

Distribution Characteristics and Driving Factors of Collichthys lucidus Species in Offshore Waters of Zhejiang Province, China

 ,
,
Abstract
1. Introduction
2. Materials and Methods
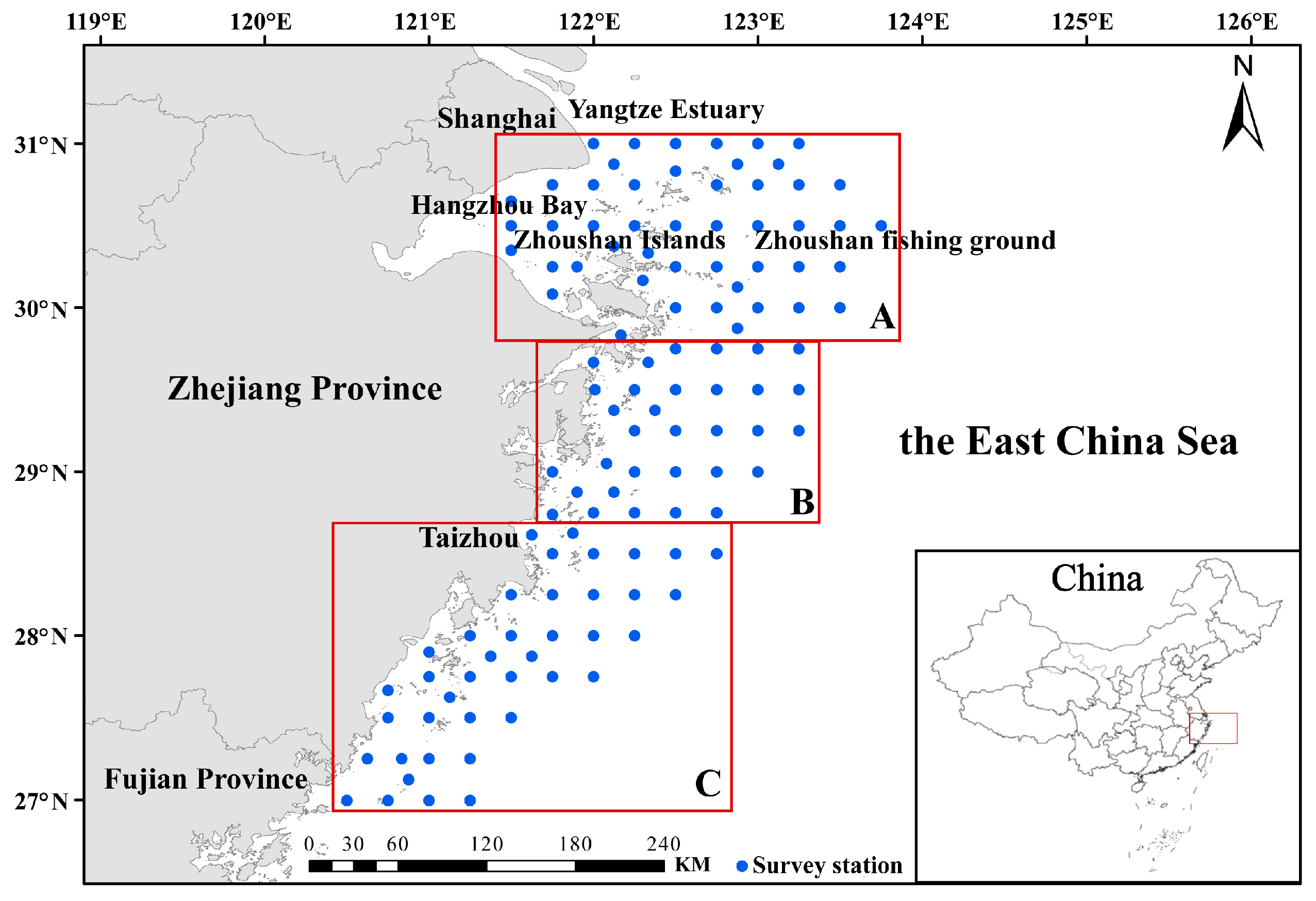
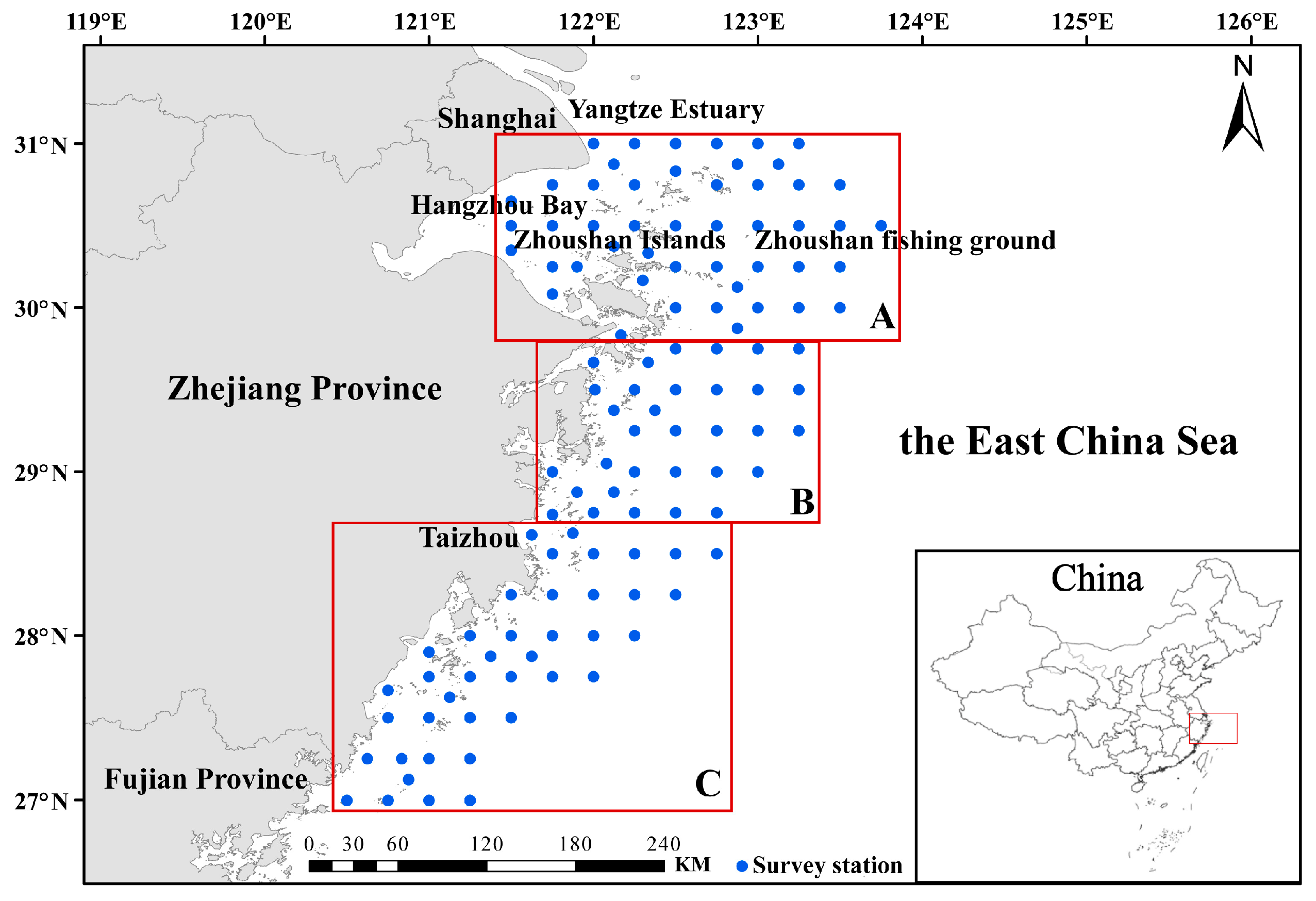
2.1. Data Source
2.2. Data Processing
2.3. Construction and Analysis of the Random Forest Model
2.4. Results Visualization
3. Results and Analysis
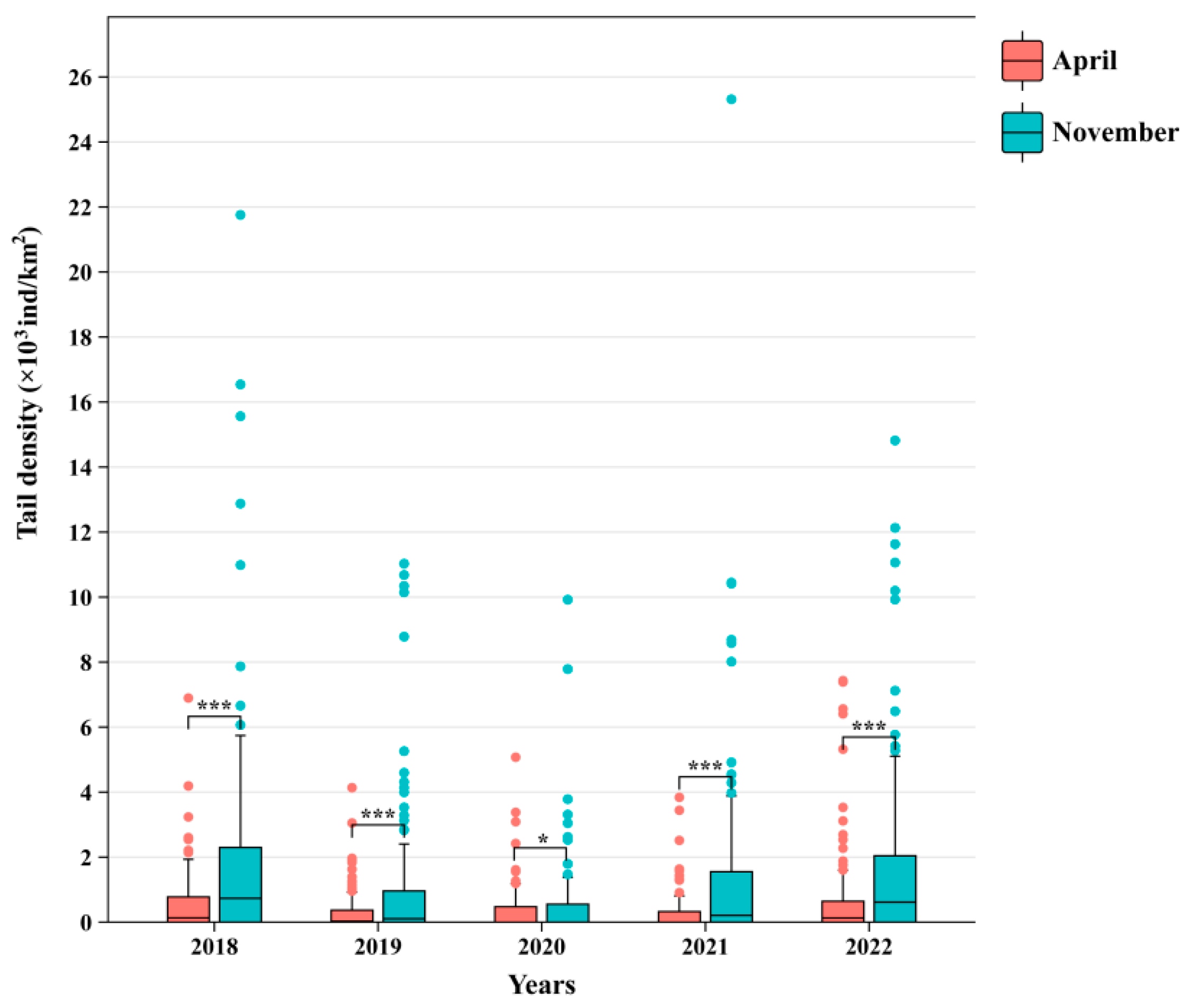
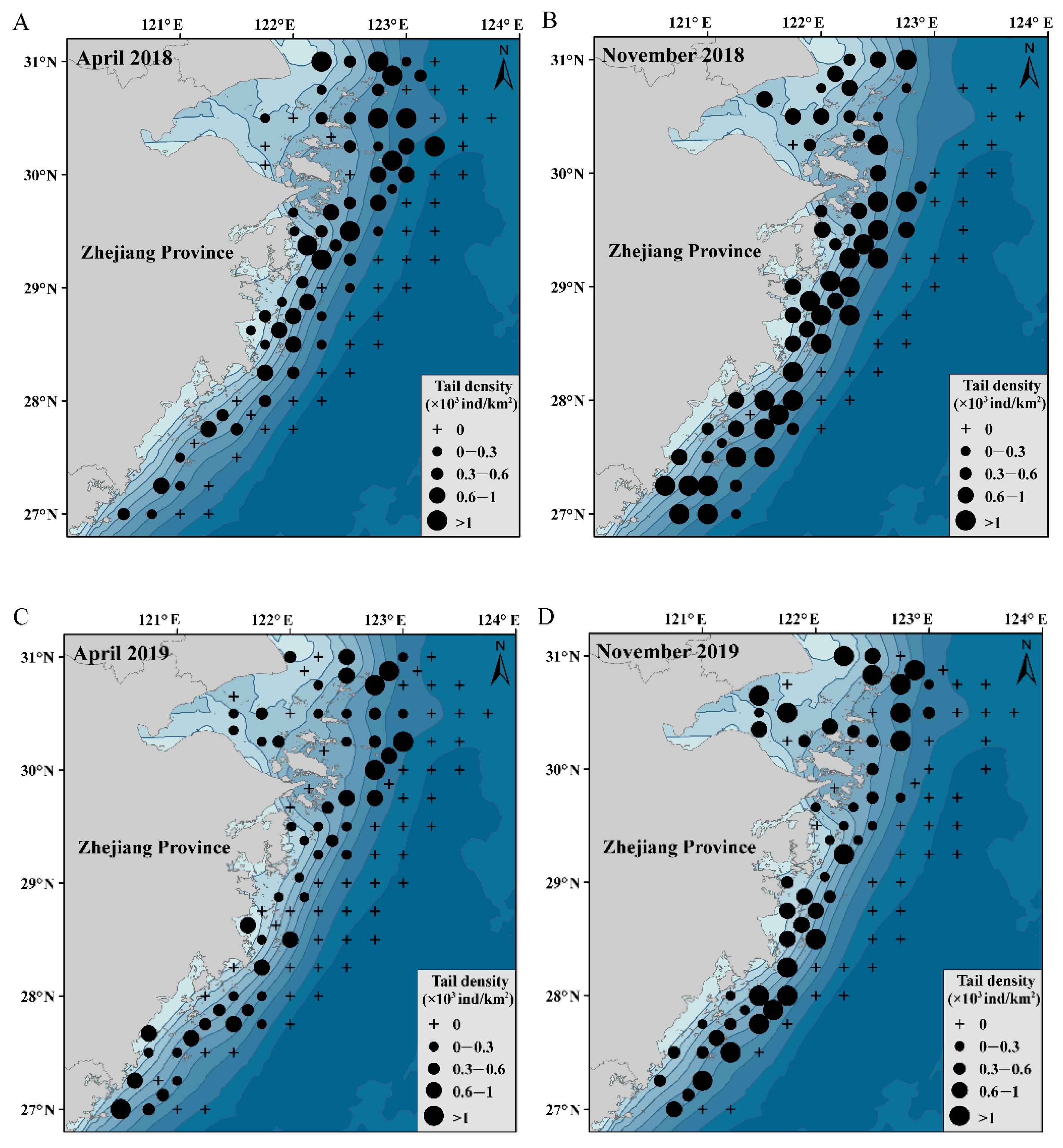
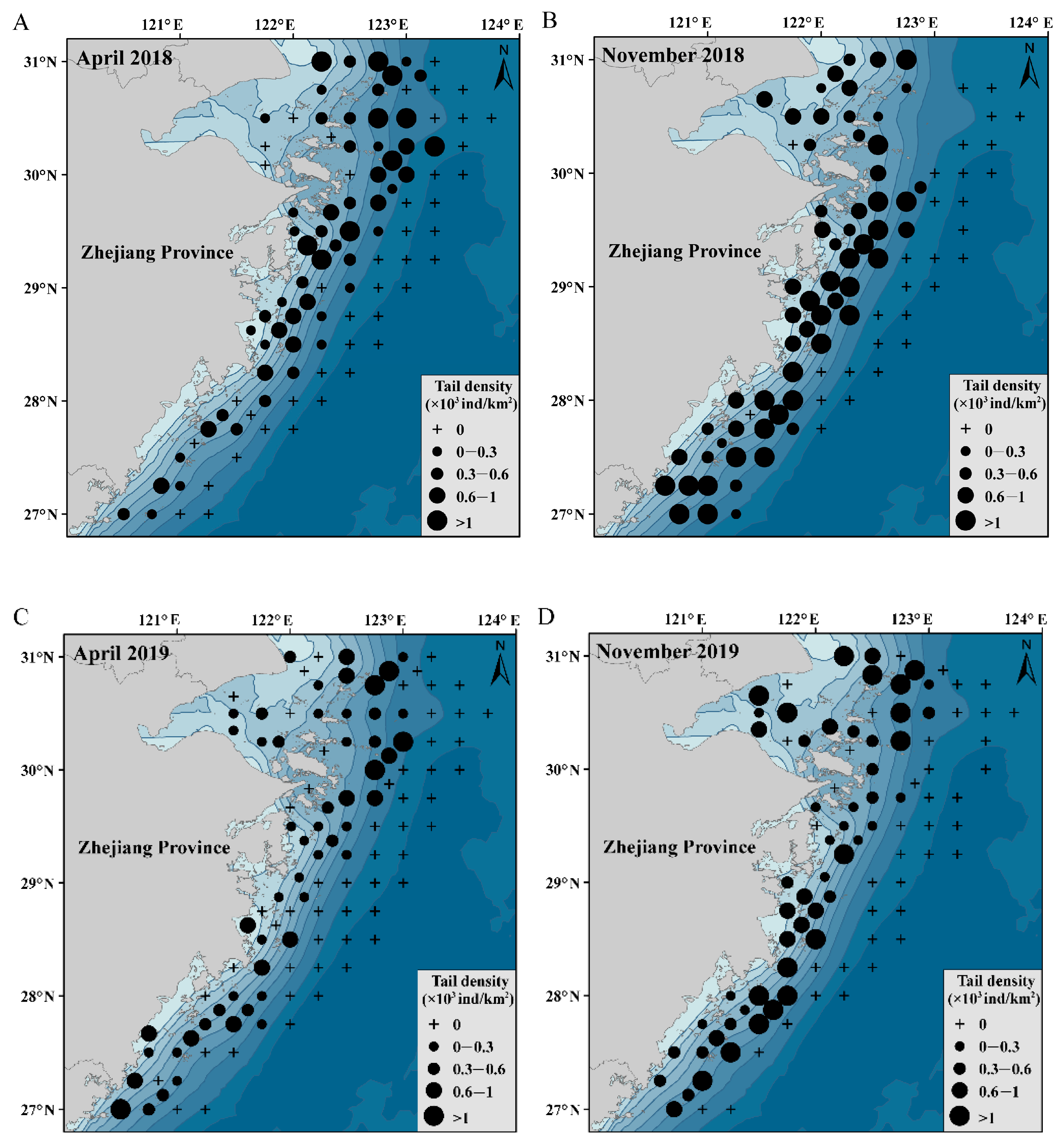
3.1. Changes in the Density of C. lucidus
3.2. Screening of Environmental Variables
3.3. Model Performance Assessment
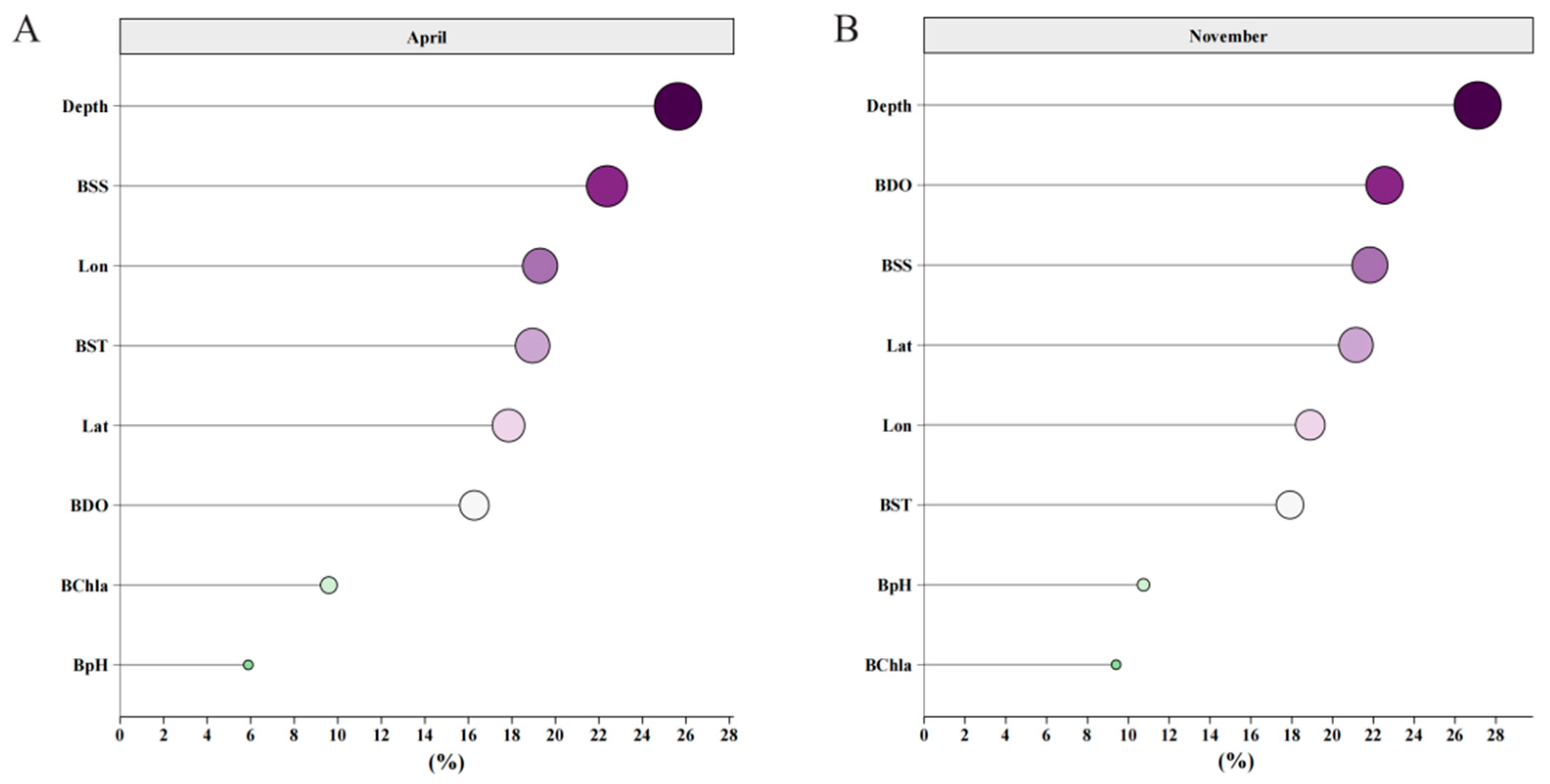
3.4. Importance Ranking of Explanatory Variables
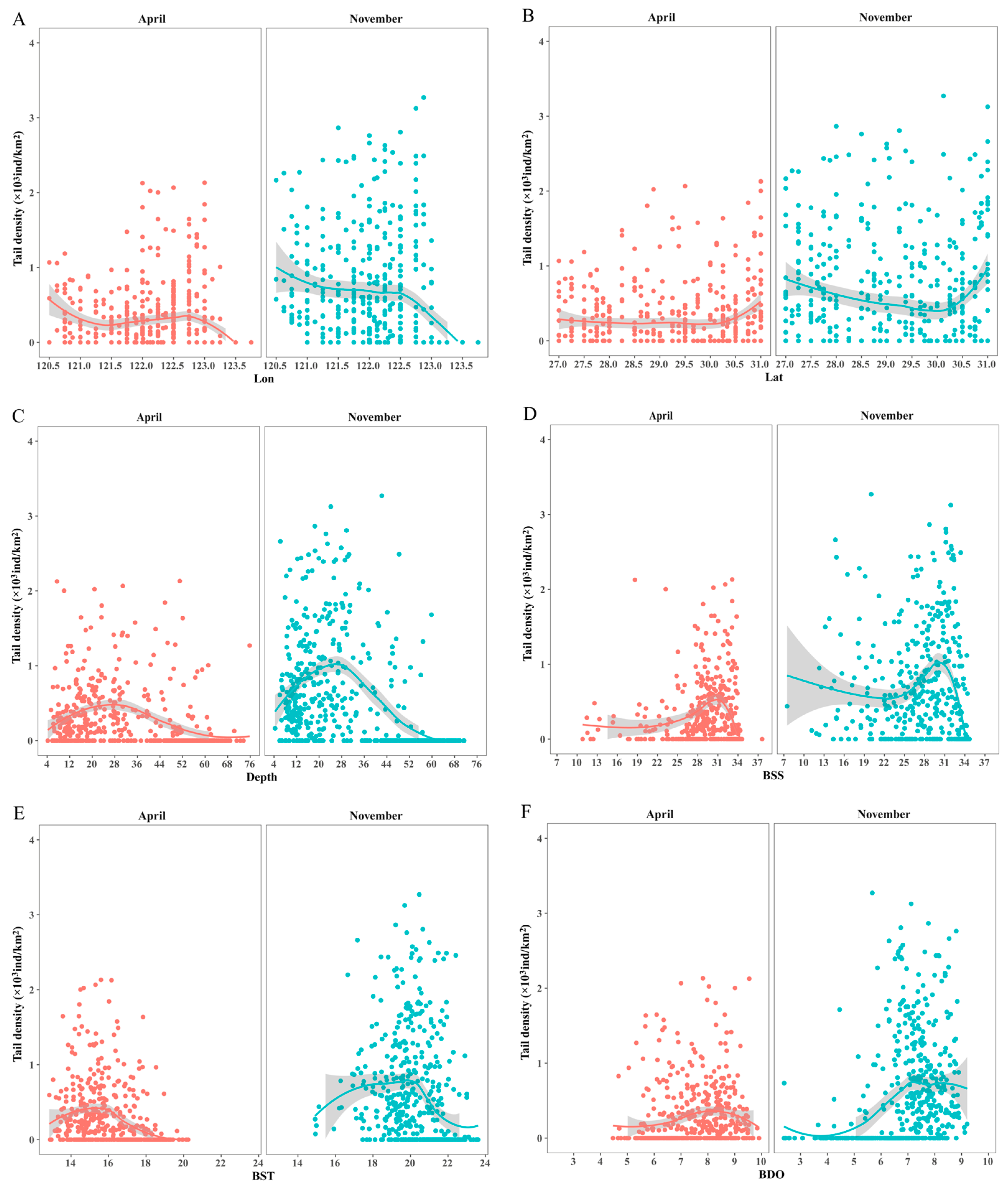
3.5. The Relationships between the Distributions of C. lucidus Species and Important Explanatory Variables
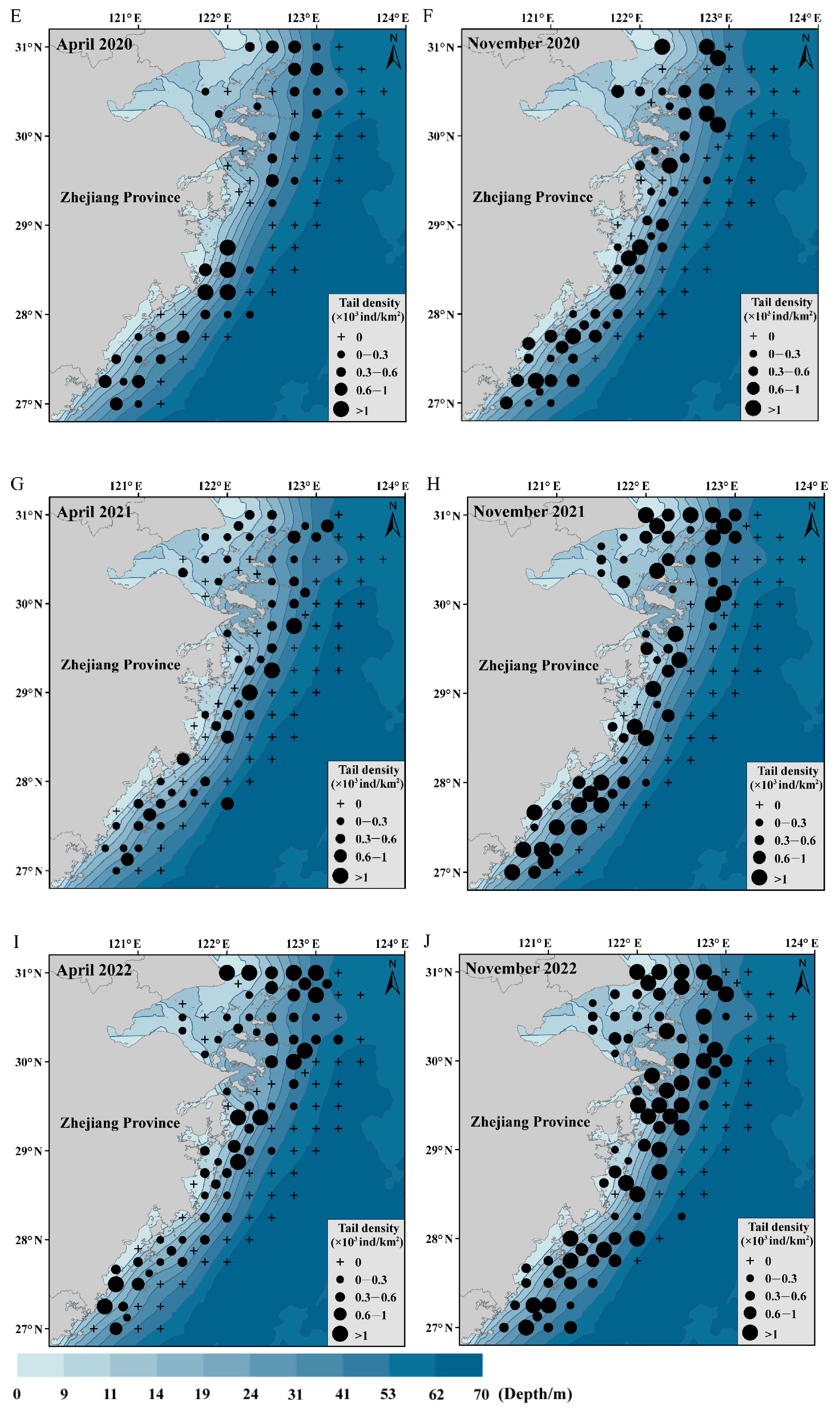
3.6. Spatial and Temporal Distributions of C. lucidus Species
4. Discussion
4.1. Spatiotemporal Changes in the Distribution of C. lucidus
4.2. Effects of Marine Variables on the Distribution of C. lucidus Species
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhao, S.L.; Xu, H.X.; Zhong, J.S.; Chen, J. Zhejiang Marine Fishes; Zhejiang Science and Technology Press: Hangzhou, China, 2016; pp. 86–92. [Google Scholar]
- Zhu, Y.D.; Zhang, C.L.; Cheng, Q.T. Fishes of East China Sea; Science Press: Beijing, China, 1963; pp. 241–243. [Google Scholar]
- Zhou, F.; Qian, Z.Y.; Liu, A.Q.; Ma, X.; Ni, X.B.; Zeng, D.Y. Recent progress on the studies of the physical mechanisms of hypoxia off the Changjiang (Yangtze River) Estuary. J. Mar. Sci. 2021, 39, 22–38. [Google Scholar] [CrossRef]
- Gao, F.; Chen, X.J.; Guan, W.J.; Li, G. A New model to forecast fishing ground of Scomber japonicus in the Yellow Sea and East China Sea. Acta Oceanol. Sin. 2016, 35, 74–81. [Google Scholar] [CrossRef]
- Hu, W.J.; Du, J.G.; Su, S.K.; Tan, H.J.; Yang, W.; Ding, L.K.; Dong, P.; Yu, W.W.; Zheng, X.Q.; Chen, B. Effects of climate change in the seas of China: Predicted changes in the distribution of fish species and diversity. Ecol. Indic. 2022, 134, 108489. [Google Scholar] [CrossRef]
- Ji, Q.; Xie, Z.L.; Gan, W.; Wang, L.M.; Song, W. Identification and Characterization of PIWI-Interacting RNAs in Spinyhead Croakers (Collichthys lucidus) by Small RNA Sequencing. Fishes 2022, 7, 297. [Google Scholar] [CrossRef]
- Song, W.; Gan, W.; Xie, Z.L.; Chen, J.; Wang, L.M. Small RNA sequencing reveals sex-related miRNAs in Collichthys lucidus. Front. Genet. 2022, 13, 955645. [Google Scholar] [CrossRef]
- Ou, Y.J.; Liao, R.; Li, J.E.; Gou, X.W. Studies on the spawning period and growth of Collichthys lucidus in estuary of Pearl River based on otolith daily annulus. J. Oceanogr. Taiwan Strait. 2012, 31, 85–88. [Google Scholar] [CrossRef]
- Zhang, H.Y.; Lin, L.S. Spatial heterogeneity of Trichiurus japonicus and small-scale fish in East China Sea and their spatial relationships. Chin. J. Appl. Ecol. 2005, 16, 708–711. [Google Scholar] [CrossRef]
- Wang, M.; Hong, B.; Zhang, Y.P.; Sun, Z.Z. Spring and summer fish community structure in northern Hangzhou bay. J. Hydrol. 2016, 37, 75–81. [Google Scholar] [CrossRef]
- Zhang, H.L.; Wang, Y.; Liang, J.; He, Z.T.; Zhou, Y.D. Seasonal Variations of the Biological Characteristics and Abundance Density of Collichthys lucidus in Zhongjieshan Islands Marine Protected Area. J. Zhejiang Ocean Univ. Nat. Sci. 2015, 34, 407–410. [Google Scholar]
- DeLuca, N.M.; Mullikin, A.; Brumm, P.; Rappold, A.G.; Hubal, E.C. Using Geospatial Data and Random Forest To Predict PFAS Contamination in Fish Tissue in the Columbia River Basin, United States. Environ. Sci. Technol. 2023, 57, 14024–14035. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.C.; Jia, M.X.; Feng, W.D.; Liu, C.D.; Huang, L.Y. Spatial-temporal distribution of Antarctic Krill (Euphausia superba)resource and its association with environment factors revealed with RF and GAM Models. J. Ocean Univ. China 2021, 51, 20–29. [Google Scholar] [CrossRef]
- Wu, T.; Liang, J.; Zhou, Y.D.; Xuan, W.D.; Fang, G.J.; Zhang, Y.Z.; Chen, F. The factors driving the spatial variation in the selection of spawning grounds for Sepiella japonica in offshore Zhejiang Province, China. Fishes 2024, 9, 20. [Google Scholar] [CrossRef]
- Zhang, S.; Li, M.; Yan, S.; Kong, X.l.; Zhu, J.F.; Xu, S.N.; Chen, Z.Z. Population genetic structure analysis of big head croaker (Collichthys lucidus) based on mitochondrial cytochrome b gene sequences. J. Fish. Sci. China 2021, 28, 90–99. [Google Scholar] [CrossRef]
- Wu, C.W.; Wang, W.H. Distribution biology and resource changes of Collichthys lucidus in Zhejiang coastal waters. Marine Fish. 1991, 13, 6–10. [Google Scholar]
- Wu, Z.X.; Wu, C.W.; Wang, W.H.; Hu, N.Q. Growth law of Collichthys lucidus in Zhejiang offshore research on group composition. Fish. Sci. Technol. Inf. 1990, 6, 170–174. [Google Scholar] [CrossRef]
- GB/T 12763.6-2007; Specifications for Oceanographic Survey-Part 6: Marine Biological Survey. General Administration of Quality Supervision, Inspection and Quarantine of the People’s Republic of China: Beijing, China, 2007.
- GB/T 17378.3-2007; The Specification for Marine Monitoring-Part 3: Sample Collection, Storage and Transportation. General Administration of Quality Supervision, Inspection and Quarantine of the People’s Republic of China: Beijing, China, 2007.
- Xiong, P.L.; Xu, S.N.; Chen, Z.Z.; Zhang, S.; Jiang, P.W.; Fan, J.T. Spatiotemporal distribution of Collichthy lucidus in the Pearl River Estuary and its relationship with environmental factors. Mar. Sci. 2022, 46, 79–87. [Google Scholar] [CrossRef]
- Zou, M.X.; Chen, Y.G.; Song, X.J.; Li, S.F.; Zhong, J.S. Distribution and drift trend of Collichthys lucidus larvae and juveniles in the coastal waters of the southern Yellow Sea. J. Fish. China 2022, 46, 557–568. [Google Scholar] [CrossRef]
- Shi, Y.R.; Chao, M.; Quan, W.M.; Tang, F.H.; Shen, X.Q.; Yuan, Q.; Huang, H.J. Spatial variation in fish community of Yangtze River estuary in spring. J. Fish. China 2011, 18, 1141–1151. [Google Scholar] [CrossRef]
- Yang, C.H.; Li, X.D.; Yang, Q.; Wang, Y.B. Application of machine learning methods for estimating the biomass of economically important crabs in the Zhoushan fishery. Mar. Sci. 2023, 47, 61–70. [Google Scholar] [CrossRef]
- Gómez, S.R.; Sánchez, R.A.; García, G.C.; Pérez, J.G. The VIF and MSE in Raise Regression. Mathematics 2020, 8, 605. [Google Scholar] [CrossRef]
- Flores, A.; Wiff, R.; Donovan, C.; Gálvez, P. Applying machine learning to predict reproductive condition in fish. Ecol. Inform. 2024, 80, 102481. [Google Scholar] [CrossRef]
- Rahman, L.F.; Marufuzzaman, M.; Alam, L.; Bari, M.A.; Sumaila, U.R.; Sidek, L.M. Application of Machine Learning to Investigate the Impact of Climatic Variables on Marine FishLandings. Natl. Acad. Sci. Lett. 2022, 45, 245–248. [Google Scholar] [CrossRef]
- Kang, H.; Jeon, D.J.; Kim, D.; Jung, K. Estimation of fish assessment index based on ensemble artificial neural network for aquatic ecosystem in South Korea. Ecol. Indic. 2022, 136, 108708. [Google Scholar] [CrossRef]
- Xu, B.D.; Zhang, C.L.; Xue, Y. Optimization of sampling effort for a fishery-independent survey with multiple goals. Environ. Monit. Assess. 2015, 187, 252. [Google Scholar] [CrossRef]
- Hitomi, O.; Satoshi, S.; Daisuke, A. Modeling the growth, transport, and feeding migration of age-0 Pacific saury Cololabis saira. Fish Sci. 2022, 88, 131–147. [Google Scholar]
- Wang, M.; Xu, K.D.; Liang, J. Preliminary analysis of biological characteristics of reproductive stocks of Collichthys lucidus in Northern Hangzhou Bay. J. Ocean Univ. Shanghai 2018, 27, 781–788. [Google Scholar] [CrossRef]
- Wang, Y.L.; Wang, J.; Zhou, Y.D.; Xu, K.D.; Jiang, R.J.; Li, Z.H.; Zhu, H.C.; Wang, J.; Cui, G.C. Spatial and temporal distribution characteristics of Larimichthys polyactis in Zhoushan fishing ground and the adjacent waters based on two-stage GAM. Fish. J. Fish. China 2022, 29, 633–641. [Google Scholar] [CrossRef]
- Papcunová, V.; Gregáňová, R.H. Global Impacts of COVID-19 on the Financing of Local Self-Governments: Evidence from Slovak municipalities. SHS Web Conf. 2021, 92, 01038. [Google Scholar] [CrossRef]
- Jackson, J.B.; Kirby, M.X.; Berger, W.H.; Bjorndal, K.A.; Botsford, L.W.; Bourque, B.J.; Bradbury, R.; Cooke, R.G.; Erlandson, J.M.; Estes, J.A.; et al. Historical overfishing and the recent collapse of coastal ecosystems. Science 2001, 293, 629–637. [Google Scholar] [CrossRef]
- Al Shehhi, M.R.; Samad, Y.A. Effects of the Covid-19 pandemic on the oceans. Remote Sens. Lett. 2021, 12, 325–334. [Google Scholar] [CrossRef]
- He, Z.T.; Zhang, Y.Z.; Xue, L.J.; Jin, H.W.; Zhou, Y.D. Seasonal and ontogenetic diet composition variation of Collichthys lucidus in inshore waters in the north of East China Sea. Mar. Fish. 2012, 34, 270–276. [Google Scholar] [CrossRef]
- Pothoven, A.S. The influence of ontogeny and prey abundance on feeding ecology of age-0 Lake Whitefish (Coregonus clupeaformis) in southeastern Lake Michigan. Ecol. Freshw. Fish 2020, 29, 103–111. [Google Scholar] [CrossRef]
- Sun, X.H.; Song, M.P.; Li, Z.G.; Song, Y.; Yuan, X.N.; Dong, B.; Zhang, L.; Zhu, L.X.; Liang, Z.L. Selective feeding of the mullet larvae Liza haematocheila during ontogeny in Laizhou Bay, Bohai Sea, China: The importance of small copepods in mesozooplankton as prey. Front. Mar. Sci. 2023, 10, 1147886. [Google Scholar] [CrossRef]
- Cai, R.S.; Chen, J.L.; Huang, R.H. The response of marine environment in the offshore area of China and its adjacent ocean to recent global climate change. Chin. J. Atmos. Sci. 2006, 30, 1019–1033. [Google Scholar]
- Xiao, J.Z.; Zou, Y.; Xiao, S.J.; Chen, J.N.; Wang, Z.Y.; Wang, Y.L.; Jie, X.M.; Cai, Y.M. Development of a PCR-based genetic sex identification method in spinyhead croaker (Collichthys lucidus). Aquac. Rep. 2020, 522, 735130. [Google Scholar] [CrossRef]
- Song, N.; Ma, G.Q.; Zhang, X.M.; Gao, T.X.; Sun, D.R. Genetic structure and historical demography of Collichthys lucidus inferred from mtDNA sequence analysis. Environ. Biol. Fish. 2014, 97, 69–77. [Google Scholar] [CrossRef]
- Francis, M.P.; Hurst, R.J.; McArdle, B.H.; Bagley, N.W.; Anderson, O.F. New Zealand demersal fish assemblages. Environ. Biol. Fish. 2002, 65, 215–234. [Google Scholar] [CrossRef]
- Jiang, R.J.; Sun, H.Q.; Li, X.F.; Zhou, Y.D.; Chen, F.; Xu, K.D.; Li, P.F.; Zhang, H.L. Habitat suitability evaluation of Harpadon nehereus in nearshore of Zhejiang province, China. Front. Mar. Sci. 2022, 9, 961735. [Google Scholar] [CrossRef]
- Bhat, M.; Nayak, V.N.; Chandran, M.D.S.; Ramachandra, T.V. Fish distribution dynamics in the Aghanashini estuary of Uttara Kannada, west coast of India. Curr. Sci. India 2014, 106, 1739–1744. [Google Scholar] [CrossRef]
- Zhang, S.; Jiang, Y.E.; Li, M.; Zhu, J.F.; Xu, S.N.; Chen, Z.Z. Life history of spinyhead croaker Collichthys lucidus (Sciaenidae) differentiated among populations from Chinese coastal waters. Aquat. Biol. 2022, 31, 65–76. [Google Scholar] [CrossRef]
- Liu, H.B.; Jiang, T.; Huang, H.H.; Shen, X.Q.; Zhu, J.B.; Yang, J. Estuarine dependency in Collichthys lucidus of the Yangtze River Estuary as revealed by the environmental signature of otolith strontium and calcium. Environ. Biol. Fish. 2015, 98, 165–172. [Google Scholar] [CrossRef]
- Lewin, W.C.; Mehner, T.; Ritterbusch, D.; Brämick, U. The influence of anthropogenic shoreline changes on the littoral abundance of fish species in German lowland lakes varying in depth as determined by boosted regression trees. Hydrobiologia 2014, 724, 293–306. [Google Scholar] [CrossRef]
- Rabalais, N.N.; Diaz, R.J.; Levin, L.A.; Turner, R.E.; Gilbert, D.; Zhang, J. Dynamics and distribution of natural and human-caused hypoxia. Biogeosciences 2010, 7, 585–619. [Google Scholar] [CrossRef]
- Maes, J.; Stevens, M.; Breine, J. Modelling the migration opportunities of diadromous fish species along a gradient of dissolved oxygen concentration in a European tidal watershed. Estuar. Coast. Shelf Sci. 2007, 75, 151–162. [Google Scholar] [CrossRef]
- Rabalais, N.N.; Cai, W.J.; Carstensen, J.; Conley, D.J.; Fry, B.; Hu, X.P.; Quiñones-Rivera, Z.; Rosenberg, R.; Slomp, C.P.; Turner, R.E.; et al. Eutrophication-Driven deoxygenation in the coastal ocean. Oceanography 2014, 27, 172–183. [Google Scholar] [CrossRef]
- Wang, H.J.; Dai, M.H.; Liu, J.W.; Kao, S.J.; Zhang, C.; Cai, W.J.; Wang, G.Z.; Qian, W.; Zhao, M.X.; Sun, Z.Y. Eutrophication-Driven hypoxia in the East China Sea off the Changjiang Estuary. Environ. Sci. Technol. 2016, 50, 2255–2263. [Google Scholar] [CrossRef]
- Zhang, Y. Analysis and evaluation of nutritional component in muscle of Collichthys lucidus. Sci. Fish Farming 2022, 43, 75–76. [Google Scholar] [CrossRef]
- Sato, K.N.; Levin, L.A.; Schiff, K. Habitat compression and expansion of sea urchins in response to changing climate conditions on the California continental shelf and slope (1994–2013). Deep Sea Res. Part II 2017, 137, 377–389. [Google Scholar] [CrossRef]
- Culter, D.R.; Edwards, T.C.; Beard, K.H.; Hess, K.T. Random forests for classification in ecology. Ecology 2007, 88, 2783–2792. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Seasons | Stations | Biomass (kg/km2) | Tail Density (×103 ind/km2) | Average Individual Relative Weight (g) | ||||
|---|---|---|---|---|---|---|---|---|
| Investigation Stations | Occurrence Stations | Maximum Value | Average Value | Maximum Value | Average Value | |||
| Spring | 2018 | 98 | 56 | 121.30 | 16.10 | 6.89 | 0.60 | 26.83 |
| 2019 | 108 | 54 | 97.47 | 9.02 | 4.14 | 0.34 | 26.16 | |
| 2020 | 79 | 39 | 220.42 | 14.53 | 5.07 | 0.43 | 34.06 | |
| 2021 | 111 | 55 | 114.78 | 7.58 | 3.84 | 0.29 | 25.93 | |
| 2022 | 114 | 65 | 148.68 | 14.47 | 7.43 | 0.68 | 21.27 | |
| Autumn | 2018 | 90 | 61 | 354.89 | 40.23 | 21.76 | 2.13 | 18.93 |
| 2019 | 94 | 53 | 269.30 | 27.27 | 11.03 | 1.21 | 22.48 | |
| 2020 | 99 | 52 | 184.02 | 15.78 | 9.92 | 0.71 | 22.31 | |
| 2021 | 110 | 61 | 611.09 | 35.79 | 25.31 | 1.59 | 22.49 | |
| 2022 | 112 | 80 | 314.96 | 34.24 | 14.82 | 1.79 | 19.17 | |
| Parameter | Month | Explanatory Variables | |||||
|---|---|---|---|---|---|---|---|
| Depth | BST | BSS | BChla | BDO | BpH | ||
| VIF | April | 2.787 | 2.188 | 2.410 | 1.113 | 2.792 | 1.152 |
| November | 2.441 | 1.805 | 1.984 | 1.214 | 1.769 | 1.057 | |
| Month | Model Fitting | Fivefold Cross-Validation | Explanation (%) | ||||
|---|---|---|---|---|---|---|---|
| RMSE | R2 | MAE | RMSE | R2 | MAE | ||
| April | 0.180 | 0.873 | 0.113 | 0.355 ± 0.060 | 0.251 ± 0.054 | 0.234 ± 0.028 | 23.26 |
| November | 0.263 | 0.912 | 0.179 | 0.554 ± 0.045 | 0.395 ± 0.072 | 0.390 ± 0.024 | 34.88 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xuan, W.; Zhang, H.; Zhang, H.; Wu, T.; Zhou, Y.; Zhu, W. Distribution Characteristics and Driving Factors of Collichthys lucidus Species in Offshore Waters of Zhejiang Province, China. Fishes 2024, 9, 83. https://doi.org/10.3390/fishes9030083
Xuan W, Zhang H, Zhang H, Wu T, Zhou Y, Zhu W. Distribution Characteristics and Driving Factors of Collichthys lucidus Species in Offshore Waters of Zhejiang Province, China. Fishes. 2024; 9(3):83. https://doi.org/10.3390/fishes9030083
Chicago/Turabian StyleXuan, Wendan, Hongliang Zhang, Haobo Zhang, Tian Wu, Yongdong Zhou, and Wenbin Zhu. 2024. "Distribution Characteristics and Driving Factors of Collichthys lucidus Species in Offshore Waters of Zhejiang Province, China" Fishes 9, no. 3: 83. https://doi.org/10.3390/fishes9030083
APA StyleXuan, W., Zhang, H., Zhang, H., Wu, T., Zhou, Y., & Zhu, W. (2024). Distribution Characteristics and Driving Factors of Collichthys lucidus Species in Offshore Waters of Zhejiang Province, China. Fishes, 9(3), 83. https://doi.org/10.3390/fishes9030083