, AIF, Ca2+ and caspase 3 and 9 mRNA and protein Attenuates ischemia-induced ↓ in mitochondrial membrane potential, SOD, GPx, Cytc | Rat cardiomyocytes | [70] |
| ↑ GAD65/67 and GABAAR subunits expression | Rat hypothalamic neurons | [22] | ||
| 30 min | 2 h post Rhy | Attenuates Aβ-induced ↑ in EphA4 phosphorylation and LTP impairment | Rat hipp. slices | [18] |
| 2–6 h | Immediate | Attenuates LPS-induced ↑ in Cox2, iNos, Ccl2 mRNAs | Rat microglia | [71] |
| ↑ Grin1 mRNA (no difference in Grin2b) | Rat hipp. neurons | [72] | ||
| 12 h | Immediate | Improves endothelial relaxation and ↑ p-Src, p-AKT and NO (in hypertensive rat arteries) and ↑ p-eNOS (in WT arteries) | Rat intrarenal arteries | [73] |
| 24 h | Immediate | Attenuates LPS-induced ↑ in p-ERK, p-38, p-IkBα, NFκBp65 Attenuates LPS-induced ↓ in IkBα Attenuates LPS-induced ↑ in culture medium MCP1, PGE2, NO, IL1β, TNFα | Rat microglia | [71] |
| 1 h | 24 h post Isorhy | * Attenuates MPP-induced ↑ in p-GSK3β Tyr297, p-FYN and ROS * ↑ nuclear NRF2 and ARE transcriptional activity | Human SH-SY5Y neuroblastoma cells | [74] |
| 2 h | 24 h post Rhy | Attenuates MPP-induced ↓ in p-GSK3β Ser9, p-AKT and MEF2D Attenuates MPP-induced ↑ in Bax/Bcl-2 ratio | Rat granule neurons | [75] |
| 48 h | Immediate | ↑ Grin1 mRNA and GluN1, and ↓ Grin2b mRNA and GluN2B | Rat hipp. neurons | [72] |
| Attenuates LPS-induced ↑ in NO, iNOS, TNFα, IL-1β, p-p38, p-ERK Attenuates LPS-induced ↓ in IkBα | N9 mouse microglia | [76] | ||
| ↓ GluN1 and ↓ ketamine-induced ↑ in GluA2/3 | PC12 cells | [77] | ||
| 72 h | Immediate | ↑ proliferation, GluN1, GluN2B, GluN3A ↓ BDNF, OXTR, and ATP Alters proliferation/differentiation related genes | Bone mesenchymal human cells | [78] |
| 24 h | 48 h post Rhy | Attenuates MPP-induced ↑ ROS, LDH, Caspase-3 activity and apoptosis; Attenuates MPP-induced ↓ Bcl2/Bax ratio and p-AKT | PC12 cells | [79] |
| SINGLE ADMINISTRATIONS | ||||
| IC | 100–600 s post Rhy | Attenuates Aβ-induced ↑ in the frequency of spontaneous discharge in CA1 | Rats | [80] |
| IV | 30 min post Rhy | Attenuates ischemia-induced ↓ in 5HIAA and DOPAC in striatum and hipp. Attenuates ischemia-induced ↑ of NE in striatum and hipp. | Rats | [81] |
| IP | 50 min post Rhy | ↓ DA in cortex, hypothalamus, and brainstem ↓ 5-HT in amygdala ↑ 5-HT in hypothalamus, and ↓ 5-HT release in hypothalamic slices ↑ 5-HT release in cortex, amygdala, and brainstem slices ↑ DA release in cortex, hypothalamus, amygdala, and brainstem slices ↓ righting reflex and spontaneous locomotor activity | Rats | [61] |
| Oral | 0–6 h post Rhy | ↓ locomotor activity and sleep latency, ↑ total sleep time ↓ number of sleep/wake cycles, ↑ total sleep time and REM sleep | Mice and Rats | [22] |
| IP | 48 h post Rhy | Attenuates stress-induced ↑ p-EphA4, p-FYN, p-Cdk5, p-Ephexin in PFC, CA3, DG Attenuates stress-induced ↓ BDNF, p-TrkB, PSD95, spines in PFC, CA3, DG | Mice | [17] |
| IP | 52 h post Rhy | Attenuates NTG-induced ↑ in EEG theta and delta activity, oxidative stress (GSH, blood CGRP), p-ERK1/2, p-JNK, p-p38, p-IκBα, and nuclear NF-κB p65 (all in trigeminal nucleus caudalis) | Rats | [82] |
| Hipp. inj | 2 w post Rhy | Attenuates Aβ-induced ↑ cell death, GluN2B, and NMDA Ca2+ influx in CA1 | Rats | [83] |
| MULTIPLE ADMINISTRATIONS | ||||
| SC for 3 days | 1–3 h after last injection | Attenuates LPS-induced ↓ in stroke volume and cardiac output Attenuates LPS-induced ↑ in IL-1β, TNFα and p-IkBα in heart, macrophages and serum | Mice | [84] |
| IP for 3 days | 3 h after last injection | * Attenuates KA-induced epileptic seizures ** Alters levels of Bdnf, Fos, Nfkbia, Map2k3, Il1b in cerebral cortex and hipp. | Rats | [85] |
| Attenuates KA-induced epileptic seizures | [86] | |||
| Attenuates KA-induced epileptic seizures and KA- induced ↑ in hippocampal p-JNK ** Attenuates KA-induced ↓ in cortical IL-6 | [87] | |||
| IP for 3 days | 12 h after last injection | Attenuates meth-induced ↑ in 5-HT, DA, TH, Glut, GluN2B, and locomotion | Zebrafishes | [88] |
| Attenuates meth-induced ↑ in GluA1 and CPP | [89] | |||
| Attenuates meth-induced ↑ in p-CREB and c-fos positive cells in CA1 and striatum | Rats | [90] | ||
| Attenuates amph-induced ↑ in CPP, glutamic acid, DA, and NE Attenuates amph-induced ↓ in GABA, endorphin, and ACh | [91] | |||
| Attenuates ketamine-induced ↑ in CPP, Nr4a2 and Bdnf mRNAs, NURR1, BDNF, p-CREB (all hipp.) | [21,92] | |||
| Attenuates amph-induced ↑ in CPP and Grin2b mRNA, and GluN2B protein in mPFC and CA1 | [20] | |||
| Attenuates meth-induced ↑ in CPP and GluN2B in brain tissue | Mice | [93] | ||
| IP for 3 days | 24 h after last injection | Attenuates KA-induced ↑ in IL-1β and BDNF positive cells in cortex and hipp. | Rats | [85] |
| Attenuates KA-induced ↑ NO scavenging activity in blood | [86] | |||
| IP for 5 days | 24 h after last injection | ↓ brain infarction and neurological deficits in a stroke model In cerebral cortex: Accentuates ischemia-induced ↑ in p-AKT and p-mTOR Attenuates ischemia-induced ↑ in TLR2,4, MyD88, caspase 3, and nuclear NF-κB Attenuates stroke-induced ↓ in p-BAD, BDNF, Bdnf and claudin-5 | [94] | |
| ICV infusion for 9 days | 33–34 h after ICV | Attenuates epilepsy-induced ↑ EC discharge frequency, neuronal death and GluN2B and Nav1.6 | Rats | [19] |
| 1 week gavage | 1 week after last gavage | Attenuates cytotoxicity-induced ↓ in TH-positive cells in substantia nigra | Mice | [79] |
| 2 weeks gavage | Immediate | ** Attenuates KA-induced neuronal death and KA-induced ↑ in spike amplitude | Rats hipp. slices | [95] |
| 3 weeks oral | Not specified | Attenuates DOI-induced ↑ TNFα, IL-6, and IL-1B (in serum and striatum); Attenuates DOI-induced ↑ p-NF-κB p65, p-IkBα, TLR2, caspase1, MyD88, DA, D2R (in striatum) Attenuates DOI-induced ↓ in p-TrkB, BDNF (in striatum), and cell viability | Rats | [96,97] |
| 3 weeks gavage | 24 h after last gavage | * Attenuates Aβ-induced ↓ in p-AKT, p-GSK3β (in brain), Bcl2/Bax in hipp., and memory * Attenuates Aβ-induced ↑ in caspases 3 and 9 in hipp. | Rats | [98] |
| 3–4 weeks gavage | Immediate | Attenuates p-EphA4 and rescues LTP in hipp. slices in APP mice | Mice | [18] |
| 3 weeks gavage | 5 days after last gavage | * Attenuates chronic mild stress-induced ↓ p-AKT, p-GSK3β, BDNF, NGF in cortex and hipp., and sucrose preference * Attenuates chronic mild stress-induced ↑ in TNFα, IL-6, nuclear NF-κB in cortex and hipp., and locomotion | Mice | [99] |
| 1 day gavage/week for 4 weeks | 24 h after last gavage | Attenuates asthma-induced ↑ in eosinophil recruitment, IL-13, IL-4, IL-5 in serum Attenuates asthma-induced ↑ TGFβ, Smad4, p-Smad2, p-Smad3, p-ERK1/2 and p-38 in lung tissue | Mice | [100] |
| 6 weeks in food | Immediate | * Attenuates cardiac hypertrophy-induced ↑ in TGFβ1, cTGF, Collagen1,3, p-ERK, p-38, p-JNK, and attenuates the induced ↓ in SOD2 * ↑ NRF2 and accentuates the induced ↑ in SOD3 | Mice | [101] |
| Effects under Baseline and/or Pathological Conditions | Sex(es) Studied | Reference | |
|---|---|---|---|
| VGCC | Baseline conditions | Males | [67,103] |
| Baseline conditions | Males and females | [104] | |
| Baseline conditions | Not indicated | [69] | |
| Pathological conditions | Not indicated | [66] | |
| VGKC | Baseline conditions | Male and female cell lines | [64] |
| NMDAR | Baseline conditions | Not indicated | [68,72,78] |
| Pathological conditions | Males | [19,88] | |
| Pathological conditions; no effect under baseline | Males | [20,83] | |
| Pathological conditions | Not indicated | [93] | |
| EPHA4 | Pathological conditions; no effect under baseline | Males and females | [18] |
| Pathological conditions; no effect under baseline | Males | [17] | |
| BDNF/TRKB | Baseline conditions | Not indicated | [78] |
| Pathological conditions | Males | [85,92,94] | |
| Pathological conditions; no effect under baseline | Males | [17,99] * | |
| Pathological conditions | Not indicated | [21] | |
| ERK/MAPK | Pathological conditions | Male cell line | [76] |
| Pathological conditions | Not indicated | [71] | |
| Pathological conditions | Female | [100] | |
| Pathological conditions | Males | [82,86,87] | |
| Pathological conditions; no effect under baseline | Males | [101] * | |
| CREB | Pathological conditions | Males | [92] |
| Pathological conditions | Not indicated | [21,90] | |
| PI3K/AKT | Pathological conditions | Males | [73] |
| Pathological conditions | Male cell line | [79] | |
| Pathological conditions | Not indicated | [70] | |
| Pathological conditions; no effect under baseline | Not indicated | [75] | |
| Pathological conditions; only one effect under baseline | Males | [101] * | |
| Pathological conditions | Males | [94] | |
| Pathological conditions; no effect under baseline | Males | [98,99] * | |
| NF-κB | Pathological conditions | Male cell line | [76] |
| Pathological conditions | Not indicated | [71] | |
| Pathological conditions | Males | [82,85,86,94,97] | |
| Pathological conditions; no effect under baseline | Males | [84] | |
| Other NTs | Baseline conditions | Not indicated | [65] |
| Baseline conditions | Males and females | [61] | |
| Pathological conditions | Males | [88,96] | |
| Pathological conditions; no effect under baseline | Not indicated | [91] | |
| GABAAR | Baseline conditions | Male neurons | [22] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ballester Roig, M.N.; Leduc, T.; Areal, C.C.; Mongrain, V. Cellular Effects of Rhynchophylline and Relevance to Sleep Regulation. Clocks & Sleep 2021, 3, 312-341. https://doi.org/10.3390/clockssleep3020020
Ballester Roig MN, Leduc T, Areal CC, Mongrain V. Cellular Effects of Rhynchophylline and Relevance to Sleep Regulation. Clocks & Sleep. 2021; 3(2):312-341. https://doi.org/10.3390/clockssleep3020020
Chicago/Turabian StyleBallester Roig, Maria Neus, Tanya Leduc, Cassandra C. Areal, and Valérie Mongrain. 2021. "Cellular Effects of Rhynchophylline and Relevance to Sleep Regulation" Clocks & Sleep 3, no. 2: 312-341. https://doi.org/10.3390/clockssleep3020020
APA StyleBallester Roig, M. N., Leduc, T., Areal, C. C., & Mongrain, V. (2021). Cellular Effects of Rhynchophylline and Relevance to Sleep Regulation. Clocks & Sleep, 3(2), 312-341. https://doi.org/10.3390/clockssleep3020020