
Disentangling Domestication from Food Production Systems in the Neotropics

 , ,
, ,  ,
,  , ,
, , 
 ,
, 
 ,
,  , , ,
, , ,
Abstract
:1. Introduction
2. Definitions of Concepts
2.1. Domestication as Process
2.2. Domestication as Result (the Domestication Syndrome)
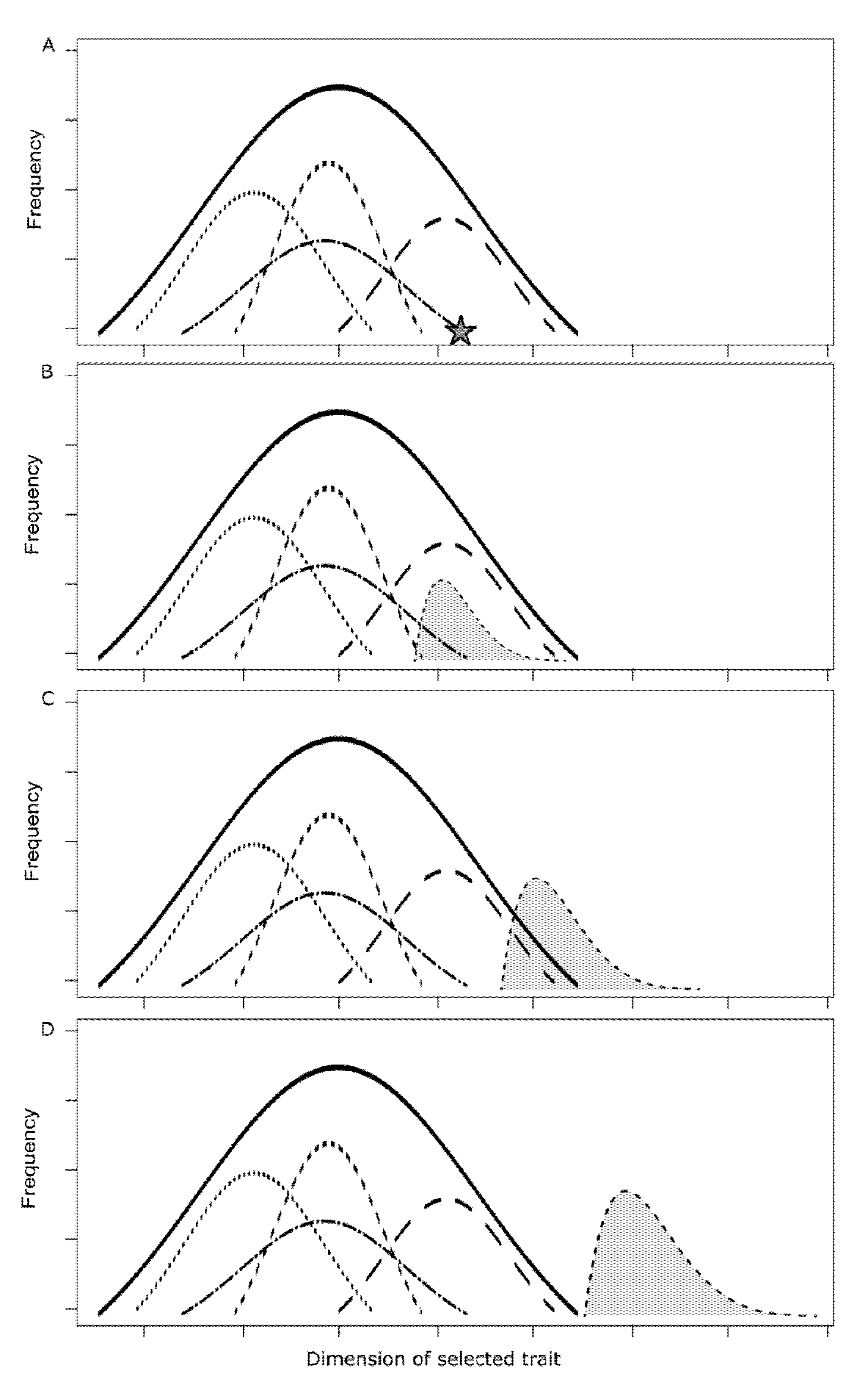
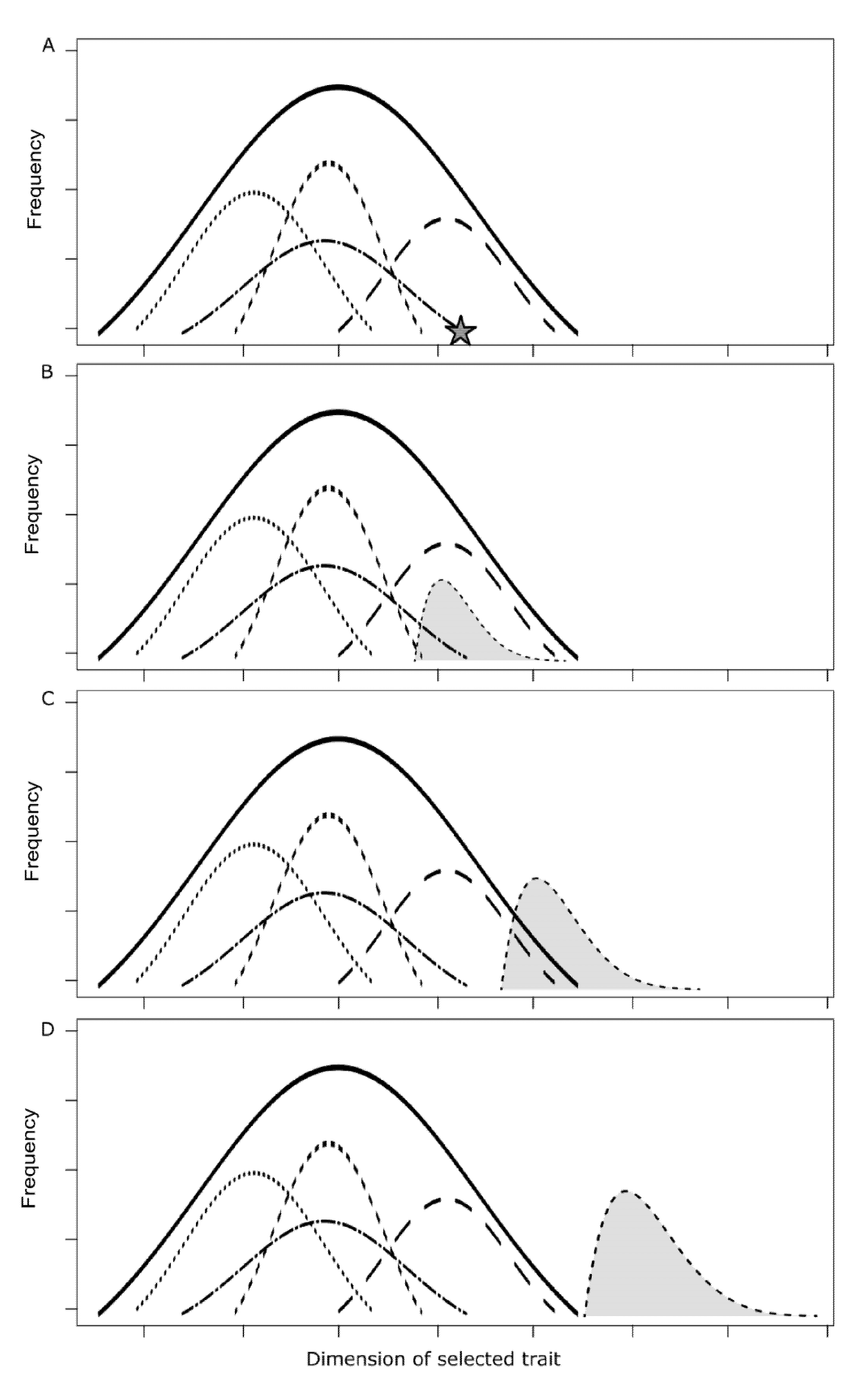
2.3. Food Production Systems
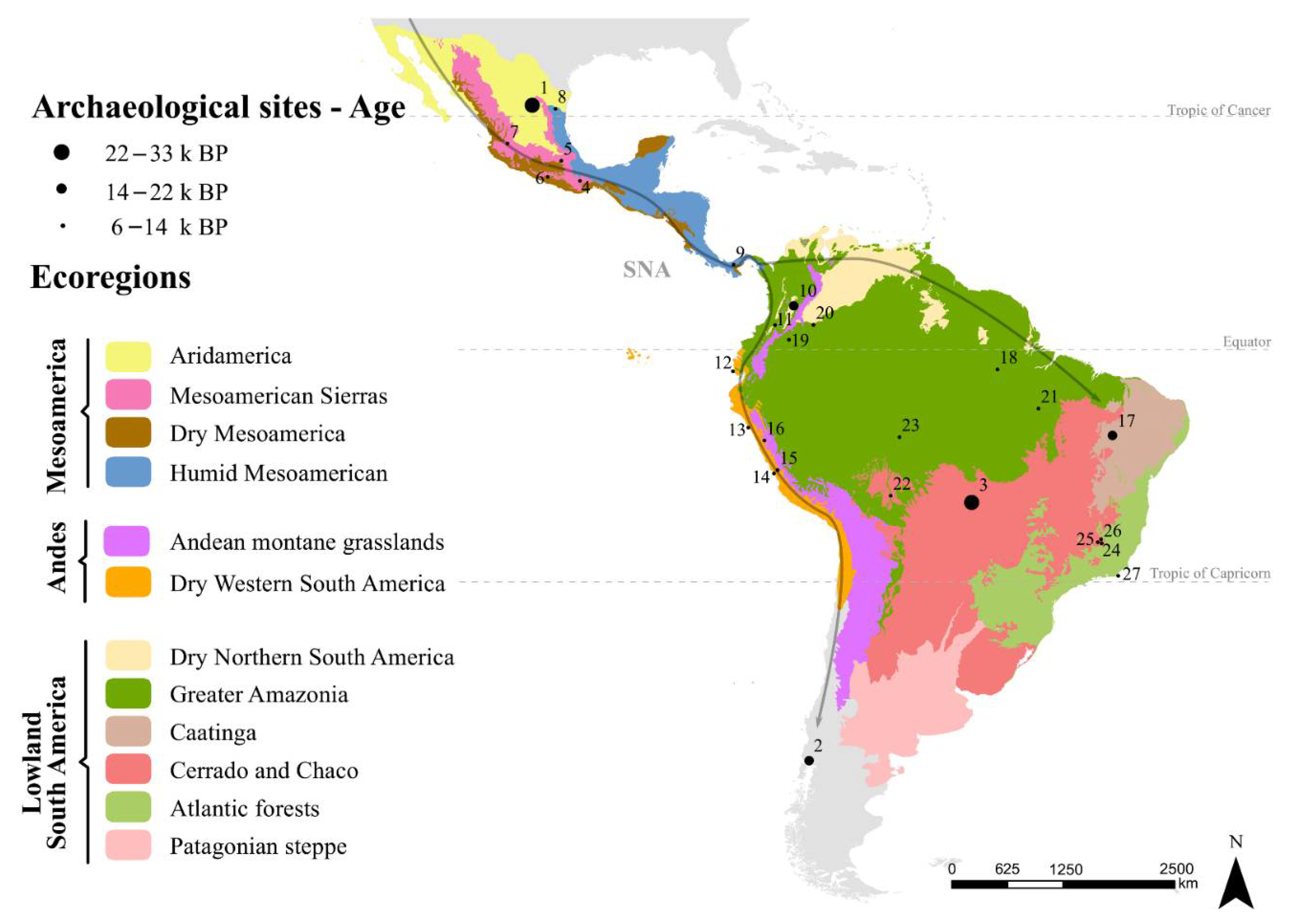
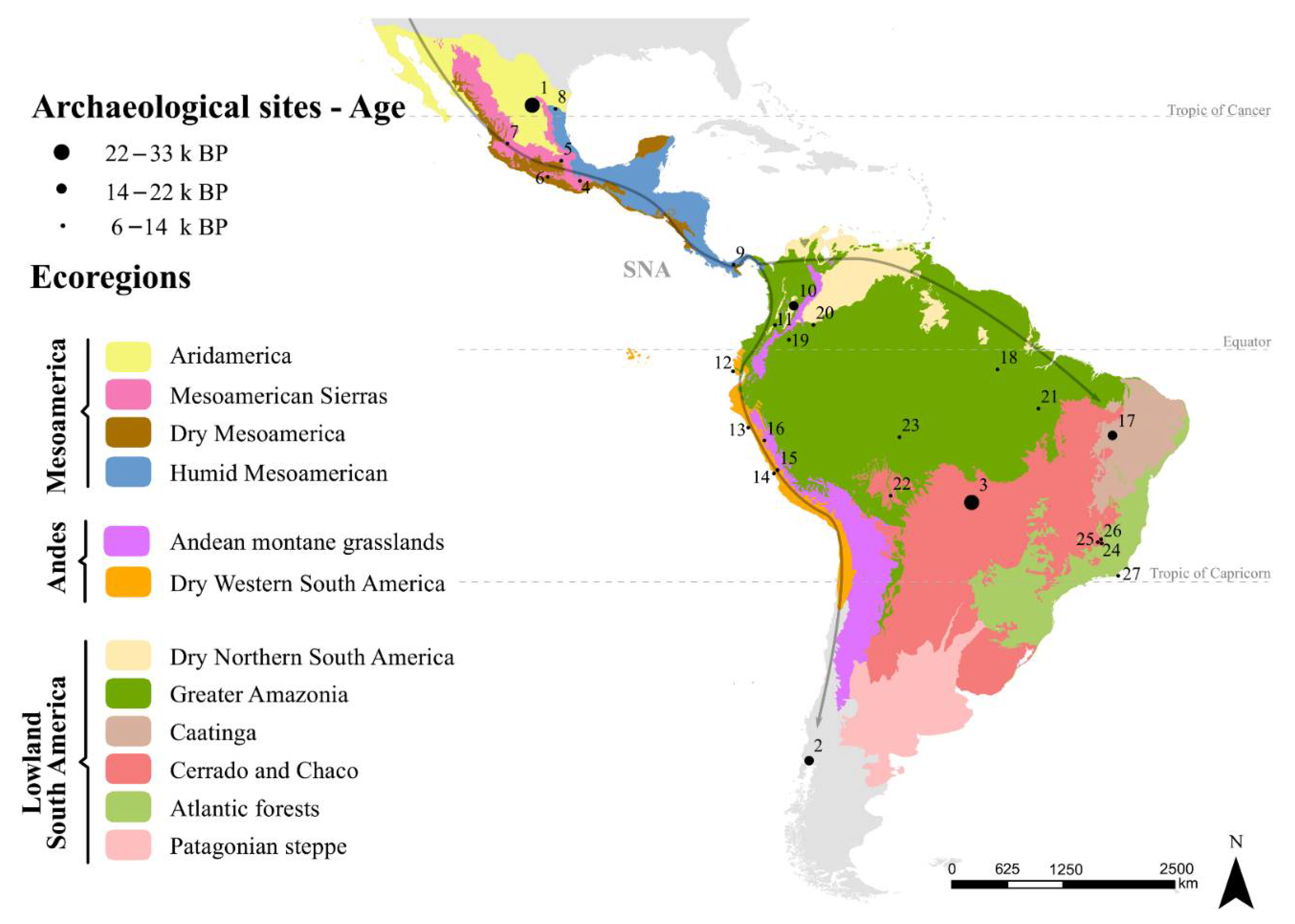
3. Early Human Entanglements in the Neotropics
4. Expansion of Plant × Human Entanglements and Domestications
4.1. Mesoamerica (Mexico and Central America)
4.2. Central Andes
4.3. South American Lowlands
4.4. Comparisons among Regions
5. Are the Neotropics Different?
6. Post-Script
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
| Definition | Author (Year) |
|---|---|
| Domestication is a co-evolutionary process that occurs when wild plants are brought into cultivation by humans. | Purugganan [270] |
| Domestication is the process of heritable genetic adaptation to human cultivation and consumption conditions. | Hufford et al. [271] |
| Domestication is an evolutionary interaction where a producer species gains new dispersal mechanisms while its performance is controlled for the benefit (commonly nutritional) of a consumer species. | Milla et al. [272] |
| Domestication can be generally considered a selection process for adaptation to human agroecological niches and, at some point in the process, human preferences. | Larson et al. [273] |
| Domestication is an evolutionary process driven by natural and human (whether conscious or unconscious) selection applied to wild plants or animals and leading to adaptation to cultivation and consumption or utilization. | Gepts [274] |
| Domestication describes genetic and morphological changes on the part of a plant population in response to selective pressures imposed by cultivation. | Fuller and Hildebrand [275] |
| These sustaining crop plants were derived, in most cases, by several thousand years or more of conscious as well as unintentional human selection, in the process transforming mostly unremarkable wild ancestors into high-yielding and otherwise useful domesticated descendants. | Olsen and Wendel [276] |
| Here, “domesticated” refers more generally to plants that are morphologically and genetically distinct from their wild ancestors as a result of artificial selection, or are no longer known to occur outside of cultivation. We define “semidomesticated” as a crop that is under cultivation and subjected to conscious artificial selection pressures. | Meyer et al. [25] |
| “Domesticated species” are those that have been genetically altered through artificial selection such that phenotypic characteristics distinguish them from wild progenitors. | Piperno [277] |
| Domestication is the outcome of a selection process that leads to plants adapted to cultivation and utilization by humans. | Brown et al. [278] |
| Domestication is generally considered to be the end-point of a continuum that starts with the exploitation of wild plants, continues through the cultivation of plants selected from the wild but not yet genetically different from wild plants, and terminates in the fixation, through human selection, of morphological and hence genetic differences distinguishing a domesticate from its wild progenitor. | Pickersgill [279] |
| Domestication is most often defined in terms of two salient characteristics: first, that the newly created “species” is observably distinct from its wild relatives; and second, that without continued human protection, it would cease to exist. | Smith [280] |
| Domestication is best viewed as an evolving of mutualism between humans and populations of plants or animals. | Zeder [56] |
| In scientific usage, “domestication” has come to mean the process by which humans transformed wild animals and plants into more useful products through control of their breeding. | Leach [58] |
| By a domesticate, I mean a species bred in captivity and thereby modified from its wild ancestors in ways making it more useful to humans who control its reproduction and (in the case of animals) its food supply. | Diamond [281] |
Appendix B
| No. | Name | References Cited |
|---|---|---|
| 1 | Chiquihuite Cave | Ardelean et al. [105]; Becerra-Valdivia and Higham [282] |
| 2 | Monte Verde | Dillehay et al. [109,137] |
| 3 | Santa Elina | Vialou et al. [110]; Scheel-Ybert and Bachelet [111] |
| 4 | GuiláNaquitz Cave | McClung de Tapia [123]; Smith [130] |
| 5 | Tehuacán Valley | Smith [128]; McClung de Tapia [123]; Debouck [126] |
| 6 | Guerrero | Piperno et al. [124] |
| 7 | Jalisco | Moreno-Letelier et al. [27] |
| 8 | Tamaulipas | McClung de Tapia [123] |
| 9 | Panama | McClung de Tapia [123]; Piperno and Pearsall [40] |
| 10 | Pubenza | van der Hammen and Urrego [129] |
| 11 | Popayán | Aceituno and Loaiza [130] |
| 12 | Santa Elena | Piperno and Stothert [132] |
| 13 | HuacaPrieta | Dillehay et al. [135] |
| 14 | Chilca Valley | Pearsall [134] |
| 15 | Tres Ventanas Cave | Pearsall [134] |
| 16 | Guitarrero Cave | Pearsall [134]; Debouck [126] |
| 17 | Serra da Capivara | Chaves [154]; Chaves and Reinhard [155]; Lahaye et al. [138] |
| 18 | Pedra Pintada | Roosevelt et al. [148]; Roosevelt [139,147]; Shock and Moraes [149] |
| 19 | Peña Roja | Morcote-Ríos et al. [141,142,145] |
| 20 | Cerro Azul | Morcote-Ríos et al. [144] |
| 21 | Carajás | Magalhães [150]; Santos et al. [151] |
| 22 | Llanos de Mojos | Lombardo et al. [152] |
| 23 | Teotônio | Watling et al. [153] |
| 24 | Lapa do Santo | Ortega [156] |
| 25 | Lapa Grande do Taquaraçu | Angeles Flores [157]; Angeles Flores et al. [158] |
| 26 | Santana do Riacho | Resende and Prous [159] |
| 27 | Forte Shell mound | Scheel-Ybert [160]; Scheel-Ybert and Boyadjian [161] |
References
- Ehrlich, P.R.; Ehrlich, A.H. The Dominant Animal: Human Evolution and the Environment; Island Press: Washington, DC, USA, 2008; p. 428. [Google Scholar]
- Harlan, J.R. Crops & Man, 2nd ed.; American Society of Agronomy & Crop Science Society of America: Madison, WI, USA, 1992. [Google Scholar]
- Scott, J.C. Against the Grain: A Deep History of the Earliest States; Yale University Press: New Haven, CT, USA, 2017; p. 336. [Google Scholar]
- Lien, M.E.; Swanson, H.A.; Ween, G.B. Naming the beast-exploring the otherwise. In Domestication Gone Wild: Politicas and Practices of Multispecies Relations; Swanson, H.A., Lien, M.E., Ween, G.B., Eds.; Duke University Press: Durham, NC, USA, 2018; pp. 1–30. [Google Scholar]
- Rindos, D. The Origins of Agriculture: An Evolutionary Perspective; Academic Press: San Diego, CA, USA, 1984; p. 325. [Google Scholar]
- Allaby, R.G.; Fuller, D.Q.; Brown, T.A. The genetic expectations of a protracted model for the origins of domesticated crops. Proc. Natl. Acad. Sci. USA 2008, 105, 13982–13986. [Google Scholar] [CrossRef] [Green Version]
- Allaby, R.G.; Kitchen, J.L.; Fuller, D.Q. Surprisingly low limits of selection in plant domestication. Evol. Bioinform. Online 2015, 11, 41–51. [Google Scholar] [CrossRef]
- Allaby, R.G.; Stevens, C.; Lucas, L.; Maeda, O.; Fuller, D.Q. Geographic mosaics and changing rates of cereal domestication. Philos. Trans. R. Soc. B Biol. Sci. 2017, 372, 20160429. [Google Scholar] [CrossRef] [Green Version]
- Rowley-Conwy, P.; Layton, R. Foraging and farming as niche construction: Stable and unstable adaptations. Philos. Trans. R. Soc. B Biol. Sci. 2011, 366, 849–862. [Google Scholar] [CrossRef]
- Fuller, D.Q.; Asouti, E.; Purugganan, M.D. Cultivation as slow evolutionary entanglement: Comparative data on rate and sequence of domestication. Veg. Hist. Archaeobot. 2011, 21, 131–145. [Google Scholar] [CrossRef]
- Fuller, D.Q.; Stevens, C.; Lucas, L.; Murphy, C.; Qin, L. Entanglements and entrapments on the pathway toward domestication. In The Archaeology of Entanglement; Der, L., Fernandini, F., Eds.; Left Coast Press: Walnut Creek, CA, USA, 2016; pp. 151–172. [Google Scholar]
- Denevan, W.M. Cultivated Landscapes of Native Amazonia and the Andes; Oxford University Press: Oxford, UK, 2001; p. 396. [Google Scholar]
- Harris, D.R. Evolution of agroecosystems: Biodiversity, origins, and differential development. In Biodiversity in Agriculture: Domestication, Evolution, and Sustainability; Gepts, P., Famula, T.R., Bettinger, R.L., Brush, S.B., Damania, A.B., McGuire, P.E., Qualset, C.O., Eds.; Cambridge University Press: Cambridge, UK, 2012; pp. 21–56. [Google Scholar]
- Casas, A.; Caballero, J.; Mapes, C.; Zárate, S. Manejo de la vegetación, domesticación de plantas y origen de la agricultura en Mesoamérica. Bot. Sci. 1997, 61, 31–47. [Google Scholar] [CrossRef]
- Casas, A.; Otero-Arnaiz, A.; Pérez-Negrón, E.; Valiente-Banuet, A. In situ management and domestication of plants in Mesoamerica. Ann. Bot. 2007, 100, 1101–1115. [Google Scholar] [CrossRef] [Green Version]
- Smith, B.D. Niche construction and the behavioral context of plant and animal domestication. Evol. Anthropol. 2007, 16, 188–199. [Google Scholar] [CrossRef]
- Smith, B.D. General patterns of niche construction and the management of ‘wild’ plant and animal resources by small-scale pre-industrial societies. Philos. Trans. R. Soc. Lond. B 2011, 366, 836–848. [Google Scholar] [CrossRef] [Green Version]
- Casas, A.; Blancas, J.; Otero-Arnaiz, A.; Cruse-Sanders, J.; Lira, R.; Avendaño, A.; Parra, F.; Guillén, S.; Figueredo, C.J.; Torres, I.; et al. Evolutionary ethnobotanical studies of incipient domestication of plants in Mesoamerica. In Ethnobotany of Mexico; Lira, R., Casas, A., Blancas, J., Eds.; Springer: New York, NY, USA, 2016; pp. 257–285. [Google Scholar] [CrossRef]
- Zeder, M.A.; Bradley, D.G.; Emshwiller, E.; Smith, B.D. Documenting Domestication: New Genetic and Archaeological Paradigms; University of California Press: Berkeley, CA, USA, 2006; p. 361. [Google Scholar]
- Cassino, M.F.; Alves, R.P.; Levis, C.; Watling, J.; Junqueira, A.B.; Shock, M.P.; Ferreira, M.J.; Andrade, V.L.C.; Furquim, L.P.; Coelho, S.D.; et al. Ethnobotany and ethnoecology applied to historical ecology. In Methods and Techniques in Ethnobiology and Ethnoecology; Albuquerque, U.P., Lucena, R.F.P., Cunha, L.V.F.C., Alves, R.R.N., Eds.; Springer: New York, NY, USA, 2019; pp. 187–208. [Google Scholar] [CrossRef]
- Reis, M.S.; Montagna, T.; Mattos, A.G.; Filippon, S.; Ladio, A.H.; Marques, A.d.C.; Zechini, A.A.; Peroni, N.; Mantovani, A. Domesticated landscapes in Araucaria Forests, southern Brazil: A multispecies local conservation-by-use system. Front. Ecol. Evol. 2018, 6, 11. [Google Scholar] [CrossRef] [Green Version]
- Willcox, G. The beginnings of cereal cultivation and domestication in Southwest Asia. In A Companion to the Archaeology of the Ancient Near East; Potts, D.T., Ed.; Blackwell: Oxford, UK, 2012; Volume 1, pp. 163–180. [Google Scholar]
- Darwin, C. On the Origin of Species by Means of Natural Selection, or the Preservation of the Favoured Races in the Struggle for Life; John Murray: London, UK, 1859. [Google Scholar]
- Hammer, K. Das domestikationssyndrom. Die Kult. 1984, 32, 11–34. [Google Scholar] [CrossRef]
- Meyer, R.S.; DuVal, A.E.; Jensen, H.R. Patterns and processes in crop domestication: An historical review and quantitative analysis of 203 global food crops. New Phytol. 2012, 196, 29–48. [Google Scholar] [CrossRef]
- Zhou, Y.; Muyle, A.; Gaut, B.S. Evolutionary genomics and the domestication of grapes. In The Grape Genome; Cantu, D., Walker, M.A., Eds.; Springer Nature: Cham, Switzerland, 2019; pp. 39–55. [Google Scholar] [CrossRef]
- Moreno-Letelier, A.; Aguirre-Liguori, J.A.; Pinero, D.; Vazquez-Lobo, A.; Eguiarte, L.E. The relevance of gene flow with wild relatives in understanding the domestication process. R. Soc. Open Sci. 2020, 7, 191545. [Google Scholar] [CrossRef] [Green Version]
- Allaby, R.G.; Ware, R.L.; Kistler, L. A re-evaluation of the domestication bottleneck from archaeogenomic evidence. Evol. Appl. 2019, 12, 29–37. [Google Scholar] [CrossRef] [Green Version]
- Brown, T.A. Is the domestication bottleneck a myth? Nat. Plants 2019, 5, 337–338. [Google Scholar] [CrossRef]
- Smith, O.; Nicholson, W.V.; Kistler, L.; Mace, E.; Clapham, A.; Rose, P.; Stevens, C.; Ware, R.; Samavedam, S.; Barker, G.; et al. A domestication history of dynamic adaptation and genomic deterioration in Sorghum. Nat. Plants 2019, 5, 369–379. [Google Scholar] [CrossRef] [Green Version]
- McKey, D.; Elias, M.; Pujol, B.; Duputié, A. Ecological approaches to crop domestication. In Biodiversity in Agriculture: Domestication, Evolution, and Sustainability; Gepts, P., Famula, T.R., Bettinger, R.L., Brush, S.B., Damania, A.B., McGuire, P.E., Qualset, C.O., Eds.; Cambridge University Press: Cambridge, UK, 2012; pp. 377–406. [Google Scholar]
- Pollan, M. The Botany of Desire: A Plant’s-Eye View of the World; Random House: New York, NY, USA, 2001; p. 271. [Google Scholar]
- van der Veen, M. The materiality of plants: Plant–people entanglements. World Arch. 2014, 46, 799–812. [Google Scholar] [CrossRef]
- Ardren, T.; Miller, S. Household garden plant agency in the creation of Classic Maya social identities. J. Anthropol. Archaeol. 2020, 60. [Google Scholar] [CrossRef]
- Viveiros de Castro, E. The transformation of objects into subjects in Amerindian ontologies. Common Knowl. 2004, 10, 463–485. [Google Scholar] [CrossRef]
- Costa, L.; Fausto, C. The return of the animists: Recent studies of Amazonian ntologies. Relig. Soc. 2010, 1, 89–109. [Google Scholar] [CrossRef]
- Fausto, C.; Neves, E.G. Was there ever a Neolithic in the Neotropics? Plant familiarisation and biodiversity in the Amazon. Antiquity 2018, 92, 1604–1618. [Google Scholar] [CrossRef] [Green Version]
- Cabral de Oliveira, J. Mundos de roças e florestas. Bol. Mus. Para. Emílio Goeldi Ciênc. Hum. 2016, 11, 115–131. [Google Scholar] [CrossRef] [Green Version]
- Descola, P. Landscape as transfiguration. Suom. Antropol. J. Finn. Anthropol. Soc. 2016, 41, 3–14. [Google Scholar]
- Piperno, D.R.; Pearsall, D.M. The Origins of Agriculture in the Lowland Neotropics; Academic Press: San Diego, CA, USA, 1998; p. 400. [Google Scholar]
- Iriarte, J. Narrowing the gap-exploring the diversity of early food production economies in the Americas. Curr. Anthropol. 2009, 50, 677–680. [Google Scholar] [CrossRef]
- Barreto, C. Social complexity in ancient Amerindian societies: Perspectives from the Brazilian lowlands. In Against Typological Tyranny in Archaeology; Gnecco, C., Langebaek, C.H., Eds.; Springer: New York, NY, USA, 2014; pp. 1–23. [Google Scholar] [CrossRef]
- Kirchoff, P. Mesoamérica: Sus límites geográficos, composición étnica y caracteres culturales. Acta Am. 1943, 1, 92–107. [Google Scholar]
- Matos-Moctezuma, E. Mesoamérica. In Historia Antigua de México; Manzanilla, L., López Luján, L., Eds.; Instituto Nacional de Antropología e Historia: Ciudad de México, Mexico, 1994; pp. 49–73. [Google Scholar]
- Watling, J.; Iriarte, J.; Mayle, F.E.; Schaan, D.; Pessenda, L.C.R.; Loader, N.J.; Street-Perrott, F.A.; Dickau, R.E.; Damasceno, A.; Ranzi, A. Impact of pre-Columbian “geoglyph” builders on Amazonian forests. Proc. Natl. Acad. Sci. USA 2017, 114, 1868–1873. [Google Scholar] [CrossRef] [Green Version]
- Smith, B.D. Low-level food production. J. Arch. Res. 2001, 9, 1–43. [Google Scholar] [CrossRef]
- Vavilov, N.I. Origin and Geography of Cultivated Plants; Cambridge University Press: New York, NY, USA, 1992. [Google Scholar]
- Vavilov, N.I. Five Continents; Rodin, L.E., Ed.; International Plant Genetic Resources Institute: Rome, Italy, 1997; p. 198. [Google Scholar]
- Clement, C.R. A center of crop genetic diversity in western Amazonia. Bioscience 1989, 39, 624–631. [Google Scholar] [CrossRef]
- Clement, C.R. 1492 and the loss of Amazonian crop genetic resources. II. Crop biogeography at contact. Econ. Bot. 1999, 53, 203–216. [Google Scholar] [CrossRef]
- Clement, C.R.; Cristo-Araújo, M.; Coppens d’Eeckenbrugge, G.; Alves Pereira, A.; Picanço-Rodrigues, D. Origin and domestication of native Amazonian crops. Diversity 2010, 2, 72–106. [Google Scholar] [CrossRef] [Green Version]
- Harlan, J.R. Agricultural origins: Centers and noncenters. Science 1971, 174, 468–474. [Google Scholar] [CrossRef] [PubMed]
- Lins Neto, E.M.d.F.; Peroni, N.; Casas, A.; Parra, F.; Aguirre, X.; Guillén, S.; Albuquerque, U.P. Brazilian and Mexican experiences in the study of incipient domestication. J. Ethnobiol. Ethnomed. 2014, 10, 33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Casas, A.; Torres-Guevara, J.; Parra-Rondinel, F. (Eds.) Domesticación en el Continente Americano. 1. Manejo de la Biodiversidad y Evolución Dirigida por las Culturas del Nuevo Mundo; Universidad Nacional Autónoma de México; Universidad Nacional Agraria La Molina (UNALM) del Perú: Morelia, Mexico, 2016; Volume 1, p. 502. [Google Scholar]
- Casas, A.; Torres-Guevara, J.; Parra-Rondinel, F. (Eds.) Domesticación en el Continente Americano. 2. Investigación para el Manejo Sustentable de Recursos Genéticos en el Nuevo Mundo; Universidad Nacional Autónoma de México; Universidad Nacional La Molina (UNALM) del Perú: Morelia, Mexico, 2017; Volume 2, p. 575. [Google Scholar]
- Zeder, M.A. Central questions in the domestication of plants and animals. Evol. Anthropol. 2006, 15, 105–117. [Google Scholar] [CrossRef]
- Viveiros de Castro, E. Who is afraid of the ontological wolf? Some comments on an ongoing anthropological debate. Camb. J. Anthropol. 2015, 33, 2–17. [Google Scholar] [CrossRef]
- Hutchinson, G.E. Concluding remarks. In Proceedings of Cold Spring Harbor Symposia on Quantitative Biology; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 1957; pp. 415–427. [Google Scholar]
- Lewontin, R.C. The Triple Helix: Gene, Organism, and Environment; Harvard University Press: Cambridge, UK, 2000; p. 136. [Google Scholar]
- Smith, B.D. A cultural niche construction theory of initial domestication. Biol. Theory 2012, 6, 260–271. [Google Scholar] [CrossRef]
- Leach, H.M. Human domestication reconsidered. Curr. Anthropol. 2003, 44, 349–368. [Google Scholar] [CrossRef]
- Hastorf, C. The cultural life of early domestic plant use. Antiquity 1998, 72, 773–782. [Google Scholar] [CrossRef]
- Casas, A.; Vázquez, M.d.C.; Viveros, J.L.; Caballero, J. Plant management among the Nahua and the Mixtec in the Balsas River Basin, Mexico: An ethnobotanical approach to the study of plant domestication. Hum. Ecol. 1996, 24, 455–478. [Google Scholar] [CrossRef]
- Blancas, J.; Casas, A.; Rangel-Landa, S.; Moreno-Calles, A.; Torres, I.; Pérez-Negrón, E.; Solís, L.; Delgado-Lemus, A.; Parra, F.; Arellanes, Y.; et al. Plant management in the Tehuacan-Cuicatlan Valley, Mexico. Econ. Bot. 2010, 64, 287–302. [Google Scholar] [CrossRef]
- Blancas, J.; Casas, A.; Pérez-Salicrup, D.; Caballero, J.; Vega, E. Ecological and socio-cultural factors influencing plant management in Náhuatl communities of the Tehuacán Valley, Mexico. J. Ethnobiol. Ethnomed. 2013, 9, 39. [Google Scholar] [CrossRef] [Green Version]
- Parra, F.; Blancas, J.; Casas, A. Landscape management and domestication of Stenocereus pruinosus (Cactaceae) in the Tehuacán Valley: Human guided selection and gene flow. J. Ethnobiol. Ethnomed. 2012, 8, 32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spengler, R.N., 3rd. Anthropogenic seed dispersal: Rethinking the origins of plant domestication. Trends Plant Sci. 2020, 25, 340–348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Posey, D.A. Indigenous management of tropical forest ecosystems: The case of the Kayapo Indians of the Brazilian Amazon. Agrofor. Syst. 1985, 3, 139–158. [Google Scholar] [CrossRef]
- Blancas, J.; Casas, A.; Lira, R.; Caballero, J. Traditional management and morphological patterns of Myrtillocactus schenckii (Cactaceae) in the Tehuacán Valley, Central Mexico. Econ. Bot. 2009, 63, 375. [Google Scholar] [CrossRef]
- Levis, C.; Flores, B.M.; Moreira, P.A.; Luize, B.G.; Alves, R.P.; Franco-Moraes, J.; Lins, J.; Konings, E.; Peña-Claros, M.; Bongers, F.; et al. How people domesticated Amazonian forests. Front. Ecol. Evol. 2018, 5, 171. [Google Scholar] [CrossRef] [Green Version]
- Anderson, E. Plants, Man and Life; Courier Dover Publications: Mineola, NY, USA, 2005; p. 251. [Google Scholar]
- Lathrap, D.W. Our father the cayman, our mother the gourd: Spinden revisited, or a unitary model for emergence of agriculture in the New World. In Origins of Agriculture; Reed, C.A., Ed.; Mouton: The Hague, The Netherlands, 1977; pp. 713–754. [Google Scholar]
- Clement, C.R.; Cassino, M.F. Landscape domestication and archaeology. In Encyclopedia of Global Archaeology; Smith, C., Ed.; Springer: New York, NY, USA, 2018; pp. 1–8. [Google Scholar] [CrossRef]
- Clement, C.R. 1492 and the loss of Amazonian crop genetic resources. I. The relation between domestication and human population decline. Econ. Bot. 1999, 53, 188–202. [Google Scholar] [CrossRef]
- Bogoni, J.A.; Graipel, M.E.; Peroni, N. The ecological footprint of Acca sellowiana domestication maintains the residual vertebrate diversity in threatened highlands of Atlantic Forest. PLoS ONE 2018, 13, e0195199. [Google Scholar] [CrossRef] [Green Version]
- Denevan, W.M. Stone vs. metal axes: The ambiguity of shifting cultivation in prehistoric Amazonia. J. Steward Anthropol. Soc. 1992, 20, 153–165. [Google Scholar]
- Denevan, W.M.; Treacy, J.M.; Alcorn, J.B.; Padoch, C.; Denslow, J.; Paitan, S.F. Indigenous agroforestry in the Peruvian Amazon: Bora Indian management of swidden fallows. Interciencia 1984, 9, 346–357. [Google Scholar]
- Bogoni, J.A.; Muniz-Tagliari, M.; Peroni, N.; Peres, C.A. Testing the keystone plant resource role of a flagship subtropical tree species (Araucaria angustifolia) in the Brazilian Atlantic Forest. Ecol. Indic. 2020, 118, 106778. [Google Scholar] [CrossRef]
- Leakey, R.R.; Tchoundjeu, Z.; Smith, R.I.; Munro, R.C.; Fondoun, J.-M.; Kengue, J.; Anegbeh, P.O.; Atangana, A.R.; Waruhiu, A.N.; Asaah, E. Evidence that subsistence farmers have domesticated indigenous fruits (Dacryodes edulis and Irvingia gabonensis) in Cameroon and Nigeria. Agrofor. Syst. 2004, 60, 101–111. [Google Scholar] [CrossRef]
- Fuller, D.Q. An emerging paradigm shift in the origins of agriculture. Gen. Anthropol. 2010, 17, 8–12. [Google Scholar] [CrossRef]
- Falconer, D.S.; MacKay, T.F. Introduction to Quantitative Genetics; Longman: Harlow, UK, 1996; p. 480. [Google Scholar]
- Cleveland, D.A.; Daniela, S.; Smith, S.E. A biological framework for understanding farmers’ plant breeding. Econ. Bot. 2000, 54, 377–394. [Google Scholar] [CrossRef]
- Leach, H.M. The terminology of agricultural origins and food production systems—A horticultural perspective. Antiquity 1997, 71, 135–148. [Google Scholar] [CrossRef]
- Bellwood, P. First Farmers: The Origins of Agricultural Societies; Blackwell Publishing: Oxford, UK, 2005; p. 360. [Google Scholar]
- van der Veen, M. Gardens and fields: The intensity and scale of food production. World Arch. 2005, 37, 157–163. [Google Scholar] [CrossRef]
- Rosenswig, R.M.; VanDerwarker, A.M.; Culleton, B.J.; Kennett, D.J. Is it agriculture yet? Intensified maize-use at 1000 cal BC in the Soconusco and Mesoamerica. J. Anthropol. Archaeol. 2015, 40, 89–108. [Google Scholar] [CrossRef]
- Ames, O. Economic Annuals and Human Cultures; Botanical Museum of Harvard University: Cambridge, UK, 1939; p. 153. [Google Scholar]
- Callicott, C.M. Interspecies communication in the Western Amazon: Music as a form of conversation between plants and people. Eur. J. Ecopsychol. 2013, 4, 32–43. [Google Scholar]
- Lima, A.G.M.d. A cultura da batata-doce: Cultivo, parentesco e ritual entre os Krahô. Mana 2017, 23, 455–490. [Google Scholar] [CrossRef] [Green Version]
- Rival, L. Seed and clone: The symbolic and social significance of bitter manioc. In Beyond the Visible and the Material: The Amerindianization of Society in the Work of Peter Rivière; Rival, L.M., Whitehead, N.L., Eds.; Oxford University Press: Oxford, UK, 2001; pp. 57–80. [Google Scholar]
- Casas, A.; Viveros, J.L.; Caballero, J. Etnobotánica Mixteca: Sociedad, Cultura y Recursos Naturales en la Montaña de Guerrero; Instituto Nacional Indigenista/Consejo Nacional para la Cultura y las Artes: Ciudad de México, Mexico, 1994. [Google Scholar]
- Barton, H.; Denham, T. Vegecultures and the social–biological transformations of plants and people. Quat. Int. 2018, 489, 17–25. [Google Scholar] [CrossRef]
- Denham, T.; Barton, H.; Castillo, C.; Crowther, A.; Dotte-Sarout, E.; Florin, S.A.; Pritchard, J.; Barron, A.; Zhang, Y.; Fuller, D.Q. The domestication syndrome in vegetatively propagated field crops. Ann. Bot. 2020, 125, 581–597. [Google Scholar] [CrossRef]
- Iriarte, J.; Elliott, S.; Maezumi, S.Y.; Alves, D.; Gonda, R.; Robinson, M.; Gregorio de Souza, J.; Watling, J.; Handley, J. The origins of Amazonian landscapes: Plant cultivation, domestication and the spread of food production in tropical South America. Quat. Sci. Rev. 2020, 248. [Google Scholar] [CrossRef]
- Oviedo y Valdés, G.F. Historia General y Natural de las Indias. [Juan Cromberger, Sevilla (1851–1855). J. A. de los Rios y Serrano (ed.)]; Imprenta de la Real Academia de la Historia: Madrid, Spain, 1535. [Google Scholar]
- Patiño, V.M. Historia y Dispersión de los Frutales Nativos del Neotrópico; Centro Internacional de Agricultura Tropical: Cali, Colombia, 2002; p. 655. [Google Scholar]
- Walker, J.H. Island, River, and Field: Landscape Archaeology in the Llanos de Mojos; University of New Mexico Press: Albuquerque, Mexico, 2018. [Google Scholar]
- Ford, A.; Nigh, R. Origins of the Maya forest garden: Maya resource management. J. Ethnobiol. 2009, 29, 213–236. [Google Scholar] [CrossRef]
- Ford, A. The Maya forest: A domesticated landscape. In The Maya World; Hutson, S.R., Ardren, T., Eds.; Routledge: New York, NY, USA, 2020; pp. 519–539. [Google Scholar]
- Smith, M.E. The Aztecs; Wiley-Blackwell: Oxford, UK, 2013; p. 416. [Google Scholar]
- Hassig, R. Aztec logistics and the unanticipated consequences of empire. In The Archaeology of Food and Warfare-Food Insecurity in Prehistory; Van Derwarker, A.M., Wilson, G.D., Eds.; Springer: Cham, Switzerland, 2016; pp. 149–160. [Google Scholar] [CrossRef]
- Heckenberger, M.J. Rethinking the Arawakan diaspora: Hierarchy, regionality, and the Amazonian formative. In Comparative Arawakan Histories: Rethinking Language Family and Culture Area in Amazonia; Hill, J.D., Santos-Granero, F., Eds.; University of Illinois Press: St. Louis, MO, USA, 2002; pp. 99–122. [Google Scholar]
- Levis, C.; Costa, F.R.C.; Bongers, F.; Peña-Claros, M.; Clement, C.R.; Junqueira, A.B.; Neves, E.G.; Tamanaha, E.K.; Figueiredo, F.O.G.; Salomão, R.P.; et al. Persistent effects of pre-Columbian plant domestication on Amazonian forest composition. Science 2017, 355, 925–931. [Google Scholar] [CrossRef]
- Shepard, G.H., Jr.; Clement, C.R.; Lima, H.P.; Mendes dos Santos, G.; Moraes, C.d.P.; Neves, E.G. Ancient and traditional agriculture in South America: Tropical lowlands. In Encyclopedia of Agriculture and the Environment; Hazlett, R., Ed.; Oxford University Press: New York, NY, USA, 2020; p. 48. [Google Scholar] [CrossRef]
- Nair, P.R. Classification of agroforestry systems. Agrofor. Syst. 1985, 3, 97–128. [Google Scholar] [CrossRef]
- Waters, M.R. Late Pleistocene exploration and settlement of the Americas by modern humans. Science 2019, 365, eaat5447. [Google Scholar] [CrossRef]
- Ardelean, C.F.; Becerra-Valdivia, L.; Pedersen, M.W.; Schwenninger, J.-L.; Oviatt, C.G.; Macias-Quintero, J.I.; Arroyo-Cabrales, J.; Sikora, M.; Ocampo-Diaz, Y.Z.E.; Rubio-Cisneros, I.I.; et al. Evidence of human occupation in Mexico around the Last Glacial Maximum. Nature 2020, 584, 87–92. [Google Scholar] [CrossRef]
- Henderson, A.; Galeano, G.; Bernal, R. A Field Guide to the Palms of the Americas; Princeton University Press: Princeton, NJ, USA, 1995. [Google Scholar]
- Rangel-Landa, S.; Rivera-Lozoya, E.; Casas, A. Uso y manejo de las palmas Brahea spp. (Arecaceae) por el pueblo ixcateco de Santa María Ixcatlán Oaxaca, México. Gaia Sci. 2014, 8, 62–78. [Google Scholar]
- Becerra-Valdivia, L.; Higham, T. The timing and effect of the earliest human arrivals in North America. Nature 2020, 584, 93–97. [Google Scholar] [CrossRef]
- Dillehay, T.D.; Ocampo, C.; Saavedra, J.; Sawakuchi, A.O.; Vega, R.M.; Pino, M.; Collins, M.B.; Scott Cummings, L.; Arregui, I.; Villagran, X.S.; et al. New archaeological evidence for an early human presence at Monte Verde, Chile. PLoS ONE 2015, 10, e0141923. [Google Scholar] [CrossRef] [Green Version]
- Vialou, D.; Benabdelhadi, M.; Feathers, J.; Fontugne, M.; Vialou, A.V. Peopling South America’s centre: The late Pleistocene site of Santa Elina. Antiquity 2017, 91, 865–884. [Google Scholar] [CrossRef] [Green Version]
- Scheel-Ybert, R.; Bachelet, C. A good place to live: Plants and people at the Santa Elina rock shelter (Central Brazil) from Late Pleistocene to the Holocene. Latin Am. Antiq. 2020, 31, 273–291. [Google Scholar] [CrossRef]
- Barbieri, C.; Barquera, R.; Arias, L.; Sandoval, J.R.; Acosta, O.; Zurita, C.; Aguilar-Campos, A.; Tito-Alvarez, A.M.; Serrano-Osuna, R.; Gray, R.D.; et al. The current genomic landscape of western South America: Andes, Amazonia, and Pacific Coast. Mol. Biol. Evol. 2019, 36, 2698–2713. [Google Scholar] [CrossRef] [Green Version]
- Campbell, L.; Grondona, V. The Indigenous Languages of South America: A Comprehensive Guide; de Gruyter Mouton: Berlin, Germany, 2012; p. 750. [Google Scholar]
- Pearsall, D.M. The origins of plant cultivation in South America. In The Origins of Agriculture: An International Perspective; Cowan, C.W., Watson, P.J., Eds.; Smithsonian Institution Press: Washington, DC, USA, 1992; pp. 173–206. [Google Scholar]
- CONABIO. Provincias Biogeográficas de México; Comisión Nacional para el Conocimiento y Uso de la Biodiversidad (CONABIO): Ciudad de México, Mexico, 1997. [Google Scholar]
- Olson, D.M.; Dinerstein, E.; Wikramanayake, E.D.; Burgess, N.D.; Powell, G.V.N.; Underwood, E.C.; D’amico, J.A.; Itoua, I.; Strand, H.E.; Morrison, J.C.; et al. Terrestrial ecoregions of the world: A new map of life on Earth. Bioscience 2001, 51, 933–938. [Google Scholar] [CrossRef]
- Romano, G.M. A high resolution shapefile of the Andean biogeographical region. Data Brief 2017, 13, 230–232. [Google Scholar] [CrossRef]
- Antonelli, A.; Zizka, A.; Carvalho, F.A.; Scharn, R.; Bacon, C.D.; Silvestro, D.; Condamine, F.L. Amazonia is the primary source of Neotropical biodiversity. Proc. Natl. Acad. Sci. USA 2018, 115, 6034–6039. [Google Scholar] [CrossRef] [Green Version]
- McClung de Tapia, E. The origins of agriculture in Mesoamerica and Central America. In The Origins of Agriculture: An International Perspective; Cowan, C.W., Watson, P.J., Eds.; Smithsonian Institution Press: Washington, DC, USA, 1992; pp. 143–171. [Google Scholar]
- Smith, B.D. The initial domestication of Cucurbita pepo in the Americas 10,000 years ago. Science 1997, 276, 932–934. [Google Scholar] [CrossRef] [Green Version]
- Lira, R.; Eguiarte, L.; Montes, S.; Zizumbo-Villarreal, D.; Marín, P.C.-G.; Quesada, M. Homo sapiens–Cucurbita interaction in Mesoamerica: Domestication, dissemination, and diversification. In Ethnobotany of Mexico—Interactions of People and Plants in Mesoamerica; Lira, R., Casas, A., Blancas, J., Eds.; Springer: New York, NY, USA, 2016; pp. 389–401. [Google Scholar] [CrossRef]
- Castellanos-Morales, G.; Paredes-Torres, L.M.; Gamez, N.; Hernandez-Rosales, H.S.; Sanchez-de la Vega, G.; Barrera-Redondo, J.; Aguirre-Planter, E.; Vazquez-Lobo, A.; Montes-Hernandez, S.; Lira-Saade, R.; et al. Historical biogeography and phylogeny of Cucurbita: Insights from ancestral area reconstruction and niche evolution. Mol. Phylogenet. Evol. 2018, 128, 38–54. [Google Scholar] [CrossRef]
- Piperno, D.R.; Ranere, A.J.; Holst, I.; Iriarte, J.; Dickau, R. Starch grain and phytolith evidence for early ninth millennium BP maize from the Central Balsas River Valley, Mexico. Proc. Natl. Acad. Sci. USA 2009, 106, 5019–5024. [Google Scholar] [CrossRef] [Green Version]
- Pickersgill, B. Domestication of plants in Mesoamerica: An archaeological review with some ethnobotanical interpretations. In Ethnobotany of Mexico—Interactions of People and Plants in Mesoamerica; Lira, R., Casas, A., Blancas, J., Eds.; Springer: New York, NY, USA, 2016; pp. 207–231. [Google Scholar] [CrossRef]
- Debouck, D.G. Your beans of the last harvest and the possible adoption of bright ideas. In Ethnobotany of Mexico: Interactions of People and Plants in Mesoamerica; Lira, R., Casas, A., Blancas, J., Eds.; Springer: New York, NY, USA, 2016; pp. 367–387. [Google Scholar]
- Rodriguez, M.; Rau, D.; Bitocchi, E.; Bellucci, E.; Biagetti, E.; Carboni, A.; Gepts, P.; Nanni, L.; Papa, R.; Attene, G. Landscape genetics, adaptive diversity and population structure in Phaseolus vulgaris. New Phytol. 2016, 209, 1781–1794. [Google Scholar] [CrossRef]
- Smith, C.E. Archeological evidence for selection in avocado. Econ. Bot. 1966, 20, 169–175. [Google Scholar] [CrossRef]
- Van der Hammen, T.; Urrego, G.C. Mastodontes en el humedal pleistocénico en el valle del Magdalena (Colombia) con evidencias de la presencia del hombre en el pleniglacial. Bol. Arqueol. 2001, 16, 4–36. [Google Scholar]
- Aceituno, F.J.; Loaiza, N. The origins and early development of plant food production and farming in Colombian tropical forests. J. Anthropol. Archaeol. 2018, 49, 161–172. [Google Scholar] [CrossRef]
- Gnecco, C. Against ecological reductionism: Late Pleistocene hunter-gatherers in the tropical forests of northern South America. Quat. Int. 2003, 109, 13–21. [Google Scholar] [CrossRef]
- Piperno, D.R.; Stothert, K.E. Phytolith evidence for early Holocene Cucurbita domestication in southwest Ecuador. Science 2003, 299, 1054–1057. [Google Scholar] [CrossRef] [Green Version]
- Capriles, J.M.; Albarracin-Jordan, J. The earliest human occupations in Bolivia: A review of the archaeological evidence. Quat. Int. 2013, 301, 46–59. [Google Scholar] [CrossRef]
- Pearsall, D.M. Plant domestication and the shift to agriculture in the Andes. In The Handbook of South American Archaeology; Silverman, H., Isbell, W.H., Eds.; Springer: New York, NY, USA, 2008; pp. 105–120. [Google Scholar] [CrossRef]
- Dillehay, T.D.; Goodbred, S.; Pino, M.; Vasquez Sanchez, V.F.; Tham, T.R.; Adovasio, J.; Collins, M.B.; Netherly, P.J.; Hastorf, C.A.; Chiou, K.L.; et al. Simple technologies and diverse food strategies of the Late Pleistocene and Early Holocene at Huaca Prieta, Coastal Peru. Sci. Adv. 2017, 3, e1602778. [Google Scholar] [CrossRef] [Green Version]
- Chiou, K.L.; Hastorf, C.A.; Bonavia, D.; Dillehay, T.D. Documenting cultural selection pressure changes on chile pepper (Capsicum baccatum L.) seed size through time in Coastal Peru (7600 BP–Present). Econ. Bot. 2014, 68, 190–202. [Google Scholar] [CrossRef]
- Dillehay, T.D.; Ramirez, C.; Pino, M.; Collins, M.B.; Rossen, J.; Pino-Navarro, J.D. Monte Verde: Seaweed, food, medicine, and the peopling of South America. Science 2008, 320, 784–786. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lahaye, C.; Hernandez, M.; Boëda, E.; Felice, G.D.; Guidon, N.; Hoeltz, S.; Lourdeau, A.; Pagli, M.; Pessis, A.-M.; Rasse, M.; et al. Human occupation in South America by 20,000 BC: The Toca da Tira Peia site, Piauí, Brazil. J. Arch. Sci. 2013, 40, 2840–2847. [Google Scholar] [CrossRef]
- Roosevelt, A.C. The Amazon and the Anthropocene: 13,000 years of human influence in a tropical rainforest. Anthropocene 2013, 4, 69–87. [Google Scholar] [CrossRef]
- Lourdeau, A.; Carbonera, M.; Santos, M.C.P.; Hoeltz, S.; Fontugne, M.; Hatté, C.; Silva, S.F.S.M.d.; Rosina, P.; Lucas, L.d.O.; da Costa, A. Pré-história na foz do rio Chapecó. Cad. CEOM Chapecó 2016, 29, 220–242. [Google Scholar] [CrossRef] [Green Version]
- Morcote-Ríos, G.; Aceituno, F.; León, T. Recolectores del Holoceno temprano en la floresta amazónica colombiana. In Antes de Orellana. Actas del 3er Encuentro Internacional de Arqueología Amazónica; Rostain, S., Ed.; Instituto Francés de Estudios Andinos: Lima, Peru, 2013; pp. 39–50. [Google Scholar]
- Morcote-Ríos, G.; Mahecha Rubio, D.; Franky Calvo, C.E. Recorrido en el tiempo: 12000 años de ocupación de la Amazonia. In Universidad y Territorio; Restrepo Zea, E., Sánchez, C.H., Silva Carrero, G., Eds.; Universidad Nacional de Colombia: Bogotá, Colombia, 2017; Volume 1, pp. 66–93. [Google Scholar]
- Aceituno, F.J.; Loaiza, N. The role of plants in the early human settlement of Northwest South America. Quat. Int. 2015, 363, 20–27. [Google Scholar] [CrossRef]
- Morcote-Ríos, G.; Aceituno, F.J.; Iriarte, J.; Robinson, M.; Chaparro-Cárdenas, J.L. Colonisation and early peopling of the Colombian Amazon during the Late Pleistocene and the Early Holocene: New evidence from La Serranía La Lindosa. Quat. Int. 2020. [Google Scholar] [CrossRef]
- Morcote-Ríos, G.; Becerra, G.C.; Rubio, D.M.; Calvo, C.E.F.; Cavalier, I. Las palmas entre los grupos cazadores-recolectores de la Amazonía colombiana. Caldasia 1998, 20, 57–74. [Google Scholar]
- Mora Camargo, S. Early Inhabitants of the Amazonian Tropical Rain Forest. A Study of Humans and Environmental Dynamics; University of Pittsburgh Press, Universidad Nacional de Colombia-Sede Leticia, Instituto Amazónico de Investigaciones IMANI: Pittsburgh, PA, USA, 2003; p. 211. [Google Scholar]
- Roosevelt, A.C. Ancient and modern hunter-gatherers of lowland South America: An evolutionary problem. In Advances in Historical Ecology; Balée, W., Ed.; Columbia University Press: New York, NY, USA, 1998; pp. 190–212. [Google Scholar]
- Roosevelt, A.C.; Lima da Costa, M.; Lopes Machado, C.; Michab, M.; Mercier, N.; Valladas, H.; Feathers, J.; Barnett, W.; Imazio da Silveira, M.; Henderson, A.; et al. Paleoindian cave dwellers in the Amazon: The peopling of the Americas. Science 1996, 272, 373–384. [Google Scholar] [CrossRef]
- Shock, M.P.; Moraes, C.d.P. A floresta é o domus: A importância das evidências arqueobotânicas e arqueológicas das ocupações humanas amazônicas na transição Pleistoceno/Holoceno. Bol. Mus. Para. Emílio Goeldi Ciênc. Hum. 2019, 14, 263–289. [Google Scholar] [CrossRef] [Green Version]
- Magalhães, M.P. A Phýsis da Origem: O Sentido da História da Amazônia; Museu Paraense Emílio Goeldi: Belém, Brazil, 2005; p. 351. [Google Scholar]
- Santos, R.S.; Lima, P.G.C.; Magalhães, M.P.; Scheel-Ybert, R.; Coelho-Ferreira, M.; Feliciano, A.L.P.; Albernaz, A.L.K.M. Plantas úteis na flora contemporânea e pretérita de Carajás. In A Humanidade e a Amazônia: 11 mil anos de Evolução Histórica em Carajás; Magalhães, M.P., Ed.; Museu Paraense Emílio Goeldi: Belém, Brazil, 2018; pp. 183–210. [Google Scholar]
- Lombardo, U.; Iriarte, J.; Hilbert, L.; Ruiz-Pérez, J.; Capriles, J.M.; Veit, H. Early Holocene crop cultivation and landscape modification in Amazonia. Nature 2020, 581, 190–193. [Google Scholar] [CrossRef]
- Watling, J.; Shock, M.P.; Mongelo, G.Z.; Almeida, F.O.; Kater, T.; Oliveira, P.E.; Neves, E.G. Direct archaeological evidence for Southwestern Amazonia as an early plant domestication and food production centre. PLoS ONE 2018, 13, e0199868. [Google Scholar] [CrossRef]
- Chaves, S.A.d.M. Novas perspectivas paleoetnobotânicas e paleovegetacionais do Planalto Central do Brasil: Uma proposta palinológica. In Pré-História do Mato Grosso; Vialou, Á.V., Ed.; EDUSP: São Paulo, Brazil, 2005; Volume 1, pp. 149–154. [Google Scholar]
- Chaves, S.A.M.; Reinhard, K.J. Critical analysis of coprolite evidence of medicinal plant use, Piauí, Brazil. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2006, 237, 110–118. [Google Scholar] [CrossRef] [Green Version]
- Ortega, D.D. Microvestígios Botânicos em Artefatos Líticos do Sítio Lapa do Santo (Lagoa Santa, Minas Gerais). Ph.D. Thesis, Universidade de São Paulo, São Paulo, Brazil, 2019. [Google Scholar]
- Angeles Flores, R. Uso de Recursos Vegetais em Lapa Grande de Taquaraçu: Evidencias Macro e Microscópicas. Master’s Thesis, Universidade de São Paulo, São Paulo, Brazil, 2015. [Google Scholar]
- Angeles Flores, R.; de Sousa, J.C.M.; de Mello Araujo, A.G.; Ceccantini, G. Before Lagoa Santa: Micro-remain and technological analysis in a lithic artifact from the Itaparica industry. J. Lithic Stud. 2016, 3, 6–29. [Google Scholar] [CrossRef] [Green Version]
- Resende, E.T.; Prous, A. Os vestígios vegetais do Grande Abrigo de Santana do Riacho. Arq. Mus. História Nat. UFMG 1991, 12, 87–111. [Google Scholar]
- Scheel-Ybert, R. Preliminary data on nonwood plant remains at Sambaquis from the Southern and Southeastern Brazilian coast: Considerations on diet, ritual, and site particularities. Cuad. Inst. Nac. Antropol. Pensam. Latinoam. Ser. Espec. 2013, 1, 60–72. [Google Scholar]
- Scheel-Ybert, R.; Boyadjian, C. Gardens on the coast: Considerations on food production by Brazilian shellmound builders. J. Anthropol. Archaeol. 2020, 60, 101211. [Google Scholar] [CrossRef]
- Iriarte, J.; Behling, H. The expansion of Araucaria forest in the southern Brazilian highlands during the last 4000 years and its implications for the development of the Taquara/Itararé Tradition. Environ. Arch. 2007, 12, 115–127. [Google Scholar] [CrossRef] [Green Version]
- Caballero, J.; Martínez-Ballesté, A.; Cortés, L. Base de Datos Etnobotánicos de México (BADEPLAM); Jardín Botánico del Instituto de Biología, Universidad Nacional Autónoma de México (UNAM): Ciudad de México, Mexico, 2020. [Google Scholar]
- Diazgranados, M.; Allkin, B.; Black, N.; Cámara-Leret, R.; Canteiro, C.; Carretero, J.; Ulian, T. World Checklist of Useful Plant Species; Royal Botanic Gardens, Kew & Knowledge Network for Biocomplexity: London, UK, 2020; p. 689. [Google Scholar]
- Hanelt, P.; IPK. Mansfeld’s Encyclopedia of Agricultural and Horticultural Crops; Institute of Plant Genetics and Crop Plant Research (IPK): Gatersleben, Germany, 2001. [Google Scholar]
- Ulloa, C.U.; Acevedo-Rodríguez, P.; Beck, S.; Belgrano, M.J.; Bernal, R.; Berry, P.E.; Brako, L.; Celis, M.; Davidse, G.; Forzza, R.C. An integrated assessment of the vascular plant species of the Americas. Science 2017, 358, 1614–1617. [Google Scholar] [CrossRef] [Green Version]
- Villaseñor, J.L. Checklist of the native vascular plants of Mexico. Rev. Mex. Biodivers. 2016, 87, 559–902. [Google Scholar] [CrossRef] [Green Version]
- Casas, A.; Parra-Rondinel, F.; Aguirre-Dugua, X.; Rangel-Landa, S.; Blancas, J.; Vallejo, M.; Moreno-Calles, A.I.; Guillén, S.; Torres-García, I.; Delgado-Lemus, A.; et al. Manejo y domesticación de plantas en Mesoamérica: Una estrategia de investigación y estado del conocimiento sobre los recursos genéticos. In Domesticación en el Continente Americano. 2. Investigación para el Manejo Sustentable de Recursos Genéticos en el Nuevo Mundo; Casas, A., Torres-Guevara, J., Parra, F., Eds.; Universidad Nacional Autónoma de México; Universidad Nacional Agraria La Molina (UNALM) del Perú: Morelia, Mexico, 2017; Volume 2, pp. 69–102. [Google Scholar]
- Casas, A.; Valiente-Banuet, A.; Viveros, J.L.; Caballero, J.; Cortés, L.; Dávila, P.; Lira, R.; Rodríguez, I. Plant resources of the Tehuacán-Cuicatlán Valley, Mexico. Econ. Bot. 2001, 55, 129–166. [Google Scholar] [CrossRef]
- Rangel-Landa, S.; Casas, A.; Rivera-Lozoya, E.; Torres-García, I.; Vallejo-Ramos, M. Ixcatec ethnoecology: Plant management and biocultural heritage in Oaxaca, Mexico. J. Ethnobiol. Ethnomed. 2016, 12, 30. [Google Scholar] [CrossRef] [Green Version]
- Rangel-Landa, S.; Casas, A.; García-Frapolli, E.; Lira, R. Socio-cultural and ecological factors influencing management of edible and non-edible plants: The case of Ixcatlán, Mexico. J. Ethnobiol. Ethnomed. 2017, 13, 59. [Google Scholar] [CrossRef] [Green Version]
- Torres, I.; Blancas, J.; León, A.; Casas, A. TEK, local perceptions of risk, and diversity of management practices of Agave inaequidens in Michoacán, Mexico. J. Ethnobiol. Ethnomed. 2015, 11, 61. [Google Scholar] [CrossRef] [Green Version]
- Casas, A.; Pickersgill, B.; Caballero, J.; Valiente-Banuet, A. Ethnobotany and domestication in Xoconochtli, Stenocereus stellatus (Cactaceae), in the Tehuacán Valley and La Mixteca Baja, México. Econ. Bot. 1997, 51, 279–292. [Google Scholar] [CrossRef]
- Arellano, E.; Casas, A. Morphological variation and domestication of Escontria chiotilla (Cactaceae) under silvicultural management in the Tehuacán Valley, Central Mexico. Genet. Resour. Crop Evol. 2003, 50, 439–453. [Google Scholar] [CrossRef]
- Carmona, A.; Casas, A. Management, phenotypic patterns and domestication of Polaskia chichipe (Cactaceae) in the Tehuacán Valley, Central Mexico. J. Arid Environ. 2005, 60, 115–132. [Google Scholar] [CrossRef]
- Parra, F.; Casas, A.; Peñaloza-Ramírez, J.M.; Cortés-Palomec, A.C.; Rocha-Ramírez, V.; González-Rodríguez, A. Evolution under domestication: Ongoing artificial selection and divergence of wild and managed Stenocereus pruinosus (Cactaceae) populations in the Tehuacan Valley, Mexico. Ann. Bot. 2010, 106, 483–496. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Figueredo, C.J.; Casas, A.; González-Rodríguez, A.; Nassar, J.M.; Colunga-GarcíaMarín, P.; Rocha-Ramírez, V. Genetic structure of coexisting wild and managed agave populations: Implications for the evolution of plants under domestication. AoB Plants 2015, 7, plv114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blancas, J.; Casas, A.; Moreno-Calles, A.I.; Caballero, J. Cultural motives of plant management and domestication. In Ethnobotany of Mexico; Lira, R., Casas, A., Blancas, J., Eds.; Springer: New York, NY, USA, 2016; pp. 233–255. [Google Scholar] [CrossRef]
- Aguirre-Dugua, X.; Pérez-Negrón, E.; Casas, A. Phenotypic differentiation between wild and domesticated varieties of Crescentia cujete L. and culturally relevant uses of their fruits as bowls in the Yucatan Peninsula, Mexico. J. Ethnobiol. Ethnomed. 2013, 9, 76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aguirre-Dugua, X.; Eguiarte, L.E.; González-Rodríguez, A.; Casas, A. Round and large: Morphological and genetic consequences of artificial selection on the gourd tree Crescentia cujete by the Maya of the Yucatan Peninsula, Mexico. Ann. Bot. 2012, 109, 1297–1306. [Google Scholar] [CrossRef]
- González-Soberanis, C.; Casas, A. Traditional management and domestication of tempesquistle, Sideroxylon palmeri (Sapotaceae) in the Tehuacán-Cuicatlán Valley, Central Mexico. J. Arid Environ. 2004, 59, 245–258. [Google Scholar] [CrossRef]
- Figueredo-Urbina, C.J.; Casas, A.; Torres-García, I. Morphological and genetic divergence between Agave inaequidens, A. cupreata and the domesticated A. hookeri. Analysis of their evolutionary relationships. PLoS ONE 2017, 12, e0187260. [Google Scholar] [CrossRef]
- Figueredo-Urbina, C.J.; Casas, A.; Martínez-Díaz, Y.; Santos-Zea, L.; Gutiérrez-Uribe, J.A. Domestication and saponins contents in a gradient of management intensity of agaves: Agave cupreata, A. inaequidens and A. hookeri in central Mexico. Genet. Resour. Crop Evol. 2018, 65, 1133–1146. [Google Scholar] [CrossRef]
- Abad-Fitz, I.; Maldonado-Almanza, B.; Aguilar-Dorantes, K.M.; Sánchez-Méndez, L.; Gómez-Caudillo, L.; Casas, A.; Blancas, J.; García-Rodríguez, Y.M.; Beltrán-Rodríguez, L.; Sierra-Huelsz, J.A.; et al. Consequences of traditional management in the production and quality of copal resin (Bursera bipinnata (Moc. & Sessé ex DC.) Engl.) in Mexico. Forests 2020, 11, 991. [Google Scholar] [CrossRef]
- Garavito, N.T.; Álvarez, E.; Caro, S.A.; Murakami, A.A.; Blundo, C.; Espinoza, T.E.B.; La Torre, M.A. Evaluación del estado de conservación de los bosques montanos en los Andes tropicales. Ecosistemas 2012, 21, 148–166. [Google Scholar] [CrossRef]
- Weberbauer, A. El Mundo Vegetal de los Andes Peruanos: Estudio Fitogeográfico; Ministerio de Agricultura: Lima, Peru, 1945; p. 776. [Google Scholar]
- Eva, H.D.; Huber, O. A Proposal for Defining the Geographical Boundaries of Amazonia; Synthesis of the Results from an Expert Consultation Workshop Organized by the European Commission in Collaboration with the Amazon Cooperation Treaty Organization-JRC Ispra; European Commision: Luxembourg, 2005. [Google Scholar]
- Brako, L.; Zarucchi, J.L. Catalogue of the Flowering Plants and Gymnosperms of Peru; Missouri Botanical Garden: Saint Louis, MO, USA, 1993; Volume 45, p. 1286. [Google Scholar]
- Brack Egg, A. Encyclopedic Dictionary of Useful Plants of Peru; Center for Andean Regional Studies “Bartolomé de las Casas”/United Nations Development Program: Cuzco, Peru, 1999; p. 500. [Google Scholar]
- Brack Egg, A. Perú: Diez Mil años de Domesticación; Bruño: Lima, Peru, 2003. [Google Scholar]
- Pancorbo-Olivera, M.; Parra Rondinel, F.A.; Torres Guevara, J.J.; Casas Fernández, A. Los otros alimentos: Plantas comestibles silvestres y arvenses en dos comunidades campesinas de los Andes centrales del Perú. Rev. Etnobiol. 2020, 18, 8–36. [Google Scholar]
- Young, K. Bosques húmedos. In Botánica Económica de los Andes Centrales; Moraes, M., Ollgaard, B., Kvist, L.P., Borchsenius, F., Balslev, H., Eds.; Universidad Mayor de San Andrés: La Paz, Bolivia, 2006; pp. 121–129. [Google Scholar]
- Kessler, M. Bosques de Polylepis. In Botánica Económica de los Andes Centrales; Moraes, M., Ollgaard, B., Kvist, L.P., Borchsenius, F., Balslev, H., Eds.; Universidad Mayor de San Andrés: La Paz, Bolivia, 2006; pp. 110–120. [Google Scholar]
- Weigend, M.; Dostert, N.; Rodríguez, E. Bosques relictos de los Andes peruanos: Perspectivas económicas. In Botánica Económica en los Andes Centrales; Moraes, M., Ollgaard, B., Kvist, L.P., Borchsenius, F., Balslev, H., Eds.; Universidad Mayor de San Andrés: La Paz, Bolivia, 2006; pp. 130–145. [Google Scholar]
- Torres-Guevara, J.; Cruz-Soriano, A.; Parra, F. La conservacion in situ en el Perú. Una experiencia dentro del Proyecto “Conservacion in situ de cultivos nativos y sus parientes silvestres en el Perú”. In Domesticación en el Continente Americano. 1. Manejo de Biodiversidad y Evolución Dirigida por las Culturas del Nuevo Mundo; Casas, A., Torres-Guevara, J., Parra, F., Eds.; UNALM & UNAM: Lima, Peru, 2016; pp. 361–386. [Google Scholar]
- De Haan, S.; Bonierbale, M.; Ghislain, M.; Nunez, J.; Trujillo, G. Indigenous biosystematics of Andean potatoes: Folk taxonomy, descriptors and nomenclature. Acta Hortic. 2007, 745, 89–134. [Google Scholar] [CrossRef]
- Bonnave, M.; Bleeckx, T.; Terrazas, F.; Bertin, P. Effect of the management of seed flows and mode of propagation on the genetic diversity in an Andean farming system: The case of oca (Oxalis tuberosa Mol.). Agric. Hum. Values 2016, 33, 673–688. [Google Scholar] [CrossRef]
- Velásquez-Milla, D.; Casas, A.; Torres-Guevara, J.; Cruz-Soriano, A. Ecological and socio-cultural factors influencing in situ conservation of crop diversity by traditional Andean households in Peru. J. Ethnobiol. Ethnomed. 2011, 7, 40. [Google Scholar] [CrossRef] [Green Version]
- Brush, S.B. Ethnoecology, biodiversity, and modernization in Andean potato agriculture. J. Ethnobiol. 1992, 12, 161–185. [Google Scholar]
- Begazo, D.; Torres, I.; Márquez-Castellanos, E.; Segovia, J.; Zarazúa, M.; Parra, F.; Cruz, A.; Torres-Guevara, J.; Casas, A. Sacha, aracca, k’ipa y cultura andina: Las bases de la diversidad de papas. Leisa Rev. Agroecol. 2019, 35, 12–15. [Google Scholar]
- Bethke, P.C.; Halterman, D.A.; Jansky, S. Are we getting better at using wild potato species in light of new tools? Crop Sci. 2017, 57, 1241–1258. [Google Scholar] [CrossRef] [Green Version]
- Bradshaw, J.E.; Bryan, G.J.; Ramsay, G. Genetic resources (including wild and cultivated Solanum species) and progress in their utilisation in potato breeding. Potato Res. 2006, 49, 49–65. [Google Scholar] [CrossRef]
- Jansky, S. Overcoming hybridization barriers in potato. Plant Breed. 2006, 125, 1–12. [Google Scholar] [CrossRef]
- Maune, J.F.; Camadro, E.L.; Erazzú, L.E. Cross-incompatibility and self-incompatibility: Unrelated phenomena in wild and cultivated potatoes? Botany 2018, 96, 33–45. [Google Scholar] [CrossRef] [Green Version]
- Scurrah, M.; Celis-Gamboa, C.; Chumbiauca, S.; Salas, A.; Visser, R.G.F. Hybridization between wild and cultivated potato species in the Peruvian Andes and biosafety implications for deployment of GM potatoes. Euphytica 2008, 164, 881–892. [Google Scholar] [CrossRef]
- Marquez-Castellanos, E.; Parra-Rondinel, F.; Lizárraga, L.; Casas, A.; Torres-Guevara, J. Conectividad entre poblaciones silvestres y cultivadas de papa. In De los Cultivos Nativos y el Cambio del Clima. Hallazgos (Huánuco y Apurímac); Torres-Guevara, J., Parra-Rondinel, F., Casas, A., Cruz-Soriano, A., Eds.; Universidad Nacional San Antonio Abad de Cusco, Universidad Nacional Agraria La Molina, Universidad Nacional Autónoma de México, Programa Nacional de Innovación Agraria, IDMA, CADEP: Lima, Peru, 2019; pp. 137–159. [Google Scholar]
- Coelho, S.D. Estudo da Relação Entre os Tamanhos Populacionais das Espécies Arbóreas na Amazônia e seus Usos pelos Humanos; Instituto Nacional de Pesquisas da Amazônia: Manaus, Brazil, 2018. [Google Scholar]
- ter Steege, H.; Pitman, N.C.A.; Sabatier, D.; Baraloto, C.; Salomao, R.P.; Guevara, J.E.; Phillips, O.L.; Castilho, C.V.; Magnusson, W.E.; Molino, J.-F.; et al. Hyperdominance in the Amazonian tree flora. Science 2013, 342, 1243092. [Google Scholar] [CrossRef] [Green Version]
- Ferreira, M.J.; Levis, C.; Iriarte, J.; Clement, C.R. Legacies of intensive management in forests around pre-columbian and modern settlements in the Madeira-Tapajós interfluve, Amazonia. Acta Bot. Bras. 2019, 33, 212–220. [Google Scholar] [CrossRef] [Green Version]
- Levis, C.; Peña-Claros, M.; Clement, C.R.; Costa, F.R.C.; Alves, R.P.; Ferreira, M.J.; Figueiredo, C.G.; Bongers, F. Pre-Columbian soil fertilization and current management maintain food resource availability in old-growth Amazonian forests. Plant Soil 2020, 450, 29–48. [Google Scholar] [CrossRef] [Green Version]
- Alves, R.P.; Levis, C.; Clement, C.R. Use and management of piquiá suggest in situ domestication along the lower Tapajos River, Brazilian Amazonia. Econ. Bot. 2016, 70, 198–202. [Google Scholar] [CrossRef]
- Francisconi, A.F.; Alves, R.P.; Clement, C.R.; Dequigiovanni, G.; Carvalho, I.A.S.; Veasey, E.A. Genetic structure and diversity identify incipient domestication of piquiá [Caryocar villosum (Aubl.) Pers.] along the lower Tapajós River, Brazilian Amazonia. Genet. Resour. Crop Evol. 2021. [Google Scholar] [CrossRef]
- Clement, C.R.; Cristo-Araújo, M.d.; Coppens d’Eeckenbrugge, G.; Reis, V.M.d.; Lehnebach, R.; Picanço-Rodrigues, D. Origin and dispersal of domesticated peach palm. Front. Ecol. Evol. 2017, 5, 148. [Google Scholar] [CrossRef] [Green Version]
- Pausas, J.G.; Keeley, J.E. A burning story: The role of fire in the history of life. Bioscience 2009, 59, 593–601. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, I.B.; Sampaio, M.B.; Figueiredo, I.B.; Ticktin, T. Fogo e artesanato de capim-dourado no Jalapão–usos tradicionais e consequências ecológicas. Biodivers. Bras. 2011, 67–85. [Google Scholar] [CrossRef]
- Mistry, J.; Berardi, A.; Andrade, V.; Krahô, T.; Krahô, P.; Leonardos, O. Indigenous fire management in the cerrado of Brazil: The case of the Krahô of Tocantíns. Hum. Ecol. 2005, 33, 365–386. [Google Scholar] [CrossRef]
- Eloy, L.; Schmidt, I.B.; Borges, S.L.; Ferreira, M.C.; Santos, T.A. Seasonal fire management by traditional cattle ranchers prevents the spread of wildfire in the Brazilian Cerrado. Ambio 2018, 48, 890–899. [Google Scholar] [CrossRef] [PubMed]
- Baldauf, C.; Santos, F.A.M. Ethnobotany, traditional knowledge, and diachronic changes in non-timber forest products management: A case study of Himatanthus drasticus (Apocynaceae) in the Brazilian savanna. Econ. Bot. 2013, 67, 110–120. [Google Scholar] [CrossRef]
- Smith, M.; Fausto, C. Sociality and diversity of pequi (Caryocar brasiliense, Caryocaraceae) among the Kuikuro of the upper Xingu River (Brazil). Bol. Mus. Para. Emílio Goeldi Ciênc. Hum. 2016, 11, 87–113. [Google Scholar] [CrossRef] [Green Version]
- Sousa Júnior, J.R.; Collevatti, R.G.; Lins Neto, E.M.F.; Peroni, N.; Albuquerque, U.P. Traditional management affects the phenotypic diversity of fruits with economic and cultural importance in the Brazilian Savanna. Agrofor. Syst. 2018, 92, 11–21. [Google Scholar] [CrossRef]
- Posey, D.A. Kayapó Ethnoecology and Culture; Routledge: London, UK, 2002; Volume 6. [Google Scholar]
- Shock, M.P. Holocene Hunter-Gatherer Plant Use and Foraging Choice: A Test from Minas Gerais, Brazil. Ph.D. Thesis, University of California, Santa Barbara, CA, USA, 2010. [Google Scholar]
- Sousa Júnior, J.R.; Albuquerque, U.P.; Peroni, N. Traditional knowledge and management of Caryocar coriaceum Wittm. (Pequi) in the Brazilian savanna, northeastern Brazil. Econ. Bot. 2013, 67, 225–233. [Google Scholar] [CrossRef]
- Lins-Neto, E.M.d.F.; Oliveira, I.F.; Britto, F.B.; Albuquerque, U.P. Traditional knowledge, genetic and morphological diversity in populations of Spondias tuberosa Arruda (Anacardiaceae). Genet. Resour. Crop Evol. 2013, 60, 1389–1406. [Google Scholar] [CrossRef]
- Lima-Nascimento, A.M.; Bento-Silva, J.S.; Lucena, C.M.; Lucena, R.F.P. Ethnobotany of native cacti in the northeast region of Brazil: Can traditional use influence availability? Acta Bot. Bras. 2019, 33, 350–359. [Google Scholar] [CrossRef] [Green Version]
- Pedrosa, K.M.; Lopes, S.F.; Carvalho, K.T.N.; Marques de Lucena, C.; Lima-Nascimento, A.M.; Lucena, R.F.P. Traditional management of Cactaceae by local populations in the semiarid region of Brazil. Ethnobiol. Conserv. 2020, 9, 1–28. [Google Scholar] [CrossRef]
- Paiva, J.R.; Barros, L.M.; Cavalcanti, J.J.V. Cashew (Anacardium occidentale L.) breeding: A global perspective. In Breeding Plantation Tree Crops: Tropical Species; Jain, S.M., Priyadarshan, P.M., Eds.; Springer: New York, NY, USA, 2009; pp. 287–324. [Google Scholar] [CrossRef] [Green Version]
- Albuquerque, U.P.; Araújo, E.d.L.; Castro, C.C.; Alves, R.R.N. People and natural resources in the Caatinga. In Caatinga: The Largest Tropical Dry Rorest Region in South America; Silva, J.M.C.d., Leal, I.R., Tabarelli, M., Eds.; Springer International Publishing: Cham, Switzerland, 2017; pp. 303–333. [Google Scholar] [CrossRef]
- Santos, J.C.; Leal, I.R.; Almeida-Cortez, J.S.; Fernandes, G.W.; Tabarelli, M. Caatinga: The scientific negligence experienced by a dry tropical forest. Trop. Conserv. Sci. 2011, 4, 276–286. [Google Scholar] [CrossRef]
- Pereira Cruz, A.; Giehl, E.L.H.; Levis, C.; Machado, J.S.; Bueno, L.; Peroni, N. Pre-colonial Amerindian legacies in forest composition of southern Brazil. PLoS ONE 2020, 15, e0235819. [Google Scholar] [CrossRef] [PubMed]
- Robinson, M.; de Souza, J.G.; Maezumi, S.Y.; Cardenas, M.; Pessenda, L.; Prufer, K.; Corteletti, R.; Scunderlick, D.; Mayle, F.E.; de Blasis, P.; et al. Uncoupling human and climate drivers of late Holocene vegetation change in southern Brazil. Sci. Rep. 2018, 8, 7800. [Google Scholar] [CrossRef] [PubMed]
- Hueck, K. As Florestas da América do Sul: Ecologia, Composição e Importância Econômica; Editora da Universidade de Brasília/Editora Polígono: São Paulo, Brazil, 1972; p. 466. [Google Scholar]
- Santos, K.L.; Peroni, N.; Guries, R.P.; Nodari, R.O. Traditional knowledge and management of feijoa (Acca sellowiana) in southern Brazil. Econ. Bot. 2009, 63, 204–214. [Google Scholar] [CrossRef]
- Adan, N.; Atchison, J.; Reis, M.S.; Peroni, N. Local knowledge, use and management of ethnovarieties of Araucaria angustifolia (Bert.) Ktze. in the plateau of Santa Catarina, Brazil. Econ. Bot. 2016, 70, 353–364. [Google Scholar] [CrossRef]
- Mattos, J.R. O Pinheiro Brasileiro, 2nd ed.; Artes Gráficas Princesa: Lages, Brazil, 1994. [Google Scholar]
- Reitz, R.; Klein, R.M. Araucariáceas; Herbário Barbosa Rodrigues: Itajaí, Brazil, 1966. [Google Scholar]
- Machado Mello, A.J.; Peroni, N. Cultural landscapes of the Araucaria forests in the northern plateau of Santa Catarina, Brazil. J. Ethnobiol. Ethnomed. 2015, 11, 51. [Google Scholar] [CrossRef] [Green Version]
- Barbizan Sühs, R.; Hettwer Giehl, E.L.; Peroni, N. Interaction of land management and araucaria trees in the maintenance of landscape diversity in the highlands of southern Brazil. PLoS ONE 2018, 13, e0206805. [Google Scholar] [CrossRef]
- Lira, R.; Casas, A.; Blancas, J. Ethnobotany of Mexico: Interactions of People and Plants in Mesoamerica; Springer: New York, NY, USA, 2016; p. 560. [Google Scholar] [CrossRef]
- Clement, C.R. Domesticação da floresta & subdesenvolvimento da Amazônia. In Grupo de Estudos Estratégicos Amazônicos, Caderno de Debates, Tomo XIV; Mendes dos Santos, G., Ed.; Instituto Nacional de Pesquisas da Amazônia: Manaus, Brazil, 2019; pp. 11–52. [Google Scholar]
- Eberhard, D.M.; Simons, G.F.; Fennig, C.D. Ethnologue: Languages of the World, 23rd ed.; SIL International: Dallas, TX, USA, 2020; Available online: http://www.ethnologue.com (accessed on 31 October 2020).
- Koch, A.; Brierley, C.; Maslin, M.M.; Lewis, S.L. Earth system impacts of the European arrival and Great Dying in the Americas after 1492. Quat. Sci. Rev. 2019, 207, 13–36. [Google Scholar] [CrossRef]
- Cordell, L.S.; McBrinn, M. Archaeology of the Southwest, 3rd ed.; Routledge: New York, NY, USA, 2016; p. 367. [Google Scholar]
- Politis, G.G. Moving to produce: Nukak mobility and settlement patterns in Amazonia. World Arch. 1996, 27, 492–511. [Google Scholar] [CrossRef]
- Caballero, J.; Casas, A.; Cortés, L.; Mapes, C. Patrones en el conocimiento, uso y manejo de plantas en pueblos indígenas de México. Estud. Atacameños 1998, 16, 181–195. [Google Scholar] [CrossRef] [Green Version]
- Coomes, O.T.; McGuire, S.J.; Garine, E.; Caillon, S.; McKey, D.; Demeulenaere, E.; Jarvis, D.; Aistara, G.; Barnaud, A.; Clouvel, P.; et al. Farmer seed networks make a limited contribution to agriculture? Four common misconceptions. Food Policy 2015, 56, 41–50. [Google Scholar] [CrossRef]
- Langlie, B.S.; Mueller, N.G.; Spengler, R.N.; Fritz, G.J. Agricultural origins from the ground up: Archaeological approaches to plant domestication. Am. J. Bot. 2014, 101, 1601–1617. [Google Scholar] [CrossRef] [PubMed]
- Casas, A.; Rangel-Landa, S.; Torres-García, I.; Pérez-Negrón, E.; Solís, L.; Parra, F.; Delgado, A.; Blancas, J.; Farfán, B.; Moreno, A. In situ management and conservation of plant resources in the Tehuacán-Cuicatlán Valley, Mexico: An ethnobotanical and ecological perspective. In Current Topics in Ethnobotany; Albuquerque, U.P., Alves-Ramos, M., Eds.; Research Signpost: Kerala, India, 2008; pp. 1–23. [Google Scholar]
- Camou-Guerrero, A.; Reyes-García, V.; Martínez-Ramos, M.; Casas, A. Knowledge and use value of plant species in a Rarámuri community: A gender perspective for conservation. Hum. Ecol. 2008, 36, 259–272. [Google Scholar] [CrossRef]
- Rival, L.M. Trekking through History: The Huaorani of Amazonian Ecuador; Columbia University Press: New York, NY, USA, 2002. [Google Scholar]
- Heckenberger, M.J.; Neves, E.G. Amazonian archaeology. Ann. Rev. Anthropol. 2009, 38, 251–266. [Google Scholar] [CrossRef] [Green Version]
- Neves, E.G. Rethinking the role of agriculture and language expansion for ancient Amazonians. In Rethinking the Andes–Amazonia Divide: A Cross-Disciplinary Exploration; Pearce, A.J., Beresford-Jones, D.G., Heggarty, P., Eds.; University College London Press: London, UK, 2020; pp. 211–220. [Google Scholar]
- Medina, J.T. (Ed.) The Discovery of the Amazon According to the Account of Friar Gaspar de Carvajal and Other Documents; The American Geographical Society: New York, NY, USA, 1934. [Google Scholar]
- Heckenberger, M.J.; Russell, J.C.; Fausto, C.; Toney, J.R.; Schmidt, M.J.; Pereira, E.; Franchetto, B.; Kuikuro, A. Pre-Columbian urbanism, anthropogenic landscapes, and the future of the Amazon. Science 2008, 321, 1214–1217. [Google Scholar] [CrossRef] [Green Version]
- MacNeish, R.S. A summary of the subsistence. In The Prehistory of the Tehuacan Valley. Volume 1. Environment and Subsistence; Byers, D.S., Ed.; University of Texas Press: Austin, TX, USA, 1967; Volume 1, pp. 290–309. [Google Scholar]
- Moreno-Calles, A.; Casas, A.; Blancas, J.; Torres, I.; Masera, O.; Caballero, J.; García-Barrios, L.; Pérez-Negrón, E.; Rangel-Landa, S. Agroforestry systems and biodiversity conservation in arid zones: The case of the Tehuacán Valley, Central México. Agrofor. Syst. 2010, 80, 315–331. [Google Scholar] [CrossRef]
- Vallejo, M.; Casas, A.; Blancas, J.; Moreno-Calles, A.I.; Solís, L.; Rangel-Landa, S.; Dávila, P.; Téllez, O. Agroforestry systems in the highlands of the Tehuacán Valley, Mexico: Indigenous cultures and biodiversity conservation. Agrofor. Syst. 2014, 88, 125–140. [Google Scholar] [CrossRef]
- Vallejo, M.; Casas, A.; Pérez-Negrón, E.; Moreno-Calles, A.I.; Hernández-Ordoñez, O.; Tellez, O.; Dávila, P. Agroforestry systems of the lowland alluvial valleys of the Tehuacán-Cuicatlán Biosphere Reserve: An evaluation of their biocultural capacity. J. Ethnobiol. Ethnomed. 2015, 11, 8. [Google Scholar] [CrossRef] [Green Version]
- Vallejo-Ramos, M.; Moreno-Calles, A.I.; Casas, A. TEK and biodiversity management in agroforestry systems of different socio-ecological contexts of the Tehuacan Valley. J. Ethnobiol. Ethnomed. 2016, 12, 31. [Google Scholar] [CrossRef] [Green Version]
- Campos-Salas, N.; Casas, A.; Moreno-Calles, A.I.; Vallejo, M. Plant management in agroforestry systems of rosetophyllous forests in the Tehuacán Valley, Mexico. Econ. Bot. 2016, 70, 254–269. [Google Scholar] [CrossRef]
- Maezumi, S.Y.; Alves, D.; Robinson, M.; Souza, J.G.; Levis, C.; Barnett, R.L.; Almeida de Oliveira, E.; Urrego, D.; Schaan, D.; Iriarte, J. The legacy of 4500 years of polyculture agroforestry in the eastern Amazon. Nat. Plants 2018, 4, 540–547. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Erickson, C.L. Historical ecology and future explorations. In Amazonian Dark Earths: Origin, Properties, Management; Lehmann, J., Kern, D.C., Glaser, B., Woods, W.I., Eds.; Kluwer: Dordrecht, The Netherlands, 2003; pp. 455–500. [Google Scholar]
- Clement, C.R.; Denevan, W.M.; Heckenberger, M.J.; Junqueira, A.B.; Neves, E.G.; Teixeira, W.G.; Woods, W.I. The domestication of Amazonia before European conquest. Proc. R. Soc. B 2015, 282, 20150813. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iriarte, J.; DeBlasis, P.; De Souza, J.G.; Corteletti, R. Emergent complexity, changing landscapes, and spheres of interaction in southeastern South America during the middle and late Holocene. J. Arch. Res. 2016, 25, 251–313. [Google Scholar] [CrossRef] [Green Version]
- Argollo, J. Aspectos geológicos. In Botánica Económica de los Andes Centrales; Moraes, M., Ollgaard, B., Kvist, L.P., Borchsenius, F., Balslev, H., Eds.; Universidad Mayor de San Andrés: La Paz, Bolivia, 2006; pp. 1–10. [Google Scholar]
- Josse, C.; Cuesta, F.; Navarro, G.; Barrena, V.; Becerra, M.T.; Cabrera, E.; Chacón-Moreno, E.; Ferreira, W.; Peralvo, M.; Saito, J.; et al. Physical geography and ecosystems in the Tropical Andes. In Climate Change and Biodiversity in the Tropical Andes; Herzog, S.K., Martínez, R., Jørgensen, P.M., Tiessen, H., Eds.; Inter-American Institute for Global Change Research (IAI) and Scientific Committee on Problems of the Environment (SCOPE): San José dos Campos, Brazil; Paris, France, 2011; pp. 152–169. [Google Scholar] [CrossRef]
- Young, K.R.; Ulloa, C.U.; Luteyn, J.L.; Knapp, S. Plant evolution and endemism in Andean South America: An introduction. Bot. Rev. 2002, 68, 4–21. [Google Scholar] [CrossRef]
- Pulgar Vidal, J. Geografía del Perú: Las ocho Regiones Naturales, la Regionalización Transversal, la Microregionalización, 9th ed.; Editorial Peisa: Lima, Peru, 1987; p. 244. [Google Scholar]
- Josse, C.; Cuesta, F.; Navarro, G.; Barrena, V.; Cabrera, E.; Chacón-Moreno, E.; Ferreira, W.; Peralvo, M.; Saito, J.; Tovar, A. Ecosistemas de los Andes del Norte y Centro-Bolivia, Colombia, Ecuador, Perú y Venezuela; Secretaría General de la Comunidad Andina: Lima, Peru, 2009. [Google Scholar]
- Purugganan, M.D. Evolutionary insights into the nature of plant domestication. Curr. Biol. 2019, 29, R705–R714. [Google Scholar] [CrossRef] [Green Version]
- Hufford, M.B.; Berny Mier y Teran, J.C.; Gepts, P. Crop biodiversity: An unfinished magnum opus of Nature. Annu. Rev. Plant Biol. 2019, 70, 727–751. [Google Scholar] [CrossRef]
- Milla, R.; Osborne, C.P.; Turcotte, M.M.; Violle, C. Plant domestication through an ecological lens. Trends Ecol. Evol. 2015, 30, 463–469. [Google Scholar] [CrossRef]
- Larson, G.; Piperno, D.R.; Allaby, R.G.; Purugganan, M.D.; Andersson, L.; Arroyo-Kalin, M.; Barton, L.; Climer Vigueira, C.; Denham, T.; Dobney, K.; et al. Current perspectives and the future of domestication studies. Proc. Natl. Acad. Sci. USA 2014, 111, 6139–6146. [Google Scholar] [CrossRef] [Green Version]
- Gepts, P. Domestication of plants. In Encyclopedia of Agriculture and Food Systems; Van Alfen, N.K., Ed.; Elsevier: San Diego, CA, USA, 2014; Volume 2, pp. 474–486. [Google Scholar]
- Fuller, D.Q.; Hildebrand, E. Domesticating plants in Africa. In The Oxford Handbook of African Archaeology; Mitchell, P., Lane, P.J., Eds.; Oxford University Press: Oxford, UK, 2013; pp. 1–19. [Google Scholar] [CrossRef]
- Olsen, K.M.; Wendel, J.F. A bountiful harvest: Genomic insights into crop domestication phenotypes. Annu. Rev. Plant Biol. 2013, 64, 47–70. [Google Scholar] [CrossRef] [Green Version]
- Piperno, D.R. The origins of plant cultivation and domestication in the New World tropics. Curr. Anthropol. 2011, 52, S453–S470. [Google Scholar] [CrossRef]
- Brown, T.A.; Jones, M.K.; Powell, W.; Allaby, R.G. The complex origins of domesticated crops in the Fertile Crescent. Trends Ecol. Evol. 2009, 24, 103–109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pickersgill, B. Domestication of plants in the Americas: Insights from Mendelian and molecular genetics. Ann. Bot. 2007, 100, 925–940. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, B.D. Documenting domesticated plants in the archaeological record. In Documenting Domestication. New Genetic and Archaeological Paradigms; Zeder, M.A., Bradley, D.G., Emshwiller, E., Smith, B.D., Eds.; University of California Press: Berkeley, CA, USA, 2006; pp. 15–24. [Google Scholar]
- Diamond, J. Evolution, consequences and future of plant and animal domestication. Nature 2002, 418, 700–707. [Google Scholar] [CrossRef] [PubMed]
| Mesoamerica | Central Andes | S. American Lowlands | |
|---|---|---|---|
| # species used | 6500 | 1206 | 6261 |
| # families | 265 | 156 | 236 |
| Main families with (# useful species; percent of # species used) | Fabaceae (699; 10.8) | Asteraceae (107; 9.4) | Fabaceae (840; 13.4) |
| Asteraceae (571; 8.8) | Fabaceae (88; 7.3) | Euphorbiaceae (262; 4.2) | |
| Cactaceae (438; 6.7) | Solanaceae (84; 7.0) | Rubiaceae (227; 3.6) | |
| Poaceae (335; 5.2) | Rubiaceae (32; 2.6) | Asteraceae (223; 3.6) | |
| Euphorbiaceae (205; 3.2) | Poaceae (28; 2.3) | Malvaceae (183, 2.9) | |
| Malvaceae (171; 2.6) | Rosaceae (28; 2.3) | Lauraceae (182; 2.9) | |
| Solanaceae (162; 2.5) | Euphorbiaceae (22; 1.8) | Poaceae (179; 2.8) | |
| Rubiaceae (159; 2.5) | Cactaceae (21; 1.7) | Myrtaceae (172; 2.7) | |
| Asparagaceae (143; 2.2) | Lamiaceae (21; 1.7) | Apocynaceae (162; 2.6) | |
| Apocynaceae (133; 2.1) | Amaryllidaceae (19; 1.6) | Annonaceae (154; 2.4) | |
| Lamiaceae (133; 2.1) | Apiaceae (18; 1.5) | Solanaceae (151; 2.4) | |
| Arecaceae (138; 2.2) | |||
| Main uses with (# species; percent of # species used) | Medicinal (3478; 53.5) | Medicinal (644; 53.0) | Medicinal (4017; 64.1) |
| Edible (1810; 27.9) | Edible (424; 34.9) | Manufacture (2175; 34.7) | |
| Fodder (1637; 25.2) | Environmental (315; 30.9) | Edible (1719; 27.4) | |
| Construction (1224; 18.8) | Manufacture (229; 18.8) | Construction (1683; 26.8) | |
| Fuel (883; 13.6) | Construction (169; 13.9) | Fodder (667; 10.7) | |
| Habit with (# species; percent of # species used) | Herbs (2619; 40.3) | Herbs (567; 47.0) | Trees (3319; 53) |
| Trees (1861; 28.6) | Shrubs (339; 28.1) | Herbs (1276; 20.4) | |
| Shrubs (1411; 21.7) | Trees (248; 20.5) | Shrubs (1002; 16) | |
| Lianas (499; 7.7) | Liana (52; 4.3) | Lianas (651; 10.4) | |
| Species gathered | 6000 (92.3%) | 778 (64.5%) | 6178 (98.6%) |
| Species managed | 1555 (23.9%) | 428 (35.5%) | 742 (11.8%) |
| Domestication | |||
| Incipient | 727 (11.2%) | Not available yet | 517 (8.2%) |
| Semi | 170 (2.7%) | 304 (25.2%) | 38 (0.6%) |
| Full | 251 (3.9%) | 124 (10.3%) | 45 (0.7%) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Clement, C.R.; Casas, A.; Parra-Rondinel, F.A.; Levis, C.; Peroni, N.; Hanazaki, N.; Cortés-Zárraga, L.; Rangel-Landa, S.; Alves, R.P.; Ferreira, M.J.; et al. Disentangling Domestication from Food Production Systems in the Neotropics. Quaternary 2021, 4, 4. https://doi.org/10.3390/quat4010004
Clement CR, Casas A, Parra-Rondinel FA, Levis C, Peroni N, Hanazaki N, Cortés-Zárraga L, Rangel-Landa S, Alves RP, Ferreira MJ, et al. Disentangling Domestication from Food Production Systems in the Neotropics. Quaternary. 2021; 4(1):4. https://doi.org/10.3390/quat4010004
Chicago/Turabian StyleClement, Charles R., Alejandro Casas, Fabiola Alexandra Parra-Rondinel, Carolina Levis, Nivaldo Peroni, Natalia Hanazaki, Laura Cortés-Zárraga, Selene Rangel-Landa, Rubana Palhares Alves, Maria Julia Ferreira, and et al. 2021. "Disentangling Domestication from Food Production Systems in the Neotropics" Quaternary 4, no. 1: 4. https://doi.org/10.3390/quat4010004
APA StyleClement, C. R., Casas, A., Parra-Rondinel, F. A., Levis, C., Peroni, N., Hanazaki, N., Cortés-Zárraga, L., Rangel-Landa, S., Alves, R. P., Ferreira, M. J., Cassino, M. F., Coelho, S. D., Cruz-Soriano, A., Pancorbo-Olivera, M., Blancas, J., Martínez-Ballesté, A., Lemes, G., Lotero-Velásquez, E., Bertin, V. M., & Mazzochini, G. G. (2021). Disentangling Domestication from Food Production Systems in the Neotropics. Quaternary, 4(1), 4. https://doi.org/10.3390/quat4010004