Abstract
Urban densification threatens biodiversity, yet conventional greenspace expansion is constrained by limited land availability. Living wall systems (LWS) offer potential biodiversity enhancement through vertical green infrastructure, though their ecological value remains underexplored. This study evaluated the biodiversity performance of three LWS in Plymouth, UK, using multi-taxa surveys to assess invertebrate communities, bird assemblages, and bat activity. A scoping review of 2638 publications revealed limited research on LWS biodiversity, with only 27% of biodiversity-focused papers referencing specific species. Field surveys employed standardised protocols including flower-visiting pollinator observations, spider assessments, soil invertebrate extraction using Tullgren funnels, acoustic bird monitoring, and bat emergence surveys across soil-based and hydroponic systems. Results demonstrated that soil-based LWS supported significantly higher invertebrate diversity than hydroponic systems, with 481 soil invertebrates recorded across 19 families. Plant species composition strongly influenced biodiversity outcomes, with Hedera helix, Erigeron karvinskianus, and Lonicera japonica attracting the most pollinator species (5 each). Bird abundance was significantly higher at LWS sites compared to control areas, with confirmed breeding by three species. However, current UK Biodiversity Net Gain frameworks undervalue LWS contributions due to their classification as artificial habitats. These findings indicate that appropriately designed soil-based LWS can deliver meaningful urban biodiversity benefits when integrated with strategic plant selection and species-based valuation approaches.
1. Introduction
Global urbanisation is accelerating, with an estimated 68% of the world’s population projected to live in urban areas by 2050 [1]. This expansion is a major driver of biodiversity loss, as natural and semi-natural habitats are replaced by high-density built infrastructure [2,3]. In the UK, this trend is driven by housing targets and brownfield redevelopment priorities [4]. In densely built environments, opportunities for conventional horizontal greenspace are limited, prompting growing interest in green infrastructure solutions that integrate vegetation into the vertical built environment [5]. Among these, living wall systems (LWS)—modular panels of vegetation with hydroponic or substrate-based rooting media attached vertically to buildings—have gained attention for their potential to contribute to urban biodiversity [6,7].
Green infrastructure is a key component of urban Nature-based Solutions, offering a wide range of human health and ecosystem service benefits [8]. When thoughtfully designed, it can support substantial biodiversity [9,10] and serve as a multifunctional response to the environmental challenges of urbanisation. In addition to enhancing urban aesthetics and mitigating thermal stress [11,12,13,14], green infrastructure contributes to vital ecosystem services such as air purification, noise reduction, natural flood management and façade protection [8,15,16,17].
Vertical surfaces make up a high percentage of the surface area in high-rise city developments, highlighting the potential scale of their contribution. Living wall systems could offer habitats for a wide range of organisms, reducing urban habitat loss, fragmentation and isolation. However, the biodiversity potential of LWS remains underexplored. A systematic review by Tiago et al. [18] found just 56 peer-reviewed papers focused on biodiversity in LWS or green roofs, with few examining multiple taxonomic groups or temperate climate contexts. Most studies focused on arthropods, and only four assessed more than one faunal group. This underscores significant research gaps, especially regarding soil invertebrates and the role of rooting substrate in shaping ecological outcomes [19,20].
While LWS are valued primarily for their visual and thermal performance, their design parameters, particularly substrate and plant choice, are critical to their ecological value [21]. Native plant species adapted to desiccating urban microclimates enhance the recruitment of native fauna and improve ecosystem resilience [7,22,23]. In contrast, incorporating non-native plants can increase the risk of invasive species spread [7,24,25,26]. Soil-based growth media outperform artificial substrates in water retention, nutrient cycling, and sustaining beneficial microbial and invertebrate communities [27,28,29]. Soil invertebrates, such as mites (Acari), springtails (Collembola), and millipedes (Diplopoda), play key roles in decomposition, nutrient regulation, and ecological function [28]. More generally, LWS design must balance biodiversity performance with ease of maintenance and water use efficiency. Modular systems allow for easy replanting, and innovations such as rainwater harvesting and greywater reuse can improve long-term sustainability [30,31].
Biodiversity Net Gain (BNG) is a concept that focuses on enriching biodiversity as part of the development process, resulting in a natural environment that is measurably better once works are completed. In February 2024, BNG became a statutory requirement in England, requiring new developments to have a 10% uplift in biodiversity above predevelopment level [32]. This compliance requirement uses Defra’s Biodiversity Metric 4.0 methodology [33] to score the pre-development extent of UKHab index areas and compare scores to proposed post-development habitat. BNG can be inset (on-site) or offset (off-site biodiversity gain compensation), with the former preferred where possible. Typically for small-scale developments, the gain is inset, with an average of 80% as on-site mitigation [34].
An alternative approach to assessing the ecological impacts of development is the Wallacea biodiversity assessment framework [35], which incorporates field-based measurements of ecological richness, abundance, and functional diversity. By capturing fine-scale biodiversity data—including that from vertical and soil systems—this method may better reflect the real-world ecological value of LWS.
This study uses field-based biodiversity measurements to evaluate how soil-based LWS can contribute to biodiversity in urban developments. Specifically, we investigate how planting choices and rooting substrates influence aboveground and soil dwelling invertebrate communities and assess the ecological value of LWS.
1.1. Objectives
- To identify knowledge gaps in current LWS biodiversity research.
- To evaluate the role of plant species choice in supporting aboveground and soil-dwelling invertebrates on LWS.
- To assess the contribution of soil-based growth media to invertebrate diversity and ecological function.
- To evaluate the biodiversity value of LWS.
1.2. Scoping Review of Living Walls Biodiversity Benefits Literature
A scoping review of peer-reviewed literature on green infrastructure was conducted using Scopus, Web of Science, PubMed, Google Scholar, and ScienceDirect, following the methodology described by Foo et al. [36]. (Figure 1). The review aimed to identify ecosystem services, biological taxa, and plant rooting substrates most frequently referenced in the living walls literature. The process adhered to PRISMA (Preferred Reporting Items for Systematic Reviews and Meta-Analyses) guidelines [37], using predefined keyword combinations shown in the results tables. Publications were identified through thematic keyword searches rather than individual relevance screening, consistent with a scoping review approach. The search yielded 2638 journal articles published between 1970 and 2025 (Table 1), with fewer than 10% published before 2014 and over half appearing in the last five years (2020–2025). A similar key-word search showed that the most frequently referred to countries (taken as country of research location) in the LWS literature in order of frequency were USA, China, Italy, Spain, South Korea, Netherlands, Sweden, Poland, Portugal and Brazil.
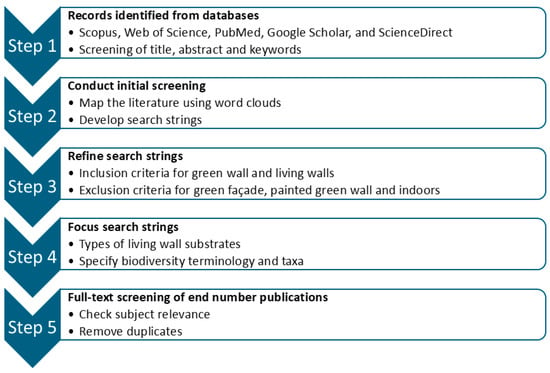
Figure 1.
Flow diagram outlining the literature scoping review process.

Table 1.
Boolean operators search returns for living wall academic publications including specific key words related to biodiversity. Inclusion criteria included peer-reviewed journal articles, conference papers, and review articles published in English from 2000 onwards. Studies included all geographical locations (see Supplementary Materials).
Boolean operators and search strings were used to select “Living* Wall*” AND “Ecosystem Service*” papers containing specific keywords in titles and abstracts from 2015–2025. The review identified climate as the most frequently reported ecosystem service benefit of LWS (43% of 46 papers) with health (39%), and biodiversity (30%) also featuring strongly. In relation to biodiversity, the search was limited exclusively to six terms, ranked by frequency of occurrence: species, birds, native plants, invertebrates, and bats (Table 1). However, out of the 500 papers reporting on living walls and biodiversity only 134 (27%) made specific reference to a species, suggesting papers make general reference to biodiversity benefits but not specific components of biodiversity.
The analysis of rooting substrates was restricted to soils, hydroponic (non-soil) systems, and artificial substrates. Of these, 81 publications referred to soils, 16 to hydroponic systems, and 20 to perlite. Additional artificial substrates (rockwool and vermiculite) were recorded at lower frequencies (Table 2). Of the mineral substrates, restricted to clay, biochar, peat, and clay pellets, clay was most frequently mentioned (13 papers). Twelve publications were identified that included the combined keywords: green wall, biodiversity, and soils.
Table 2.
Boolean operators search returns for living wall academic publications including specific key words related to the plant root substrates. Inclusion criteria included peer-reviewed journal articles, conference papers, and review articles published in English from 2000 onwards.
1.3. Floristic Composition and Growth Form of Temperate Living Wall Vegetation
A survey was carried out on the plant species composition of 15 LWS in the UK (Supplementary Materials). Of the 173 plant species commonly planted, the majority (65%) were non-native, with 35% being native. In the UK, the predominant LWS plant growth form is perennial evergreen, followed by perennial deciduous (Figure 2).
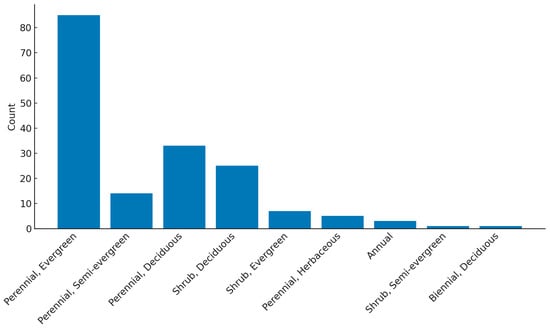
Figure 2.
Growth form and life history composition of plants in temperate living wall systems. Results are based on 173 plant species recorded across 15 LWS in the UK.
2. Study Sites and Living Wall Systems
Plymouth, located in south-west England, has a temperate oceanic climate [38]. Three distinct living wall systems (LWS) within the city were evaluated: the Sustainability Hub LWS, the Genesis Building LWS, and the Sherford LWS (Figure 3).
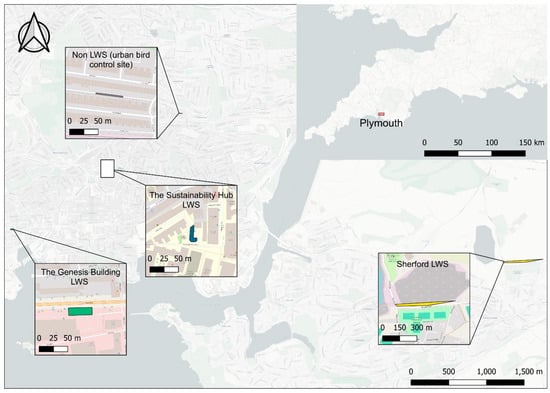
Figure 3.
Study area map showing locations of three living wall installations and a control site for bird surveys (without a living wall installation) in the city of Plymouth, UK.
The Sustainability Hub LWS is installed on a two-storey building at the University of Plymouth. Established in 2019, it comprises a modular Fytotextile system designed by Scotscape [39]. The system uses a multi-layered fabric structure with a waterproof inner lining, an absorptive middle layer, and a permeable felt outer layer, forming discrete planting pockets filled with general-purpose potting compost. These 1.5 × 1 m panels were mounted on a wooden frame fixed to the building’s exterior.
The Genesis Building LWS, on a four-storey office building, employs a modular, soil-free, hydroponic plug-plant system developed by Biotexture [40]. Plants are supported in a highly absorbent synthetic rooting medium, with irrigation and nutrients delivered by a pump-driven drip system. The Sustainability Hub and Genesis Building systems both operate with this irrigation approach.
The Sherford LWS, by contrast, is a large-scale, soil-based system constructed on a retaining embankment along Hercules Road. It consists of a geogrid framework with soil-filled pockets and is entirely rainfed. The wall reaches up to 10 m in height, extends approximately 380 m in length, and accommodates around 26,000 plants.
Across all three LWS, planting combined native and non-native evergreen shrubs and herbaceous species typical of temperate climates. Species were chosen for their durability, low maintenance requirements, and visual impact.
2.1. Survey Methods for Multi-Taxa Living Wall Biodiversity Assessments
A multi-taxa biodiversity assessment approach was used to evaluate the ecological value of Local Wildlife Sites (LWS), following the Wallacea Method [35], which recommends using at least five biodiversity metrics tailored to the habitat context. This approach captures species richness and abundance across faunal groups representing distinct ecological functions and habitat interactions [41,42]. The selected taxa included pollinating bees and hoverflies, predatory spiders, below-ground soil invertebrates, bats, and birds—groups commonly used as ecological indicators due to their biodiversity relevance, mobility, and the ecosystem services they provide (e.g., pollination, pest control, soil health) [18,41].
Sampling effort was tailored to the ecological traits and seasonal activity of each group. Above-ground invertebrates (bees, hoverflies, spiders) were surveyed using ten replicated 2 × 2 m plots at each of two soil-based LWS: Sustainability Hub and Sherford. Below-ground invertebrates were sampled at one LWS (Sustainability Hub) using five planting pockets per plant species across four selected species to capture root and detritivore interactions. Bird and bat surveys were conducted at all three LWS, with bird monitoring also including a control urban site.
Identification was conducted to species level where feasible, or to morpho-species level based on shared morphological traits. Analysis was carried out on taxonomic groupings (e.g., bees, spiders), ecological functions (such as pollination and predation), and plant–invertebrate associations, enabling a comprehensive assessment of biodiversity and ecosystem service provision across the LWS.
2.2. Invertebrate Surveys
For above-ground invertebrate assessments, three major arthropod taxa were selected: bees (Insecta: Hymenoptera), hoverflies (Insecta: Diptera), and spiders (Araneae). These groups are taxonomically diverse and provide essential ecosystem services, including pollination and biological control. Bees and hoverflies, which may be either resident or transient on LWS, serve as effective indicators of floral resource availability and habitat connectivity. Spiders, typically resident predators, are valuable indicators of local invertebrate community health and trophic structure.
Below-ground invertebrates were also surveyed due to their critical roles in soil health, nutrient cycling, and plant–soil interactions. To capture a representative range of plant–invertebrate associations, eight commonly used non-native and one native plant species were selected based on their growth forms and flowering phenology (Table 3).
Table 3.
Surveyed plant species used in invertebrate assessments, categorised by flowering time, growth form, and native status.
Surveys of both above-ground (flower-visiting bees and hoverflies, and spiders) and below-ground (root- and detritivore-feeding) invertebrates were conducted during the primary activity period for most UK invertebrates, from May to November. Adult specimens were identified to species level where possible, or to morpho-species level when species identification was difficult in the field. Morpho-species refer to taxonomically unresolved groups distinguished by shared morphological traits, typically classified at the family to genus level.
2.3. Assessment of Above-Ground Flower-Visiting Bees, Hoverflies and Predatory Spiders
For Hymenoptera (specifically bees) and Diptera (specifically hoverflies), three repeated surveys were conducted during June and July 2021. Surveys were carried out under optimal UK weather conditions, with temperatures ranging from 17 to 27 °C, mostly sunny skies with partial cloud cover, and wind speeds between 7 and 13 mph. Ten 2 × 2 m survey plots were established at each of the two soil-based LWS. Each survey consisted of three repeated 5-min observation periods per plot.
During each observation period, the number of individual bees and hoverflies landing within the 2 × 2 m plots was recorded, along with species identity, associated plant species, and behavioural observations. Photographs were taken to support later identification and verification.
For Araneae (spiders), the same ten 2 × 2 m plots per soil-based LWS were surveyed once per month from June to November. When spider webs were present, individuals were gently coaxed out by triggering the web to facilitate observation. The number of individual spiders observed within each plot was recorded, and individuals were photographed in situ to aid identification and to document their location within the LWS, whether on plant material or structural components [43,44].
2.4. Below-Ground Soil Meso- and Macrofaunal Invertebrate Sampling Methods
Soil invertebrate sampling was conducted on the Sustainability Hub LWS, focusing on four plant species representing three distinct growth forms. These included the herbaceous ornamental Erigeron karvinskianus, the grass-like Carex oshimensis, and two evergreen shrubs, Fatsia japonica and Sarcococca confusa (Table 3). For each plant species, five planting pockets on the lower (accessible) section of the wall were randomly selected.
Once selected, the above-ground plant material was carefully removed. The roots and associated soil from each pocket were transferred into individual Tullgren funnels. Glass collection bottles containing 50 mL of 70% Industrial Denatured Alcohol (IDA) were placed beneath each funnel to collect invertebrates displaced by the heat and light. Funnels were illuminated with 20-watt light bulbs for a continuous 14-day period. Collection bottles were checked every two days, and IDA levels topped up regularly to maintain preservation quality.
At the end of the extraction period, the collection bottles were removed and the invertebrates sorted into major taxonomic groups (e.g., Acariformes, Collembola, Hemiptera, Hymenoptera). Earthworms were identified to family level (Lumbricidae). Where possible, other soil invertebrates (e.g., Acariformes) were also classified to family level to enable calculation of family-level diversity indices.
2.5. Avian Acoustic Monitoring Methods
Bird song recordings were conducted at all three LWS sites and at a control site representing a typical urban street in Plymouth (terraced housing) without green infrastructure. Surveys were carried out on three separate occasions between 26 July and 20 September 2023.
Acoustic data were collected using the Merlin ID app [45] on a mobile device. Recordings began at dawn and continued for 90 min under suitable weather conditions. Data included species identification, number of individuals, and behavioural codes based on Merlin classifications. Visual aids were used to assist with bird identification in the surrounding area. Seagulls (Laridae) were excluded from the analysis due to their wide-ranging behaviour and ubiquitous presence in coastal cities such as Plymouth.
2.6. Bat Emergence Surveys
Three bat dusk emergence and dawn re-entry surveys were carried out on all three LWS in July to September 2023. Dusk surveys began 15 to 30 min before sunset and continued for up to two hours. Dawn surveys began two hours before sunrise and continued for up to 30 min after sunrise. Surveys were only conducted in suitable weather conditions, avoiding rain, strong winds, and temperatures below 10 ° C. Evidence of bat emergence was observed visually using a night vision scope and bat activity recorded using an Anabat SD2 full-spectrum bat detector.
2.7. Statistical Analysis of Biodiversity Indicators
All statistical analyses were conducted using R (version 4.3.2; R Core Team, 2023), statistical software [46]. Data visualisations were produced using the ggplot2 package [47]. A significance level of p < 0.05 was applied to all tests.
Differences in biodiversity indicators among the three LWS were assessed using one-way analysis of variance (ANOVA). Prior to analysis, data were checked for normality and equality of variances across groups using the Shapiro-Wilk test of residuals [48]. Where necessary, non-normally distributed data were log-transformed to meet the assumptions of ANOVA. Post hoc pairwise comparisons were performed using Tukey’s Honest Significant Difference (HSD) test to identify significant differences between LWS sites.
3. Results
3.1. Above-Ground Plant-Invertebrate Associations on Living Walls
Visiting pollinator diversity varied among the seven plant species on the two urban LWS (Figure 4). Erigeron karvinskianus (Mexican fleabane), Hedera helix (Ivy) and Lonicera japonica (Japanese honeysuckle) attracted the highest number of pollinator species (5 species each). Although H. helix is the only native species, E. karvinskianus and Lonicera japonica are both fully naturalised species in the UK with similarities to closely related native species. Notably, visitation rates to E. karvinskianus were 2.9× higher than to the other two plant species, but this was driven by a clear preference of Apis mellifera for E. karvinskianus. Geranium cantabrigiense (cranesbill) attracted the largest number of pollinator visits, but only from two Bombus species, with visitation dominated by Bombus pratorum (Early Bumblebee; 22 individuals; Figure 4). Eighty-four per cent of the pollinators were observed to be visiting flowers (pollinator behaviour) with 13% resting and 3% involved in other behaviours.
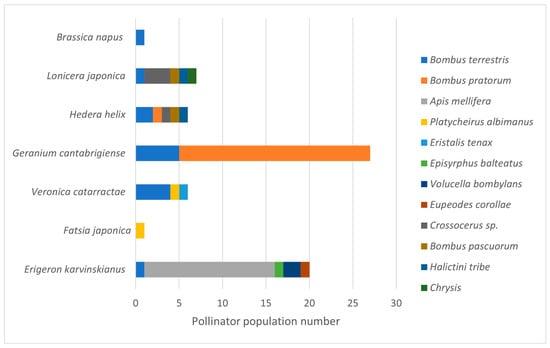
Figure 4.
Pollinator bee and hoverfly visitors to native and non-native plant species with data collected from two soil-based living wall systems with shared plant species. Data represents the total numbers from three repeats of surveys on two LWS in June and July 2021.
Similarly, spider abundance and diversity varied across the two soil-based LWS surfaces and 12 plant species (Figure 5). Overall, 274 adult spider specimens from 12 taxa were sampled, with the three most abundant spider (Araneid) taxa being Araneus diadematus (28.5% of the total spider abundance), Zygiella x-notata (27.5%) and Steatoda nobilis (14.2%).
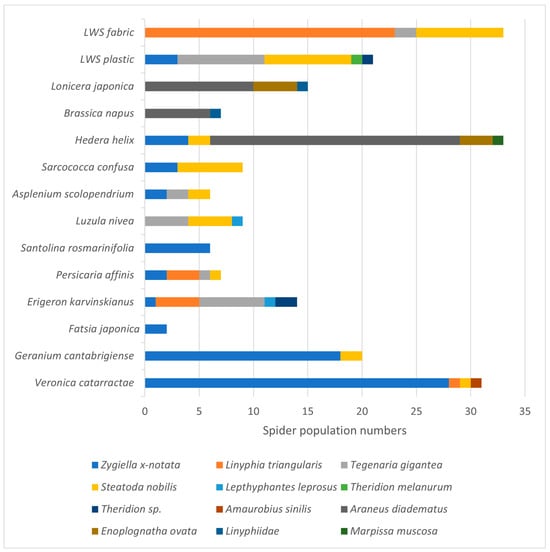
Figure 5.
Resident spiders associated with common native and non-native plant species grown in soil-based LWS. Data represents the totals from six repeated monthly single surveys of two LWS, June and November 2021.
H. helix, E. karvinskianus and the plastic surround of the LWS hosted the most resident araneid taxa (5 species each; Figure 5). H. helix and the textile fabric of the LWS supported the largest number of spiders (33 individuals each), with the native H. helix hosting the largest numbers of Araneus diadematus (European garden spider). Veronica catarractae (Parahebe ‘Avalanche’) hosted the next highest abundance of spiders (31 individuals from 4 taxa). Parahebe (see Table 3) has a rigid but open structure favoured by web spinning spiders, as shown by the high numbers of the missing sector orb weaver (Zygiella x-notata) recorded on this plant.
3.2. Soil Invertebrates Associated with Living Wall Plants
Tullgren sampling of soil from LWS planting pockets from four plant species yielded a total of 481 individual invertebrates across 19 different families. Over half of the individuals (258) belonged to the Mesostigmata (soil mites), making it the most abundant order, followed by the (60) Collembola (springtails). Plant species had a significant effect on soil invertebrate community composition (Figure 6). Invertebrate abundance was found to vary significantly with plant species (Figure 6a; F3, 55 = 24.8, p ≤ 0.001). Soil from Carex oshimensis hosted significantly more soil invertebrates (272) than all other plant species, with Sarcococca confusa hosting an intermediate (144), but significantly higher (p ≤ 0.05) soil invertebrate abundance than both E. karvinskianus (53) and Fatsia japonica (12).
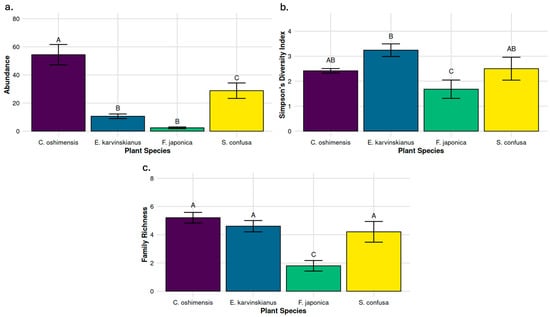
Figure 6.
The effect of plant species on: (a) below-ground invertebrate abundance; (b) Simpson’s diversity metric of below-ground communities using counts at the family level; (c) Below-ground family richness (number of families observed). Values represent means (n = 5) and bars show +/− standard error. Significant differences (p ≤ 0.05) between plant species means are denoted by differences in superscript capital letters.
There was a significant difference in Simpson’s Diversity of soil invertebrates between plant species (Figure 6b; F3,16 = 3.91, p < 0.05). E. karvinskianus had a significantly higher Simpson’s Diversity score than that of F. japonica (p = 0.017) but did not significantly differ from the other two plant species. Finally, soil invertebrate family richness (number of families observed) also showed a significant difference across plant species (Figure 6c; F3,16 = 9.07, p < 0.001). F. japonica showed significantly lower family richness than E. karvinskianus (p = 0.005), C. oshimensis (p = 0.001), and S. confusa (p = 0.016).
3.3. Avian Acoustic Survey and Bat Survey Results
Acoustic monitoring recorded a total of 489 individuals and 32 bird species. Over half of the individuals (285) belonged to the genus Larus (seagulls). Seagulls were excluded from the analysis because of their wide-roaming and ubiquitous presence in coastal cities.
Significant differences in bird abundance were observed among sites (Figure 7a; F3, 55 = 24.8, p ≤ 0.001). The Sherford and Genesis Building LWS attracted significantly (p ≤ 0.001) more birds, 28 and 24, respectively. The Sustainability Hub LWS showed an intermediate (15) but significantly higher (p ≤ 0.001) bird abundance than the non-LWS control area (2).
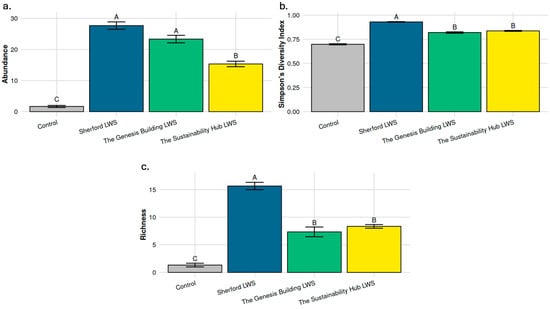
Figure 7.
Bird species abundance, diversity and number associated with three Plymouth living wall sites and a control urban street. (a) Bird abundance; (b) Simpson’s diversity Index; (c) Species number. Values represent means (n = 3) and bars show +/− standard error. Significant differences (p ≤ 0.05) between LWS means are denoted by differences in superscript capital letters.
Bird diversity, measured by Simpson’s Diversity Index, also differed significantly between sites (Figure 7b; F3,16 = 3.91, p < 0.05). The Sherford LWS had significantly higher diversity than the Genesis and Sustainability Hub LWS (p = 0.017), which supported intermediate but significantly higher bird diversity than the non LWS control area (p ≤ 0.001).
Species richness followed a similar pattern (Figure 7c; F3,16 = 9.07, p < 0.001). The Sherford LWS supported the greatest number of species (p = 0.017), significantly higher than Genesis Building and Sustainability Hub, which were intermediate but significantly greater (p ≤ 0.001) than the non-LWS control area.
Three species were observed using LWS for nesting or feeding: House sparrow (Passer domesticus), with four pairs confirmed breeding on the Genesis Building LWS, and Eurasian robin (Erithacus rubecula) and Eurasian blackbird (Turdus merula) as possible breeders. These findings suggest that LWS not only attract more birds but also provide resources for breeding and foraging, with Sherford offering the greatest habitat complexity and resource availability for birds.
Bat surveys found no individuals using the green wall, despite being present in the surrounding area.
4. Discussion
Biodiversity is particularly important for human health and wellbeing in urban environments where access to green spaces can be limited [49,50,51]. LWS in high-density urban conurbations represent an opportunity for biodiversity enhancement without taking up valuable space for development [18]. Our results demonstrate that LWS can provide significant biodiversity benefits, but that this depends on their design from choices about planting to underlying substrates for plant growth.
LWS have the potential to enhance urban biodiversity by acting as ecological steppingstones or corridors that facilitate wildlife movement across fragmented landscapes [52,53]. These structures can connect isolated habitat patches, thereby improving ecological resilience in urban areas facing pressures from climate change, habitat loss, and local species extinction [53]. While such connectivity can support native species, it may also inadvertently aid the spread of non-native organisms, as urban environments often facilitate the establishment and dispersal of invasive species through reduced biotic resistance and enhanced connectivity [54,55,56].
Evidence from Partridge and Clark [57] indicates that green roofs support greater abundance and richness of birds and arthropods compared to conventional roofs, particularly during migration and breeding seasons. This suggests that vegetated vertical structures, such as LWS, may similarly contribute to habitat quality and connectivity in urban environments [50,58,59].
In addition to supporting general biodiversity, LWS may play a specific role in conserving native pollinators. Tonietto et al. [60] demonstrated that green roofs planted with diverse native forbs provide valuable foraging and nesting resources for native bees, highlighting their potential as pollinator habitats in cities. Braaker et al. [61] demonstrated that arthropod diversity on green roofs increased with greater habitat connectivity and plant species richness, regardless of substrate depth or roof height. If similar principles apply to LWS, these findings suggest that plant diversity is a key driver of biodiversity, but not the height above ground.
These findings underscore the importance of integrating LWS into urban planning frameworks, particularly within the context of biodiversity objectives, where enhancing ecological connectivity and supporting functional species groups are key priorities [18].
4.1. Optimising Living Wall Systems (LWS) for Biodiversity: The Role of Plant Species Selection
Plant composition strongly shapes invertebrate communities in LWS. Insect-pollinated flowering plants provide nectar and pollen for pollinators, while others support high densities of predatory taxa such as spiders or soil-dwelling invertebrates [24,62]. Conversely, some species, such as Fatsia japonica, offer low ecological value, suggesting that alternative species should be prioritised. Perennial evergreens, particularly Hedera helix (ivy), although an important late summer pollen/nectar source, can dominate LWS and suppress plant diversity by outcompeting annuals, deciduous perennials, and shrubs for light and space. This was evident at the Sherford LWS, where ivy covered more than 80% of the installation within three years of planting.
Plant selection in LWS is not determined solely by ecological function but also by practical and aesthetic considerations. Year-round visual appeal, species availability, and maintenance requirements strongly influence design choices. In the UK, evergreen and non-deciduous species dominate LWS installations because their persistent foliage increases visual appeal. Perennials are generally favoured over larger shrubs that require regular trimming. The predominance of non-native species in LWS reflects both the limited pool of native evergreen plants in the UK flora and the urban greening preference for evergreen perennials and shrubs [62,63].
Birds, invertebrates, and soil organisms are widely recognised as indicators of habitat quality. The structural diversity hypothesis predicts that species richness and abundance increase with ecosystem complexity [64,65]. This highlights the importance of thoughtful species selection in LWS design [63]. While differences in biodiversity value between native and non-native plants are evident, they may be less pronounced in urban environments dominated by generalist species. Nonetheless, plant choice remains a critical determinant of the ecological value of LWS.
4.2. Soil-Based Growth Media and Its Contribution to Biodiversity in Living Wall Systems
Our results clearly demonstrated that LWS soils can support extensive belowground invertebrate communities. Unlike inert substrates, soil-based media support microbial-driven soil processes such as organic matter decomposition and nutrient cycling create resource heterogeneity and sustain diverse trophic networks [27,66,67]. Symbiotic microbes, including mycorrhizal fungi and nitrogen-fixing bacteria, enhance plant vigour and diversity, reinforcing the structural and functional stability of LWS [67]. This ecological foundation underpins the greater biodiversity value of soil-based systems compared with artificial media [18,19,27].
Collectively, these mechanisms establish a multifunctional soil ecology that supports invertebrate communities, improves nutrient cycling, and provides prey for higher trophic groups such as birds and bats [18,68,69]. Furthermore, plant species selection strongly influences rhizosphere microbial composition and associated soil fauna, making plant choice critical for sustaining biodiversity and enabling microbial colonisation [70].
Design choices also influence biodiversity outcomes. Soil-based substrates offer richer organic resources and structurally more complex habitats than artificial growth media [14]. For example, Madre et al. [19] found that modular pocket systems filled with sphagnum moss supported significantly higher spider richness and abundance than felt-layer facades, climbing plant facades, or bare walls. These results were attributed to the moist microhabitats and detritivore communities sustained by the moss substrate. Beyond invertebrates, soil-based LWS provide habitat value for vertebrates. Urban birds utilise them for foraging, nesting, and shelter, while crevice-inhabiting bat species may exploit cavities behind installations [18,69]. Systems incorporating diverse plantings further enhance these opportunities, strengthening their role in urban wildlife conservation [9,18,71].
Finally, soil biodiversity underpins multiple ecosystem functions fundamental to urban greenspaces, including carbon cycling, organic matter decomposition, water regulation, and pathogen control [72]. These processes are closely linked to the richness and functional diversity of soil invertebrates, reinforcing the ecological importance of soil-based substrates within LWS.
4.3. Possible Approaches for Biodiversity Valuation of LWS
Our findings show that living wall systems (LWS), particularly soil-based designs, can support diverse assemblages of plants, birds, and invertebrates. Yet, both in the UK and internationally, no biodiversity valuation framework exists that adequately accounts for the ecological potential of human-designed habitats such as LWS.
Current biodiversity valuation frameworks used in development mitigation fall broadly into two categories. The first, represented by the UK Biodiversity Net Gain (BNG) metric [33], is habitat-based: biodiversity value is determined by habitat classification, quality (e.g., naturalness or potential for creation), and area. The second category is species-based, exemplified by the Wallacea biodiversity assessment method [35], the most widely applied global credit system for biodiversity recovery. This framework uses composite scoring based on species richness and abundance across at least five indicator groups, including functional taxa such as soil invertebrates and zoological groups such as birds and bees.
Development-led compliance mechanisms have significant potential to fund LWS and deliver biodiversity gains in high-density urban areas where ground space is limited. However, under the BNG metric, LWS are classified within the category Urban–Built-up Areas and Gardens, an artificial habitat type associated with non-native species and assigned low distinctiveness scores. Consequently, LWS receive minimal credit for biodiversity enhancement [73] and are currently unsuitable for offsetting habitat losses arising from new developments [74]. This reveals a clear disconnect between the demonstrated ecological value of LWS and their treatment in planning compliance frameworks.
Urban habitats that provide food and shelter for rare or legally protected species may be particularly valuable. In the UK, all bat species and their roosts are protected under the Wildlife and Countryside Act (1981) [75]. While bats were observed foraging near the LWS in Plymouth, no individuals were recorded entering or emerging from the structures. This suggests that current LWS designs do not yet provide suitable roosting opportunities. Nevertheless, modular frame systems, such as the Sustainability Hub installation, show potential for supporting crevice-dwelling bats, with possible future use as hibernacula or maternity roosts [76]. Further research is required to evaluate the conservation value of LWS for bats in urban environments.
In the absence of an appropriate valuation system, developers are unlikely to adopt LWS as biodiversity interventions and will instead seek alternative offsite options, representing a missed opportunity for urban biodiversity enhancement. Species-based valuation methods may offer a more appropriate means of assessing LWS, especially where all species are treated equally without weighting for rarity, naturalness, or native status. While these methods can be more time-intensive and costly than area-based assessments, advances in AI-powered photo and bioacoustic recognition technologies are making species surveys increasingly cost-effective and scalable [77].
Future valuation frameworks should also consider the full suite of ecosystem services provided by LWS, including climate regulation, aesthetic value, air pollution mitigation, natural flood management, and biodiversity support [78]. Although LWS are expensive to install and maintain and vulnerable to system failure—particularly from irrigation breakdowns—optimising their biodiversity value requires an integrated approach. This should combine careful design, species selection, faunal habitat provision, water efficiency, and maintenance, all of which are critical for building resilient, multifunctional LWS in urban landscapes.
5. Conclusions
Urban biodiversity is increasingly threatened by development, making innovative strategies essential for ecological resilience. LWS offer a viable solution in high-density cities where ground space is limited, but their potential remains underutilised due to weak policy integration, inadequate valuation frameworks, and limited developer funding. To unlock this potential, urban planning should:
- Embed LWS into statutory planning policy with enforceable biodiversity standards and clear design guidelines [74].
- Integrate LWS into wider green infrastructure networks to enhance habitat connectivity and species movement.
- Adopt valuation frameworks that recognise functional and species-level contributions, such as Wallacea’s composite methodology [35].
- Prioritise native plantings, complemented by selected non-native evergreens for year-round cover and climate resilience.
- Mandate long-term monitoring and maintenance plans to ensure sustained ecological performance.
Our findings indicate that LWS can deliver meaningful biodiversity benefits when designed with ecological principles in mind. Soil-based systems, in particular, support more complex food webs, enhance microclimatic stability, and provide critical resources for microbial, detritivore, and higher trophic communities. Plant species composition is equally important, with structural and functional diversity strongly influencing above- and belowground invertebrate assemblages and, by extension, habitat quality for birds and bats.
Future research should address long-term biodiversity outcomes, the role of LWS in conserving priority taxa, and structural innovations that support bat roosting in urban environments. This study was limited by a single replicate per LWS design and short duration, constraining generalisation. However, it provides a framework for biodiversity assessment. Observations for birds and bats should be considered indicative. Future studies should include multiple replicates and extend temporal and spatial coverage to strengthen inference.
Overall, LWS represent a viable but underused tool for enhancing urban biodiversity. Their inclusion in planning policy, supported by robust valuation frameworks and long-term monitoring, will be key to ensuring urban development contributes positively to ecological resilience and human well-being.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/urbansci9120519/s1.
Author Contributions
Conceptualisation, P.H.L. and T.R.M. Methodology, P.H.L., T.R.M. and J.B. Software G.T.; Validation, P.H.L., T.R.M. and J.B.; Formal Analysis, P.H.L., T.R.M., G.T., E.C. and S.M.; Investigation, G.T., E.C. and S.M.; Resources, P.H.L., T.R.M. and J.B.; Data Curation, E.C., S.M. and G.T.; Writing—Original Draft Preparation, P.H.L. and T.R.M.; Writing—Review & Editing, P.H.L., T.R.M. and J.B.; visualisation, P.H.L. and G.T. Supervision, P.H.L., T.R.M. and J.B.; Project Administration, P.H.L. and T.R.M.; Funding Acquisition, P.H.L. All authors have read and agreed to the published version of the manuscript.
Funding
Funding was provided by the Sustainability Hub: Low Carbon Devon project as part of a European Regional Development award (Ref: 16R17P02120).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The original contributions presented in this study are included in the article/Supplementary Material. Further inquiries can be directed to the corresponding author.
Acknowledgments
We also thank the efforts of the following undergraduate students, Jennifer Poole and Matthew Philpott who contributed to data collection efforts, as well as Spencer Collins and Jane Akerman for assistance with species ID.
Conflicts of Interest
The authors declare no conflict of interest.
References
- United Nations. World Urbanization Prospects 2018; Department of Economics and Social Affairs—Population Dynamics: New York, NY, USA, 2019; Available online: https://population.un.org/wup/assets/Publications/WUP2018-Report.pdf/ (accessed on 26 November 2025).
- Seto, K.C.; Güneralp, B.; Hutyra, L.R. Global Forecasts of Urban Expansion to 2030 and Direct Impacts on Biodiversity and Carbon Pools. Proc. Natl. Acad. Sci. USA 2012, 109, 16083–16088. [Google Scholar] [CrossRef]
- Li, G.; Fang, C.; Li, Y. Global Impacts of Future Urban Expansion on Terrestrial Vertebrate Diversity. Nat. Commun. 2022, 13, 1628. [Google Scholar] [CrossRef]
- Ministry of Housing, Communities and Local Government. National Planning Policy Framework—Revised. December 2024. Available online: https://assets.publishing.service.gov.uk/media/67aafe8f3b41f783cca46251/NPPF_December_2024.pdf/ (accessed on 26 November 2025).
- Tzoulas, K.; Korpela, K.; Venn, S.; Yli-Pelkonen, V.; Kazmierczak, A.; Niemelä, J.; James, P. Promoting Ecosystem and Human Health in Urban Areas Using Green Infrastructure: A Literature Review. Landsc. Urban Plan. 2007, 81, 167–178. [Google Scholar] [CrossRef]
- Ottelé, M.; Perini, K.; Fraaij, A.L.A.; Haas, E.M.; Raiteri, R. Comparative Life Cycle Analysis for Green Façades and Living Wall Systems. Energy Build. 2011, 43, 3419–3429. [Google Scholar] [CrossRef]
- Francis, R.A.; Lorimer, J. Urban Reconciliation Ecology: The Potential of Living Roofs and Walls. J. Environ. Manag. 2011, 92, 1429–1437. [Google Scholar] [CrossRef] [PubMed]
- Sommese, F. Nature-Based Solutions to Enhance Urban Resilience in the Climate Change and Post-Pandemic Era: A Taxonomy for the Built Environment. Buildings 2024, 14, 2190. [Google Scholar] [CrossRef]
- Aronson, M.F.J.; La Sorte, F.A.; Nilon, C.H.; Katti, M.; Goddard, M.A.; Lepczyk, C.A.; Warren, P.S.; Williams, N.S.G.; Cilliers, S.; Clarkson, B.; et al. A Global Analysis of the Impacts of Urbanization on Bird and Plant Diversity Reveals Key Anthropogenic Drivers. Proc. R. Soc. B Biol. Sci. 2014, 281, 20133330. [Google Scholar] [CrossRef] [PubMed]
- Ives, C.D.; Lentini, P.E.; Threlfall, C.G.; Ikin, K.; Shanahan, D.F.; Garrard, G.E.; Bekessy, S.A.; Fuller, R.A.; Mumaw, L.; Rayner, L.; et al. Cities Are Hotspots for Threatened Species. Glob. Ecol. Biogeogr. 2016, 25, 117–126. [Google Scholar] [CrossRef]
- Perini, K.; Ottelé, M.; Fraaij, A.L.A.; Haas, E.M.; Raiteri, R. Vertical Greening Systems and the Effect on Air Flow and Temperature on the Building Envelope. Build. Environ. 2013, 46, 2287–2294. [Google Scholar] [CrossRef]
- Cameron, R.W.F.; Taylor, J.E.; Emmett, M.R. What’s “Cool” in the World of Green Façades? How Plant Choice Influences the Cooling Properties of Green Walls. Build. Environ. 2014, 73, 198–207. [Google Scholar] [CrossRef]
- Fox, M.; Morewood, J.; Murphy, T.; Lunt, P.; Goodhew, S. Living Wall Systems for Improved Thermal Performance of Existing Buildings. Build. Environ. 2022, 207, 108491. [Google Scholar] [CrossRef]
- Lunt, P.H.; Fuller, K.; Fox, M.; Goodhew, S.; Murphy, T.R. Comparing the Thermal Conductivity of Three Soil Types under Differing Moisture and Density Conditions for Use in Green Infrastructure. Soil Use Manag. 2022, 39, 260–269. [Google Scholar] [CrossRef]
- Köhler, M. Green Facades—A View Back and Some Visions. Urban Ecosyst. 2008, 11, 423–436. [Google Scholar] [CrossRef]
- Rondeau, A.; Mandon, A.; Malhautier, L.; Poly, F.; Richaume, A. Biopurification of Air Containing a Low Concentration of TEX: Comparison of Removal Efficiency Using Planted and Non-Planted Biofilters. J. Chem. Technol. Biotechnol. 2012, 87, 746–750. [Google Scholar] [CrossRef]
- Ismail, M.R. Quiet Environment: Acoustics of Vertical Green Wall Systems of the Islamic Urban Form. Front. Archit. Res. 2013, 2, 162–171. [Google Scholar] [CrossRef]
- Tiago, P.; Leal, A.I.; Matos Silva, C. Assessing Ecological Gains: A Review of How Arthropods, Bats and Birds Benefit from Green Roofs and Walls. Environments 2024, 11, 76. [Google Scholar] [CrossRef]
- Madre, F.; Clergeau, P.; Machon, N.; Vergnes, A. Building Biodiversity: Vegetated Façades as Habitats for Spider and Beetle Assemblages. Glob. Ecol. Conserv. 2015, 3, 222–233. [Google Scholar] [CrossRef]
- Fernández-Cañero, R.; Pérez Urrestarazu, L.; Perini, K. Vertical Greening Systems: Classifications, Plant Species, Substrates. In Nature Based Strategies for Urban and Building Sustainability; Pérez, G., Perini, K., Eds.; Butterworth-Heinemann: Oxford, UK, 2018; pp. 45–54. ISBN 9780128121504. [Google Scholar]
- Salisbury, A.; Blanusa, T.; Bostock, H.; Perry, J.N. Careful Plant Choice Can Deliver More Biodiverse Vertical Greening (Green Façades). Urban For. Urban Green. 2023, 89, 128118. [Google Scholar] [CrossRef]
- Dunnett, N.; Kingsbury, N. Planting Green Roofs and Living Walls; Timber Press: Portland, OR, USA, 2008; ISBN 978-0881929119. [Google Scholar]
- Weiler, S.K.; Scholz-Barth, K. Green Roof Systems: A Guide to the Planning, Design, and Construction of landscapes Over Structure; John Wiley & Sons: Hoboken, NJ, USA, 2009; ISBN 978-0471674955. [Google Scholar]
- Tartaglia, E.S.; Aronson, M.F.J. Plant Native: Comparing Biodiversity Benefits, Ecosystem Services Provisioning, and Plant Performance of Native and Non-Native Plants in Urban Horticulture. Urban Ecosyst. 2024, 27, 2587–2611. [Google Scholar] [CrossRef]
- Andrade, M.; Fernandes, C.; Figueiredo, A. Are Urban Green Spaces’ Attributes Relevant to Explain the Occurrence of Invasive Species Within Urban Green Infrastructure? Urban Sci. 2025, 9, 260. [Google Scholar] [CrossRef]
- Russo, A.; Esperon-Rodriguez, M.; St-Denis, A.; Tjoelker, M.G. Native vs. Non-Native Plants: Public Preferences, Ecosystem Services, and Conservation Strategies for Climate-Resilient Urban Green Spaces. Land 2025, 14, 954. [Google Scholar] [CrossRef]
- Bustami, R.A.; Belusko, M.; Ward, J.; Beecham, S. A Statistically Rigorous Approach to Experimental Design of Vertical Living Walls for Green Buildings. Urban Sci. 2019, 3, 71. [Google Scholar] [CrossRef]
- Uthappa, A.R.; Shishira, D.; Chavan, S.; Kumar, M. Soil Arthropods and Their Role in Soil Health Sustenance. Available online: https://www.researchgate.net/publication/362301891_Soil_arthropods_and_their_role_in_soil_health_sustenance (accessed on 13 August 2025).
- Lewandowski, D.; Cortina-Segarra, J.; Klein, T.; Früh, B. Bioreceptivity of Living Walls: Interactions between Building Materials and Biological Colonization. Build. Environ. 2023, 241, 110719. [Google Scholar] [CrossRef]
- Manso, M.; Castro-Gomes, J. Green Wall Systems: A Review of Their Characteristics. Renew. Sustain. Energy Rev. 2015, 41, 863–871. [Google Scholar] [CrossRef]
- Pérez-Urrestarazu, L.; Fernández-Cañero, R.; Franco-Salas, A.; Egea, G. Vertical Greening Systems and Sustainable Cities. J. Urban Technol. 2016, 23, 59–79. [Google Scholar] [CrossRef]
- Environment Act 2021. UK Public General Acts c. 30. 2021. Available online: https://www.legislation.gov.uk/ukpga/2021/30/part/6/crossheading/biodiversity-gain-in-planning/enacted (accessed on 8 August 2025).
- Natural England. The Biodiversity Metric 4.0 User Guide; Natural England Joint Publication JP039; Natural England. March 2023. Available online: https://www.researchgate.net/publication/373453757_The_Biodiversity_Metric_40_User_Guide?channel=doi&linkId=64eda1adc4f9ac00607473d2&showFulltext=true (accessed on 26 November 2025).
- Defra. Understanding Biodiversity Net Gain: Guidance on What BNG Is and How It Affects Land Managers, Developers and Local Planning Authorities: GOV.UK. 2025. Available online: https://www.gov.uk/guidance/understanding-biodiversity-net-gain (accessed on 26 November 2025).
- Wallacea Trust. Species-Level Biodiversity Assessment Methodology. Available online: https://wallaceatrust.org (accessed on 7 August 2025).
- Foo, J.; O’Dea, R.E.; Koricheva, J.; Nakagawa, S.; Lagisz, M. A Practical Guide to Question Formation, Systematic Searching and Study Screening for Literature Reviews in Ecology and Evolution. Methods Ecol. Evol. 2021, 12, 1705–1720. [Google Scholar] [CrossRef]
- Shamseer, L.; Moher, D.; Clarke, M.; Ghersi, D.; Liberati, A.; Petticrew, M.; Shekelle, P.; Stewart, L.A.; PRISMA-P Group. Preferred Reporting Items for Systematic Review and Meta-Analysis Protocols (PRISMA-P) 2015: Elaboration and Explanation. BMJ 2015, 349, g7647. [Google Scholar] [CrossRef]
- Murphy, T.R.; Hanley, M.E.; Ellis, J.S.; Lunt, P.H. Deviation between Projected and Observed Precipitation Trends Greater with Altitude. Clim. Res. 2019, 79, 77–89. [Google Scholar] [CrossRef]
- Scotscape. Living Wall Systems: Modular Green Infrastructure for Urban Environments. Available online: https://www.scotscape.co.uk/our-products/living-walls (accessed on 20 August 2025).
- Biotecture Ltd. Why Hydroponics? Available online: https://www.biotecture.uk.com/design-and-specify/why-hydroponics/ (accessed on 7 August 2025).
- Pinho, P.; Correia, O.; Lecoq, M.; Munzi, S.; Vasconcelos, S.; Gonçalves, P.; Rebelo, R.; Antunes, C.; Silva, P.; Freitas, C.; et al. Evaluating Green Infrastructure in Urban Environments Using a Multi-Taxa and Functional Diversity Approach. Environ. Res. 2016, 147, 601–610. [Google Scholar] [CrossRef]
- Arizaga, J.; Garin-Barrio, I.; Etxeberria, M.; García del Bao, A.; Laso, M. The Value of Multi-Taxon Approaches to Improve Biodiversity Assessment at Municipal Scales. Anim. Biodivers. Conserv. 2024, 47, 161–170. [Google Scholar] [CrossRef]
- Rooney, T.; Roberson, E. Deer Herbivory Reduces Web-Building Spider Abundance by Simplifying Forest Vegetation Structure. PeerJ 2016, 4, e2538. [Google Scholar] [CrossRef]
- Natural History Museum of Bern. World Spider Catalog. Available online: https://wsc.nmbe.ch/statistics/ (accessed on 7 April 2022).
- Cornell Lab of Ornithology. Merlin Bird ID App. Available online: https://merlin.allaboutbirds.org (accessed on 20 August 2025).
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2025; Available online: https://www.R-project.org/ (accessed on 20 August 2025).
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016; Available online: https://ggplot2.tidyverse.org (accessed on 26 November 2025).
- Fox, J.; Weisberg, S. An R Companion to Applied Regression; Sage Publications: Thousand Oaks, CA, USA, 2019; ISBN 9781544336473. [Google Scholar]
- Fuller, R.A.; Irvine, K.N.; Devine-Wright, P.; Warren, P.H.; Gaston, K.J. Psychological Benefits of Green Space Increase with Biodiversity. Biol. Lett. 2007, 3, 390–394. [Google Scholar] [CrossRef]
- Collins, R.; Schaafsma, M.; Hudson, M.D. The Value of Green Walls to Urban Biodiversity. Land Use Policy 2017, 64, 114–123. [Google Scholar] [CrossRef]
- Goel, M.; Jha, B.; Khan, S. Living Walls Enhancing the Urban Realm: A Review. Environ. Sci. Pollut. Res. 2022, 29, 38715–38734. [Google Scholar] [CrossRef]
- Mayrand, F.; Clergeau, P. Green Roofs and Green Walls for Biodiversity Conservation: A Contribution to Urban Connectivity? Sustainability 2018, 10, 985. [Google Scholar] [CrossRef]
- Hecht, K.; Haan, L.; Wösten, H.A.B.; Hamel, P.; Swaminathan, S.; Jain, A. Rocks and Walls: Biodiversity and Temperature Regulation of Natural Cliffs and Vertical Greenery Systems. Build. Environ. 2025, 268, 112308. [Google Scholar] [CrossRef]
- Glen, A.S.; Pech, R.P.; Byrom, A.E. Connectivity and Invasive Species Management: Towards an Integrated Landscape Approach. Biol. Invasions 2013, 15, 2127–2138. [Google Scholar] [CrossRef]
- Boscutti, F.; Lami, F.; Pellegrini, E.; Buccheri, M.; Busato, F.; Martini, F.; Sibella, R.; Sigura, M.; Marini, L. Urban Sprawl Facilitates Invasions of Exotic Plants across Multiple Spatial Scales. Biol. Invasions 2022, 24, 1497–1510. [Google Scholar] [CrossRef]
- Gaertner, M.; Larson, B.M.H.; Irlich, U.M.; Holmes, P.M.; Stafford, L.; Richardson, D.M. Managing Invasive Species in Cities: A Framework from Cape Town, South Africa. Landsc. Urban Plan. 2016, 151, 1–9. [Google Scholar] [CrossRef]
- Partridge, D.R.; Clark, J.A. Urban Green Roofs Provide Habitat for Migrating and Breeding Birds and Their Arthropod Prey. PLoS ONE 2018, 13, e0202298. [Google Scholar] [CrossRef] [PubMed]
- Benedict, M.A.; McMahon, E.T. Green Infrastructure: Linking Landscapes and Communities; Island Press: Washington, DC, USA, 2006; ISBN 9781559635585. [Google Scholar]
- Belcher, R.N.; Fornasari, L.; Menz, S.; Schroepfer, T. Birds’ Use of Vegetated and Non-Vegetated High-Density Buildings—A Case Study of Milan. J. Urban Ecol. 2018, 4, juy001. [Google Scholar] [CrossRef]
- Tonietto, R.; Fant, J.; Ascher, J.; Ellis, K.; Larkin, D. A Comparison of Bee Communities of Chicago Green Roofs, Parks and Prairies. Landsc. Urban Plan. 2011, 103, 102–108. [Google Scholar] [CrossRef]
- Braaker, S.; Ghazoul, J.; Obrist, M.K.; Moretti, M. Habitat Connectivity and Local Conditions Shape Taxonomic and Functional Diversity of Arthropods on Green Roofs. J. Anim. Ecol. 2017, 86, 521–531. [Google Scholar] [CrossRef]
- Salisbury, A.; Armitage, J.; Bostock, H.; Perry, J.; Tatchell, M.; Thompson, K. Enhancing Gardens as Habitats for Plant-Associated Invertebrates: Should We Plant Native or Exotic Species? Biodivers. Conserv. 2017, 26, 2657–2673. [Google Scholar] [CrossRef]
- Law, C.M.Y.; Wong, N.H.; Tan, C.L.; Lim, H.S.; Wong, D.K.W. Data-Driven Approach for Optimising Plant Species Selection and Planting Design on Outdoor Modular Green Walls with Aesthetic, Maintenance, and Water-Saving Goals. Sustainability 2025, 17, 3528. [Google Scholar] [CrossRef]
- Siemann, E.; Tilman, D.; Haarstad, J. Experimental Tests of the Impacts of Plant Diversity on Community Structure and Ecosystem Functioning. Ecol. Monogr. 1998, 68, 427–442. [Google Scholar] [CrossRef]
- Voigt, A.; Kabisch, N.; Wurster, D.; Haase, D.; Breuste, J. Structural Diversity: A Multi-Dimensional Approach to Assess Recreational Services in Urban Parks. Ambio 2014, 43, 480–491. [Google Scholar] [CrossRef]
- Droz, A.G.; Koyama, A.; Zhang, J.; Stephan, R.; Schindlbacher, A.; Rousk, J.; Frey, S.D. Drivers of Fungal Diversity and Community Biogeography Differ in Green Roofs vs. Ground Soils. Environ. Microbiol. 2022, 24, 5781–5797. [Google Scholar] [CrossRef]
- Wang, X.; Chi, Y.; Song, S. Important Soil Microbiota’s Effects on Plants and Soils: A Comprehensive 30-Year Systematic Literature Review. Front. Microbiol. 2024, 15, 1347745. [Google Scholar] [CrossRef]
- Bray, N.; Wickings, K. The Roles of Invertebrates in the Urban Soil Microbiome. Front. Ecol. Evol. 2019, 7, 359. [Google Scholar] [CrossRef]
- Chiquet, C.; Dover, J.W.; Mitchell, P. Birds and the Urban Environment: The Value of Green Walls. Urban Ecosyst. 2013, 16, 453–462. [Google Scholar] [CrossRef]
- Fulthorpe, R.; MacIvor, J.S.; Jia, P.; Yasui, S.-L.E. The Green Roof Microbiome: Improving Plant Survival for Urban Roof Ecosystems. Front. Ecol. Evol. 2018, 6, 5. [Google Scholar] [CrossRef]
- Viritopia. Living Walls and Biodiversity: How Vertical Gardens Support Urban Wildlife. Available online: https://www.viritopia.com/living-wall (accessed on 8 August 2025).
- Delgado-Baquerizo, M.; Reich, P.B.; Trivedi, C.; Eldridge, D.J.; Abades, S.; Alfaro, F.D.; Bastida, F.; Berhe, A.A.; Cutler, N.A.; Gallardo, A.; et al. Multiple Elements of Soil Biodiversity Drive Ecosystem Functions across Biomes. Nat. Ecol. Evol. 2020, 4, 210–220. [Google Scholar] [CrossRef] [PubMed]
- Natural England. Biodiversity Net Gain: How to Use the Biodiversity Metric. GOV.UK. Available online: https://www.gov.uk/guidance/biodiversity-net-gain (accessed on 19 August 2025).
- Rampling, E.E.; Bull, J.W.; zu Ermgassen, S.O.S.E.; Hawkins, I. Achieving Biodiversity Net Gain by Addressing Governance Gaps Underpinning Ecological Compensation Policies. Conserv. Biol. 2024, 38, e14198. [Google Scholar] [CrossRef] [PubMed]
- Wildlife and Countryside Act 1981. UK Public General Acts c. 69. 1981. Available online: https://www.legislation.gov.uk/ukpga/1981/69 (accessed on 8 August 2025).
- Lehrer, E.W.; Gallo, T.; Fidino, M.; Kilgour, R.J.; Wolff, P.J.; Magle, S.B. Urban Bat Occupancy Is Highly Influenced by Noise and the Location of Water: Considerations for Nature-Based Urban Planning. Landsc. Urban Plan. 2021, 210, 104063. [Google Scholar] [CrossRef]
- Microsoft Research. Accelerating Biodiversity Surveys with AI. Available online: https://www.microsoft.com/en-us/research/project/accelerating-biodiversity-surveys/ (accessed on 8 August 2025).
- Larcher, F.; Battisti, L.; Bianco, L.; Giordano, R.; Montacchini, E.; Serra, V.; Tedesco, S. Sustainability of Living Wall Systems Through an Ecosystem Services Lens. In Urban Horticulture; Nandwani, D., Ed.; Springer: Cham, Switzerland, 2018; pp. 31–51. ISBN 978-331-967-017-1. [Google Scholar]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).