1. Introduction
The biology of elasmobranchs, including sharks, have intrigued marine biologists and paleontologists for several hundred years [
1,
2]. One of the intriguing points is the appearance of so-called placoid scales, or dermal denticles, in the skin of the sharks. Since the pioneering work of Wolf-Ernst Reif initiated in the 1970s, summarized e.g., in [
3,
4], the putative drag-reducing effects of these dermal denticles of the fastest-swimming sharks such as
Lamniformes have triggered engineers to produce surfaces with structural elements that although enlarged in scale, were inspired by the shapes of these dermal denticles [
5,
6]. This putative drag-reducing function is seen in the fastest-swimming sharks and is supposedly optimized for their maximum swimming speed, obtained during bursting motion [
3]. It is probably the most sought-after function in biomimetic engineering work inspired by shark skin. The putative drag-reducing effect of some of these engineered structures has, however, been shown to be limited [
7] and for some conditions even drag increasing [
7]. In recent studies, surfaces that were replicas of the natural structures were investigated [
8,
9], and discussions of the design include the importance of the flexibility of the substrate in which the dermal denticles reside [
8,
10] and the ability of the dermal denticles to bristle [
11,
12,
13,
14]. Also, the spacing and modifications of the patterning of the denticle-like structures are a topic of investigations related to the drag-reducing properties of the structured surface [
10,
15,
16].
According to the original investigations of dermal denticles and resulting conjectures by Reif [
3,
17], the structure and role of dermal denticles is, however, richer than that of a drag-reducing surface. The paleontological investigations of Reif point to division of sharks into four groups, according to the putative function of their dermal denticles, as summarized in Figure 18 in [
3]. In addition to drag reduction, dermal denticles of other groups of sharks are thus suggested to provide abrasion strength, defense, and luminescence. The latter is actually not achieved by the dermal denticles themselves but rather by photophores in other parts of the skin of the shark, and recent literature [
18] suggests excluding the luminescent function and thus reduce the number of groups to three.
Dermal denticles believed to provide abrasion strength are postulated mainly for demersal species of sharks living near rocky surfaces whereas both abrasion strength and defense against ectoparasites are suggested for demersal species living in sandy or muddy areas [
3]. Finally, the dermal denticles of slowly moving sharks living in open water are suggested to provide defense against ectoparasites [
3]. It is unclear how defense should be understood in this context, and one may speculate of a relation to anti-fouling. Anti-fouling properties of direct replicas of dermal denticles from slowly moving sharks have been investigated [
19], but the dermal denticle structures of fast-moving sharks have also inspired the creation of bioengineered surfaces that have been demonstrated to reduce fouling of small bacteria [
20]. The structures of dermal denticles of sharks add to other natural structures in the quest for microstructured surfaces that would reduce fouling [
20,
21,
22,
23]. Naturally, the anti-fouling properties depend on flow conditions and the full understanding of the phenomenon is as yet unclear [
19]. Thus, it appears that there is more to the story than the original hypotheses put forward by Reif while grouping species of sharks into four [
3].
Our conjecture is that the nature of the flow pattern at length scales ranging from well below the size of the dermal denticle to several times the denticle size plays a role for the function of the dermal denticles. Thus, visualization of the flow surrounding the dermal denticles is relevant and interesting, and to perform this visualization, the method of micro-PIV is applied, to complement investigations in the literature visualizing the flow at larger scale using otherwise comparable technology [
7,
14,
24,
25]. In Particle Image Velocimetry (PIV), flow velocity fields are measured by imaging tracer particles in two consecutive photos taken in a short time. Velocities can be measured based on the correlation extracted from the image pair [
26]. Micro-PIV is the microfluidic version [
27] using microspheres and imaging through microscope optics. More elaborate techniques like stereoscopic micro-PIV or tomographic micro-PIV have been developed, see e.g., [
28], but for these first reported micro-PIV measurements near natural shark skin samples, we apply the originally developed micro-PIV technology as commercially available, and discussed further in
Section 5.
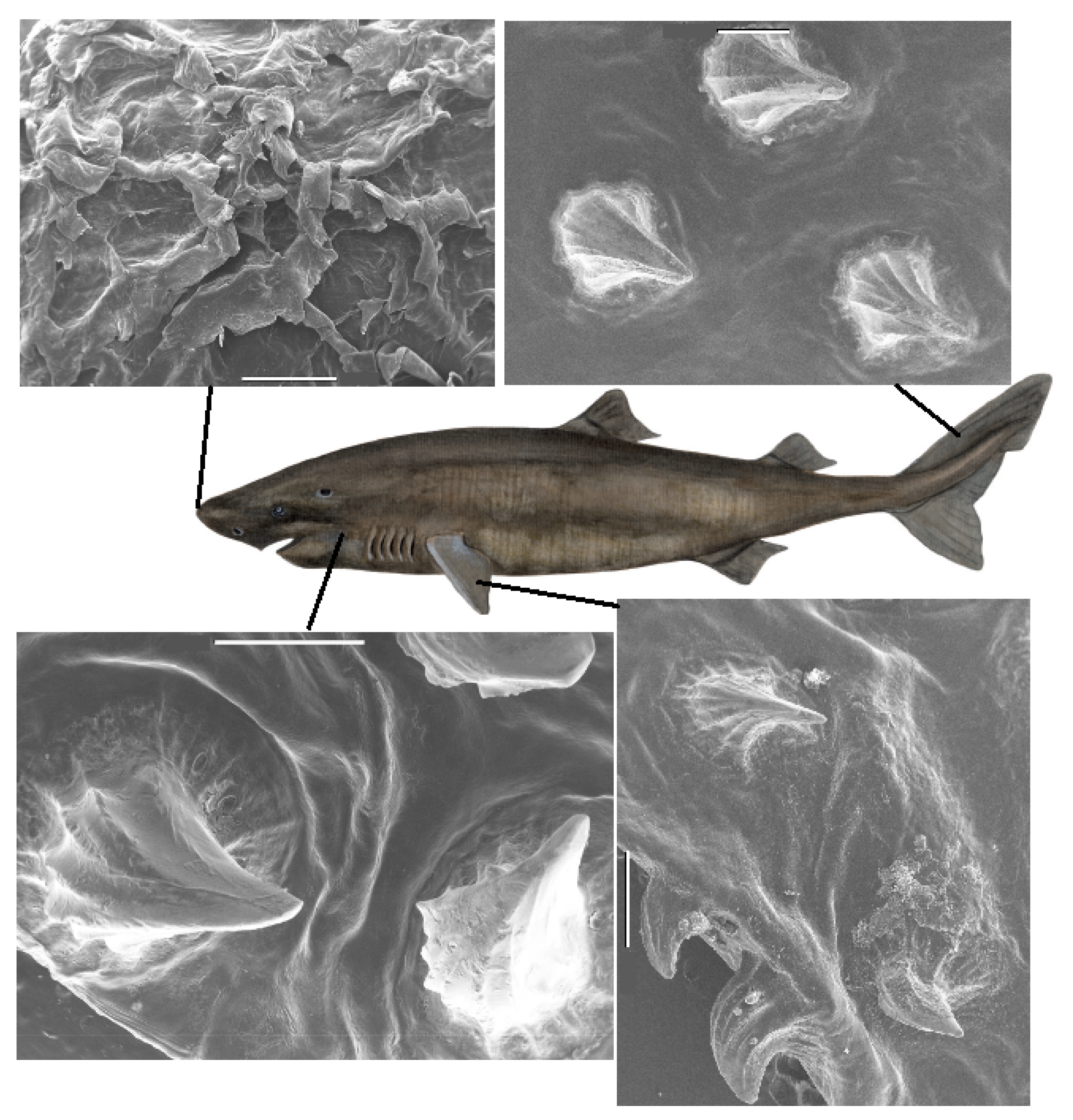
The exact size and distribution of dermal denticles depend on the actual position on the body of the shark. Therefore samples from sharks at selected positions on the body of the shark were investigated in detail, following an extension of the map in [
3], similar to that applied by other authors [
12,
13], see
Supplementary Figure S1. The detailed visualization of the structures of dermal denticles in the skin of the shark samples, and of their distribution and areal density, is conducted by use of scanning electron microscopy (SEM) and optical microscopy.
2. Results of Microscopy of Dermal Denticles
For the small-spotted catshark, detailed images of the dermal denticles were obtained by SEM with examples shown in
supplementary information, Figure S2, and less detailed by optical microscopy. Indicative variations over the body are apparent from
Figure 1, showing the dermal denticles visualized by optical microscopy. Sizes and areal densities of the dermal denticles were based on the optical microscopy images. From the microscope images of the denticles, the length of a few denticles and the total number of denticles in each frame were noted. Our investigations indicated variations in the crown length of the dermal denticles between 300
m and 1 mm, with areal densities between 400 cm
and 2000 cm
. As one could expect, the longer the denticles, the lower their areal density. However, the relation between the length and the density is not described well by any power law, for this species. Smaller dermal denticles are seen on the fins and near gills whereas larger dermal denticles are found on the body (see
supplementary information, Figure S6).
In addition to SEM and optical microscopy to determine density and size of the dermal denticles, we investigated one alive, adult shark, in an attempt to decide if bristling of the dermal denticles may take place; bristling has been investigated in recent literature and may be important for the hydrodynamic properties of the skin [
11,
12]. The shark was briefly positioned with its fin or tail under a stereo microscope and bending was induced to search for signs of bristling. We did not see evidence for this type of repositioning of the dermal denticles (see
supplementary information). More detailed investigations as in [
11] were not possible with an alive specimen.
Also, for the Greenland shark, detailed images of the dermal denticles were obtained by SEM. Variations over the body are illustrated in
Figure 2, with additional examples in
supplementary information, Figure S3. The dermal denticles of the Greenland shark are particularly large, and on the microscopic scale, appear more sparsely distributed than those of the other shark species.
To determine the sizes and densities of the dermal denticles for determination of areal density and average size, skin samples were visualized with optical microscopy, with additional sample images available in the
supplementary information (Figures S4 and S5). The logarithm of the density appears roughly inversely proportional to the length of the denticles. Also, for the Greenland shark, smaller dermal denticles are found on the fins and near the gills and larger denticles on the body (see
supplementary information, Figure S6). Based on the shape of these dermal denticles, bristling is in our view unlikely.
The dermal denticles of the spiny dogfish were comparably smaller and all images were obtained by SEM, with examples shown in
Figure 3. Also, for this species, the larger the denticle, the lower the areal density (see
supplementary information, Figure S7). The possibility for bristling was not investigated as samples had dried in the freezer before our experiments were conducted.
4. Discussion
In
Section 2, microscopy images from 31 positions on samples from each of the three shark species were analyzed for areal density and size distribution. Dermal denticles near gills and on the fins were smallest and most densely distributed, whereas the largest dermal denticles were found on the back side of the sharks. One might have suspected a power law dependence of density to dimension, but our data do not show such behavior in a convincing manner.
In
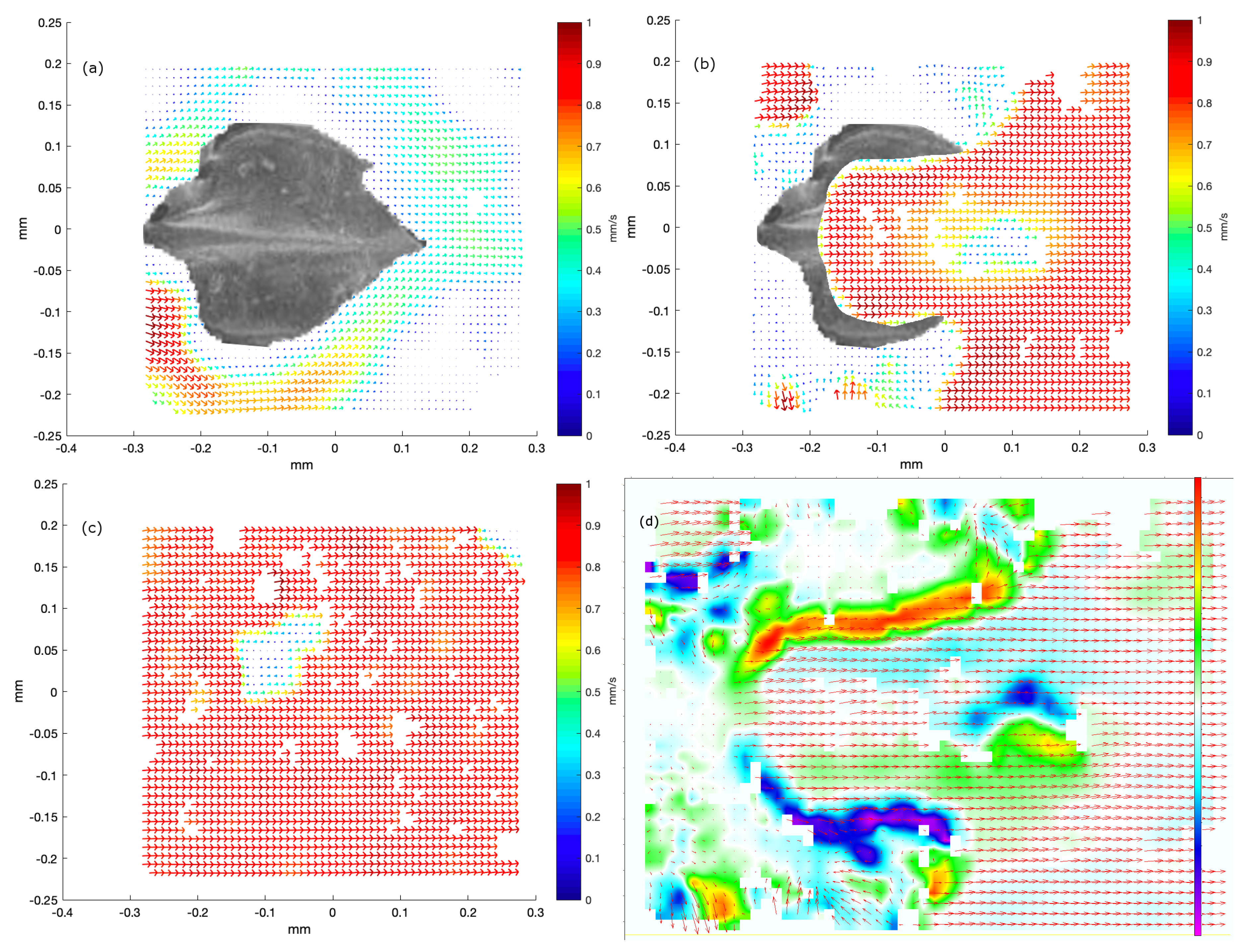
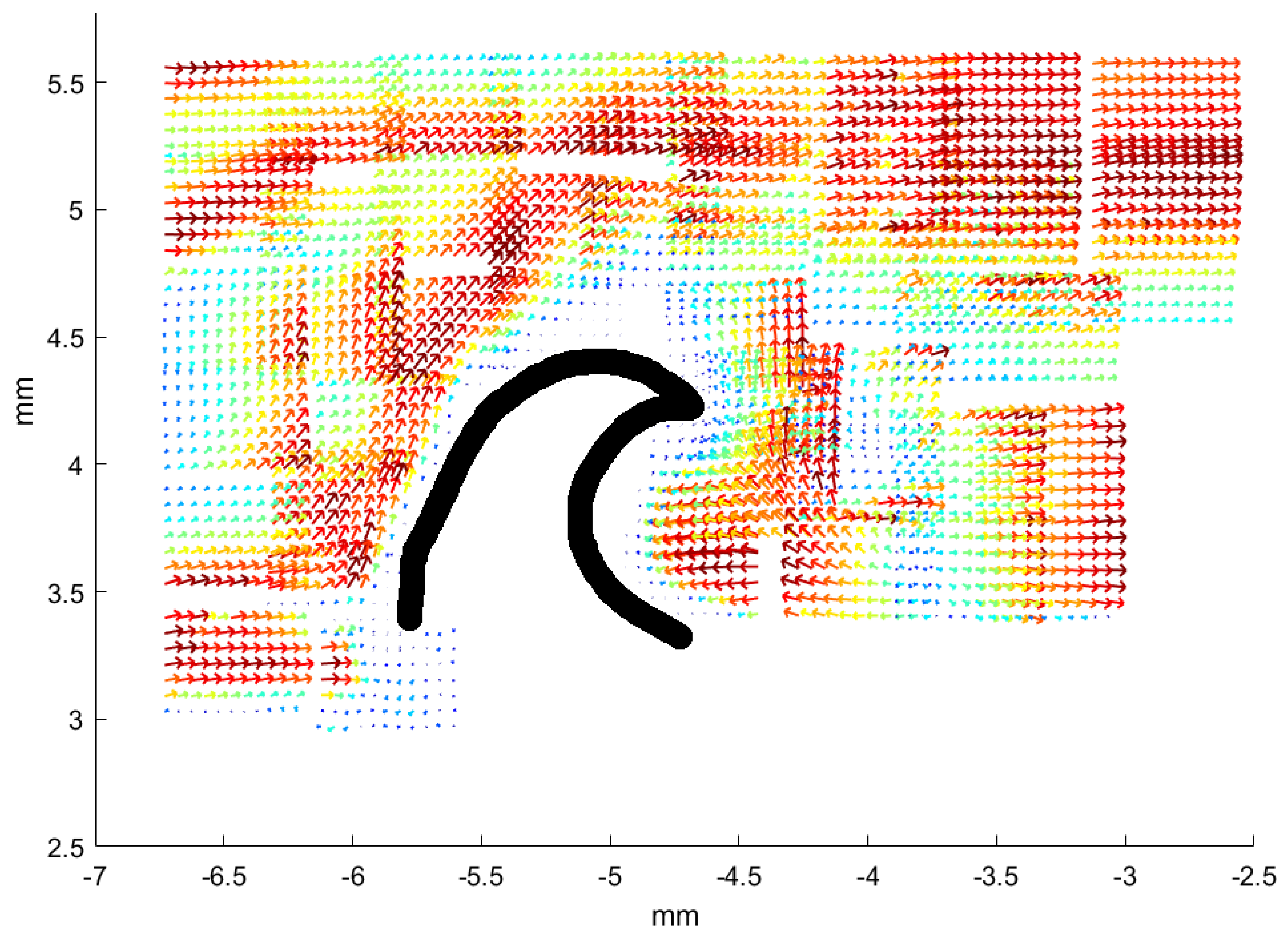
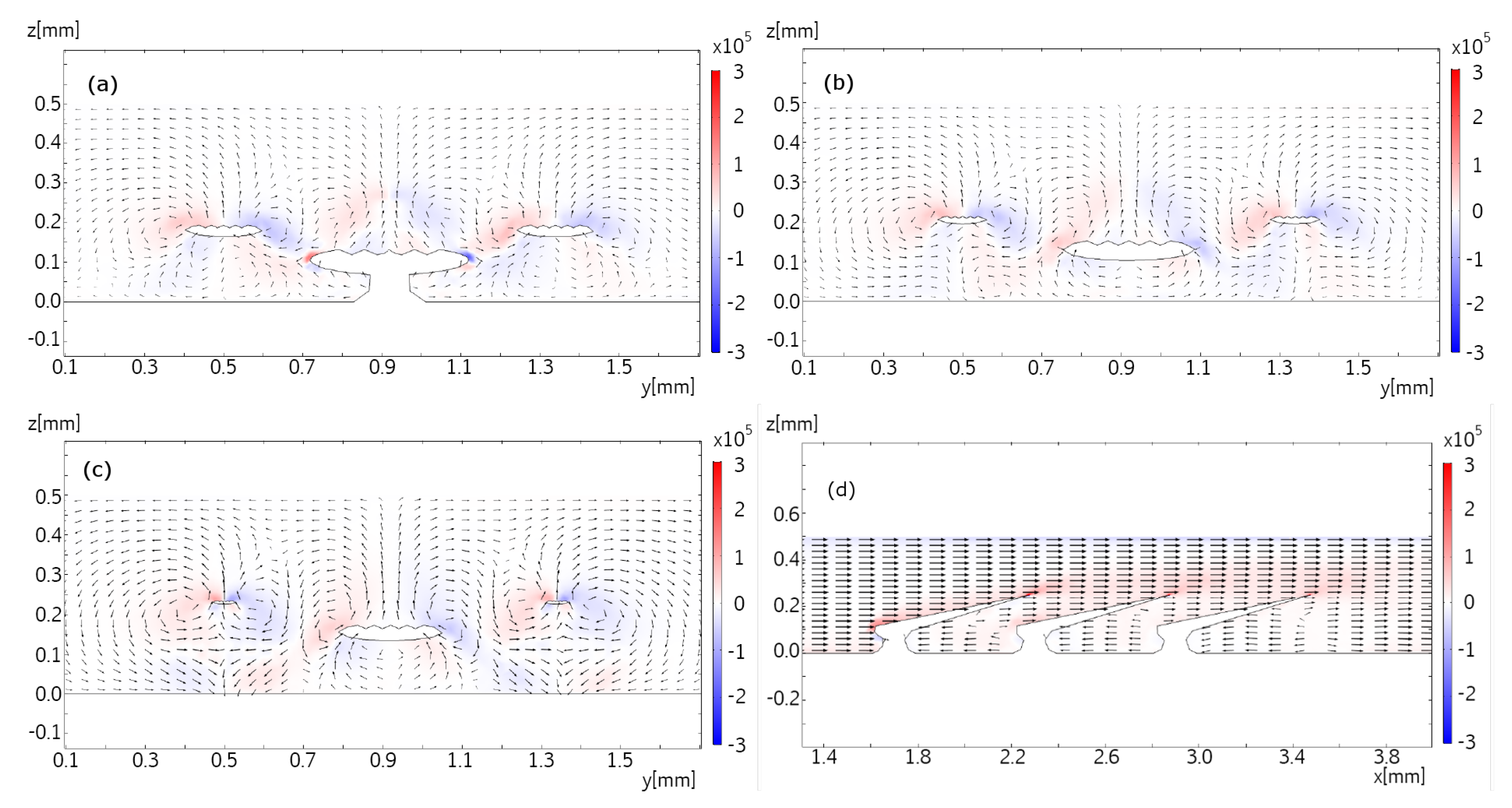
Section 3, we selected a few representative examples from the many samples to demonstrate that micro-PIV measurements very near shark skin surfaces are possible and, with the experimental assay discussed here, the micro-PIV measurements provide a close-up view of flow above or around actual dermal denticles. Our study reveals the flow patterns near real shark skin, rather than artificial mock-ups, and we find coherent structures such as the recirculation bubble and coherent eddies convected with the flow. Similar patterns were found in a numerical model, although at much higher Reynolds number. We acknowledge, though, that the microfluidic chambers constructed here do not provide a completely realistic view on the flow patterns near the dermal denticles of a swimming shark, both due to the closeness of nearby surfaces and due to the fact that the skin sample is held flat and rigid.
According to the conjectures in the literature from the 1980s [
3], the dermal denticles of the three species that we have investigated should provide defense against ectoparasites or abrasion strength or both in the case of the small-spotted catshark. One study in the literature discusses the possible role of the dermal denticles of this shark species for anti-fouling [
19], and demonstrates that structures provide reduced fouling for particular flow speeds. Interestingly, other literature on shark-mimetic structures that show anti-fouling behavior [
20,
29]—possibly corresponding to the suggested function of defense against ectoparasites—are inspired by the perhaps better-known dermal denticles of fast-moving sharks. Mytilid shells that have also inspired biomimicry for anti-fouling purposes [
30] also have ribbed structures, of dimensions similar to the riblets in e.g., dermal denticles of a scalloped hammerhead shark (
Sphyrna lewini). It is, therefore, tempting to speculate on requirements for dermal denticles to provide anti-fouling function.
The microscopy images in
Figure 1 and
Figure 3 suggest that the dermal denticles of these two species densely cover the skin of the shark. All dermal denticles also have a few riblets. However, as shown in
Figure 2, as well as
Figures S3–S5, the dermal denticles of the Greenland shark have a pointed structure with riblets on the denticle surface, ending at the pointed tip. In addition, the density of dermal denticles is sufficiently low that a very large fraction of the skin of Greenland shark, at the
m-mm scale, is without dermal denticles and thus exposed to parasites, if one assumes that the riblets alone provide the anti-fouling function. At several occasions, large 3–4 cm crustacean ectoparasites have been observed on the skin of Greenland shark from both Greenland and Norway [
31], but we found no published literature on the matter. It therefore appears that the large dermal denticles do not necessarily seem to protect the Greenland shark completely from such infections. However, as discussed in detail below, it is possible that the vortex flow generated across the skin protect from infections of smaller bacteria or parasites, and future and more detailed and targeted investigations on this particular effect of Greenland shark dermal denticles are called for.
Our alternative hypothesis suggests that the actual structure of the denticles is not decisive for the ability of the denticle to provide anti-fouling, at least not for small fouling agents: Dermal denticles introduce a flow unsteadiness which leads to larger peak velocities intermittently. During a velocity peak, the wall shear stress also increases. We speculate that small fouling agents may be washed away during such a velocity peak. It just requires that only one or a few peaks in the velocity could wash away a parasite that perhaps manages to stay on the shark for lower speeds. Further work is required to investigate the idea, but we provide here first estimates that agree with our speculation: Fouling agents with characteristic sizes of order
m tend to follow the local flow unless they have a way to attach themselves to the shark. This can be estimated from their Stokes number
where
is the characteristic momentum response timescale and
is the characteristic flow timescale which can be thought of as the eddy turnover time. If
, a free fouling agent will follow the local flow (as e.g., airborne seeds), and if
, the fouling agent will not react to the flow (ballistic flight) [
32].
The flow time scale,
, can be estimated from the length scale
l and eddy velocity scale
as
. For
m/s and
μm, one finds
s. To estimate the momentum response time of the fouling agent, we assume it to be spherical. Then Newton’s law with the hydrodynamic force is
We introduce the characteristic momentum response timescale
in which the fouling agent reacts to the local flow velocity as
. Solving for
gives:
Introducing the Reynolds number of the fouling agent
we get
where the first fraction approaches unity for Stokes flow and
for typical microorganisms in water. The momentum response time can now be estimated as
The detailed geometrical form of the fouling agent changes the numerical factor 18, but not the basic scaling. For water
m
/s. For
μm, we find the momentum response timescale to be
s. The Stokes number is
As for these parameters, a fouling agent will react to local eddies and flow unsteadiness introduced by dermal denticles. Future work is called for to test this hypothesis.
In summary, we have presented detailed microscopical investigations of dermal denticles from three slowly swimming shark species for which, according to original hypotheses by Reif [
3], the dermal denticles provide defense and possibly abrasion strength. While not addressing the postulated overall fluidic function of the dermal denticles, we investigated selected samples from these shark species through micro-PIV measurements of the flow near the dermal denticles, at low Re. The experimental investigations were supplemented by numerical simulations at high Re. In both cases, recirculation bubbles and vortex structures appear in the flow.
5. Experimental Methods
5.1. Microscopy
Scanning Electron Microscopy (SEM) was performed with electron microscopes at the student facility Nanoteket at DTU Physics (JEOL JCM-6000). Shark skin samples were cut into small pieces (about 5 mm × 5 mm) and positioned in copper stubs for the electron microscope. On some samples, element analysis was carried out as well, based on Energy Dispersive X-ray Spectroscopy available with the electron microscopy equipment. These results agree with the known composition of dermal denticles.
For the determination of areal density of dermal denticles in spiny dogfish, we visualized an area of 3.98 mm × 2.98 mm in the SEM and counted the number of dermal denticles within this area. In addition, measurements of width and length of selected dermal denticles were performed.
For small-spotted catshark and Greenland sharks, the magnification allowed by an optical microscope (Leica DM 2000 LED) equipped with a CCD camera (Leica ICC50 HD) was sufficient. A pre-defined square was selected and the dermal denticles within this area were counted and measurements of denticle width and length performed. Furthermore, we conducted one microscopy session using an optical stereomicroscope (Leica MZ125) equipped with a digital camera (Canon D5 Mk2).
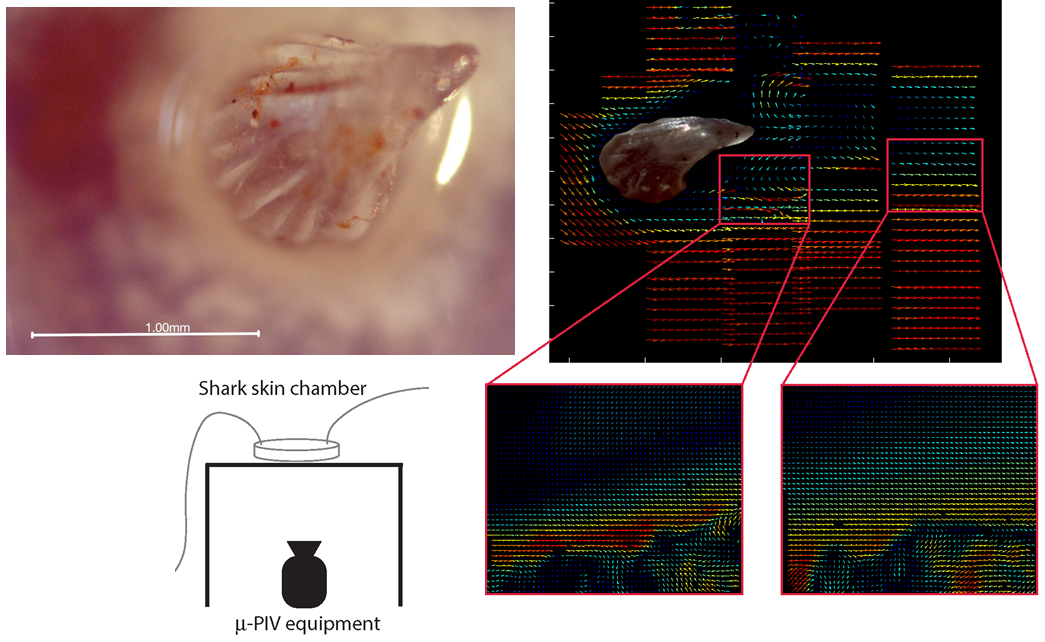
5.2. Micro-PIV and Microfluidic Setup
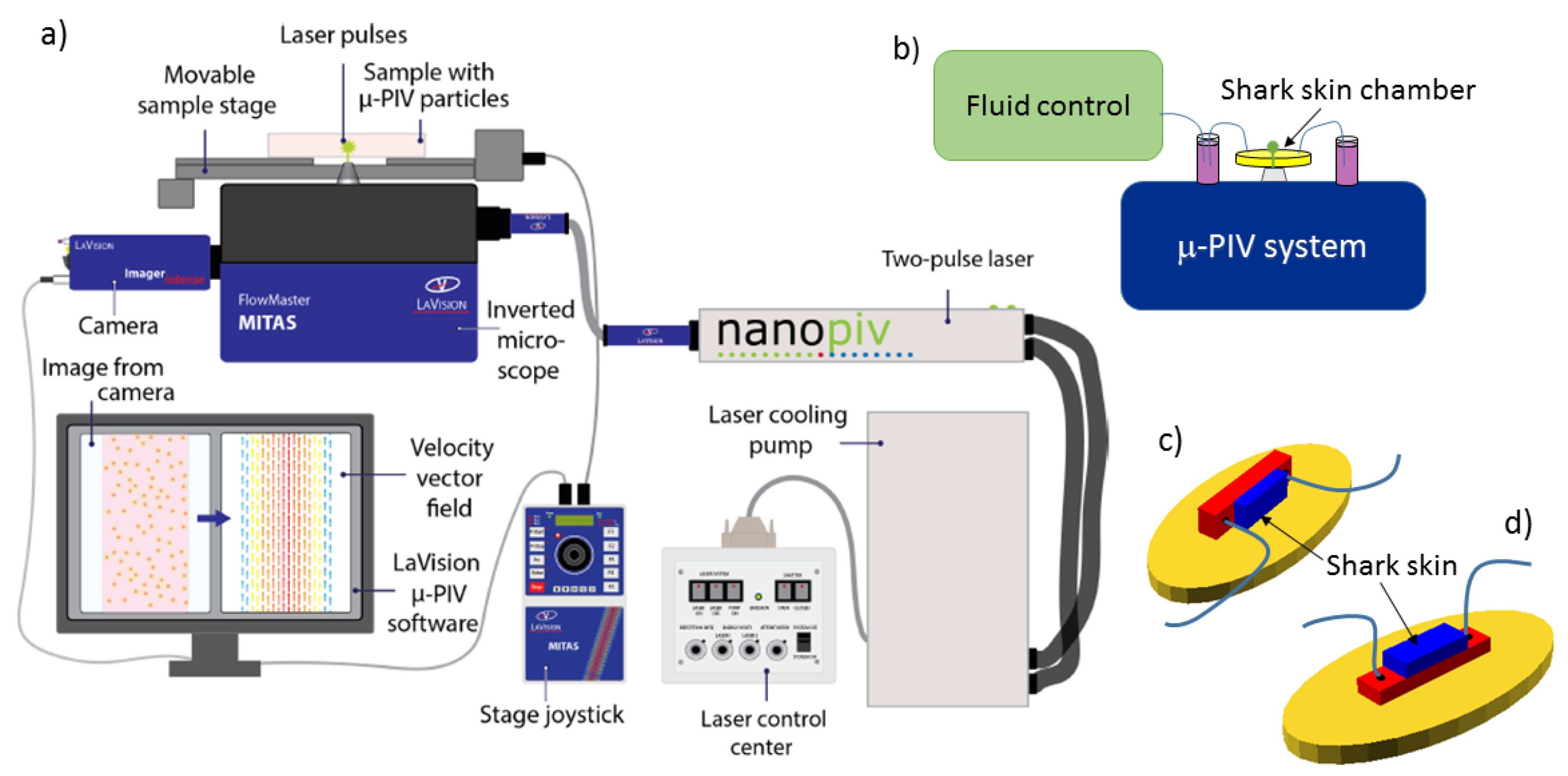
Micro-PIV experiments were performed with a commercial Flowmaster Mitas system from LaVision, based on an Nd:YAG double-cavity laser (Litron nano s65—15PIV) and a CCD camera (LaVision Imager Intense). The laser illuminates 0.86
m or 2
m diameter fluorescent microspheres (Fluoro-Max R900 and R0200, Thermo Scientific). Imaging was performed with either a 10× or 20× objective (Zeiss, LD Plan-Neofluar, long working distance), in addition to the magnification optics embedded in the Flowmaster Mitas system. Actual dimensions were calibrated using a calibration plate. Image processing and velocity maps were calculated with the software DaVis 8.2. Time-averaged velocity maps were obtained through the method sum of correlation [
33], as implemented in DaVis, and recommended as the best averaging method with our experimental conditions. The setup is illustrated in
Figure 9a,b.
Microfluidic chambers were based on custom-made 3D printed molds that were cast by soft lithography in polydimethylsiloxane polymer (PDMS; Sylgard 184 Silicone elastomer kit). The PDMS chamber contains a rectangular indentation to create a flow chamber. Within the flow chamber, another indentation is designed to fit a piece of shark skin, 4 mm × 10 mm large, glued to the PDMS with standard two-component epoxy glue. Chambers constructed for control experiments are prepared without the latter indentation. The flow chamber was sealed by a standard microscope slide glued with double sticky tape (ARcare 90106) to the PDMS chamber. Tubing (Teflon tubing, 1.6 mm outer diameter, VWR 228-0738) was mounted in the chamber by drilling holes through the PDMS part prior to gluing the shark skin and to sealing with the microscope slide. PDMS chambers were constructed to allow for 2D micro-PIV measurements both in the plane of the shark skin and with a side view on the dermal denticles. Chamber construction is illustrated in
Figure 9c,d.
Microfluidic flow was generated by a pressure-driven pump (Fluigent MFCS-EZ, 4 channel, 345 mbar), and for some experiments, the flow speed was monitored by the Fluigent Flow Rate Platform (Flowboard) with a flowunit size XL.
Control experiments confirmed the overall flow velocity and Poiseuille-flow behavior, with agreement of the numerical value of the average velocity within 50%. These studies also confirmed very low velocities near the channel walls, consistent with a no-slip boundary condition.
5.3. Experimental Geometry and Limitations
The flow chamber used for visualization of the flow has dimensions of 4 mm × 18 mm × 1–4 mm, fitting shark skin samples of size 4 mm × 10 mm. To fit the large dermal denticles of the Greenland shark, a chamber depth of 4mm was necessary for those samples. Smaller chamber depths were chosen for the other sharks, i.e., 3 mm for the small-spotted catshark and 1mm for the spiny dogfish. The flow rate platform allows a theoretical limit of maximum 5 mL/min and the pump a maximum applied pressure of 345 mbar.
The natural swimming speeds of the three shark species are not well known. For the Greenland shark, a cruising speed of 0.34 m/s has been suggested [
34]. We aimed to perform the experiments at a flow speed that would mimic this estimated cruising speed of a Greenland shark, but were limited by the performance of the pumping system and the requirement for the microfluidic chambers to stay tightly sealed.
6. Numerical Methods
Numerical simulations of the flow in the vicinity of dermal denticles aid the understanding of flow phenomena resulting from a geometry with dermal denticles as well as the interpretation of the micro-PIV measurements. Whereas the micro-PIV measurements are two-dimensional, the numerical simulations give access to the flow velocities and vorticities in three dimensions, complementing the measurements. Furthermore, the numerical simulations allow for visualization of the flow patterns in a larger region of space than that investigated microscopically, and with higher Reynolds numbers.
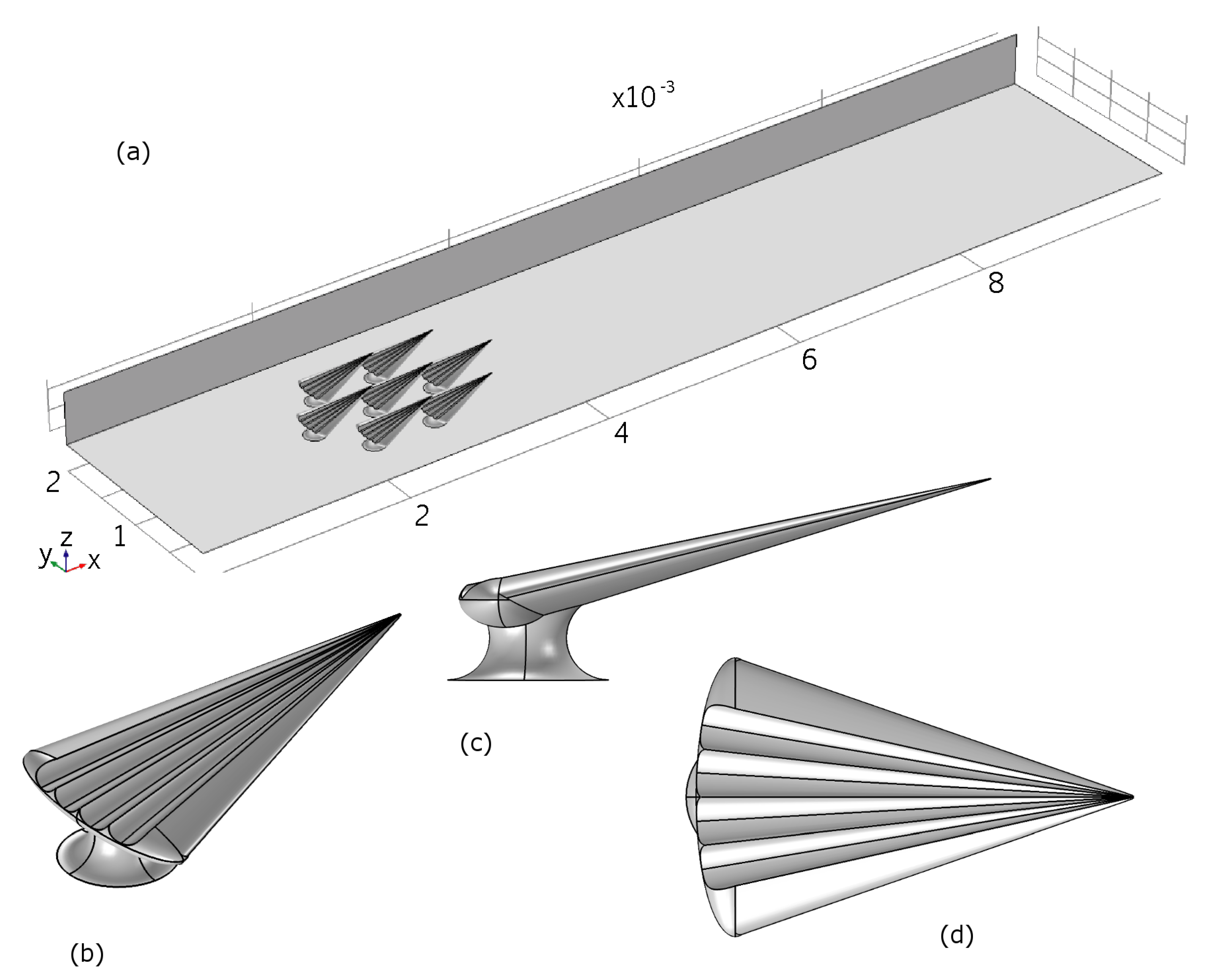
We generated a CAD model simulating an archetypical dermal denticle, modelled to resemble the dermal denticles of the small-spotted catshark. Seven identical dermal denticle models were positioned with a spacing and pattern similar to that observed from SEM microscopy. The areal density of the dermal denticles is 7/mm
. SEM images such as those in
Figure S2 were used to build a realistic model of the shape of the dermal denticle including the ribbed structures aligned with the flow on the water-facing side. Details of the geometry of the denticles are provided in
Table 1 and
Table 2.
The dermal denticles were placed in a channel with the dimensions of the micro-PIV experiment. The simulation model is illustrated in
Figure 10, which also illustrates the size of the domain, namely 10 mm along the direction of fluid flow (
x), 2 mm wide (
y) and 0.5 mm high (
z). In the simulations, mirror symmetry in the
-plane at
mm is assumed, to reduce computational effort, resulting in a final physical domain of 10 mm by 4 mm by 0.5 mm. No-slip boundary conditions are assumed at top (
-plane at
mm), bottom (
-plane at
mm) and sides (
-planes at
mm and
mm), as well as at the surface of the dermal denticles. The center of the first dermal denticle is positioned 2 mm after the inlet. At the inlet, a fully developed turbulent velocity profile is assumed, and deduced from a simulation of the steady-state solution in a domain of the same cross-sectional area and boundary conditions, but without dermal denticles. At the outlet, flux-conserving zero-gradient conditions on the flow velocities were assumed.
The size of the domain is chosen to fit the experimental conditions, and thus boundary effects that may influence the flow measured would also be found in the numerical model.
The flow is simulated using the incompressible Reynolds-averaged Navier-Stokes equations of Newtonian fluids which are closed using a standard k--turbulence model. The Reynolds number is Re∼5000 based on typical length scales of dermal denticles and flow velocities, providing a Reynolds number orders of magnitude larger than the limitations imposed in the experiments. The equations were discretized using the finite element method implemented in the commercially available software COMSOL. We used an unstructured grid with tetrahedral cells close to the dermal denticles and a structured grid downstream. The grid size was determined after numerical grid convergence tests. In all, the grid had just over 500.000 cells.

 ,
, 











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}