Abstract
The effects of various levels of alkalinity stress (0, 18.25, 35.41, 52.53 and 69.74 mmol/L) in Chinese mitten crab (Eriocheir sinensis) were investigated by means of measuring hepatopancreas antioxidant system and serum nonspecific immune system-related indices at 0, 12, 24, 48 and 96 h, hepatopancreas tissue structure at 96 h. The activities of superoxide dismutase (SOD), catalase (CAT), glutathione (GSH) and aspartate aminotransferase (AST) in each concentration group generally showed a trend of a first increasing and then decreasing during the 96-h stress process (p < 0.05), while no significant changes were observed in the blank group during this period (p > 0.05). The activities of CAT, GSH and AST in all treatment reached peak values at 24–48 h. At 96 h of alkalinity stress, the activities of GSH, alanine aminotransferase (ALT), acid phosphatise (ACP) and alkaline phosphatise (AKP) in the 18.25 mmol/L group were not significantly different with the control group (p > 0.05). The activities of SOD and CAT in the 52.53 and 69.74 mmol/L treatment were significantly lower than those in the control group (p < 0.05), and the level of total antioxidant capacity(T-AOC), ALT, ACP and AKP in the 69.74 mmol/L group were significantly higher than those in the other groups (p < 0.05). Hepatopancreatic histological observation showed that the hepatopancreas of E. sinensis in the control group was normal. With increasing alkalinity, the basal membrane of the hepatopancreas fell off or even ruptured. Additionally, the number of hepatopancreas vacuoles increased, the volume of B cells and their internal transport vesicles increased, epithelial cells disintegrated, and the nucleus gradually shrank. E. sinensis can activate antioxidant and nonspecific immune systems to adapt to alkalinity stress. However, oxidative stress, immune system damage and hepatopancreas structure damage were caused when the stress exceeded the adaptive capacity of the body.
1. Introduction
Approximately 33.3% of the total land area worldwide consists of saline-alkaline land [1,2]. It has been estimated that saline-alkaline water covers approximately 45.87 million hectares in China alone, saline-alkali water resources are abundant and widely distributed [3]. Saline-alkali water has the characteristics of high pH, high ion coefficient and main ion imbalance, resulting in an extremely low utilization rate [4]. In the carbonate saline-alkali water environment, with increasing carbonate alkalinity, the pH and HCO3− content in the water body increased accordingly. These ions directly act on the surface epithelial cells of the gills of aquatic animals, causing the gill epithelium to be separated or ruptured and chloride cells to proliferate or experience hypertrophy, thereby affecting the ion exchange system on the cell surface [5]. In addition, HCO3− affected the balance system of carbon dioxide in the blood to harm aquatic animals [6]. The discharge of domestic and industrial sewage and the unreasonable utilization of water resources lead to the decreasing aquaculture spaces [7]. The research showed that as a result of global climate changes, the process of water environment salinization and alkalinization might continue to deepen, and the available spaces for freshwater aquaculture might steadily shrink [8]. Therefore, the development and utilization of saline-alkali water resources through fishery methods is of great significance for expanding aquaculture space and ensuring aquatic products. At present, there are few studies on the effect of alkalinity on aquatic animals. Carbonate alkalinity suppresses growth, survival and reproduction in aquaculture [2,9,10,11,12], substantiated with inflammation and abnormality in immunoregulation, aerobic and ammonia excretion rates in some species [13,14].
Chinese mitten crab, Eriocheir sinensis, (Phylum Arthropoda, Class Crustacea, Order Decapoda, Family Grapsidae), commonly known as river crabs. The life cycle of E. sinensis is completed by amphidromous migration [15]. As an important aquaculture economic crab in China, with the rapid expansion of their culture area, they have become one of the most important economic crabs in China. Crab farming is developing in deep inland area with arid climate conditions, because crab farming doesn’t require much water resources [16,17]. E. sinensis has a strong adaptability to stress conditions such as ammonia nitrogen, pH and heavy metal ions, and it has been used as a target crab for culture in saline and alkaline waters in deep inland areas in China under arid climate conditions [18,19]. In the present study, the effects of alkalinity on the antioxidant and nonspecific immune systems as well as hepatopancreas tissue structure of E. sinensis were studied under different alkalinity stresses of carbonate to provide a practical basis for the development of E. sinensis culture technology in saline and alkaline waters in the arid areas.
2. Materials and Methods
2.1. Crabs and Rearing Conditions
The E. sinensis used in the experiment originated from a crab culture farm in Jiangsu Province, China. Healthy, neat and complete juvenile crabs with an average body weight of 12.6 ± 1.5 g were selected for the experiment. Before experimentation, healthy juvenile E. sinensis were acclimated for 15 days in cycling filtered glass tanks (64 cm × 38 cm × 43 cm) containing 30 L aerated tap water. The temperature was maintained at a range of 22.0 to 23.0 °C, with dissolved oxygen concentration of >6.0 mg/L, and a pH of 7.0 to 8.0. During acclimation, the crabs were daily fed with a commercial formulated diet (Nanjing Aohua Biotechnology Co., Ltd., Nanjing, China) accounting for 5% of body weight. Residuals and faces were removed, and one-third of the water was renewed per day. Feeding was stopped for 24 h before experimentation.
Based on the 96 h median lethal dose (LC50, 69.74 mmol/L) and safe concentration (SC, 18.25 mmol/L) of alkalinity in E. sinensis [20], four concentration gradient groups (18.25, 35.41, 52.53 and 69.74 mmol/L) were set up using equal logarithmic distance method [21]. Aerated freshwater and analytical pure sodium bicarbonate (NaHCO3) were used to adjust the target alkalinity, a control group without NaHCO3 (alkalinity, 0.49 mmol/L) were carried out in this experiment. Three parallel groups were set for each concentration gradient in each group and 30 crabs were placed in each parallel group. The healthy acclimated individuals were randomly put into each experimental tank for 96 h. During the experiment, no feed was given, the temperature was maintained at 22.0–23.0 °C. Methyl orange hydrochloride calibration method was used to measure and adjust the alkalinity for ensuring the concentration stable in each group.
2.2. Sample Preparations
Chinese mitten crabs (three crabs were taken randomly per glass tank) in each treatment were sampled at 0, 12, 24, 48, and 96 h. Haemolymph was extracted from the soft membrane of the joint at its root of the third paraeiopod with a sterilized 1 mL syringe and was treated without anticoagulant. All the haemolymphs were centrifuged with 8000 r/min for 10 min after been kept 24 h at 4 °C, and then the supernatant was transferred to a centrifuge tube and cryopreserved at −80 °C for further examination for immunoenzyme activity analysis. At the same time, the hepatopancreas of each crab was dissected and stored in a −80 °C refrigerator after freezing with liquid nitrogen for antioxidant enzyme activity analysis. Each hepatopancreas sample was homogenized after adding 0.86% normal saline at a weight/volume ratio of 1:9. The hepatopancreas homogenate was centrifuged with 3500 r/min for 15 min at 4 °C, and the upper lipid layer was discarded. The supernatant was carefully collected and immediately used for measuring enzymatic activity.
2.3. Biochemical Analysis
The activities of superoxide dismutase (SOD), catalase (CAT), and glutathione (GSH), the level of total antioxidant capacity (T-AOC) in hepatopancreas were determined using commercially available assay kits according to the manufacturer’s protocols (Jiancheng Ltd., Nanjing, China). Protein concentration was determined by a Coomassie Brilliant Blue protein assay kit (Jiancheng Ltd., Nanjing, China). The results were read using a microplate reader (Synergy HTX, BioTek, Winooski, VT, USA) and a spectrophotometer (YoKe, Shanghai, China). The activities of acid phosphatise (ACP), alkaline phosphatise (AKP), aspartate aminotransferase (AST) and alanine aminotransferase (ALT) in serum were determined using a fully automatic bioanalysis machine (Mindray BS-240Vet, Shenzhen, China).
2.4. Hepatopancreas Histological Analysis
Three crabs were randomly obtained from the four treatments and the control group at 96 h of stress, and the hepatopancreas was quickly removed. The hepatopancreas was maintained in 10% formalin fixative, and the tissues were embedded, sliced, and stained with haematoxylin-eosin (HE). The sections were observed under a microscope.
2.5. Statistical Analysis
All data in the figures are expressed as the mean ± standard deviation (S.D.). The experimental data was conducted with SPSS 19.0 software. A homogeneity test of variances was performed, followed by one-way analysis of variance, and a multiple comparison of the Duncan test was used to determine significant differences among all groups. Differences were considered significant at p < 0.05. GraphPad Prism 5.0 software was used for mapping.
3. Results
3.1. Antioxidant Enzymes
During the whole experiment, the hepatopancreas SOD activities in the control did not change significantly (p > 0.05), while the SOD and CAT activities of the experimental group showed a trend of first increasing and then decreasing except in the 35.41 mmol/L concentration group (p < 0.05; Figure 1 and Figure 2). The activities of SOD and CAT in the experimental group reached a peak at 24–48 h. In the 35.41 mmol/L concentration group, the hepatopancreas SOD activity significantly increased (p < 0.05), and hepatopancreas CAT activity increased significantly and reached a peak at 24 h (p < 0.05), then decreased significantly from 24 h to 96 h. The CAT peak level in 18.25 mmol/L group at 12 h was significantly lower than that in 35.41 mmol/L group (p < 0.05). After 96 h of stress, the CAT activity in 18.25 mmol/L group returned to the initial level (p > 0.05), while the SOD and CAT activities in 35.41 mmol/L group were significantly higher than those in control (p < 0.05), the SOD and CAT activities in 52.53 and 69.74 mmol/L groups were significantly lower than those in control group (p < 0.05).
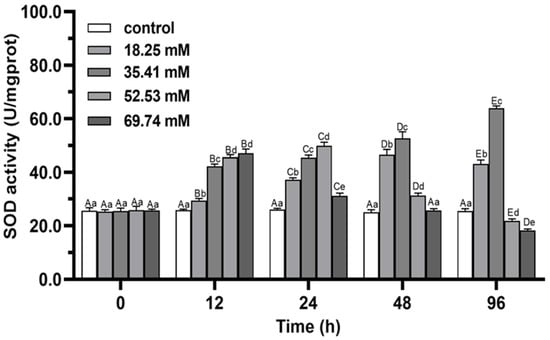
Figure 1.
Effects of alkalinity stress on SOD enzyme activity in the hepatopancreas of Eriocheir sinensis. Different capital letters indicate significant differences in the same alkalinity group (p < 0.05), and different lowercase letters indicate significant differences at the same time point (p < 0.05).
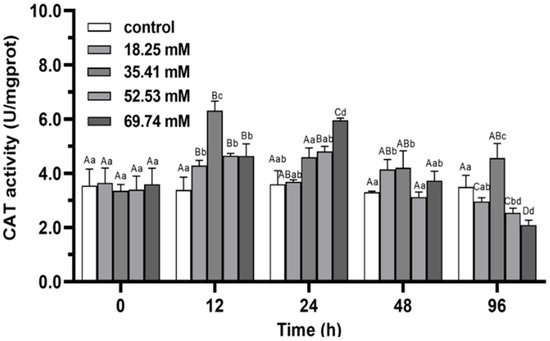
Figure 2.
Effects of alkalinity stress on CAT enzyme activity in the hepatopancreas of Eriocheir sinensis. Different capital letters indicate significant differences in the same alkalinity group (p < 0.05), and different lowercase letters indicate significant differences at the same time point (p < 0.05).
Within 96 h, the level of T-AOC in 18.25 and 35.41 mmol/L groups showed a trend of first increasing and then decreasing (p < 0.05; Figure 3), and reached a peak at 24 h and 12 h, respectively. The level of T-AOC in 52.53 and 69.74 mmol/L groups showed the trend of volatility within the range of alkalinity stress for 96 h (Figure 3), and the level of T-AOC in both groups were significantly lower than that in control group at 12 h (p < 0.05). At 96 h, the level of T-AOC of the 18.25 and 35.41 mmol/L groups were significantly lower than that in the control group (p < 0.05), while the level of T-AOC in the 69.74 mmol/L group was significantly higher than control (p < 0.05).
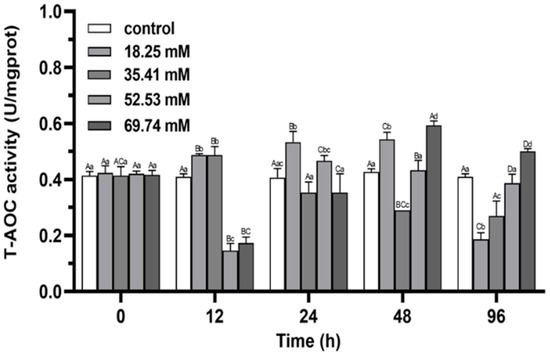
Figure 3.
Effects of alkalinity stress on T-AOC enzyme activity in the hepatopancreas of Eriocheir sinensis. Different capital letters indicate significant differences in the same alkalinity group (p < 0.05), and different lowercase letters indicate significant differences at the same time point (p < 0.05).
The activity of GSH in 35.41 mmol/L groups showed the trend of volatility in the range of alkalinity stress for 96 h, and the activity was induced to the peak level at 24 h (Figure 4). The GSH activities in the 18.25, 52.53 and 69.74 mmol/L groups showed a trend of first increasing and then decreasing (p < 0.05), and reached a peak at 48, 48 and 24 h, respectively. At 96 h of stress, GSH activity of 18.25 mmol/L group was close to that of control group (p > 0.05), while it was significantly higher than that in other groups in 35.41 mmol/L group (p < 0.05).
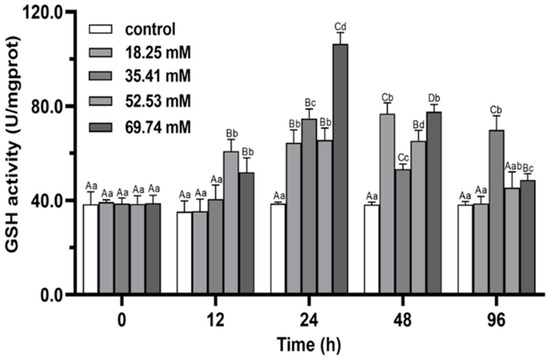
Figure 4.
Effects of alkalinity stress on GSH enzyme activity in the hepatopancreas of Eriocheir sinensis. Different capital letters indicate significant differences in the same alkalinity group (p < 0.05), and different lowercase letters indicate significant differences at the same time point (p < 0.05).
3.2. Immune Enzymes
During the whole stress process, the AST activity of serum in treatment first increased and then decreased, and all of them reached the peak level at 12 h (p < 0.05; Figure 5). The AST peak level in the 69.74 mmol/L group was significantly higher than that in the other groups (p < 0.05). At 96 h of alkalinity stress, the AST activity in 18.25 mmol/L group was significantly lower than control (p < 0.05), while it recovered to the initial level in 35.41 mmol/L group (p > 0.05). The AST activities of 52.53 and 69.74 mmol/L groups were significantly higher than control (p < 0.05).
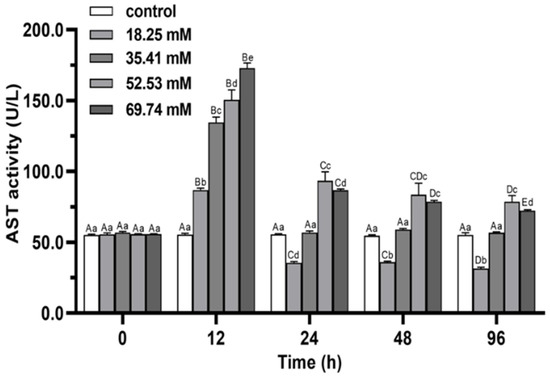
Figure 5.
Effects of alkalinity stress on AST enzyme activity in the hepatopancreas of Eriocheir sinensis. Different capital letters indicate significant differences in the same alkalinity group (p < 0.05), and different lowercase letters indicate significant differences at the same time point (p < 0.05).
The ALT activities in 18.25 and 52.53 mmol/L groups first increased and then decreased during 96 h of stress (p < 0.05), and reached the peak levels at 12 and 48 h, respectively (Figure 6). The peak level of ALT activity at 48 h in the 52.53 mmol/L group was significantly higher than that in the other groups (p < 0.05). The ALT activity in the 69.74 mmol/L group increased continuously during the whole experiment (p < 0.05). At 96 h, the ALT activity in 18.25 mmol/L group returned to the initial level (p > 0.05), while the ALT activities in the other three groups still significantly higher than that in the control group (p < 0.05). The serum ALT activity increased significantly with increasing alkalinity at 96 h (p < 0.05).
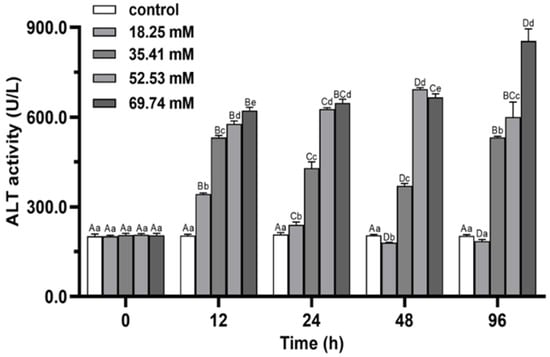
Figure 6.
Effects of alkalinity stress on ALT enzyme activity in the hepatopancreas of Eriocheir sinensis. Different capital letters indicate significant differences in the same alkalinity group (p < 0.05), and different lowercase letters indicate significant differences at the same time point (p < 0.05).
During 96 h of alkalinity stress, the activities of ACP and AKP in 18.25 and 35.41 mmol/L groups first increased and then decreased (p < 0.05; Figure 7 and Figure 8) and peaked at 12 h in both groups. ACP and AKP activities significantly induced in 52.53 mmol/L group (p < 0.05). The activity of ACP in the 69.74 mmol/L group showed a continuous increasing trend during the whole stress process (p < 0.05). At 96 h of stress, the activities of ACP and AKP in the 18.25 mmol/L group recovered to the initial level (p > 0.05), and the levels of AKP was not significant difference between the 35.41 mmol/L group and the control group (p > 0.05). The ACP and AKP activities of the 52.53 and 69.74 mmol/L concentration groups were significantly higher than control at 96 h of stress (p < 0.05).
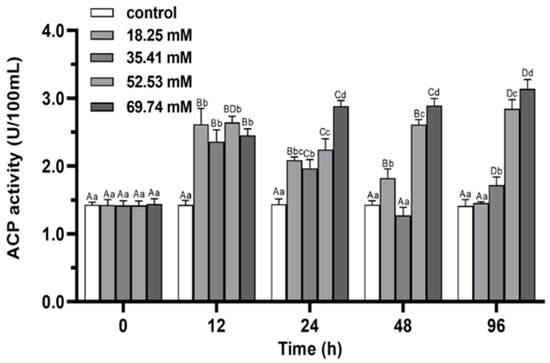
Figure 7.
Effects of alkalinity stress on ACP enzyme activity in the hepatopancreas of Eriocheir sinensis. Different capital letters indicate significant differences in the same alkalinity group (p < 0.05), and different lowercase letters indicate significant differences at the same time point (p < 0.05).
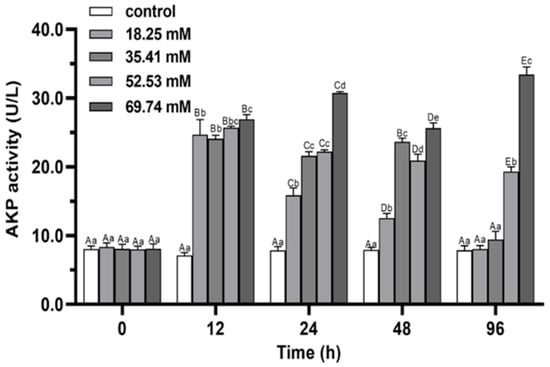
Figure 8.
Effects of alkalinity stress on AKP enzyme activity in the hepatopancreas of Eriocheir sinensis. Different capital letters indicate significant differences in the same alkalinity group (p < 0.05), and different lowercase letters indicate significant differences at the same time point (p < 0.05).
3.3. Hepatopancreatic Histological Observation
The hepatic tubule structure in the control group was shown in Figure 9A. In the 18.25 mmol/L group, the hepatic tubule structure was relatively normal, and part of the basal membrane was separated from the epithelial cells (Figure 9B). In the 35.41 mmol/L group, the cells showed vacuoles, part of the basal membrane was detached, the lumen and transport vesicles were enlarged, and the nuclei of some epithelial cells were disintegrated and contracted (Figure 9C). In the 52.53 mmol/L group, the lumen of hepatic tubules became larger, the number of vacuoles increased, some epithelial cells disintegrated, nuclear pyknosis, and part of basement membrane of hepatic tubules ruptured (Figure 9D). In the 69.74 mmol/L group, the number of vacuoles increased. Some hepatopancreas cells in the hepatopancreas duct were necrotic and disintegrated, and some of the basal membrane and epithelial cells fell off to form cavities or even ruptured, and the epithelial cells also fell off, and nuclear contraction occurred (Figure 9E).
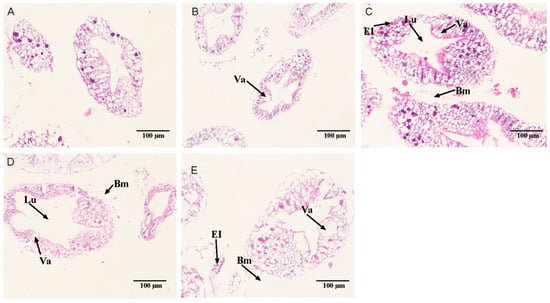
Figure 9.
Effect of alkalinity stress on the hepatopancreas tissue structure of Eriocheir sinensis. (A) Control; (B) 18. 25 mmol/L treatment group; (C) 35.41 mmol/L treatment group; (D) 52.53 mmol/L treatment group; (E) 69.74 mmol/L treatment group. The arrows point to lumen (Lu), the basement membrane (Bm), vacuole (Va) and Epithelial Cell (EI).
4. Discussion
4.1. Antioxidant Capacity and Non-Specific Immune Functions of E. sinensis under Alkalinity Stress
In crustaceans, reactive oxygen species (ROS) play an important role in cellular defense. However, excessive ROS can cause oxidative damage to cells and tissues, such as DNA, cell membranes, proteins and enzymes. The body’s antioxidant defense system is activated to prevent excessive ROS from causing oxidative damage to the body [22,23]. Antioxidant enzymes such as SOD, CAT, T-AOC can remove reactive oxygen species, enhance the body’s defense ability and remove toxins in antioxidant defense [24,25]. Free radicals in a normal crab are in a dynamic balance, and when the body is exposed to external stimuli, it will trigger a normal stress response, and its activity rises within a certain range [26]. However, the crustaceans’ antioxidant system has a baseline level of protection against oxidative stress and decreases after a certain intensity, which is a protective mechanism developed by the body [27,28,29]. The antioxidant enzyme system of white shrimp could be activated to adapt to the external environment under acute carbonate alkalinity stress [10]. A previous studies showed that the activity of antioxidant enzymes in E. sinensis after deltamethrin treatment increased to remove ROS, and reduced environmental stress [30]. In the present study, the activities of SOD, CAT in the hepatopancreas of E. sinensis were significantly increased within 24 h of alkalinity stress compared with the control, and the activities of CAT returned to the initial level after 96 h of low-concentration treatment, indicating that the body’s antioxidant enzyme system played a protective role in resisting environmental damage to the body when the body was subjected to alkalinity stress. In addition, the body gradually adapted to the water environment with alkalinities of 18.25 mmol/L and 35.41 mmol/L, which was the result of E. sinensis adjusting their physiological state to adapt to the external environment, as similar to the research from Zhang et al. [29].
In the present study, after 96 h of alkalinity treatment, the activities of SOD and CAT in high alkalinity stress group were significantly lower than those in the control, while the level of T-AOC was significantly higher than that in the control, which was similar to the results that the regulation of melatonin on hepatopancreas antioxidant enzymes in E. sinensis [31]. When an organism suffered mild stress, the organism responded accordingly to adapt to environmental changes, and the enzyme activity tended to increase [32]. The activity of antioxidant enzymes in the high alkalinity stress was significantly inhibited, indicating the crab might decompose and remove the reactive oxygen species generated in the metabolic process of the body to resist the pressure of the external environment. However, due to the fact that long-term high alkalinity exposure resulted in excessive accumulation of ROS or H2O2 in the crab, the endogenous antioxidant defense system could not completely counteract them. Eventually, SOD was further inactivated. In addition to CAT, GSH can also prevent ROS production by neutralizing H2O2 [33]. Enhanced GSH activity would help clear accumulated H2O2 during the response to stress [34]. In this study, the activity of GSH in the alkalinity exposure group was significantly higher than that in the control group. Therefore, the change of CAT level in this study may be related to the compensatory change of GSH activity.
Some nonspecific immune enzymes in E. sinensis could reflect its adaptability to different alkalinity stresses. AST and ALT are important amino acid transaminases that widely exist in animal cytoplasm and mitochondria. They play an important role in the protein metabolism of crustaceans and are sensitive indicators of immune function [35]. Impaired AST and ALT activity is directly related to the extent of damage caused by toxic compounds [36]. In this study, AST and ALT in the serum of the 18.25 mmol/L treatment group increased first and then gradually decreased and returned to the initial level during the stress process. This is consistent with consistent with previous studies [25,31]. In the low concentration group, the activity of AST and ALT increased briefly under stress so that the body could adapt to changes in the external environment. AST and ALT in the serum of crustaceans mainly come from the hepatopancreas and are sensitive inducers of hepatocyte injury induced by oxidative stress of exogenous compounds [37]. The present study showed that the AST and ALT activities was significantly higher under high alkalinity stress all the time, indicating that the hepatopancreas of crab might be damaged, which was consistent with the results of tissue sections.
Phosphatase has a variety of physiological functions, such as immunomodulation and antioxidation, involved in protein and lipid metabolism [38]. Phosphatases (ACP and AKP) are involved in the protein and lipid metabolism, immune regulation and antioxidant functions [39], and cell damage is usually accompanied by an increase in phosphatase activity [40]. The results of this study showed that the activities of ACP and AKP first increased and then recovered to their initial levels under low alkalinity stress. ACP and AKP activities increased significantly during the experiment with high alkalinity and were significantly higher than those of the control group at 96 h. These results suggested that high alkalinity stress might disrupt the permeability and integrity of the hepatopancreatic cell membrane of the crab, then leading to impair immune function. It may cause disturbance of cell metabolism and immune function. These results also indicated that ACP and AKP were involved in the immune response to alkalinity exposure of E. sinensis. Similar results were obtained in a previous study dealing with agro-chemical stresses [31].
4.2. Effects of Alkalinity on the Hepatopancreas Structure of E. sinensis
The hepatopancreas is an important digestive and immune organ of E. sinensis, as well as other crustaceans. The histological changes of the hepatopancreas often reflect the degree of toxicity. In this study, the hepatopancreas structure of E. sinensis were distinctly damaged under four different alkalinity stresses, and the higher the alkalinity was, the more serious the hepatopancreas structure was damaged. The main pathological changes were the enlargement of cell vacuoles, the abscission and rupture of the basal membrane, the disintegration of epithelial cells and the enlargement of transport vacuoles. This is similar to the symptoms of E. sinensis under agro-chemical stresses [41,42]. It is speculated that hepatopancreas epithelial cell vacuoles may expel HCO3− and H+ from the body to achieve detoxification. The proportion of hepatopancreatic cells in crustaceans will undergo adaptive changes under external environmental stress [43]. The results of this study were consistent with the phenomenon that E. sinensis also showed increased vacuoles in B cells after ammonia nitrogen stress [27]. Studies have shown the presence of p-glycoprotein in the differentiation of F cells into B cells, which neutralize the toxicity of certain compounds [44]. B cells not only secrete but also digest and absorb nutrients. Under environmental stress, crustaceans reduce their activity level and use more stored nutrients to maintain basic physiological metabolism [29]. In this study, alkalinity stress caused E. sinensis to require more nutrients to maintain basic physiological metabolism, and the increase in the number and volume of transport vesicles inside B cells may contribute to the metabolism and absorption of nutrients in liver tubules and promote detoxification. However, when the alkalinity of carbonate exceeds the detoxification ability of hepatopancreas B cells, the structure of hepatopancreas cells will be abnormal, and even the whole structure of hepatopancreas cells will be destroyed.
5. Conclusions
The effects of acute alkalinity stress on the antioxidant and nonspecific immune systems, and hepatopancreas structure of E. sinensis were studied. The results showed that under low alkalinity (18.25 mmol/L) stress, E. sinensis could activate the antioxidant system and nonspecific immune system to resist the environment, and then the physiological indices returned to normal levels to realize adaptation to the environment. However, high alkalinity (69.74 mmol/L) stress can induce oxidative stress in the hepatopancreas of E. sinensis, thereby disrupting antioxidant capacity and immune-related responses. The results of this study are expected to provide a practical basis for the development of culture technology for E. sinensis extending to saline and alkaline waters under arid climate conditions in deep inland areas in China.
Author Contributions
Conceptualization, M.L., Z.Z. and Y.Y.; methodology, M.L.; software, M.L.; validation, M.L., Z.Z. and Y.Y.; formal analysis, S.W., L.L. and L.Z.; investigation, R.Z.; resources, K.G.; data curation, M.L., S.W. and K.G.; writing—original draft preparation, M.L.; writing—review and editing, M.L. and Z.Z.; visualization, M.L.; supervision, Z.Z. and Y.Y.; project administration, L.L.; funding acquisition, M.L., Z.Z. and Y.Y. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by National Key R&D Program of China (Grant No. 2020YFD0900402, 2019YFD0900404, 2020YFD0900304), Central Public-interest Scientific Institution Basal Research Fund, CAFS (Grant No. 2021XT05, 2019XT0502, 2020TD56).
Institutional Review Board Statement
The studies in Eriocheir sinensis were reviewed and approved by the Committee for the Welfare and Ethics of Laboratory Animals of Heilongjiang River Fisheries Research Institute (Approval numbers: 20210501-002, approved on 7 May 2021).
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Acknowledgments
We want to thank all the students in our team for their help on the collection of samples.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Sumner, M.E.; Naidu, R. Sodic Soils: Distribution, Properties, Management, and Environmental Consequences; Oxford University Press: New York, NY, USA, 1998; pp. 1–232. [Google Scholar]
- Shang, X.; Geng, L.; Yang, J.; Zhang, Y.; Xu, W. Transcriptome analysis reveals the mechanism of alkalinity exposure on spleen oxidative stress, inflammation and immune function of Luciobarbus capito. Ecotoxicol. Environ. Saf. 2021, 225, 112748. [Google Scholar] [CrossRef] [PubMed]
- Dang, Y.F.; Xu, W.; Geng, L.W.; Bai, Y.Y. A review of effects of saline-alkalinity and pH on growth and development in fish. Chin. J. Fish. 2012, 225, 62–64. [Google Scholar] [CrossRef]
- Wang, P.; Liu, J.Y.; Yao, Z.L.; Lai, Q.F.; Yang, P.H.; Wang, H.; Wu, G.X. Advances in saline-alkali adaptation physiology of aquatic animals. J. Yangtze Univ. (Nat. Sci. Ed.) 2015, 12, 44–47. [Google Scholar] [CrossRef]
- Galat, D.L.; Post, G.; Keefe, T.J.; Bouck, G.R. Histological changes in the gill, kidney and liver of Lahontan cutthroat trout, Salmo clarki henshawi, living in lakes of different salinity-alkalinity. J. Fish Biol. 2010, 27, 533–552. [Google Scholar] [CrossRef]
- Wang, H.; Fang, W.H.; Lai, Q.F. Effects of concentrations of Ca2+ and Mg2+ on survival and growth of Penaeus chinensis. J. Fish. Sci. China 2000, 7, 82–86. [Google Scholar] [CrossRef]
- Zhang, L.P.; Xia, J.; Hu, Z.F. Situation and problem analysis of water resources security in China. Resour. Environ. Yangtze Basin 2009, 18, 116–120. [Google Scholar]
- Ondrasek, G.; Rengel, Z. Environmental salinization processes: Detection, implications & solutions. Sci. Total Environ. 2021, 754, 142432. [Google Scholar] [CrossRef]
- Liang, C.F. Tolerance, Growth and Branchal mRNA Expression of Na+/HCO3− Cotransporter and CA of the First Selected Generation of Oreochromis Niloticus in Saline and Alksaline Water; Shanghai Ocean University: Shanghai, China, 2015; pp. 17–23. [Google Scholar]
- Liu, F.; Li, J.; Li, J.T.; Ge, Q.Q.; Ge, H.X.; Shen, M.M. Effects of carbonate alkalinity stress on the survival, growth, reproduction, and immune enzyme activities of Exopalaemon carinicauda. J. Fish. Sci. China 2016, 23, 1137–1147. [Google Scholar] [CrossRef]
- Yang, F.Y.; Li, X.J.; Zhao, C.S.; Chen, Y.; Yang, X.Q.; Sun, L.M. Factors influencing the growth of Litopenaeus vannamei in carbonate alkaline waters of northeast China. J. Hydroecol. 2007, 27, 42–46. [Google Scholar]
- Lin, T.T.; Lai, Q.F.; Yao, Z.L.; Lu, J.X.; Zhou, K.; Wang, H. Combined effects of carbonate alkalinity and pH on survival, growth and haemocyte parameters of the Venus clam Cyclina sinensis. Fish Shellfish Immunol. 2013, 35, 525–531. [Google Scholar] [CrossRef]
- Zeng, F.Y.; Luo, K.; Luan, S.; Cao, B.X.; Lu, X.; Tan, J. Analysis of growth and survival among different families of Litopenaeu vannamei in the chloride typed alkaline water. J. Fish. Sci. China 2018, 25, 308–315. [Google Scholar] [CrossRef]
- Zheng, W.G.; Zhang, Z.Q.; Zhang, M.Z. Study on tolerance of Carassius auratus Pengze fingerlings to salinity and alkalinity. J. Jimei Univ. (Nat. Sci.) 2004, 2, 127–130. [Google Scholar] [CrossRef]
- Qi, T.T.; Liu, J.; Zhao, P.S.; Ge, B.M.; Liu, Q.N.; Jiang, S.H.; Wang, Z.; Zhang, H.; Tang, B.; Ding, G.; et al. A novel modulation of physiological regulation in cultured Chinese mitten crab (Eriocheir japonica sinensis) in response to consistent salinity changes. Gene 2020, 756, 144914. [Google Scholar] [CrossRef]
- Wang, X.D.; Huang, Z.P.; Wang, C.L.; Qi, C.L.; Gu, Z.M.; Li, E.C.; Qin, J.G.; Chen, L.Q. A comparative study on growth and metabolism of Eriocheir sinensis juveniles under chronically low and high pH stress. Front. Physiol. 2020, 11, 885. [Google Scholar] [CrossRef]
- Qiang, J.; Duan, X.J.; Zhu, C.K.; He, J.; Bao, J.W.; Tao, Y.F.; Zhu, H.J.; Xu, P. Selenium-cultured potamogeton maackianus in the diet can alleviate oxidative stress and immune suppression in Chinese mitten crab (Eriocheir sinensis) under copper exposure. Front. Physiol. 2020, 11, 713. [Google Scholar] [CrossRef]
- Zhao, X.J.; Yang, Z.G.; Cheng, Y.X. Effects of cadmium alone and in combination with pH on bioaccumulation, tissue structure, and enzyme activity of the Chinese mitten crab, Eriocheir sinensis. Comp. Biochem. Physiol. C 2021, 245, 109025. [Google Scholar] [CrossRef]
- Wang, T.Y.; Yang, C.; Zhang, T.T.; Liang, H.L.; Ma, Y.C.; Wu, Z.X.; Sun, W.T. Immune defense, detoxification, and metabolic changes in juvenile Eriocheir sinensis exposed to acute ammonia. Aquat. Toxicol. 2021, 240, 105989. [Google Scholar] [CrossRef]
- Yang, Y.H.; Li, M.S.; Luo, L.; Wang, S.H.; Zhang, R.; Guo, K.; Liu, J.Y.; Li, H.T.; Zhao, Z.G. Study on toxicity of salinity and alkalinity on Eriocheir sinensis. J. Northeast Agric. Univ. 2022, 53, 36–41. [Google Scholar] [CrossRef]
- Wang, H.; Lai, Q.; Fang, W.; Wang, J. Study on Aquaculture in Different Type of Saline Alkali Water. In Proceedings of the Forum on Fishery Science and Technology, Guangzhou, China, 22–25 September 2003; p. 303. [Google Scholar]
- Athanasios, V.; Thomais, V.; Manos, D.; Michael, S. Molecular biomarkersof oxidative stress in aquatic organisms in relation to toxic environmental pollutants. Ecotoxicol. Environ. Saf. 2006, 64, 178–189. [Google Scholar] [CrossRef]
- Wu, Y.S.; Lee, M.C.; Huang, C.T.; Kung, T.C.; Huang, C.Y.; Nan, F.H. Effects of traditional medical herbs “minor bupleurum decoction” on the non-specific immune responses of white shrimp (Litopenaeus vannamei). Fish Shellfish Immunol. 2017, 64, 218–225. [Google Scholar] [CrossRef]
- Li, Y.J.; Chai, X.; Wu, H.; Jing, W.X.; Wang, L. The response of metallothionein and malondialdehyde after exclusive and combined Cd/Zn exposure in the crab Sinopotamon henanense. PLoS ONE 2013, 8, e80475. [Google Scholar] [CrossRef] [PubMed]
- Fu, L.L.; Zhou, G.; Pan, J.L.; Li, Y.H.; Lu, Q.P.; Zhou, J.; Li, X.G. Effects of Astragalus polysaccharides on antioxidant abilities and non-specific immune responses of Chinese mitten crab, Eriocheir sinensis. Aquac. Int. 2017, 25, 1333–1343. [Google Scholar] [CrossRef]
- Zhang, Y.; Sun, K.; Li, Z.Y.; Chai, X.X.; Fu, X.Y.; Kholodkevich, S.; Kuznetsova, T.; Chen, C.; Ren, N.Q. Effescts of acute diclofenac exposure on intestinal histology, antioxidant defense, and microbiota in freshwater crayfish (Procambarus clarkii). Chemosphere 2021, 263, 128130. [Google Scholar] [CrossRef] [PubMed]
- Hong, M.L.; Chen, L.Q.; Gu, S.Z.; Liu, C.; Long, Z.Q.; Zhang, W. Effects of ammonia exposure on immunity indicators of haemolymph a histological structure of hepatopancreas in Chinese mitten crab (Eriocheir sinensis). J. Fish. Sci. China 2007, 3, 412–418. [Google Scholar] [CrossRef]
- Castex, M.; Lemaire, P.; Wabete, N.; Chim, L. Effect of dietary probiotic Pediococcus acidilacticion antioxidant defences and oxidative stress status of shrimp Litopenaeus stylirostris. Aquaculture 2009, 294, 306–313. [Google Scholar] [CrossRef]
- Zhang, F.; Wei, J.J.; Li, Q.; Jiang, R.L.; Yu, N.; Qin, J.G.; Chen, L.Q. Effects of perfluorooctane sulfonate on the immune responses and expression of immune-related genes in Chinese mitten-handed crab Eriocheir sinensis. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2015, 172–173, 13–18. [Google Scholar] [CrossRef]
- Hong, Y.H.; Huang, Y.; Yan, G.G.; Pan, C.; Zhang, J.L. Antioxidative status, immunological responses, and heat shock protein expression in hepatopancreas of Chinese mitten crab, Eriocheir sinensis under the exposure of glyphosate. Fish Shellfish Immunol. 2019, 86, 840–845. [Google Scholar] [CrossRef]
- Zhang, C.; Zhang, Q.; Pang, Y.Y.; Song, X.Z.; Zhou, N.; Wang, J.; He, L.; Lv, J.H.; Song, Y.M.; Cheng, Y.X.; et al. The protective effects of melatonin on oxidative damage and the immune system of the Chinese mitten crab (Eriocheir sinensis) exposed to deltamethrin. Sci. Total Environ. 2019, 653, 1426–1434. [Google Scholar] [CrossRef]
- Hong, Y.H.; Huang, Y.; Yan, G.W.; Huang, Z.Q. Effects of deltamethrin on the antioxidant defense and heat shock protein expression in Chinese mitten crab, Eriocheir sinensis. Environ. Toxicol. Pharmacol. 2019, 66, 1–6. [Google Scholar] [CrossRef]
- Noguchi, N.; Watanabe, A.; Shi, H. Diverse functions of antioxidants. Free Radic. Res. 2000, 33, 809–817. [Google Scholar] [CrossRef]
- Bagnyukova, T.V.; Storey, K.B.; Lushchak, V.I. Adaptive response of antioxidant enzymes to catalase inhibition by aminotriazole in goldfifish liver and kidney. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2005, 142, 335–341. [Google Scholar] [CrossRef]
- Pan, L.Q.; Wu, Z.W.; Zhang, H.X. Effects of heavy metal ions on transaminases activities of Litopenaeu vannamei. J. Ocean Univ. China 2005, 35, 195–198. [Google Scholar] [CrossRef]
- Dey, S.; Samanta, P.; Pal, S.; Mukherjee, A.K.; Kole, D.; Ghosh, A.R. Integrative assessment of biomarker responses in teleostean fishes exposed to glyphosate-based herbicide (Excel Mera 71). Emerg. Contam. 2016, 2, 191–203. [Google Scholar] [CrossRef]
- Qin, Q.; Qin, S.J.; Wang, L.; Lei, W.W. Immune responses and ultrastructural changes of hemocytes in freshwater crab Sinopotamon henanense exposed to elevated cadmium. Aquat. Toxicol. 2012, 106, 140–146. [Google Scholar] [CrossRef]
- Ajima, M.N.O.; Kumar, K.; Poojary, N.; Pandey, P.K. Oxidative stress biomarkers, biochemical responses and Na+-K+-ATPase activities in Nile tilapia, Oreochromis niloticus exposed to diclofenac. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2021, 240, 108934. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Cheng, Z.Z. Effects of the anesthetic MS-222 on the AKP, CAT and ACP activities in goldfish. J. Shanghai Ocean Univ. 2010, 19, 327–332. [Google Scholar]
- Zhao, L.L.; Yang, X.Z.; Cheng, Y.X.; Liang, P.; Zhang, J.B.; Hong, Y.H.; Wang, C.; Yang, Z. Effects of Histamine on Survival and Immune Parameters of the Chinese mitten crab, Eriocheir sinensis. J. Shellfish Res. 2012, 31, 827–834. [Google Scholar] [CrossRef]
- Yang, Z.Y.; Yang, Y.B.; Zhang, Y.L.; Hu, K.; Zeng, L.G.; Liu, L.S.; Yan, Z.J.; Yang, X.L.; Chang, O.Q. Effects of avermectin exposure on oxidative stress and histological structure of hepatopancreas in Eriocheir sinensis. Asian J. Ecotoxicol. 2017, 12, 337–347. [Google Scholar] [CrossRef]
- Zhang, X.J.; Lu, H.D.; Tian, Q.Q.; Jia, X.X.; Ren, F.F. Effects of deltamethrin on histopathology of Chinese mitten crab Eriocheir sinensis. Asian J. Ecotoxicol. 2018, 13, 342–351. [Google Scholar] [CrossRef]
- Tao, Y.F.; Qiang, J.; Wang, H. Acute toxicity of low-pH stress and its effect on enzyme activity and histological structure of gill and hepatopancreas in Procambarus clarkia. J. Fish. Sci. China 2016, 23, 1279–1289. [Google Scholar] [CrossRef]
- Köhler, A.; Lauritzen, B.; Jansen, D.; Böttcher, P.; Tegoliwa, L.; Krüner, G.; Broeg, K. Detection of P-Glycoprotein Mediated MDR/MXR in Carcinus Maenas Hepatopancreas by Immuno-Gold-Silver Labeling. In Proceedings of the Agu Fall Meeting, AGU Fall Meeting Abstracts, San Francisco, CA, USA, 6–10 December 1998; Volume 387, pp. 175–180. [Google Scholar]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).