
Comparative Analysis of Macro/Microstructures and Constituents of Sorghum and Reed Straw



Abstract
1. Introduction
2. Materials and Methods
2.1. Samples
2.2. Observation Positions
2.3. Stereomicroscopy
2.4. Scanning Electron Microscopy (SEM) and Energy Dispersive Spectrometer (EDS)
2.5. Infrared Spectra Analysis
3. Results and Discussion
3.1. Comparison of Macrostructures
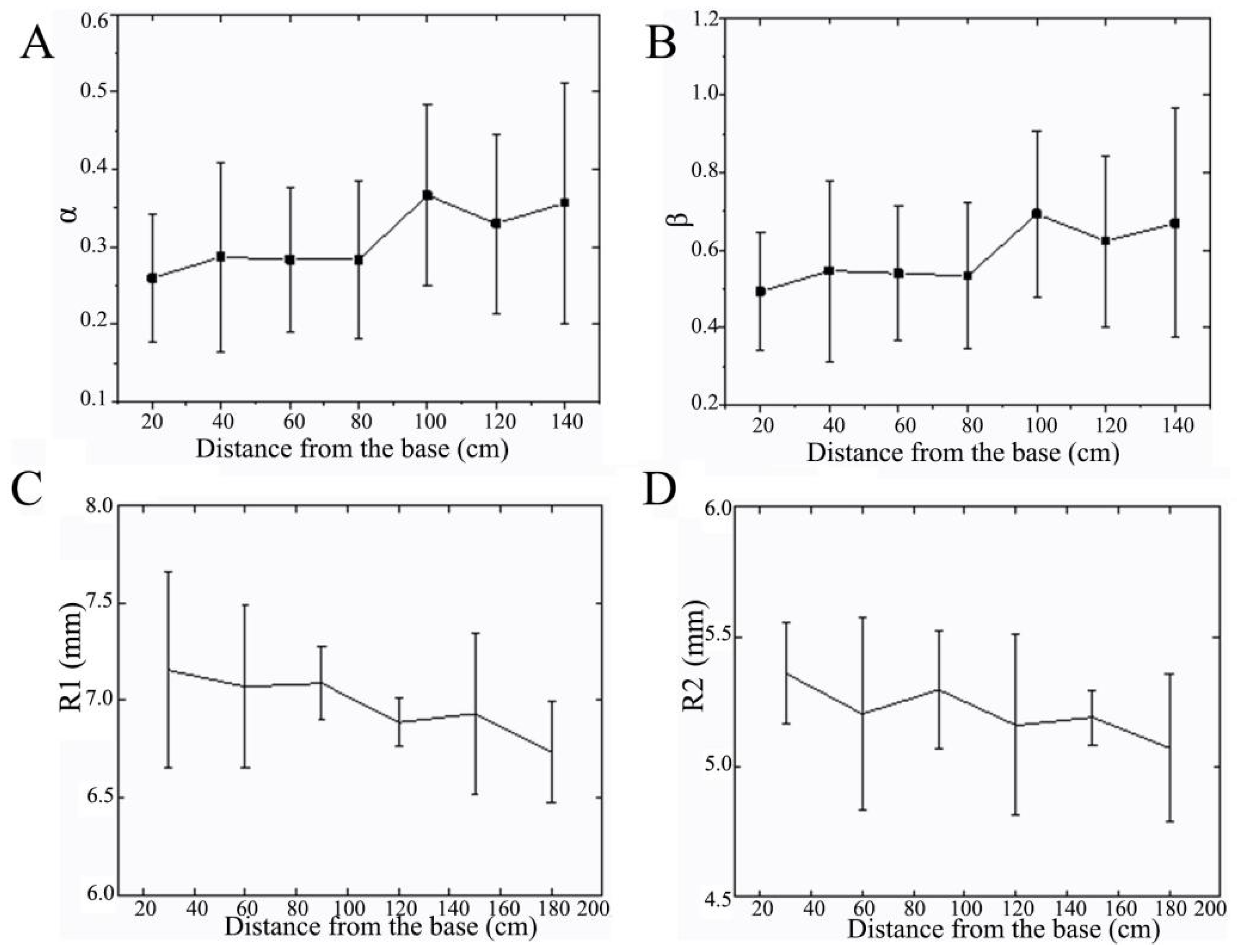
3.1.1. Changing Measurements of Diameter
3.1.2. Wall Thickness Changes
3.1.3. Changing Measurements of Internode Spacing
3.1.4. Cross-Sectional Properties
3.2. Comparison of Microstructures
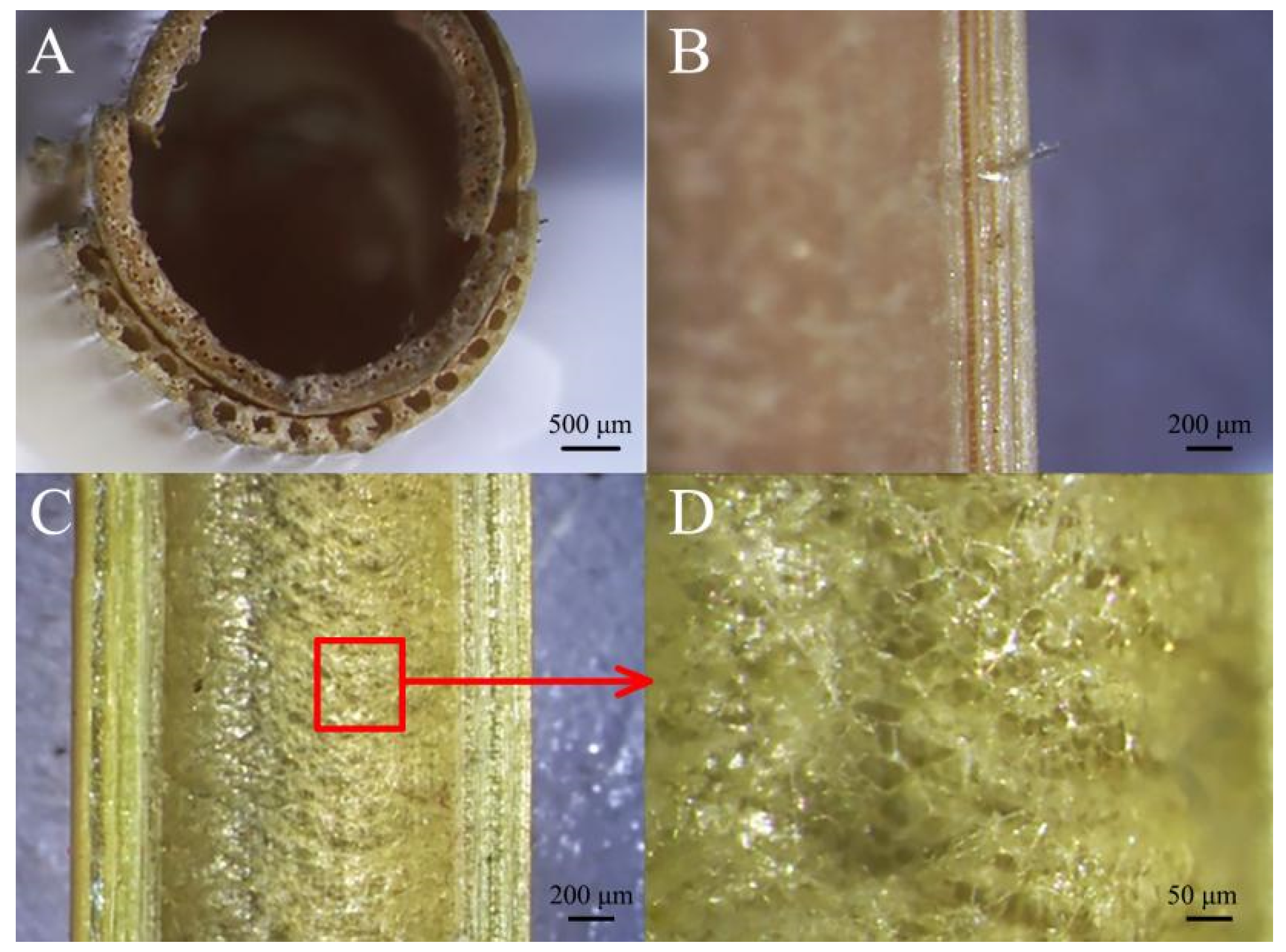
3.2.1. Structural Analysis under a Microscope
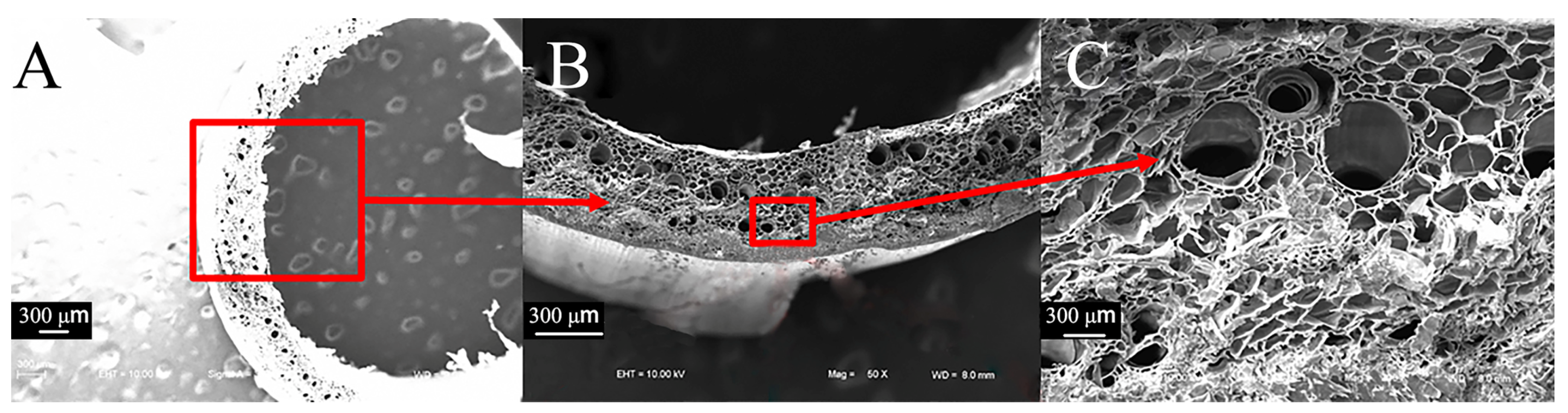
3.2.2. Comparison of Microscopic Transverse Cross-Sections
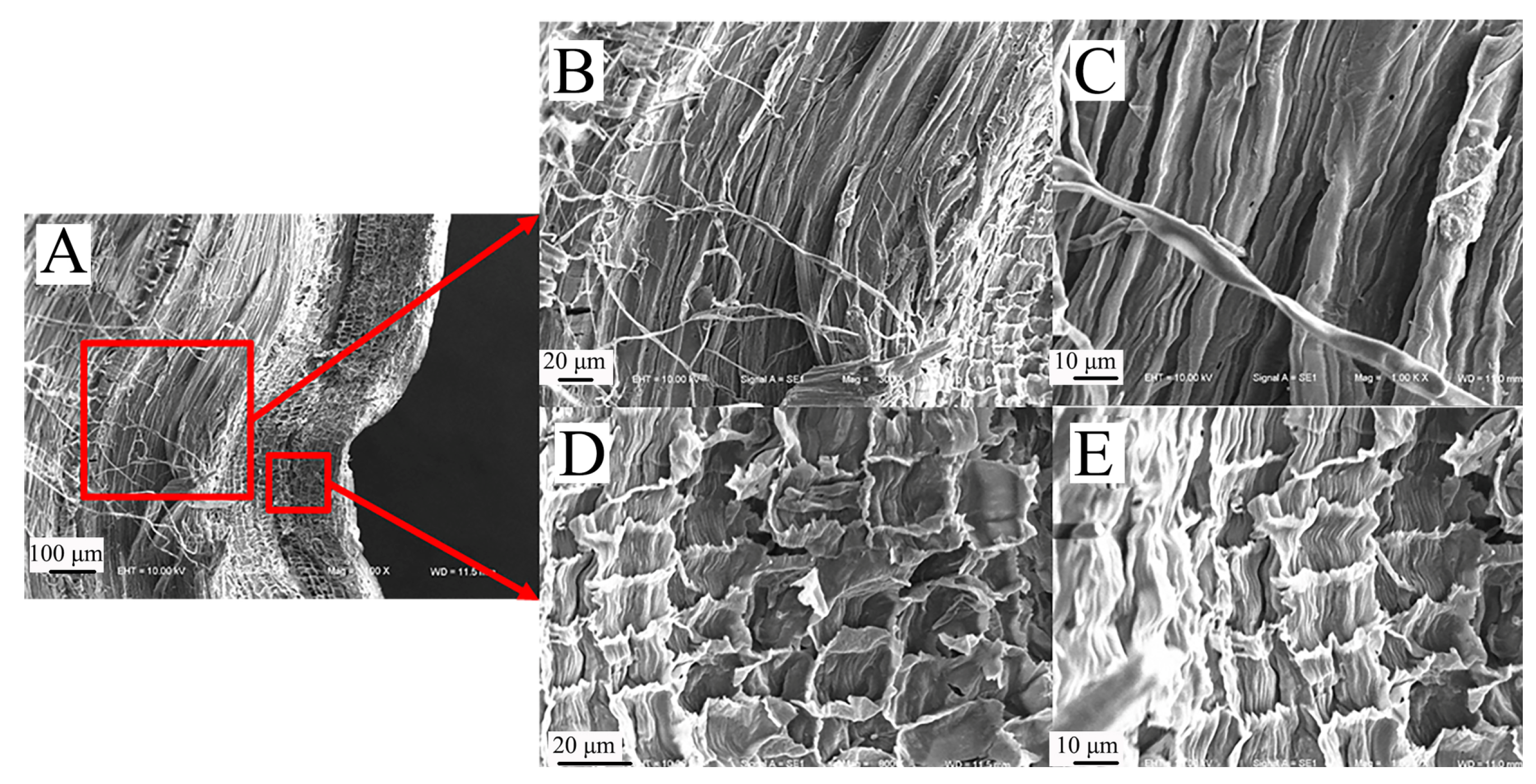
3.2.3. Comparison of Microscopic Longitudinal Cross-Sections
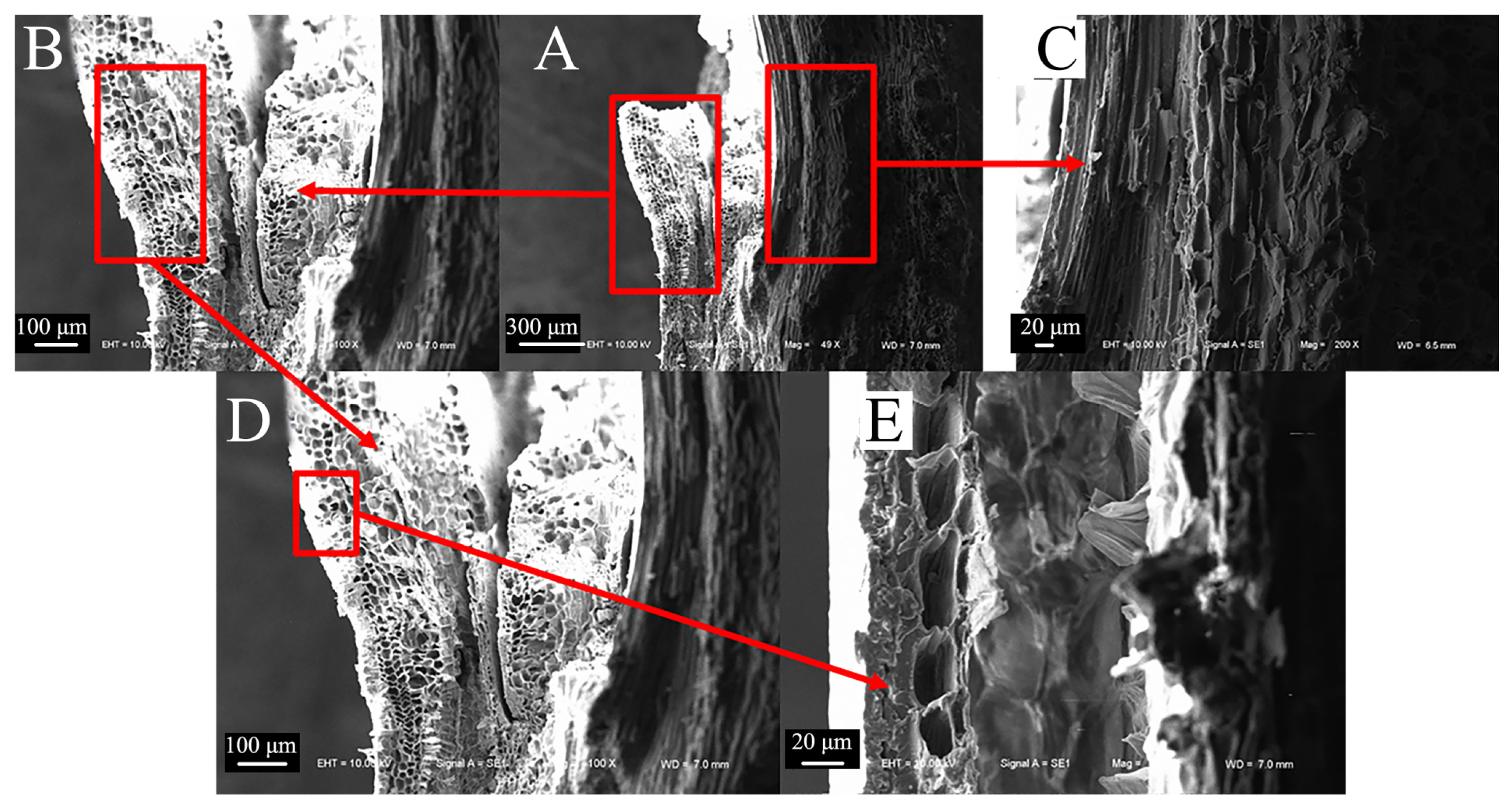
3.2.4. Comparison of Microscopic Node Characteristics
3.3. Comparison of Chemical Composition and Functional Groups
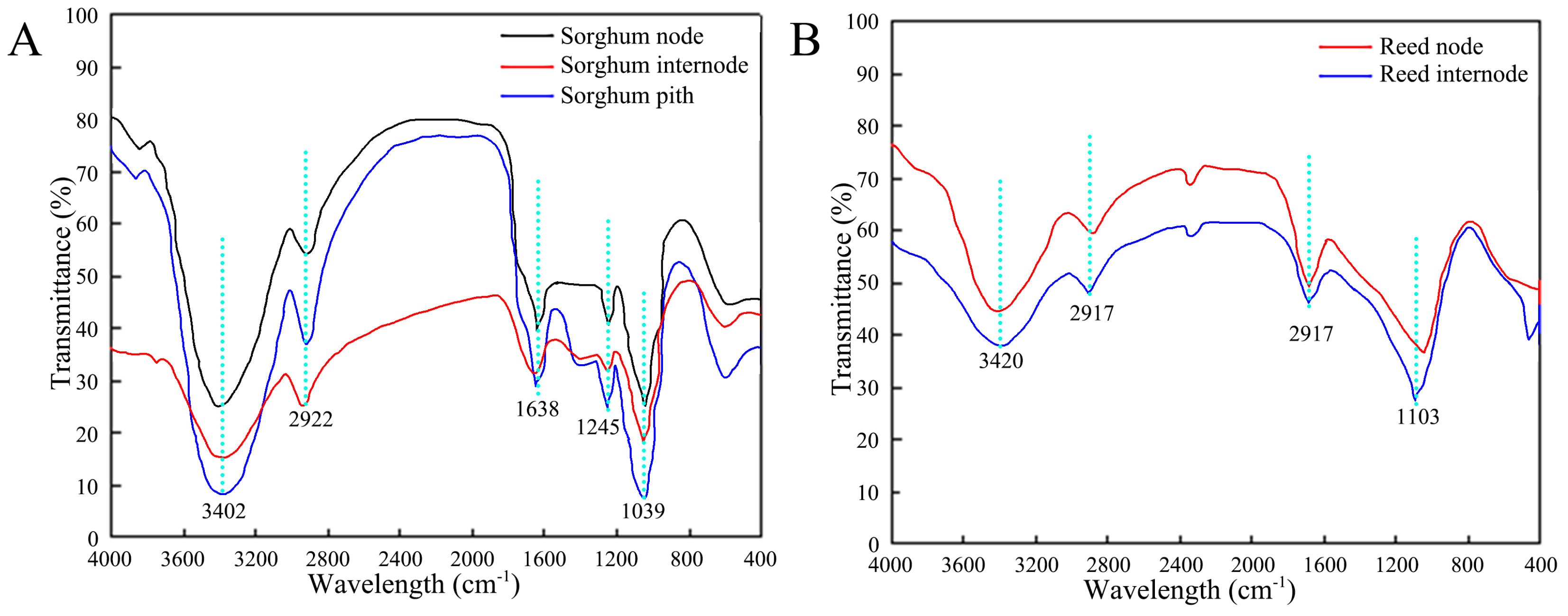
3.3.1. Functional Groups
3.3.2. Element Composition
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Dong, S.L. The Revelation of the Jellyfish, the Sea’s Thousand-Mile Ear. Reading 2022, 86, 39–41. [Google Scholar]
- Qin, Y.; Wang, H. Prototype, Diversity, and Mixture: The Experience of Commercial Complex Development in Singapore. New Archit. 2023, 4, 76–81. [Google Scholar]
- Chen, H.R. “Bird’s Nest” throws off the support and raises its “spine”. China Economic Herald (Newspaper), 21 September 2006; p. C01. [Google Scholar]
- Melati, R.B.; Sass, D.C.; Pagnocca, F.C.; Brienzo, M. Anatomic influence of sugarcane biomass on xylan solubilization. Ind. Crops Prod. 2021, 164, 113357. [Google Scholar] [CrossRef]
- Wang, X.; Shi, Y.; Wang, Q.; Li, Y. Dorsoventral regulation of photosynthetic functions related to morphological and anatomical structures in field-grown sorghum. Photosynthetica 2019, 57, 607–616. [Google Scholar] [CrossRef]
- Zou, M.; Xu, S.; Wei, C.; Wang, H.; Liu, Z. A bionic method for the crashworthiness design of thin-walled structures inspired by bamboo. Thin-Walled Struct. 2016, 101, 222–230. [Google Scholar] [CrossRef]
- Marinho, N.P.; Nascimento, E.M.; Nisgoski, S.; Valarelli, I.D. Some physical and mechanical properties of medium-density fiberboard made from giant bamboo. Mater. Res. 2013, 16, 1387–1392. [Google Scholar] [CrossRef]
- Hirai, Y.; Inoue, E.; Mori, K.; Hashiguchi, K. PM—Power and Machinery: Investigation of Mechanical Interaction between a Combine Harvester Reel and Crop Stalks. Biosyst. Eng. 2002, 83, 307–317. [Google Scholar] [CrossRef]
- Crook, M.J.; Ennos, A. The mechanics of root lodging in winter wheat, Triticum aestivum L. J. Exp. Bot. 1993, 44, 1219–1224. [Google Scholar] [CrossRef]
- Crook, M.; Ennos, A. Stem and root characteristics associated with lodging resistance in four winter wheat cultivars. J. Agric. Sci. 1994, 123, 167–174. [Google Scholar] [CrossRef]
- Skubisz, G. Development of studies on the mechanical properties of winter rape stems. Int. Agrophys. 2001, 15, 197–200. [Google Scholar]
- Jiao, Y.; Li, X.; Liang, L.; Takeuchi, K.; Okuro, T.; Zhang, D.; Sun, L. Indigenous ecological knowledge and natural resource management in the cultural landscape of China’s Hani Terraces. Ecol. Res. 2012, 27, 247–263. [Google Scholar] [CrossRef]
- Mangieri, M.A.; Mantese, A.I.; Schürmann, A.A.; Chimenti, C.A. Effects of ethephon on anatomical changes in sunflower (Helianthus annuus L.) stems associated with lodging. Crop Pasture Sci. 2016, 67, 988–999. [Google Scholar] [CrossRef]
- Liu, J.; Zhao, D.; Zhao, J. Study of the cutting mechanism of oil tree peony stem. Forests 2020, 11, 760. [Google Scholar] [CrossRef]
- Kong, E.; Liu, D.; Guo, X.; Yang, W.; Sun, J.; Li, X.; Zhan, K.; Cui, D.; Lin, J.; Zhang, A. Anatomical and chemical characteristics associated with lodging resistance in wheat. Crop J. 2013, 1, 43–49. [Google Scholar] [CrossRef]
- Shah, L.; Yahya, M.; Shah, S.M.A.; Nadeem, M.; Ali, A.; Ali, A.; Wang, J.; Riaz, M.W.; Rehman, S.; Wu, W. Improving lodging resistance: Using wheat and rice as classical examples. Int. J. Mol. Sci. 2019, 20, 4211. [Google Scholar] [CrossRef]
- Salter, B.; Bonnett, G.; Lawn, R. Morphology of young sugarcane stalks produced at different stages of crop development. Aust. J. Agric. Res. 2008, 59, 149–156. [Google Scholar] [CrossRef]
- Jung, H.-J.G. Maize stem tissues: Ferulate deposition in developing internode cell walls. Phytochemistry 2003, 63, 543–549. [Google Scholar] [CrossRef]
- Genet, M.; Stokes, A.; Salin, F.; Mickovski, S.B.; Fourcaud, T.; Dumail, J.-F.; van Beek, R. The influence of cellulose content on tensile strength in tree roots. Plant Soil 2005, 278, 1–9. [Google Scholar] [CrossRef]
- Zhang, R.; Jia, Z.; Ma, X.; Ma, H.; Zhao, Y. Characterising the morphological characters and carbohydrate metabolism of oat culms and their association with lodging resistance. Plant Biol. 2020, 22, 267–276. [Google Scholar] [CrossRef]
- Yin, H.; Xiao, Y.; Wen, G.; Qing, Q.; Wu, X. Crushing analysis and multi-objective optimization design for bionic thin-walled structure. Mater. Des. 2015, 87, 825–834. [Google Scholar] [CrossRef]
- Liu, Q.; Ma, J.; He, Z.; Hu, Z.; Hui, D. Energy absorption of bio-inspired multi-cell CFRP and aluminum square tubes. Compos. B Eng. 2017, 121, 134–144. [Google Scholar] [CrossRef]
- Tasdemirci, A.; Akbulut, E.F.; Guzel, E.; Tuzgel, F.; Yucesoy, A.; Sahin, S.; Guden, M. Crushing behavior and energy absorption performance of a bio-inspired metallic structure: Experimental and numerical study. Thin Wall. Struct. 2018, 131, 547–555. [Google Scholar] [CrossRef]
- Tabarsa, T.; Chui, Y.H. Characterizing microscopic behavior of wood under transverse compression. Part II. Effect of species and loading direction. Wood Fiber Sci. 2001, 223–232. [Google Scholar] [CrossRef]
- Kronbergs, E. Mechanical strength testing of stalk materials and compacting energy evaluation. Ind. Crops Prod. 2000, 11, 211–216. [Google Scholar] [CrossRef]
- Zhao, C.; Wu, J.; Cheng, Z.; Shi, S. Research on mechanical properties and chemical compositions of bottom stems of medicago sativa in harvesting period. In Proceedings of the International Conference on Information Science and Technology, Nanjing, China, 26–28 March 2011. [Google Scholar]
- Mackinnon, B.; Easton, H.; Barry, T.; Sedcole, J. The effect of reduced leaf shear strength on the nutritive value of perennial ryegrass. J. Agric. Sci. 1988, 111, 469–474. [Google Scholar] [CrossRef]
- Robertson, D.J.; Julias, M.; Gardunia, B.W.; Barten, T.; Cook, D.D. Corn stalk lodging: A forensic engineering approach provides insights into failure patterns and mechanisms. Crop Sci. 2015, 55, 2833–2841. [Google Scholar] [CrossRef]
- Shah, D.U.; Reynolds, T.P.; Ramage, M.H. The strength of plants: Theory and experimental methods to measure the mechanical properties of stems. J. Exp. Bot. 2017, 68, 4497–4516. [Google Scholar] [CrossRef]
- Yue, J.Z.; Jing, Y.Y.; Zhang, Z.P.; Zhang, Q.G. Effect of Particle Size on the Water-Soluble Substances and Microscopic Structure of Sorghum Straw Powder. Adv. Mater. Res. 2012, 512–515, 384–387. [Google Scholar] [CrossRef]
- Pardales, J.J.; Kono, Y.; Yamauchi, A. Epidermal cell elongation in sorghum seminal roots exposed to high root-zone temperature. Plant Sci. 1992, 81, 143–146. [Google Scholar] [CrossRef]
- Brugnerotto, J.; Lizardi, J.; Goycoolea, F.; Argüelles-Monal, W.; Desbrieres, J.; Rinaudo, M. An infrared investigation in relation with chitin and chitosan characterization. Polymer 2001, 42, 3569–3580. [Google Scholar] [CrossRef]
- Krimm, S.; Bandekar, J. Vibrational spectroscopy and conformation of peptides, polypeptides, and proteins. Adv. Protein Chem. 1986, 38, 181–364. [Google Scholar] [PubMed]
- Zdarta, J.; Klapiszewski, Ł.; Wysokowski, M.; Norman, M.; Kołodziejczak-Radzimska, A.; Moszyński, D.; Ehrlich, H.; Maciejewski, H.; Stelling, A.L.; Jesionowski, T. Chitin-lignin material as a novel matrix for enzyme immobilization. Mar. Drugs 2015, 13, 2424–2446. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Element | C | O | Cl | K | Ca | Mg | Br |
|---|---|---|---|---|---|---|---|
| Fiber bundles of sorghum | 53 | 45 | 1 | 1 | 0 | 0 | 0 |
| Basic tissues of sorghum | 50 | 46 | 1 | 1 | 1 | 1 | 0 |
| Mechanical tissues of reed | 51 | 49 | 0 | 0 | 0 | 0 | 0 |
| Thin-walled tissues of reed | 51 | 48 | 0 | 0 | 0 | 0 | 1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Song, J.; Li, G.; Liu, Y.; Zou, M. Comparative Analysis of Macro/Microstructures and Constituents of Sorghum and Reed Straw. Biomimetics 2024, 9, 107. https://doi.org/10.3390/biomimetics9020107
Song J, Li G, Liu Y, Zou M. Comparative Analysis of Macro/Microstructures and Constituents of Sorghum and Reed Straw. Biomimetics. 2024; 9(2):107. https://doi.org/10.3390/biomimetics9020107
Chicago/Turabian StyleSong, Jiafeng, Guoyu Li, Yansong Liu, and Meng Zou. 2024. "Comparative Analysis of Macro/Microstructures and Constituents of Sorghum and Reed Straw" Biomimetics 9, no. 2: 107. https://doi.org/10.3390/biomimetics9020107
APA StyleSong, J., Li, G., Liu, Y., & Zou, M. (2024). Comparative Analysis of Macro/Microstructures and Constituents of Sorghum and Reed Straw. Biomimetics, 9(2), 107. https://doi.org/10.3390/biomimetics9020107