
Exploring the Internal Defense System of Cerastoderma glaucum (Bruguière, 1789) Exposed to Pristine Microplastics: The Sentinel Role of Haemocytes as Biomarkers

 ,
,  ,
, 
 ,
,  , ,
, ,  , , and
, , and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Contaminants
2.2. Selection of Concentrations
2.3. Animal Preparation
2.4. Experimental Design
2.5. Tissue Preparation
2.6. Histological Analysis
2.7. Immunofluorescence
2.8. Confocal Microscopy
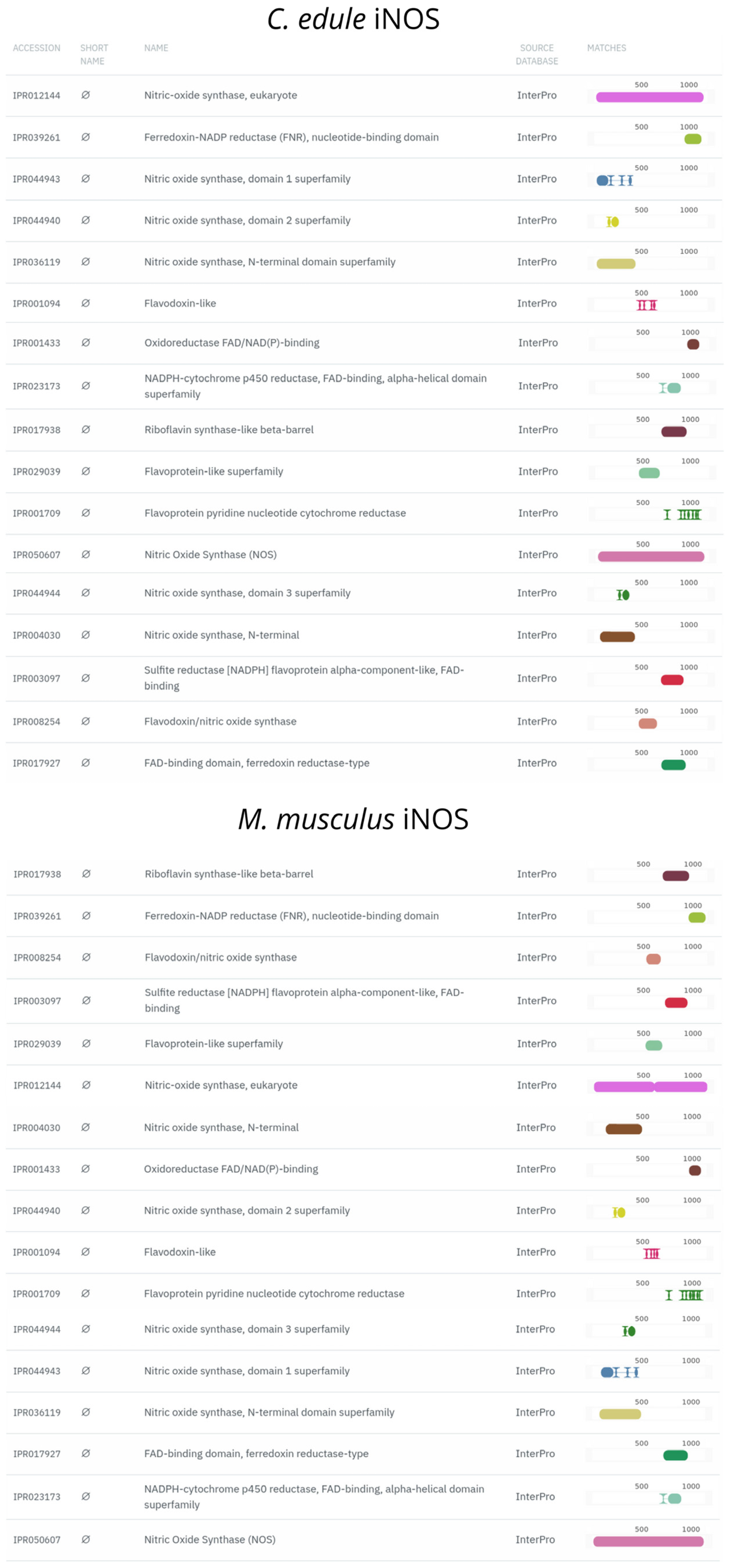
2.9. In Silico Prediction of TLR2 and iNOS Orthologs in the Genome of the Close Relative Cerastoderma edule (Linnaeus, 1758)
2.10. Statistical Analysis
3. Results
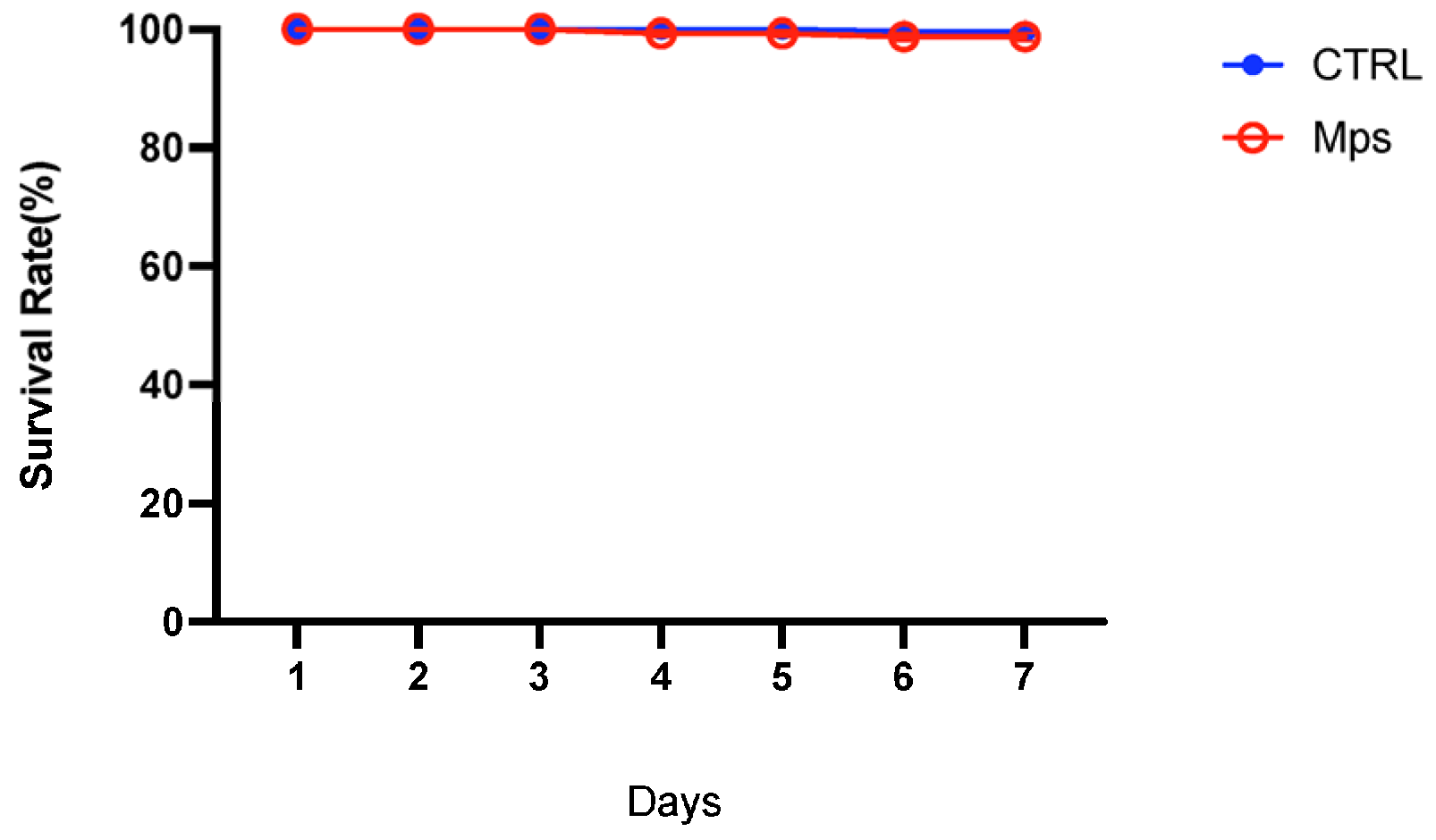
3.1. Mortality
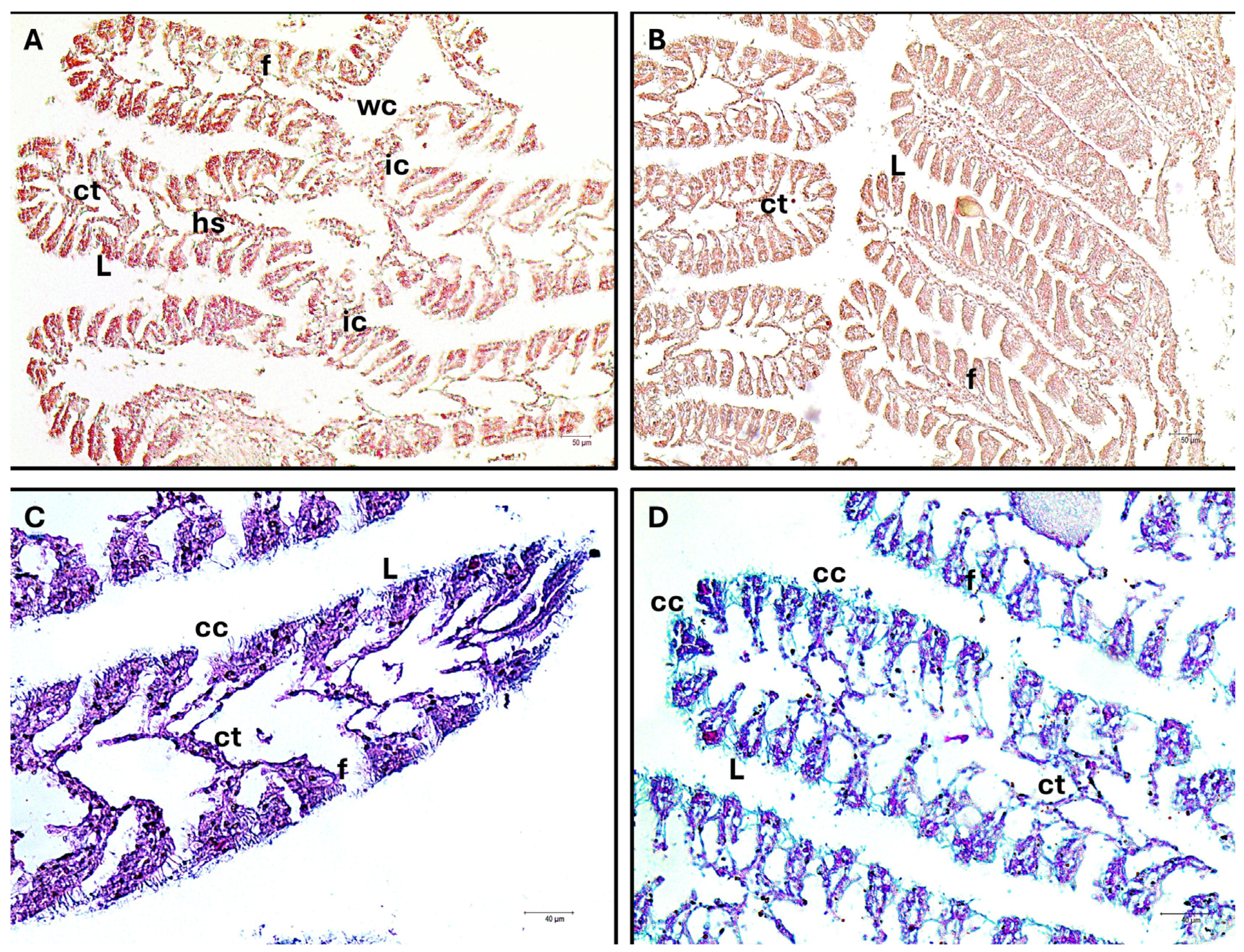
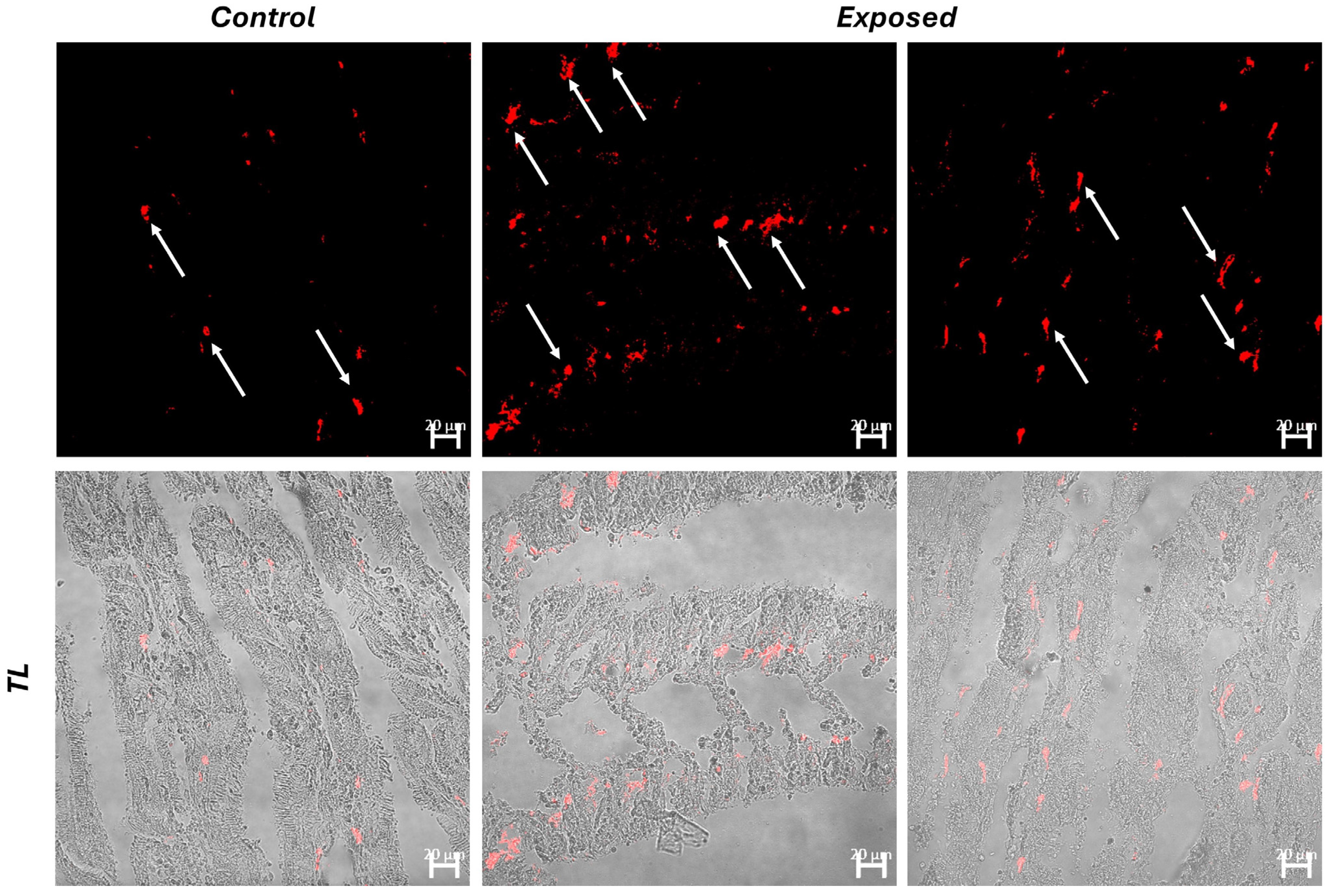
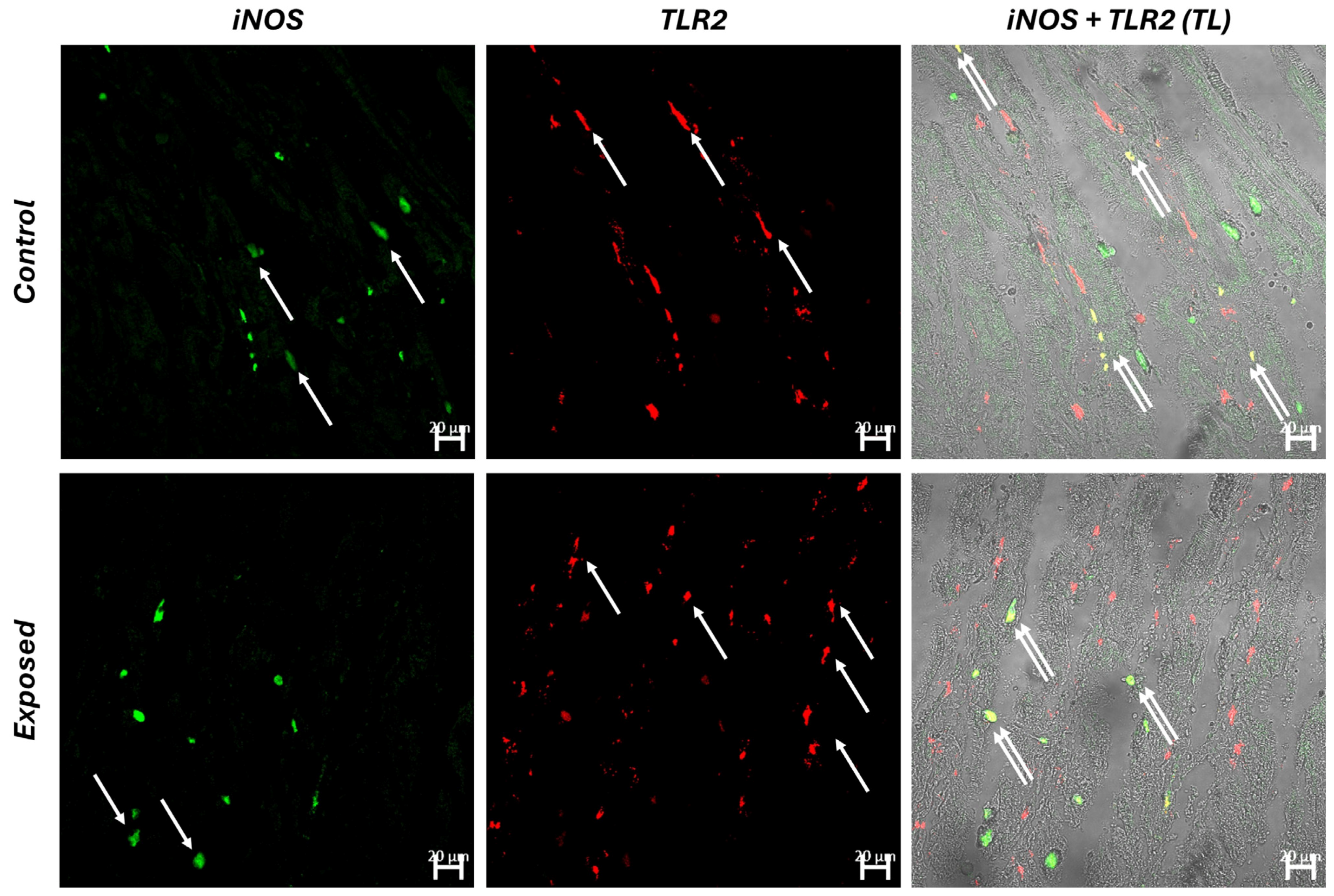
3.2. Histology and Immunofluorescence
3.3. TLR2 and iNOS Orthologs Are Found in the Cerastoderma edule Genome
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bertocci, I.; Dell’Anno, A.; Musco, L.; Gambi, C.; Saggiomo, V.; Cannavacciuolo, M.; Lo Martire, M.; Passarelli, A.; Zazo, G.; Danovaro, R. Multiple Human Pressures in Coastal Habitats: Variation of Meiofaunal Assemblages Associated with Sewage Discharge in a Post-Industrial Area. Sci. Total Environ. 2019, 655, 1218–1231. [Google Scholar] [CrossRef]
- Sharifinia, M.; Bahmanbeigloo, Z.A.; Keshavarzifard, M.; Khanjani, M.H.; Lyons, B.P. Microplastic Pollution as a Grand Challenge in Marine Research: A Closer Look at Their Adverse Impacts on the Immune and Reproductive Systems. Ecotoxicol. Environ. Saf. 2020, 204, 111109. [Google Scholar] [CrossRef] [PubMed]
- Larsen, A.K.; Holen, E.; Segner, H. Editorial: Immunity and Disease of Aquatic Organisms under the Combined Impact of Anthropogenic Stressors: Mechanisms and Disease Outcomes. Front. Mar. Sci. 2023, 10, 1291639. [Google Scholar] [CrossRef]
- Bijma, J.; Pörtner, H.-O.; Yesson, C.; Rogers, A.D. Climate Change and the Oceans—What Does the Future Hold? Mar. Pollut. Bull. 2013, 74, 495–505. [Google Scholar] [CrossRef] [PubMed]
- Crain, C.M.; Kroeker, K.; Halpern, B.S. Interactive and Cumulative Effects of Multiple Human Stressors in Marine Systems. Ecol. Lett. 2008, 11, 1304–1315. [Google Scholar] [CrossRef] [PubMed]
- Segner, H.; Schmitt-Jansen, M.; Sabater, S. Assessing the Impact of Multiple Stressors on Aquatic Biota: The Receptor’s Side Matters. Environ. Sci. Technol. 2014, 48, 7690–7696. [Google Scholar] [CrossRef] [PubMed]
- Byers, J.E. Marine Parasites and Disease in the Era of Global Climate Change. Annu. Rev. Mar. Sci. 2021, 13, 397–420. [Google Scholar] [CrossRef] [PubMed]
- Hutson, K.S.; Davidson, I.C.; Bennett, J.; Poulin, R.; Cahill, P.L. Assigning Cause for Emerging Diseases of Aquatic Organisms. Trends Microbiol. 2023, 31, 681–691. [Google Scholar] [CrossRef] [PubMed]
- De Souza Machado, A.A.; Kloas, W.; Zarfl, C.; Hempel, S.; Rillig, M.C. Microplastics as an Emerging Threat to Terrestrial Ecosystems. Glob. Change Biol. 2018, 24, 1405–1416. [Google Scholar] [CrossRef]
- Sutherland, W.J.; Broad, S.; Caine, J.; Clout, M.; Dicks, L.V.; Doran, H.; Entwistle, A.C.; Fleishman, E.; Gibbons, D.W.; Keim, B.; et al. A Horizon Scan of Global Conservation Issues for 2016. Trends Ecol. Evol. 2016, 31, 44–53. [Google Scholar] [CrossRef]
- Liu, M.; Liu, J.; Xiong, F.; Xu, K.; Pu, Y.; Huang, J.; Zhang, J.; Pu, Y.; Sun, R.; Cheng, K. Research Advances of Microplastics and Potential Health Risks of Microplastics on Terrestrial Higher Mammals: A Bibliometric Analysis and Literature Review. Env. Geochem. Health 2023, 45, 2803–2838. [Google Scholar] [CrossRef] [PubMed]
- Lyons, B.P.; Cowie, W.J.; Maes, T.; Le Quesne, W.J.F. Marine Plastic Litter in the ROPME Sea Area: Current Knowledge and Recommendations. Ecotoxicol. Environ. Saf. 2020, 187, 109839. [Google Scholar] [CrossRef] [PubMed]
- Galloway, T.S.; Cole, M.; Lewis, C. Interactions of Microplastic Debris throughout the Marine Ecosystem. Nat. Ecol. Evol. 2017, 1, 0116. [Google Scholar] [CrossRef] [PubMed]
- Porter, A.; Lyons, B.P.; Galloway, T.S.; Lewis, C. Role of Marine Snows in Microplastic Fate and Bioavailability. Environ. Sci. Technol. 2018, 52, 7111–7119. [Google Scholar] [CrossRef] [PubMed]
- Su, L.; Deng, H.; Li, B.; Chen, Q.; Pettigrove, V.; Wu, C.; Shi, H. The Occurrence of Microplastic in Specific Organs in Commercially Caught Fishes from Coast and Estuary Area of East China. J. Hazard. Mater. 2019, 365, 716–724. [Google Scholar] [CrossRef] [PubMed]
- Kataoka, C.; Kato, Y.; Ariyoshi, T.; Takasu, M.; Narazaki, T.; Nagasaka, S.; Tatsuta, H.; Kashiwada, S. Comparative Toxicities of Silver Nitrate, Silver Nanocolloids, and Silver Chloro-Complexes to Japanese Medaka Embryos, and Later Effects on Population Growth Rate. Environ. Pollut. 2018, 233, 1155–1163. [Google Scholar] [CrossRef] [PubMed]
- Kataoka, C.; Kashiwada, S. Ecological Risks Due to Immunotoxicological Effects on Aquatic Organisms. Int. J. Mol. Sci. 2021, 22, 8305. [Google Scholar] [CrossRef] [PubMed]
- Sarma, H.; Hazarika, R.P.; Kumar, V.; Roy, A.; Pandit, S.; Prasad, R. Microplastics in Marine and Aquatic Habitats: Sources, Impact, and Sustainable Remediation Approaches. Environ. Sustain. 2022, 5, 39–49. [Google Scholar] [CrossRef]
- Fauser, P.; Vorkamp, K.; Strand, J. Residual Additives in Marine Microplastics and Their Risk Assessment—A Critical Review. Mar. Pollut. Bull. 2022, 177, 113467. [Google Scholar] [CrossRef]
- Eerkes-Medrano, D.; Thompson, R.C.; Aldridge, D.C. Microplastics in Freshwater Systems: A Review of the Emerging Threats, Identification of Knowledge Gaps and Prioritisation of Research Needs. Water Res. 2015, 75, 63–82. [Google Scholar] [CrossRef]
- Paul-Pont, I.; Lacroix, C.; González Fernández, C.; Hégaret, H.; Lambert, C.; Le Goïc, N.; Frère, L.; Cassone, A.-L.; Sussarellu, R.; Fabioux, C.; et al. Exposure of Marine Mussels Mytilus Spp. to Polystyrene Microplastics: Toxicity and Influence on Fluoranthene Bioaccumulation. Environ. Pollut. 2016, 216, 724–737. [Google Scholar] [CrossRef] [PubMed]
- Duncan, E.M.; Broderick, A.C.; Fuller, W.J.; Galloway, T.S.; Godfrey, M.H.; Hamann, M.; Limpus, C.J.; Lindeque, P.K.; Mayes, A.G.; Omeyer, L.C.M.; et al. Microplastic Ingestion Ubiquitous in Marine Turtles. Glob. Change Biol. 2019, 25, 744–752. [Google Scholar] [CrossRef] [PubMed]
- Raju, P.; Gunabal, S.; Santhanam, P. The Impact of Microplastics on Marine Copepods. In Basic and Applied Zooplankton Biology; Santhanam, P., Begum, A., Pachiappan, P., Eds.; Springer: Singapore, 2019; pp. 429–442. ISBN 978-981-10-7952-8. [Google Scholar]
- Ivar Do Sul, J.A.; Costa, M.F. The Present and Future of Microplastic Pollution in the Marine Environment. Environ. Pollut. 2014, 185, 352–364. [Google Scholar] [CrossRef]
- Xu, S.; Ma, J.; Ji, R.; Pan, K.; Miao, A.-J. Microplastics in Aquatic Environments: Occurrence, Accumulation, and Biological Effects. Sci. Total Environ. 2020, 703, 134699. [Google Scholar] [CrossRef]
- Mirzoeva, A.T.; Demchenko, N.A. Morphological Response of Lagoon Cockle Cerastoderma glaucum (Poiret, 1789) to Eutrophication in the Sea of Azov. IOP Conf. Ser. Earth Environ. Sci. 2022, 1049, 012059. [Google Scholar] [CrossRef]
- Velez, C.; Pires, A.; Sampaio, L.; Cardoso, P.; Moreira, A.; Leandro, S.; Figueira, E.; Soares, A.M.V.M.; Freitas, R. The Use of Cerastoderma glaucum as a Sentinel and Bioindicator Species: Take-Home Message. Ecol. Indic. 2016, 62, 228–241. [Google Scholar] [CrossRef]
- Karray, S.; Tastard, E.; Moreau, B.; Delahaut, L.; Geffard, A.; Guillon, E.; Denis, F.; Hamza-Chaffai, A.; Chénais, B.; Marchand, J. Transcriptional Response of Stress-Regulated Genes to Industrial Effluent Exposure in the Cockle Cerastoderma glaucum. Environ. Sci. Pollut. Res. 2015, 22, 17303–17316. [Google Scholar] [CrossRef]
- Machreki-Ajmi, M.; Hamza-Chaffai, A. Assessment of Sediment/Water Contamination by in Vivo Transplantation of the Cockles Cerastoderma glaucum from a Non Contaminated to a Contaminated Area by Cadmium. Ecotoxicology 2008, 17, 802–810. [Google Scholar] [CrossRef] [PubMed]
- González-Fernández, C.; Albentosa, M.; Campillo, J.A.; Viñas, L.; Fumega, J.; Franco, A.; Besada, V.; González-Quijano, A.; Bellas, J. Influence of Mussel Biological Variability on Pollution Biomarkers. Environ. Res. 2015, 137, 14–31. [Google Scholar] [CrossRef]
- Paul-Pont, I.; Gonzalez, P.; Baudrimont, M.; Nili, H.; De Montaudouin, X. Short-Term Metallothionein Inductions in the Edible Cockle Cerastoderma Edule after Cadmium or Mercury Exposure: Discrepancy between mRNA and Protein Responses. Aquat. Toxicol. 2010, 97, 260–267. [Google Scholar] [CrossRef]
- Hamza-Chaffai, A. Usefulness of Bioindicators and Biomarkers in Pollution Biomonitoring. Int. J. Biotech. Well. Indus. 2014, 3, 19–26. [Google Scholar] [CrossRef]
- Kandeel, K.E.; Mohammed, S.Z.; Mostafa, A.M.; Abd-Alla, M.E. Reproductive Biology of the Cockle Cerastoderma glaucum (Bivalvia: Cardiidae) from Lake Qarun, Egypt. Egypt. J. Aquat. Res. 2013, 39, 249–260. [Google Scholar] [CrossRef]
- WoRMS—World Register of Marine Species—Cerastoderma Glaucum (Bruguière, 1789). Available online: https://www.marinespecies.org/aphia.php?p=taxdetails&id=138999 (accessed on 15 March 2024).
- Rollins-Smith, L.A. Amphibian Immunity–Stress, Disease, and Climate Change. Dev. Comp. Immunol. 2017, 66, 111–119. [Google Scholar] [CrossRef]
- Palmer, C.V. Immunity and the Coral Crisis. Commun. Biol. 2018, 1, 91. [Google Scholar] [CrossRef]
- Canesi, L.; Ciacci, C.; Balbi, T. Invertebrate Models for Investigating the Impact of Nanomaterials on Innate Immunity: The Example of the Marine Mussel Mytilus Spp. Curr. Bionanotechnol. 2017, 2, 77–83. [Google Scholar] [CrossRef]
- Alesci, A.; Capillo, G.; Fumia, A.; Albano, M.; Messina, E.; Spanò, N.; Pergolizzi, S.; Lauriano, E.R. Coelomocytes of the Oligochaeta Earthworm Lumbricus terrestris (Linnaeus, 1758) as Evolutionary Key of Defense: A Morphological Study. Zool. Lett. 2023, 9, 5. [Google Scholar] [CrossRef] [PubMed]
- Alesci, A.; Fumia, A.; Albano, M.; Messina, E.; D’Angelo, R.; Mangano, A.; Miller, A.; Spanò, N.; Savoca, S.; Capillo, G. Investigating the Internal System of Defense of Gastropoda Aplysia depilans (Gmelin, 1791): Focus on Hemocytes. Fish Shellfish Immunol. 2023, 137, 108791. [Google Scholar] [CrossRef]
- Espinosa, C.; Cuesta, A.; Esteban, M.Á. Effects of Dietary Polyvinylchloride Microparticles on General Health, Immune Status and Expression of Several Genes Related to Stress in Gilthead Seabream (Sparus aurata L.). Fish. Shellfish. Immunol. 2017, 68, 251–259. [Google Scholar] [CrossRef]
- Espinosa, C.; García Beltrán, J.M.; Esteban, M.A.; Cuesta, A. In Vitro Effects of Virgin Microplastics on Fish Head-Kidney Leucocyte Activities. Environ. Pollut. 2018, 235, 30–38. [Google Scholar] [CrossRef]
- Marques-Santos, L.F.; Grassi, G.; Bergami, E.; Faleri, C.; Balbi, T.; Salis, A.; Damonte, G.; Canesi, L.; Corsi, I. Cationic Polystyrene Nanoparticle and the Sea Urchin Immune System: Biocorona Formation, Cell Toxicity, and Multixenobiotic Resistance Phenotype. Nanotoxicology 2018, 12, 847–867. [Google Scholar] [CrossRef]
- Tang, J.; Ni, X.; Zhou, Z.; Wang, L.; Lin, S. Acute Microplastic Exposure Raises Stress Response and Suppresses Detoxification and Immune Capacities in the Scleractinian Coral Pocillopora damicornis. Environ. Pollut. 2018, 243, 66–74. [Google Scholar] [CrossRef] [PubMed]
- Gustafson, H.H.; Holt-Casper, D.; Grainger, D.W.; Ghandehari, H. Nanoparticle Uptake: The Phagocyte Problem. Nano Today 2015, 10, 487–510. [Google Scholar] [CrossRef] [PubMed]
- Alesci, A.; Di Paola, D.; Fumia, A.; Marino, S.; D’Iglio, C.; Famulari, S.; Albano, M.; Spanò, N.; Lauriano, E.R. Internal Defense System of Mytilus galloprovincialis (Lamarck, 1819): Ecological Role of Hemocytes as Biomarkers for Thiacloprid and Benzo[a]Pyrene Pollution. Toxics 2023, 11, 731. [Google Scholar] [CrossRef] [PubMed]
- Alesci, A.; Cicero, N.; Fumia, A.; Petrarca, C.; Mangifesta, R.; Nava, V.; Lo Cascio, P.; Gangemi, S.; Di Gioacchino, M.; Lauriano, E.R. Histological and Chemical Analysis of Heavy Metals in Kidney and Gills of Boops Boops: Melanomacrophages Centers and Rodlet Cells as Environmental Biomarkers. Toxics 2022, 10, 218. [Google Scholar] [CrossRef] [PubMed]
- Zelikoff, J.T.; Raymond, A.; Carlson, E.; Li, Y.; Beaman, J.R.; Anderson, M. Biomarkers of Immunotoxicity in Fish: From the Lab to the Ocean. Toxicol. Lett. 2000, 112–113, 325–331. [Google Scholar] [CrossRef] [PubMed]
- Torrealba, D.; More-Bayona, J.A.; Wakaruk, J.; Barreda, D.R. Innate Immunity Provides Biomarkers of Health for Teleosts Exposed to Nanoparticles. Front. Immunol. 2019, 9, 3074. [Google Scholar] [CrossRef]
- Hook, S.E.; Gallagher, E.P.; Batley, G.E. The Role of Biomarkers in the Assessment of Aquatic Ecosystem Health. Int. Environ. Assess. Manag. 2014, 10, 327–341. [Google Scholar] [CrossRef] [PubMed]
- Dubaish, F.; Liebezeit, G. Suspended Microplastics and Black Carbon Particles in the Jade System, Southern North Sea. Water Air Soil. Pollut. 2013, 224, 1352. [Google Scholar] [CrossRef]
- Albano, M.; Panarello, G.; Di Paola, D.; Capparucci, F.; Crupi, R.; Gugliandolo, E.; Spanò, N.; Capillo, G.; Savoca, S. The Influence of Polystyrene Microspheres Abundance on Development and Feeding Behavior of Artemia salina (Linnaeus, 1758). Appl. Sci. 2021, 11, 3352. [Google Scholar] [CrossRef]
- Sharma, S.; Sharma, V.; Chatterjee, S. Microplastics in the Mediterranean Sea: Sources, Pollution Intensity, Sea Health, and Regulatory Policies. Front. Mar. Sci. 2021, 8, 634934. [Google Scholar] [CrossRef]
- Lauriano, E.R.; Żuwała, K.; Kuciel, M.; Budzik, K.A.; Capillo, G.; Alesci, A.; Pergolizzi, S.; Dugo, G.; Zaccone, G. Confocal Immunohistochemistry of the Dermal Glands and Evolutionary Considerations in the Caecilian, Typhlonectes natans (Amphibia: Gymnophiona). Acta Zool. 2016, 97, 154–164. [Google Scholar] [CrossRef]
- Hoff, K.J.; Lange, S.; Lomsadze, A.; Borodovsky, M.; Stanke, M. BRAKER1: Unsupervised RNA-Seq-Based Genome Annotation with GeneMark-ET and AUGUSTUS. Bioinformatics 2016, 32, 767–769. [Google Scholar] [CrossRef] [PubMed]
- Bruna, T.; Hoff, K.J.; Lomsadze, A.; Stanke, M.; Borodovsky, M. BRAKER2: Automatic Eukaryotic Genome Annotation with GeneMark-EP+ and AUGUSTUS Supported by a Protein Database. NAR Genom. Bioinform. 2021, 3, lqaa108. [Google Scholar]
- Hoff, K.J.; Lomsadze, A.; Borodovsky, M.; Stanke, M. Whole-Genome Annotation with BRAKER. In Gene Prediction; Kollmar, M., Ed.; Methods in Molecular Biology; Springer: New York, NY, USA, 2019; Volume 1962, pp. 65–95. ISBN 978-1-4939-9172-3. [Google Scholar]
- Buchfink, B.; Xie, C.; Huson, D.H. Fast and Sensitive Protein Alignment Using DIAMOND. Nat. Methods 2015, 12, 59–60. [Google Scholar] [CrossRef]
- Stanke, M.; Diekhans, M.; Baertsch, R.; Haussler, D. Using Native and Syntenically Mapped cDNA Alignments to Improve de Novo Gene Finding. Bioinformatics 2008, 24, 637–644. [Google Scholar] [CrossRef]
- Stanke, M.; Schöffmann, O.; Morgenstern, B.; Waack, S. Gene Prediction in Eukaryotes with a Generalized Hidden Markov Model That Uses Hints from External Sources. BMC Bioinform. 2006, 7, 62. [Google Scholar] [CrossRef] [PubMed]
- Dainat, J.; Hereñú, D.; Murray, K.D.; Davis, E.; Crouch, K.; LucileSol; Agostinho, N.; Pascal-Git; Zollman, Z. Tayyrov NBISweden/AGAT: AGAT-v1.2.0. 2023. Available online: https://zenodo.org/records/8178877 (accessed on 7 March 2024).
- Emms, D.M.; Kelly, S. OrthoFinder: Phylogenetic Orthology Inference for Comparative Genomics. Genome Biol. 2019, 20, 238. [Google Scholar] [CrossRef] [PubMed]
- Paysan-Lafosse, T.; Blum, M.; Chuguransky, S.; Grego, T.; Pinto, B.L.; Salazar, G.A.; Bileschi, M.L.; Bork, P.; Bridge, A.; Colwell, L. InterPro in 2022. Nucleic Acids Res. 2023, 51, D418–D427. [Google Scholar] [CrossRef]
- Dobal, V.; Suárez, P.; Ruiz, Y.; García-Martín, O.; San Juan, F. Activity of Antioxidant Enzymes in Mytilus galloprovincialis Exposed to Tar: Integrated Response of Different Organs as Pollution Biomarker in Aquaculture Areas. Aquaculture 2022, 548, 737638. [Google Scholar] [CrossRef]
- Urban-Malinga, B.; Jakubowska, M.; Białowąs, M. Response of Sediment-Dwelling Bivalves to Microplastics and Its Potential Implications for Benthic Processes. Sci. Total Environ. 2021, 769, 144302. [Google Scholar] [CrossRef]
- Sıkdokur, E.; Belivermiş, M.; Sezer, N.; Pekmez, M.; Bulan, Ö.K.; Kılıç, Ö. Effects of Microplastics and Mercury on Manila Clam Ruditapes Philippinarum: Feeding Rate, Immunomodulation, Histopathology and Oxidative Stress. Environ. Pollut. 2020, 262, 114247. [Google Scholar] [CrossRef] [PubMed]
- Added, A.; Khalloufi, N.; Khazri, A.; Harrath, A.H.; Mansour, L.; Nahdi, S.; Boufahja, F.; Aldahmash, W.; Alrefaei, A.F.; Dellali, M. Effects of an Endocrine Disruptor Triclosan on Ruditapes Decussatus: Multimarker and Histological Approaches. Animals 2023, 13, 402. [Google Scholar] [CrossRef] [PubMed]
- Smolowitz, R. Mollusca: Bivalvia. In Invertebrate Histology; LaDouceur, E.E.B., Ed.; Wiley: New York, NY, USA, 2021; pp. 163–183. ISBN 978-1-119-50765-9. [Google Scholar]
- Liquin, F.; Hünicken, L.A.; Arrighetti, F.; Davies, D.; Paolucci, E.M.; Sylvester, F. Parasitism and Fitness of Invaders: Oligochaete Chaetogaster Limnaei Produces Gill Damage and Increased Respiration Rates in Freshwater Asian Clams. Hydrobiologia 2021, 848, 2213–2223. [Google Scholar] [CrossRef]
- Jemaà, M.; Morin, N.; Cavelier, P.; Cau, J.; Strub, J.M.; Delsert, C. Adult Somatic Progenitor Cells and Hematopoiesis in Oysters. J. Exp. Biol. 2014, 217, 3067–3077. [Google Scholar] [CrossRef] [PubMed]
- De Sales-Ribeiro, C.; Brito-Casillas, Y.; Fernandez, A.; Caballero, M.J. An End to the Controversy over the Microscopic Detection and Effects of Pristine Microplastics in Fish Organs. Sci. Rep. 2020, 10, 12434. [Google Scholar] [CrossRef] [PubMed]
- Karray, S.; Marchand, J.; Geffard, A.; Rebai, T.; Denis, F.; Chénais, B.; Hamza-Chaffai, A. Metal Contamination and Biomarkers in Cerastoderma glaucum: A Multi-Level Approach. Arch. Environ. Contam. Toxicol. 2023, 84, 484–503. [Google Scholar] [CrossRef] [PubMed]
- Mona, M.H.; El-Khodary, G.M.; Abdel-Halim, K.Y.; Omran, N.E.; Abd El-Aziz, K.K.; El-Saidy, S.A. Histopathological Alterations Induced by Marine Environmental Pollutants on the Bivalve Cerastoderma glaucum (Bruguière 1789) from Temsah Lake, Suez Canal, Egypt. Environ. Sci. Pollut. Res. 2022, 29, 9971–9989. [Google Scholar] [CrossRef] [PubMed]
- Ben Youssef-Dridi, S.; Magalhães, L.; Soares, A.M.V.M.; Pereira, E.; Freitas, R.; Gargouri, L. Trace Elements Assessment in Cerastoderma glaucum from Port Areas in the Tunisian Mediterranean Coast: The Influence of Parasites on Bioaccumulation. Mar. Pollut. Bull. 2024, 198, 115831. [Google Scholar] [CrossRef] [PubMed]
- Virginie Cuvillier-Hot; Céline Boidin-Wichlacz; Aurélie Tasiemski Polychaetes as Annelid Models to Study Ecoimmunology of Marine Organisms. J. Mar. Sci. Technol. 2014, 22, 2. [CrossRef]
- Gomiero, A.; Strafella, P.; Pellini, G.; Salvalaggio, V.; Fabi, G. Comparative Effects of Ingested PVC Micro Particles with and without Adsorbed Benzo (a) Pyrene vs. Spiked Sediments on the Cellular and Sub Cellular Processes of the Benthic Organism Hediste Diversicolor. Front. Mar. Sci. 2018, 5, 99. [Google Scholar] [CrossRef]
- Ellis, R.P.; Parry, H.; Spicer, J.I.; Hutchinson, T.H.; Pipe, R.K.; Widdicombe, S. Immunological Function in Marine Invertebrates: Responses to Environmental Perturbation. Fish. Shellfish. Immunol. 2011, 30, 1209–1222. [Google Scholar] [CrossRef] [PubMed]
- Bergami, G.; Bertini, F.; Montesi, D. On Approximate Nesting of Multiple Social Network Graphs: A Preliminary Study. In Proceedings of the 23rd International Database Applications & Engineering Symposium—IDEAS’19, Athens, Greece, 10–12 June 2019; ACM Press: Athens, Greece, 2019; pp. 1–5. [Google Scholar]
- Chen, J.-C.; Chen, M.-Y.; Fang, C.; Zheng, R.-H.; Jiang, Y.-L.; Zhang, Y.-S.; Wang, K.-J.; Bailey, C.; Segner, H.; Bo, J. Microplastics Negatively Impact Embryogenesis and Modulate the Immune Response of the Marine Medaka Oryzias melastigma. Mar. Pollut. Bull. 2020, 158, 111349. [Google Scholar] [CrossRef] [PubMed]
- Revel, M.; Yakovenko, N.; Caley, T.; Guillet, C.; Châtel, A.; Mouneyrac, C. Accumulation and Immunotoxicity of Microplastics in the Estuarine Worm Hediste Diversicolor in Environmentally Relevant Conditions of Exposure. Environ. Sci. Pollut. Res. 2020, 27, 3574–3583. [Google Scholar] [CrossRef] [PubMed]
- Franzenburg, S.; Fraune, S.; Künzel, S.; Baines, J.F.; Domazet-Lošo, T.; Bosch, T.C.G. MyD88-Deficient Hydra reveal an Ancient Function of TLR Signaling in Sensing Bacterial Colonizers. Proc. Natl. Acad. Sci. USA 2012, 109, 19374–19379. [Google Scholar] [CrossRef] [PubMed]
- Škanta, F.; Roubalová, R.; Dvořák, J.; Procházková, P.; Bilej, M. Molecular Cloning and Expression of TLR in the Eisenia andrei Earthworm. Dev. Comp. Immunol. 2013, 41, 694–702. [Google Scholar] [CrossRef] [PubMed]
- Lauriano, E.R.; Aragona, M.; Alesci, A.; Lo Cascio, P.; Pergolizzi, S. Toll-like Receptor 2 and α-Smooth Muscle Actin Expressed in the Tunica of a Urochordate, Styela Plicata. Tissue Cell 2021, 71, 101584. [Google Scholar] [CrossRef] [PubMed]
- Alesci, A.; Pergolizzi, S.; Savoca, S.; Fumia, A.; Mangano, A.; Albano, M.; Messina, E.; Aragona, M.; Lo Cascio, P.; Capillo, G.; et al. Detecting Intestinal Goblet Cells of the Broadgilled Hagfish Eptatretus cirrhatus (Forster, 1801): A Confocal Microscopy Evaluation. Biology 2022, 11, 1366. [Google Scholar] [CrossRef]
- Alesci, A.; Capillo, G.; Fumia, A.; Messina, E.; Albano, M.; Aragona, M.; Lo Cascio, P.; Spanò, N.; Pergolizzi, S.; Lauriano, E.R. Confocal Characterization of Intestinal Dendritic Cells from Myxines to Teleosts. Biology 2022, 11, 1045. [Google Scholar] [CrossRef]
- Alesci, A.; Albano, M.; Savoca, S.; Mokhtar, D.M.; Fumia, A.; Aragona, M.; Lo Cascio, P.; Hussein, M.M.; Capillo, G.; Pergolizzi, S. Confocal Identification of Immune Molecules in Skin Club Cells of Zebrafish (Danio rerio, Hamilton 1882) and Their Possible Role in Immunity. Biology 2022, 11, 1653. [Google Scholar] [CrossRef]
- Lauriano, E.R.; Alesci, A.; Aragona, M.; Pergolizzi, S.; Miller, A.; Zuwala, K.; Kuciel, M.; Zaccone, G.; Germanà, A.; Guerrera, M.C. Immunohistochemistry of the Gut-Associated Lymphoid Tissue (GALT) in African Bonytongue (Heterotis niloticus, Cuvier 1829). Int. J. Mol. Sci. 2023, 24, 2316. [Google Scholar] [CrossRef]
- Alesci, A.; Pergolizzi, S.; Fumia, A.; Calabrò, C.; Lo Cascio, P.; Lauriano, E.R. Mast Cells in Goldfish (Carassius auratus) Gut: Immunohistochemical Characterization. Acta Zool. 2023, 104, 366–379. [Google Scholar] [CrossRef]
- Ghosh, J.; Lun, C.M.; Majeske, A.J.; Sacchi, S.; Schrankel, C.S.; Smith, L.C. Invertebrate Immune Diversity. Dev. Comp. Immunol. 2011, 35, 959–974. [Google Scholar] [CrossRef]
- Sigh, J.; Lindenstrom, T.; Buchmann, K. The Parasitic Ciliate Ichthyophthirius multifiliis Induces Expression of Immune Relevant Genes in Rainbow Trout, Oncorhynchus mykiss (Walbaum). J. Fish. Dis. 2004, 27, 409–417. [Google Scholar] [CrossRef]
- Davies, S.-A.; Dow, J.A.T. Modulation of Epithelial Innate Immunity by Autocrine Production of Nitric Oxide. Gen. Comp. Endocrinol. 2009, 162, 113–121. [Google Scholar] [CrossRef]
- Rivero, A. Nitric Oxide: An Antiparasitic Molecule of Invertebrates. Trends Parasitol. 2006, 22, 219–225. [Google Scholar] [CrossRef]
- Wang, N.; Wang, T.; Zhao, X.; Chen, Y.; Liu, R.; Fang, Y.; Zhang, R. Molecular Characterization of the Nitric Oxide Synthase Gene and Its Immunomodulation of Nitric Oxide Production in the Triangle Shell Mussel (Hyriopsis cumingii). Dev. Comp. Immunol. 2021, 122, 104136. [Google Scholar] [CrossRef] [PubMed]
- Alesci, A.; Fumia, A.; Mastrantonio, L.; Marino, S.; Miller, A.; Albano, M. Functional Adaptations of Hemocytes of Aplysia depilans (Gmelin, 1791) and Their Putative Role in Neuronal Regeneration. Fishes 2024, 9, 32. [Google Scholar] [CrossRef]
- Alesci, A.; Albano, M.; Fumia, A.; Messina, E.; Miller, A.; Di Fresco, D.; De Oliveira Fernandes, J.M.; Spanò, N.; Savoca, S.; Capillo, G. Shell Formation in Two Species of Bivalves: The Role of Mantle Cells and Haemocytes. Zool. J. Linn. Soc. 2023, 200, 980–993. [Google Scholar] [CrossRef]
- Oliveira-Nascimento, L.; Massari, P.; Wetzler, L.M. The Role of TLR2 in Infection and Immunity. Front. Immun. 2012, 3, 79. [Google Scholar] [CrossRef]
- Taban, Q.; Mumtaz, P.T.; Masoodi, K.Z.; Haq, E.; Ahmad, S.M. Scavenger Receptors in Host Defense: From Functional Aspects to Mode of Action. Cell Commun. Signal 2022, 20, 2. [Google Scholar] [CrossRef] [PubMed]
- Underhill, D.M.; Gantner, B. Integration of Toll-like Receptor and Phagocytic Signaling for Tailored Immunity. Microbes Infect. 2004, 6, 1368–1373. [Google Scholar] [CrossRef]
- Ikuta, T.; Tame, A.; Takahashi, T.; Nomaki, H.; Nakajima, R. Microplastic Particles Are Phagocytosed in Gill Cells of Deep-Sea and Coastal Mussels. Front. Mar. Sci. 2022, 9, 1034950. [Google Scholar] [CrossRef]
- Sun, M.; Li, S.; Zhang, X.; Xiang, J.; Li, F. Isolation and Transcriptome Analysis of Three Subpopulations of Shrimp Hemocytes Reveals the Underlying Mechanism of Their Immune Functions. Dev. Comp. Immunol. 2020, 108, 103689. [Google Scholar] [CrossRef]
- De La Ballina, N.R.; Villalba, A.; Cao, A. Shotgun Analysis to Identify Differences in Protein Expression between Granulocytes and Hyalinocytes of the European Flat Oyster Ostrea edulis. Fish. Shellfish. Immunol. 2021, 119, 678–691. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antibody | Supplier | Product Number | Dilution | Animal Source |
|---|---|---|---|---|
| TLR2 | Active Motif, La Hulpe, Belgium | 40981 | 1:200 | Rabbit |
| iNOS | Santa Cruz Biotechnology, Inc., Dallas, TX, USA | sc-7271 | 1:200 | Mouse |
| Alexa Fluor 488 Donkey anti-Mouse IgG FITC conjugated | Molecular Probes, Invitrogen, Eugene, OR, USA | A21202 | 1:300 | Donkey |
| Alexa Fluor 594 Donkey anti-Rabbit IgG TRITC conjugated | Molecular Probes, Invitrogen, Eugene, OR, USA | A21207 | 1:300 | Donkey |
| Control | Pristine MPs | |
|---|---|---|
| TLR2 | 362.21 ± 67.91 * | 704.84 ± 96.54 ** |
| iNOS | 234.72 ± 65.73 * | 263.91 ± 43.41 * |
| TLR2 + iNOS | 123.12 ± 48.37 ** | 141.64 ± 39.21 * |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alesci, A.; Di Paola, D.; Marino, S.; De Gaetano, F.; Albano, M.; Morgante, S.; Rigano, G.; Giuffrè, L.; Kotanska, M.; Spanò, N.; et al. Exploring the Internal Defense System of Cerastoderma glaucum (Bruguière, 1789) Exposed to Pristine Microplastics: The Sentinel Role of Haemocytes as Biomarkers. Fishes 2024, 9, 241. https://doi.org/10.3390/fishes9060241
Alesci A, Di Paola D, Marino S, De Gaetano F, Albano M, Morgante S, Rigano G, Giuffrè L, Kotanska M, Spanò N, et al. Exploring the Internal Defense System of Cerastoderma glaucum (Bruguière, 1789) Exposed to Pristine Microplastics: The Sentinel Role of Haemocytes as Biomarkers. Fishes. 2024; 9(6):241. https://doi.org/10.3390/fishes9060241
Chicago/Turabian StyleAlesci, Alessio, Davide Di Paola, Sebastian Marino, Federica De Gaetano, Marco Albano, Silvana Morgante, Gabriele Rigano, Letterio Giuffrè, Magdalena Kotanska, Nunziacarla Spanò, and et al. 2024. "Exploring the Internal Defense System of Cerastoderma glaucum (Bruguière, 1789) Exposed to Pristine Microplastics: The Sentinel Role of Haemocytes as Biomarkers" Fishes 9, no. 6: 241. https://doi.org/10.3390/fishes9060241
APA StyleAlesci, A., Di Paola, D., Marino, S., De Gaetano, F., Albano, M., Morgante, S., Rigano, G., Giuffrè, L., Kotanska, M., Spanò, N., & Lauriano, E. R. (2024). Exploring the Internal Defense System of Cerastoderma glaucum (Bruguière, 1789) Exposed to Pristine Microplastics: The Sentinel Role of Haemocytes as Biomarkers. Fishes, 9(6), 241. https://doi.org/10.3390/fishes9060241