Late Middle—Early Late Pleistocene Mammoths from the Lower Don River Region (Russia)


Abstract
1. Introduction
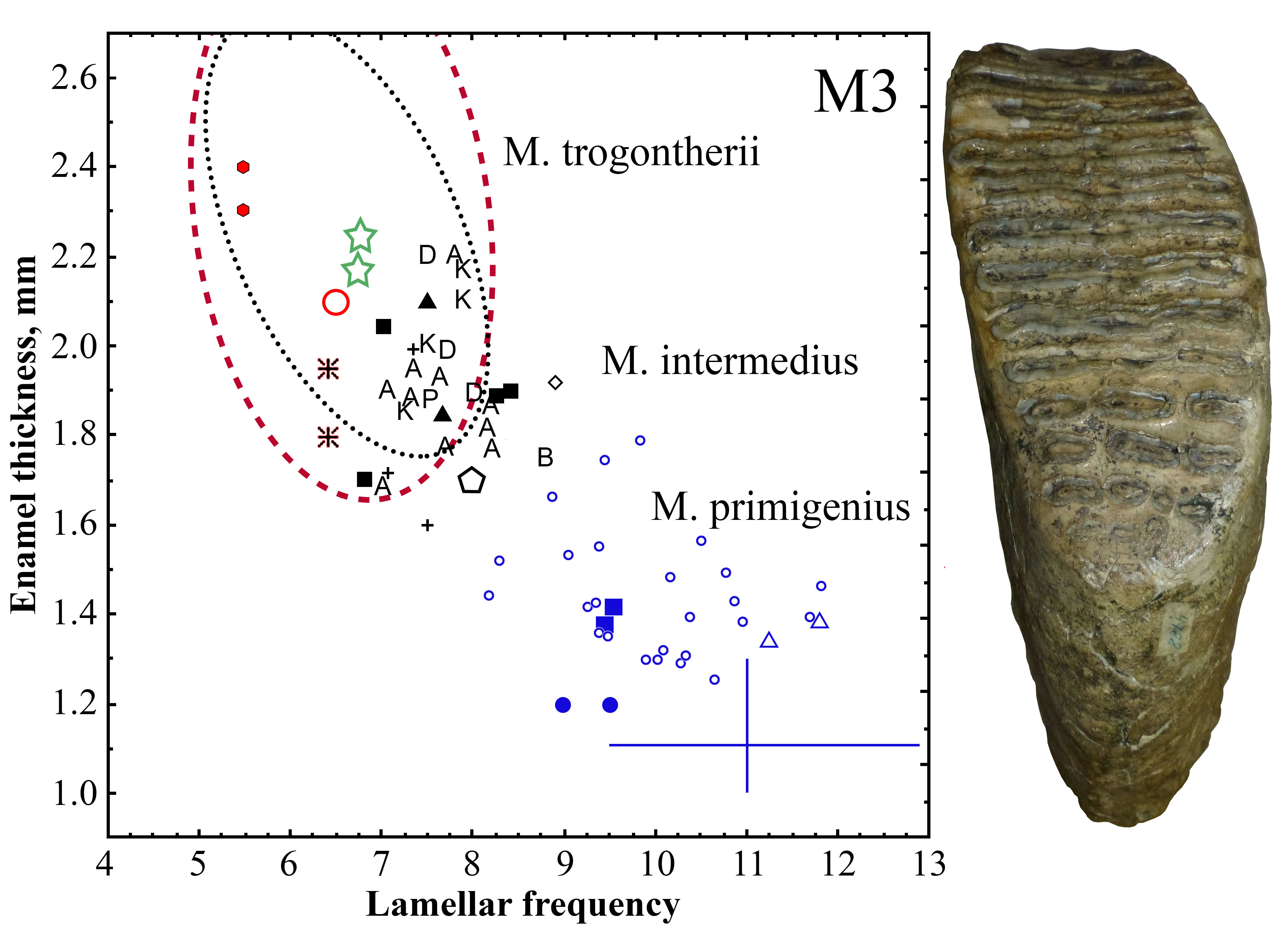
2. Materials and Methods
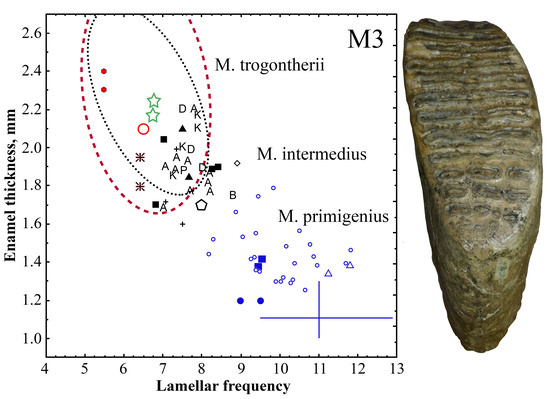
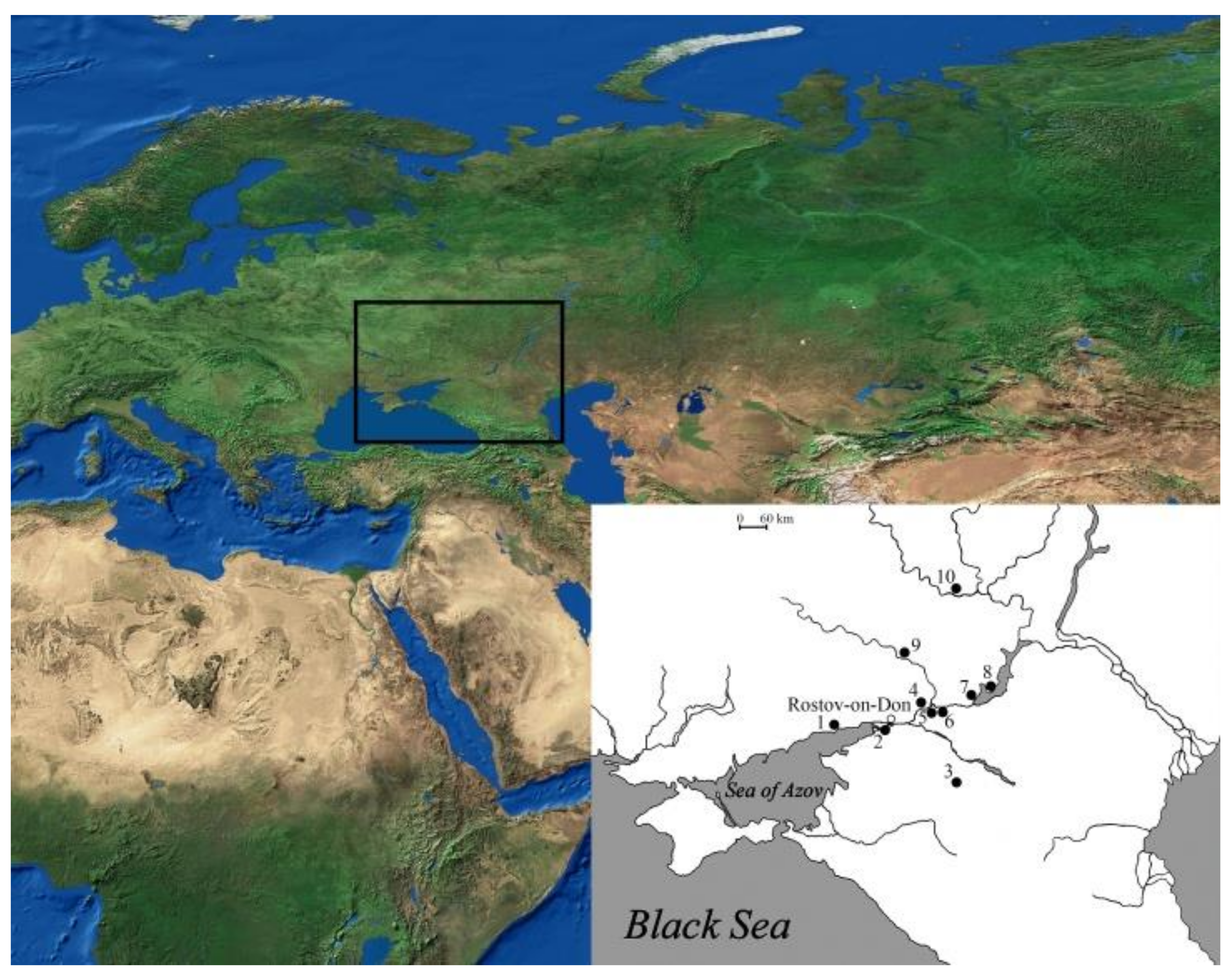
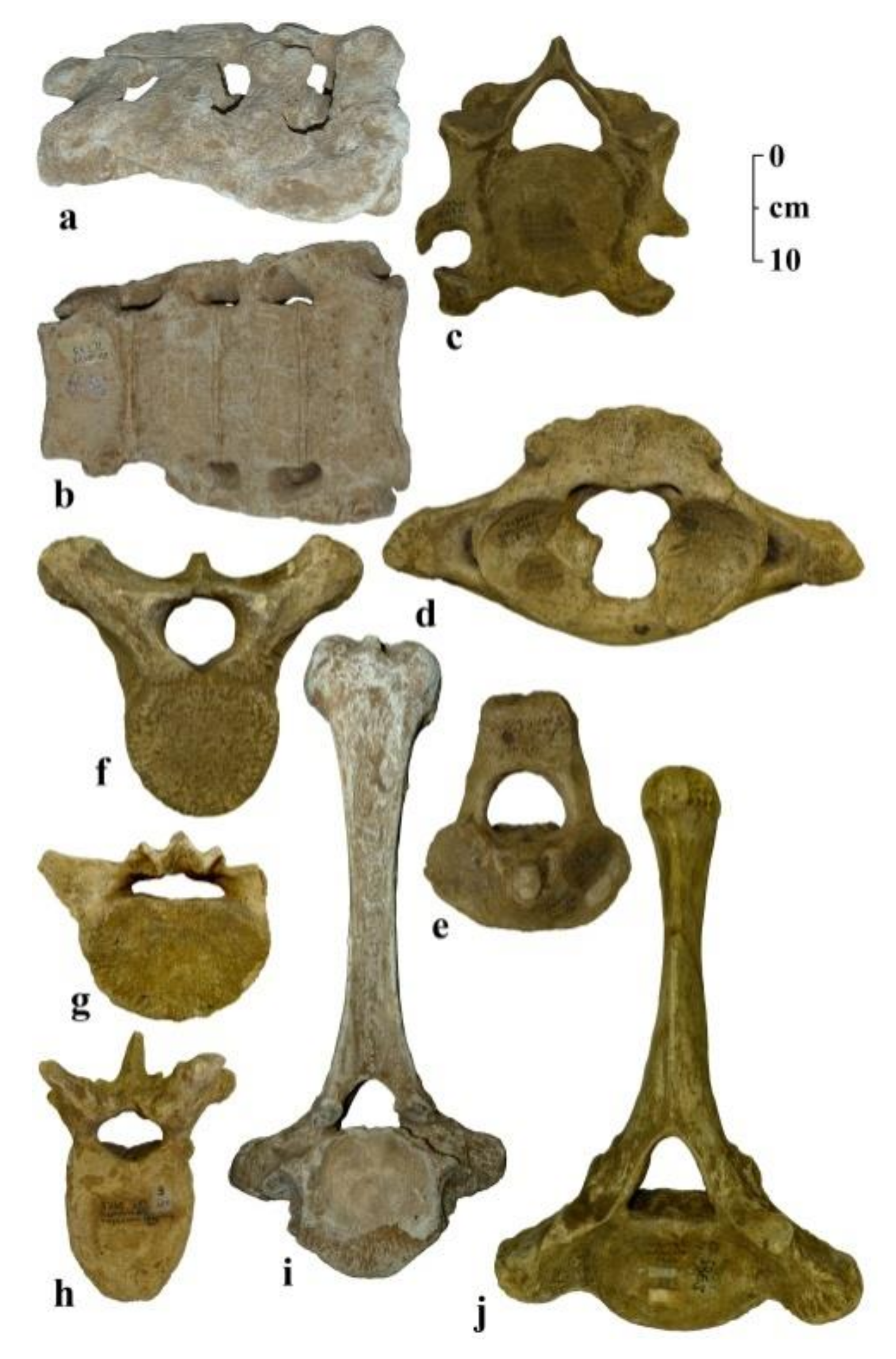
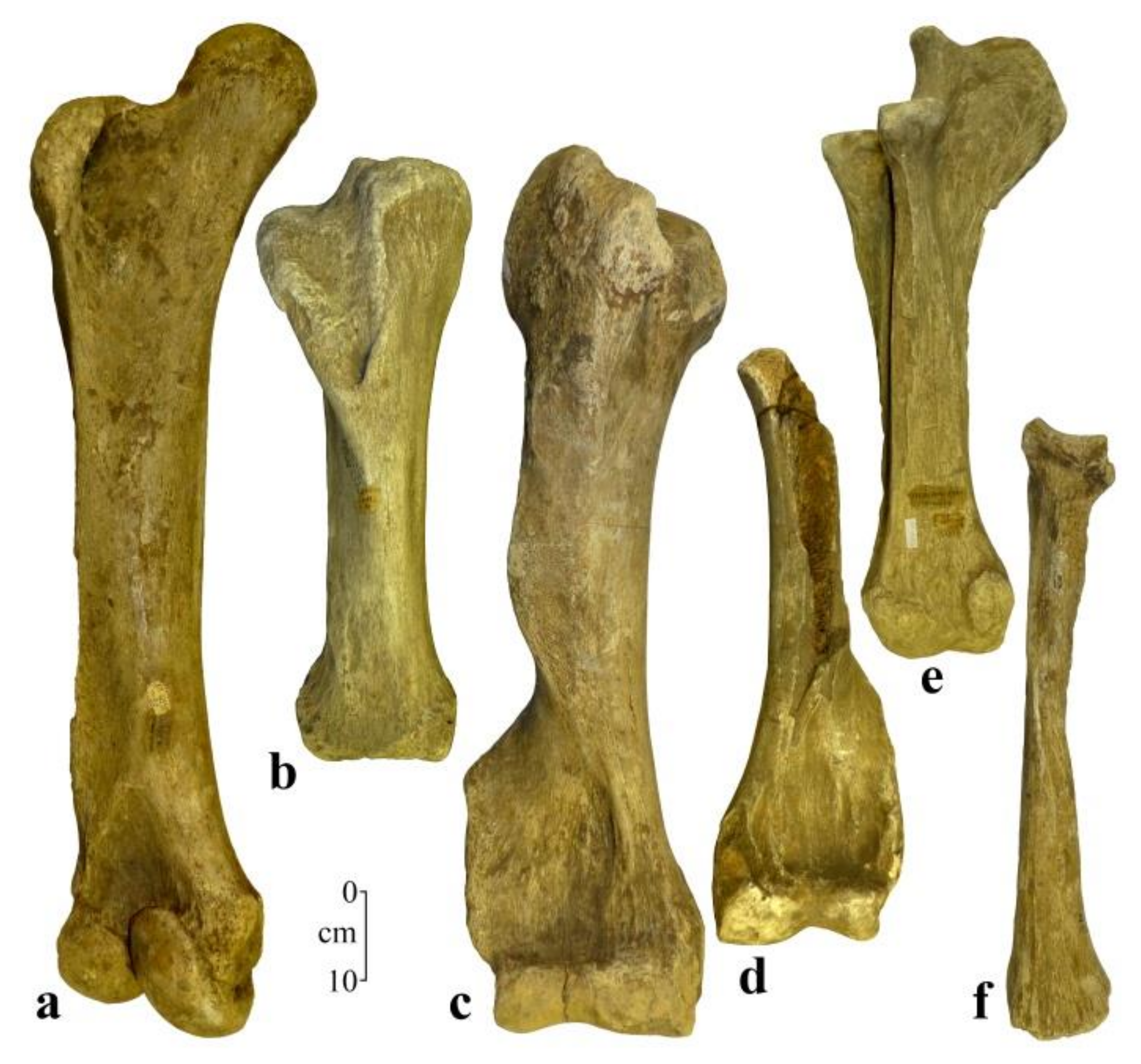
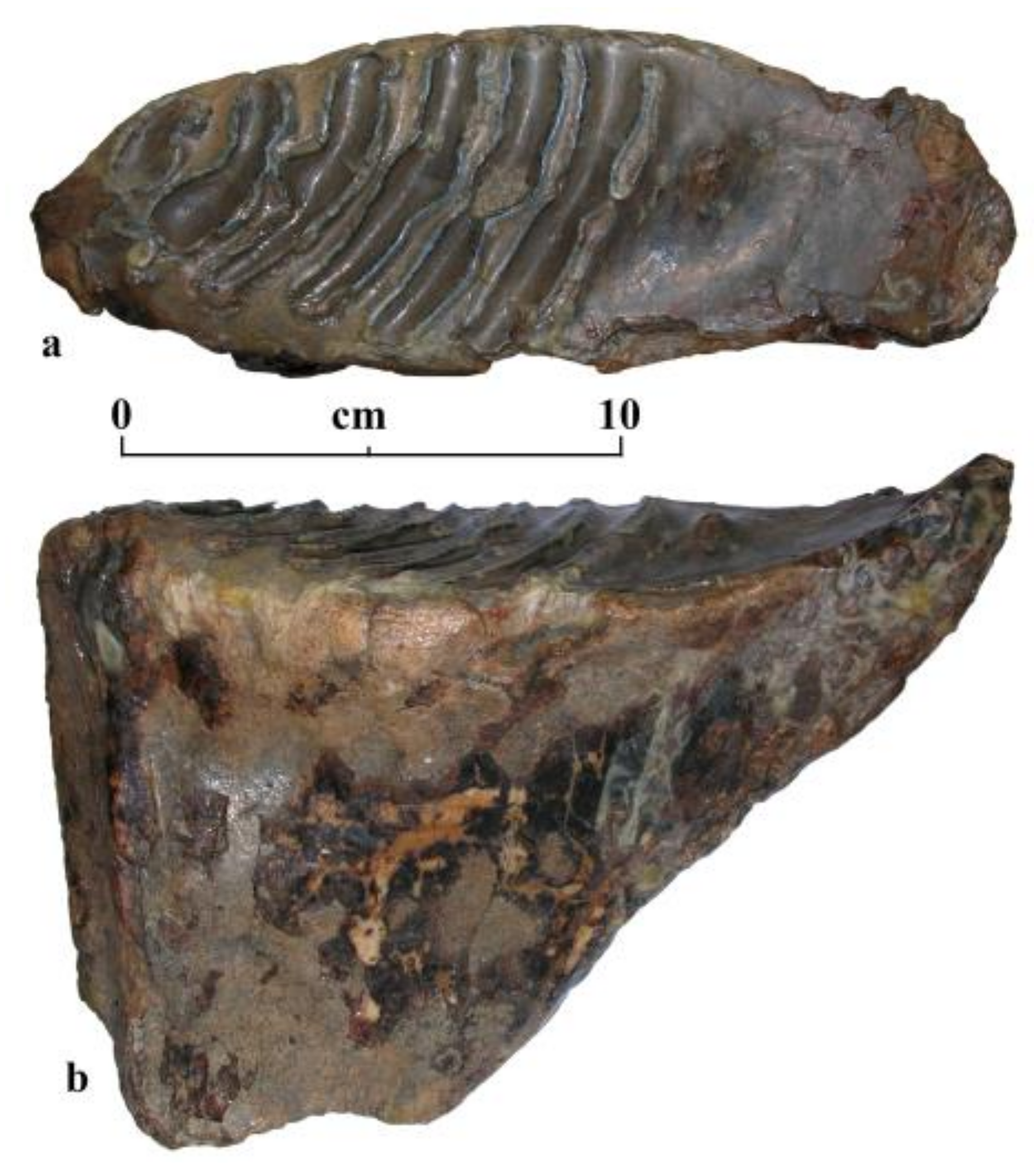
3. Results
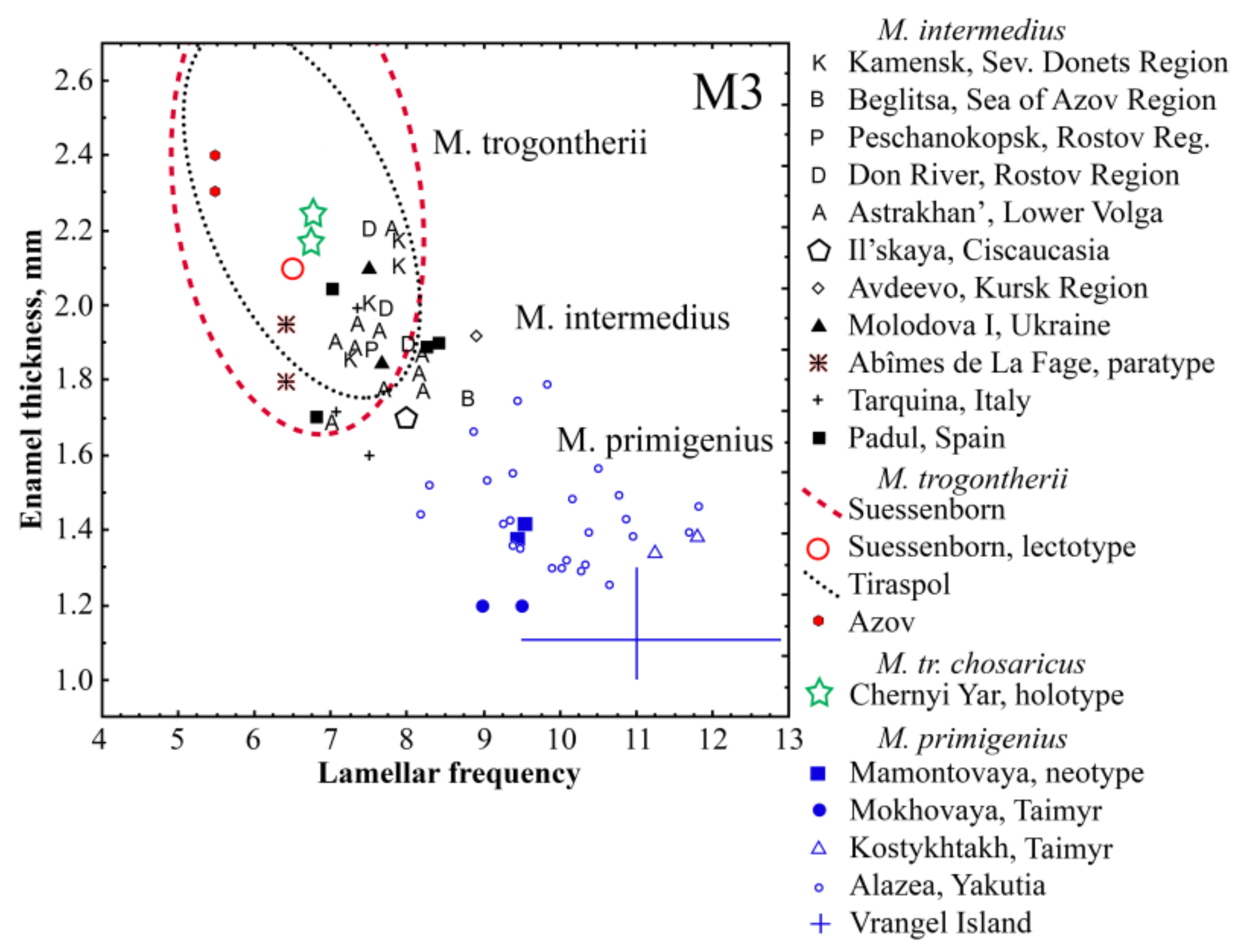
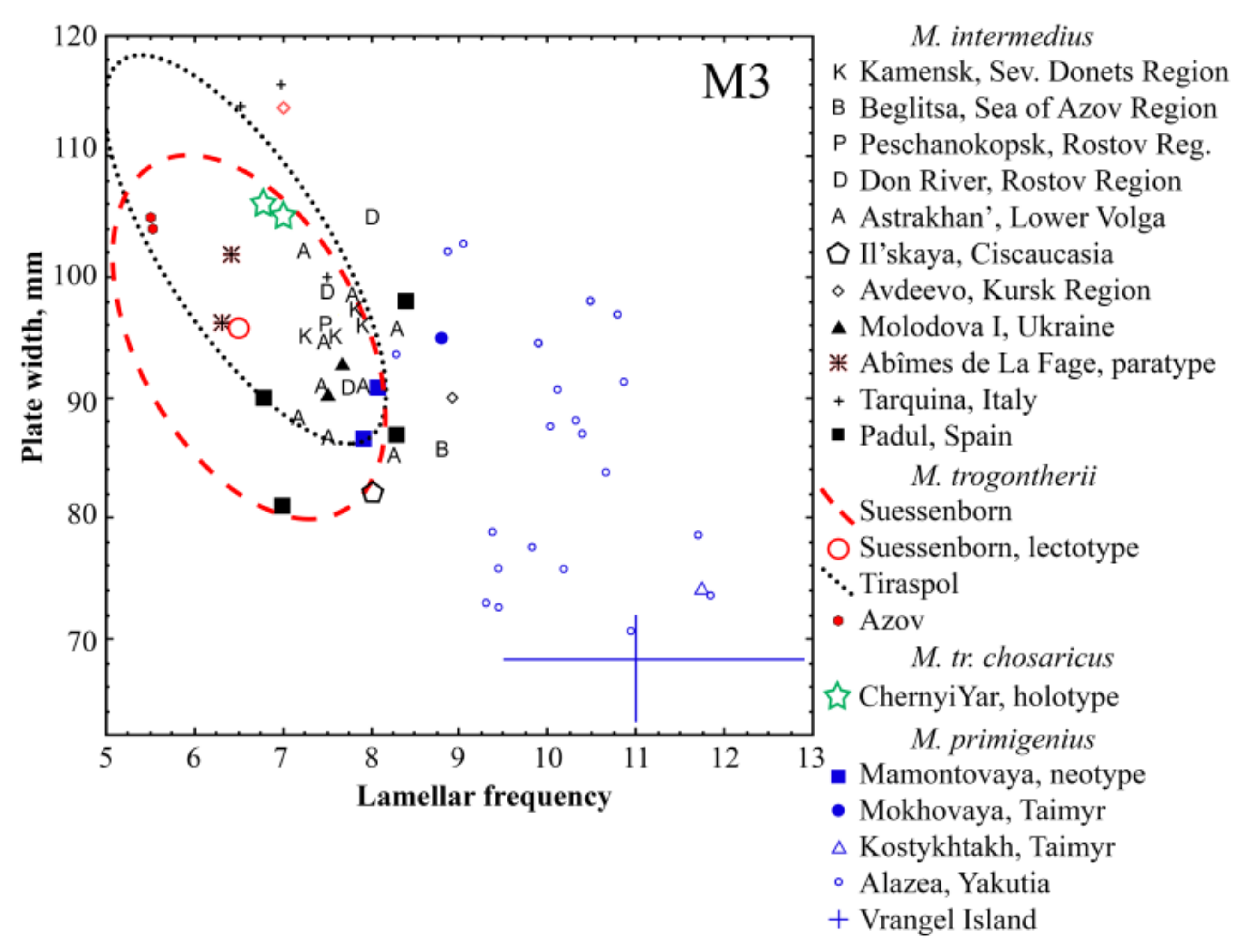
4. Comparison and Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| AMZ | Azov Historical, Archaeological, and Paleontological Museum-Reserve, Azov, Rostov Region; |
| AVIM | Aksai Military-Historical Museum, Aksai, Rostov Region; |
| NMDC | Novocherkassk Museum of History of the Don Cossacks, Novocherkassk, Rostov Region, Pal—the paleontological division of the collection; |
| PC | private collection of A.S. Basan, Taganrog, Rostov Region; |
| ROMK | Rostov Regional Studies Museum, Rostov-on-Don, Rostov Region; |
| SSC RAS | Southern Scientific Centre of the Russian Academy of Sciences, Rostov-on-Don. |
References
- Lister, A.M. Evolution and taxonomy of Eurasian mammoths. In The Proboscidea. Evolution and Palaeontology of Elephants and Their Relatives; Shoshani, J., Tassy, P., Eds.; Oxford University Press: Oxford, UK; New York, NY, USA; Tokyo, Japan, 1996; pp. 204–213. [Google Scholar]
- Lister, A.M.; Sher, A.V. The origin and evolution of the woolly mammoth. Science 2001, 294, 1094–1097. [Google Scholar] [CrossRef] [PubMed]
- Lister, A.M.; Sher, A.V.; Van Essen, H.; Wei, G. The pattern and process of mammoth evolution in Eurasia. Quat. Int. 2005, 126, 49–64. [Google Scholar] [CrossRef]
- Garutt, V.E. To the history of identifying of the intermediate links in the Archidiskodon–Mammuthus elephants phylogenetic line. In Chronology of the Glacial Age, Materials of Symposium, March 1971, Leningrad, USSR.; Geography Society of the USSR: Leningrad, Russia, 1971; pp. 78–88. [Google Scholar]
- Pavlow, M. Les elephants fossiles de la Russie. Nouv. Mem. Soc. Impér. Nat. Mosc. 1910, 17, 1–56. [Google Scholar]
- Dubrovo, I.A. Systematical position of the elephant of Khozar faunistic complex. Byull. Kom. Izuc. Chetvertichnogo Period. 1966, 32, 63–74. (In Russian) [Google Scholar]
- Garutt, V.E. The origin of elephants and ways of their phylogeny. Trudy Zoologich. Inst. AN SSSR 1986, 149, 15–32. (In Russian) [Google Scholar]
- Dietrich, W.O. Elephas primigenius fraasi, eine schwäbische Mammutraase. Jahresh. Ver. Vaterl. Naturkd. Württemberg 1912, 77, 1–64. [Google Scholar]
- Garutt, V.E.; Averianov, A.O.; Vartanyan, S.L. On the systematic position of the Holocene population of mammoths Mammuthus primigenius (Blumenbach, 1799) from Wrangel Island (Northeast Siberia). Dokl. Akad. Nauk. SSSR 1993, 332, 799–801. (In Russian) [Google Scholar]
- Foronova, I.V.; Zudin, A.N. The structure of the lineage Archidiskodon-Mammuthus in Eurasia and peculiarities of its evolution. Deinsea 1999, 6, 103–118. [Google Scholar]
- Foronova, I.V. Quaternary Mammals of the South-East of Western Siberia (Kuznetsk Basin): Phylogeny, Biostratigraphy, and Palaeoecology; Publishing House of Siberian Branch, Russian Academy of Sciences: Novosibirsk, Russia, 2001; 243p, ISBN 5-7692-0411-7. (In Russian) [Google Scholar]
- Foronova, I.V. Thin-enamel dental specialization in mammoth evolution: An example of direction selection. Quat. Int. 2007, 169, 95–104. [Google Scholar] [CrossRef]
- Jourdan, C. Des terrains sidérolitiques. CR Acad. Sci. 1861, 53, 1009–1014. [Google Scholar]
- Lortet, L.; Chantre, E. Études paléontologiques dans le Bassin du Rhône: Période quaternaire. Arch. Mus. Hist. Nat. 1872, 1, 59–130. [Google Scholar]
- Beden, M.; Guérin, C. Les Proboscidiens du gisement pléistocène moyen des Abîmes de La Fage à Noailles (Corrèze). Nouv. Arch. Mus. Hist. Nat. 1975, 13, 69–87. [Google Scholar]
- Labe, B.; Guérin, C. Réhabilitation de Mammuthus intermedius (Jourdan, 1861), un mammouth (Mammalia, Elephantidae) du Pléistocène moyen récent d’Europe. CR Palevol. 2005, 4, 235–242. [Google Scholar] [CrossRef]
- Obadă, T. Evidence of contact between Mammuthus intermedius (Jourdan, 1861) and ancient humans from Duruitoarea Veche, Republic of Moldova: Preliminary data. In Abstracts of VIth International Conference on Mammoths and their Relatives; Spec. Vol. 102; S.A.S.G.: Thessaloniki, Greece, 2014; pp. 145–146. [Google Scholar]
- Golovachev, M.V.; Titov, V.V. Remains of Mammuthus intermedius Jourdan, 1861 from the type localities of the Khazar fauna of the Lower Volga Region. Trudy Zoologich. Inst. RAN 2018, 322, 268–284. (In Russian) [Google Scholar] [CrossRef][Green Version]
- Foronova, I.V. Mammoths of the Molodova V Paleolithic site (Dniester Basin): The case of dental thin-enamel specialization and paleoecological adaptation. Quat. Int. 2014, 326, 235–242. [Google Scholar] [CrossRef]
- Baigusheva, V.S. Mammoth Mammuthus primigenius Blum. from the left bank of Severskiy Donets River. Trudy Zoologich. Inst. AN SSSR 1980, 93, 75–80. (In Russian) [Google Scholar]
- Baigusheva, V.S. About some palaeontological finds from the collections of the Museum of Don Cossacks history. Kraevedcheskie Zapiski 1999, 4, 94–115. (In Russian) [Google Scholar]
- Averianov, A.O.; Baryshnikov, G.F.; Garutt, V.E.; Garutt, N.V.; Fomicheva, N.L. The Volga Fauna of Pleistocene Mammals in the Geological and Mineralogical Museum of Kazan University; Kazan University Publishes: Kazan, Russia, 1992; 164p. (In Russian) [Google Scholar]
- Kazanov, I.A.; Titov, V.V. About the find of mammoth in the Belaya Kalitva district of Rostov Region. In Basic Problems of Quaternary, Research Results, and the Main Trends of Future Studies. Proceedings of VIII All-Russian Conference on Quaternary Research, Rostov-on-Don, Russia, 10–15 June 2013; SSC RAS Publishers: Rostov-on-Don, Russia, 2013; pp. 259–260. (In Russian) [Google Scholar]
- Zastrozhnov, A.S.; Danukalova, G.A.; Golovachev, M.V.; Titov, V.V.; Tesakov, A.S.; Simakova, A.N.; Osipova, E.M.; Trofimova, S.S.; Zynoviev, E.V.; Kurmanov, R.G. Singil Deposits in the Quaternary Scheme of the Lower Volga Region: New Data. Stratigr. Geol. Correl. 2018, 26, 647–685. [Google Scholar] [CrossRef]
- Korniets, N.L. About the reasons of mammoth’s extinction at the territory of Ukraine. In Fossil Faunae of Ukraine and Adjacent Territories; Pidoplichko, I.G., Ed.; Naukova Dumka: Kiew, USSR, 1962; pp. 93–169. (In Ukranian) [Google Scholar]
- Baryshnikov, G. Mammuthus primigenius from the Crimea and the Caucasus. Deinsea 2003, 9, 41–56. [Google Scholar]
- Zamyatnin, S.N. Notes about the Palaeolithic of the Donets Basin and Sea of Azov Region. Sbornik Muzeya Antropol. Etnographii 1953, 14, 231–255. (In Russian) [Google Scholar]
- Velichko, A.A.; Morozova, T.D.; Borisova, O.K.; Timireva, S.N.; Semenov, V.V.; Kononov, Y.M.; Titov, V.V.; Tesakov, A.S.; Konstantinov, E.A.; Kurbanov, R.N. Development of the steppe zone in Southern Russia based on the reconstruction from the loess-soil formation in the Don–Azov Region. Dokl. Earth Sci. 2012, 445, 999–1002. [Google Scholar] [CrossRef]
- Velichko, A.A.; Borisova, O.K.; Kononov, Y.M.; Konstantinov, E.A.; Kurbanov, R.N.; Morozova, T.D.; Panin, P.G.; Semenov, V.V.; Tesakov, A.S.; Timireva, S.N.; et al. Reconstruction of Late Pleistocene Events in the Periglacial Area in the Southern Part of the East European Plain. Dokl. Earth Sci. 2017, 475, 895–899. [Google Scholar] [CrossRef]
- Tesakov, A.S.; Titov, V.V.; Leonova, N.B.; Velichko, A.A.; Simakova, A.N.; Zastrozhnov, A.S.; Frolov, P.D. Quaternary Stratigraphy and Paleontology of the Southern Russia: Connections between Europe, Africa, and Asia. Programme and Guidebook of Excursions of 2010 Annual Meeting INQUA-SEQS, Rostov-on-Don, Russia, June 21–26 2010; SSC RAS Publishers: Rostov-on-Don, Russia, 2010; 48p. [Google Scholar]
- Tesakov, A.S.; Titov, V.V.; Baygusheva, V.S.; Velichko, A.A.; Timireva, S.N.; Simakova, A.N.; Borisova, O.K.; Morozova, T.D.; Semenov, V.V.; Syromyatnikova, E.V.; et al. Basic Problems of Quaternary, Research Results, and the Main Trends of Future Studies. VIII All-Russian Conference on Quaternary Research: Field Excursions Guidebook, Rostov-on-Don, Russia, 10–15 June 2013; SSC RAS Publishers: Rostov-on-Don, Russia, 2013; 48p. (In Russian) [Google Scholar]
- Byakov, A.S.; Parada, S.G.; Lev, I.E. The rare find of late Pleistocene mammoth at the south of Russia (Rostov Region). In Proceedings of the II Intern. Conference, Volume 1, Novocherkassk, Russia, 21–23 October 1999; pp. 97–98. (In Russian). [Google Scholar]
- Baygusheva, V.S.; Titov, V.V.; Tesakov, A.S.; Syromyatnikova, E.V.; Kurshakov, S.V.; Frolov, P.D. Middle Pleistocene fauna of Veshenskaya (middle Don River, Rostov Region, Russia). In The Quaternary of the Urals: Global trends and Pan-European Quaternary Records. Abstract Book of Intern. Conf., Ekaterinburg, Russia, 10–16 February 2014; UrFU Publishers: Ekaterinburg, Russia, 2014; pp. 15–17. [Google Scholar]
- Dodonov, A.E.; Tesakov, A.S.; Titov, V.V.; Inozemtcev, S.A.; Simakova, A.N.; Nikolskiy, P.A.; Trubikhin, V.M. New data on stratigraphy of Pliocene-Pleistocene deposits of lower Don River, cross sections of Tsimliansk reservoir. In Geological Events of Neogene and Quaternary of Russia: Recent Condition of Stratigraphical Schemes and Palaeogeographical Reconstructions. Proceedings of All-Russian Conference; GEOS: Moscow, Russia, 2007; pp. 43–53. (In Russian) [Google Scholar]
- Titov, V.V. Habitat conditions for Camelus knoblochi and factors in its extinction. Quat. Int. 2008, 179, 120–125. [Google Scholar] [CrossRef]
- Dubrovo, I.A. Ancient elephants of the SSSR. Trudy Paleontologich. Inst. AN SSSR 1960, 85, 78. (In Russian) [Google Scholar]
- Aguirre, E. Evolutionary history of the elephant. Science 1969, 164, 1366–1376. [Google Scholar] [CrossRef]
- Garutt, V.E.; Foronova, I.V. Research on the Teeth of Extant Elephants: Methodological Recommendations; Institute of Geology and Geophysics Publishers: Novosibirsk, Russia, 1976; p. 36. (In Russian) [Google Scholar]
- Janis, C.M. An Estimation of Tooth Volume and Hypsodonty Indices in Ungulate Mammals, and the Correlation of these Factors with Dietary Preference. Mem. Mus. Nat. Hist. Nat. Paris Ser. C 1988, 53, 367–387. [Google Scholar]
- Sher, A.V.; Garutt, V.E. New data on morphology of elephants’ molars. Dokl. Akad. Nauk. SSSR 1985, 285, 221–225. (In Russian) [Google Scholar]
- Dubrovo, I.A. Mammuthus trogontherii trogontherii (Pohlig) aus dem Pleistozän von Tiraspol. Quartärpaläontologie 1975, 1, 125–155. [Google Scholar]
- Titov, V.V.; Golovachev, M.V. The skeleton of mammoth Mammuthus trogontherii cf. chosaricus Dubrovo, 1966 from terminal Middle Pleistocene of the Lower Volga region (Russia). Russian J. Theriol. 2017, 16, 15–29. [Google Scholar] [CrossRef]
- Osborn, H.F. Proboscidea. A Monograph of the Discovery, Evolution, Migration and Extinction of the Mastodonts and Elephants of the World, Vol. II. Stegodontoidea Elaphantoidea; American Museum Press: New York, NY, USA, 1942; 828p. [Google Scholar]
- Kahlke, R.D. Zur festlegung des lectotypus von Mammuthus trogontherii (Pohlig, 1885) (Mammalia, Proboscidea). Quartärpaläontologie 1990, 8, 119–124. [Google Scholar]
- Averianov, A.O. Kutomanov’s Mammoth. Trudy Zoologich. Inst. AN SSSR. 1994, 256, 111–135. (In Russian) [Google Scholar]
- Vereshchagin, N.K. Berelekhskoe “cemetery” of mammoth. Trudy Zoologich. Inst. AN SSSR 1977, 72, 5–50. (In Russian) [Google Scholar]
- Maschenko, E.N.; Gablina, S.S.; Tesakov, A.S.; Simakova, A.N. The Sevsk woolly mammoth (Mammuthus primigenius) site in Russia: Taphonomic, biological and behavioral interpretations. Quat. Int. 2006, 142, 147–165. [Google Scholar] [CrossRef]
- Kirillova, I.V.; Shidlovskiy, F.K.; Titov, V.V. Kastykhtakh mammoth from Taimyr (Russia). Quat. Int. 2012, 276, 269–277. [Google Scholar] [CrossRef]
- Kirillova, I.V.; Borisova, O.K.; Chernova, O.F.; Van Kolfschoten, T.; van der Lubbe, J.J.L.; Panin, A.V.; Pečnerová, P.; van der Plicht, J.; Shidlovskiy, F.K.; Titov, V.V.; et al. ‘Semi-dwarf’ woolly mammoths from the East Siberian Sea coast, continental Russia. Boreas 2020, 49, 269–285. [Google Scholar] [CrossRef]
- Álvarez-Lao, D.J.; García, N. Comparative revision of the Iberian woolly mammoth (Mammuthus primigenius) record into a European context. Quat. Sci. Rev. 2012, 32, 64–74. [Google Scholar] [CrossRef]
- Hoffecker, J.F.; Baryshnikov, G.; Potapova, O. Vertebrate Remains from the Mousterian Site of Il’skaya I (Northern Caucasus, U.S.S.R.): New Analysis and Interpretation. J. Archaeol. Sci. 1991, 18, 113–147. [Google Scholar] [CrossRef]
- Obadă, T.; Ivanov, S.; Kolavenko, S.; Paşenciuc, N. New findings from the Paleolithic site of Costesti I in the north-western Moldova. Bul. Ştiinţ. Rev. Etnogr. Ştiinţ. Naturii Muzeol. (Ser. Nouă) 2018, 28, 44–54. [Google Scholar]
- Maglio, V.J. Origin and evolution of the Elephantidae. Trans. Am. Philos. Soc. 1973, 63, 149. [Google Scholar] [CrossRef]
- Garutt, V.E.; Gentry, A.; Lister, A.M. Mammuthus Brookes, 1828 (Mammalia, Proboscidea): Proposed conservation, and Elephas primigenius Blumenbach, 1799 (currently Mammuthus primigenius): Proposed designation as the type species of Mammuthus, and designation of a neotype. Bull. Zoolog. Nomencl. 1990, 47, 38–44. [Google Scholar] [CrossRef]
- Lister, A.M.; Sher, A.V. Evolution and dispersal of mammoths across the Northern Hemisphere. Science 2015, 350, 805–809. [Google Scholar] [CrossRef]
- Palombo, M.R.; Ferretti, M.P. Elephant fossil record from Italy: Knowledge, problems, and perspectives. Quat. Int. 2005, 126, 107–136. [Google Scholar] [CrossRef]
- Baygusheva, V.S.; Titov, V.V. Palaeoloxodon elephant from the Sea of Azov Region (Russia). In Abstr. 6th Meeting of the European Association of Vertebrate Palaeontologists. Slovakia, 30 June–5 July 2008; Spišská Nová: Ves, Slovakia, 2008; pp. 12–14. [Google Scholar]
- Puzachenko, A.Y.; Titov, V.V.; Kosintsev, P.A. Evolution of the European regional large mammals assemblages in the end of the Middle Pleistocene—The first half of the Late Pleistocene (MIS 6–MIS 4). Quat. Int. 2021. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Measurements | M3 | M3 | M3 | M3 | m3 | m3 | m3 |
|---|---|---|---|---|---|---|---|
| NMDC Pal-83 | NMDC Pal-83 | NMDC Pal-91 | NMDC Pal-19 | NMDC Pal-85 | NMDC Pal-85 | NMDC Pal-100 | |
| dex | sin | dex | sin | sin | dex | sin | |
| Length, mm | >248 | >245 | 303.0 | 305.0 | >280 | >248 | >209 |
| Width, mm | 95.0 | 95 | 97.6 | 96.0 | 91.8 | 90.0 | 94.0 |
| Height, mm | - | - | 170.0 | 170.0 | - | - | - |
| Tooth formula (number of plates and talons) | ~16t | ~15t | 24t | 22t | ~17t | ~17t | −10t |
| Lamellar frequency | 7.25 | 7.5 | 7.875 | 7.875 | 6.88 | 7.125 | 7.0 |
| Length of single plate, mm | 13.4 | 12.7 | 12.4 | 15.03 | 15.74 | 14.84 | 15.1 |
| Enamel thickness, mm | 1.85 | 2.0 | 2.1 | 2.17 | 2.16 | 2.2 | 2.1 |
| Wear stage | 5 | 5 | 3 | 4 | 4 | 4 | 6 |
| Measurements, mm | NMDC P-58 sin | NMDC P-59 sin, juv | NMDC P-60 dex | NMDC P-61 dex | NMDC P-68 | NMDC P-220 dex |
|---|---|---|---|---|---|---|
| Maximum length | >850 | >680 | >940 | 990 | - | - |
| Maximum width of the proximal end (at the level of epiphyseal suture) | 147.6 | 98.2 | 131.6 | 128 | - | - |
| Maximum diameter of the humeral head | - | - | - | 215 | - | - |
| Minimum width of diaphysis | 145 | 100 | 130 | 132 | - | 128 |
| Width of the distal end at the level of epiphyseal suture | 290 | 230 | 290 | 250 | (270) | 285 |
| Diameter of a distal medial condyle | - | 140 | 180 | 170 | 185 | 180 |
| Ratio of diaphysis’ width to the bone’s length | - | - | - | 13.3 | - | - |
| Measurements, mm | NMDC P-210 sin | NMDC P-213 sin | NMDC P-214 dex | NMDC P-219 sin |
|---|---|---|---|---|
| Maximum length of bone | 848 | 808 | 818 | - |
| Maximum width of the proximal end | 238 | 211 | 217 | 232 |
| Width of the articular surface for ulna | 230 | 205 | - | - |
| Maximum width of the distal end (at the level of epiphyseal suture) | - | 190 | >153 | - |
| Minimum width of diaphysis | 115 | 90 | 102 | 119.4 |
| Ratio of the width of diaphysis to the length of the bone | 13.56 | 11.14 | 12.47 | - |
| Measurements, mm | NMDC P-57 sin |
|---|---|
| Maximum length | 1130 |
| Minimum diameter of the femoral head | 160 |
| Width of the distal end at the level of epiphyseal suture | 210 |
| Minimum width of diaphysis | 148.8 |
| Width of the articular surface for the kneecap | 101.4 |
| Ratio of the width of diaphysis to the maximum length of the bone | 13.1 |
| Measurements, mm | NMDC P-185 sin | NMDC P-186 dex | NMDC P-187 sin | NMDC P-189 dex | NMDC P-188 sin | NMDC P-192 sin | NMDC P-191 sin, juv | NMDC P-190 dex |
|---|---|---|---|---|---|---|---|---|
| Maximum length | 605 | 589 | 605 | 595 | >660 | >629 | >587 | >582 |
| Maximum width of the proximal end | 219.8 | 219 | 219 | 220 | 221 | 212 | 225 | 227 |
| Maximum width of the distal end at the level of epiphyseal suture | 172 | 165 | 171.8 | 176 | 185 | 180 | 172 | 173 |
| Minimum width of diaphysis | 107 | 105 | 98 | 104.4 | 112.2 | 114.6 | 112 | 106 |
| Ratio of the width of diaphysis to the maximum length of the bone | 17.69 | 17.83 | 16.20 | 17.55 | - | - | - | - |
| Measurements | Beglitsa | Peschanokopskoe | Peschanokopskoe | Vyoshenskaya | Vyoshenskaya | Vyoshenskaya | Tsymlyansk | Tsymlyansk | Don River | Don River | Don River | Don River | Don River | Razdorskaya |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| M3 dex | M3 sin | m3 sin | m1 sin | m2 sin | m2 dex | m3 sin | M1–2 | M3 sin | M3 sin | M3 dex | m2 dex | m3 dex | m3 sin | |
| PC | Coll. ROMK | Coll. AMZ, KP-27350/1b | ROMK M-1757 | ROMK M-1757 | – | AMZ KP-27500/1 | SSC RAS No no | NMDC no No | NMDC-KP-10576 | NMDC no No | NMDC no No | AVIM-290 | Coll. ROMK | |
| Length, mm | 226 | >212 | >211.5 | – | 260 | – | >267 | – | >270 | 270 | >203 | 255 | >294 | >197 |
| Width, mm | 85.8 | 95.8 | 94 | 72 | 82 | 76 | 75 | 93 | 99 | 105 | 91 | 101 | 95.4 | 73.6 |
| Height, mm | 149 13 | – | – | – | 114 | 101 | 146 | 166 | – | 152 | – | – | – | |
| Tooth formula (number of plates and talons) | t21t | ~13t | ~11t | ~11t | t16t | –15t | t7– | ~21t | 23t | ~15t | 18t | ~19t | ~10t | |
| Lamellar frequency | 8.8 | 7.5 | 7.0 | 8.0 | 8.0 | 8.0 | 8.0 | 7.75 | 7.5 | 8.0 | 7.75 | 8.25 | 6.7 | 6.0 |
| Length of single plate, mm | 10.99 | 13.82 | 15.4 | 12.6 | 15.0 | 15.0 | ||||||||
| Enamel thickness, mm | 1.75 | 1.88 | 1.7 | 2.0 | 2.2 | 2.0 | 1.7 | 2.1 | 2.2 | 1.9 | 2.0 | 1.7 | 1.8 | 2.1 |
| Wear stage | 4 | 5–6 | 6 | 6 | 1 | 4–5 | 3 | 1 | 5 | 4 | 5–6 | 4–5 | 4 | 6 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Baigusheva, V.S.; Titov, V.V. Late Middle—Early Late Pleistocene Mammoths from the Lower Don River Region (Russia). Quaternary 2021, 4, 5. https://doi.org/10.3390/quat4010005
Baigusheva VS, Titov VV. Late Middle—Early Late Pleistocene Mammoths from the Lower Don River Region (Russia). Quaternary. 2021; 4(1):5. https://doi.org/10.3390/quat4010005
Chicago/Turabian StyleBaigusheva, Vera S., and Vadim V. Titov. 2021. "Late Middle—Early Late Pleistocene Mammoths from the Lower Don River Region (Russia)" Quaternary 4, no. 1: 5. https://doi.org/10.3390/quat4010005
APA StyleBaigusheva, V. S., & Titov, V. V. (2021). Late Middle—Early Late Pleistocene Mammoths from the Lower Don River Region (Russia). Quaternary, 4(1), 5. https://doi.org/10.3390/quat4010005