Abstract
The present work was aimed at identifying the IgE-binding epitopic regions on the surface of the Cup s 3 allergen from the common cypress Cupressus sempervirens, that are possibly involved in the IgE-binding cross-reactivity reported between Cupressaceae species. Three main IgE-binding epitopic regions were mapped on the molecular surface of Cup s 3, the PR-5 thaumatin-like allergen of common cypress Cupressus sempervirens. They correspond to exposed areas containing either electropositive (R, K) or electronegative (D, E) residues. A coalescence occurs between epitopes #1 and #2, that creates an extended IgE-binding regions on the surface of the allergen. Epitope #3 contains a putative N-glycosylation site which is actually glycosylated and could therefore comprise a glycotope. However, most of the allergenic potency of Cup s 3 depends on non-glycosylated epitopic peptides. The corresponding regions of thaumatin-like allergens from other closely related Cupressaceae (Cryptomeria, Juniperus, Thuja) exhibit a very similar conformation that should account for the IgE-binding cross-reactivity observed among the Cupressaceae allergens.
1. Introduction
Allergy to common cypress (Cupressus sempervirens) pollen is widely distributed over Mediterranean countries, especially in Italy [1], Israel [2], Greece [3], Spain [4], and the south of France [5]. Pollens from other closely related Cupressaceae species are responsible for seasonal rhinitis in Japan (Japanese cypress Chamaecyparis obtusa, Japanese cedar Cryptomeria japonica) [6], Australia (Callitris glaucophylla) [7], USA (Mountain cedar Juniperus ashei, Arizona cypress Hesperocyparis arizonica) [8], France (Northern whitecedar Thuja occidentalis) [9], and Mediterranean countries (prickly juniper Juniperus oxycedrus) [10]. The exposure to Cupressaceae pollen allergens and consequently allergy levels have increased these last 50 years and is still increasing for several reasons. Firstly, besides the classical geographical distribution, Cupressaceae trees are spreading over many other regions due to their increasing use as ornamental trees, especially in urban and suburban areas [11]. Secondly, because of global warming, the pollination period of cypresses and other Cupressaceae species lasts longer, which promotes the currently observed spreading of their associated allergies, especially in northern Europe [12,13]. Thirdly, the expression of a group 3 allergen of Cupressaceae that belongs to the thaumatin-like protein family was reported to increase under an increasingly polluted environment [14].
Different allergens have been characterized in the pollen of Chamaecyparis obtusa (Cha o 1, Cha o 2, Cha o 3), Cryptomeria japonica (Cry j 1, Cry j 2, Cry j 7), Cupressus arizonica (Cup a 1), C. sempervirens (Cup s 1, Cup s 2, Cup s 3, Cup s 7), Juniperus ashei (Jun a 1, Jun a 2, Jun a 3, Jun a 7), J. oxycedrus (Jun o 1, Jun o 4), J. sabinoides (Jun s 1), and J. virginiana (Jun v 1, Jun v 3) (WHO/IUIS Allergen Nomenclature Sub-Committee, www.allergen.org, assessed on 15 October 2022). They essentially correspond to ubiquitous proteins such as pectate lyase (group 1 allergens), polygalacturonase (group 2 allergens), PR-5 thaumatin-like protein (group 3 allergens), and Ca2+ binding protein (calmodulin, group 4 allergen). Besides the WHO/IUIS certified allergens, about 20 additional allergens have been described in Cupressaceae pollen [13]. Recently, three additional allergens belonging to the family of Gibberellin-Regulated Proteins (GRP), Cup s 7 from Cupressus sempervirens [15], Cry j 7 from Cryptomeria japonica [16], and Jun a 7 from Juniperus ashei [17], have completed the list of pollen allergens from the Cupressaceae. Interestingly, these pollen GRP allergens display some IgE-binding cross-reactivity with the corresponding GRPs from fruits and vegetable [18,19]. According to the ubiquitous character of the major Cupressaceae allergens, IgE-binding cross-reactivity has been reported to occur between closely related Cupressaceae pollens [20]. PR-5 thaumatin-like proteins of Cupressaceae pollen offer an example of conserved allergens able to trigger IgE-binding cross-reactions and thus constitute an appropriate model to decipher the structural basis of this IgE-binding cross-reactivity [14,21,22,23,24,25].
Here, we report on the characterization of IgE-binding epitopic peptides of the PR-5 thaumatin-like allergen Cup s 3 [26] and their structural features responsible for the reported IgE-binding cross-reactivity among different Cupressaceae species. In addition, the identification of IgE-binding epitopic regions of pollen allergens is an exciting goal since synthetic peptides mimicking the IgE-binding epitopic stretches identified on the molecular surface should be used instead of recombinant allergens for the purpose of diagnosis, e.g., in immobilized epitope micro-arrays.
2. Materials and Methods
2.1. Cypress Pollen Extract
A crude protein extract was prepared from common cypress (Cupressus sempervirens) pollen (Allergon, Angelholm, Denmark) by suspending 1 vol. of pollen in 10 vol. of 50 mM NH4HCO3 (pH 6.5). After stirring for 3 h at 4 °C, the slurry was centrifuged at 20,000× g for 30 min and the supernatant was collected and kept frozen at −20 °C until used. The protein content of the protein extract was evaluated using the bicinchoninic acid kit reagent (Pierce) [27], with the banana TLP Mus a 4 [28] as a standard. The occurrence of both Cup s 1 (pectate lyase) and Cup s 3 in the protein extract was checked by sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE) in 15% polyacrylamide gels using Tris-glycine as trailing ion [29] and staining with Coomassie blue or silver nitrate.
2.2. Sera from Allergic Patients
Blood samples were collected after informed consent of patients selected according to symptoms as rhino-conjunctivitis and/or asthma to Cupressaceae pollen and individual sera were used in SPOT experiments and ELISA. All the used cypress pollen allergic patient (CPAP) sera interacted with Cupressaceae pollen extracts (ImmunoCAP ® t23, Thermo Fisher Scientific, Phadia 67400 Illkirch, France) (Table 1). Sera from this list are the same as previously reported in [30].

Table 1.
List of sera from patients allergic to cypress pollen, used in SPOT and ELISA inhibition experiments.
2.3. IgE-Binding Epitope Mapping
Overlapping 15-mer peptides, frame shifted by three residues, corresponding to the entire amino acid sequences of Cup s 3 (Accession number Uniprot. Q69CS2), were synthesized using the SPOT technique [31]. For these experiments we used the Multipep automatic SPOT synthesizer (Intavis Bioanalytical instruments, Cologne, Germany). After different treatments to bind the peptides as described previously [32] the membranes were soaked overnight in tris-buffered saline (TBS) containing 2 mL blocking buffer (Roche-Diagnostics, Meylan, France) and 1 g sucrose (pH 7.0). Then, membranes were incubated (2 h) with CPAP serum (1:10 v/v) in the presence of an anti-protease cocktail (Roche) in a moist chamber. Membranes were then soaked in a 1:4000 dilution of mAb anti human-IgE coupled to Alkaline Phosphatase (AP) (Sigma-Aldrich, St Louis, MO, USA) for 1 h. After, the interacting peptide spots were stained for 30 min by adding the AP substrate, 5-Bromo-4-Chloro-3-Indolyl Phosphate (BCIP, Promega corporation, Madison, WI, USA). Then the membranes were washed three times with deionized water and dried for scanning. Negative controls were obtained by using either TBS without serum or a serum from non-allergic patients to Cupressaceae pollen. Between each incubation step, 3 washes with TBS containing 0.1% (v/v) tween 20 (TBS-Tw) were performed. Membranes can be used repeatedly after regeneration steps consisting of 1–3 washes in dimethylformamide for 10 min each, 3 washes in deionized water, 3 washes in 8M urea containing 1% (w/v) SDS and 1% (w/v) β-mercaptoethanol and finally 3 washes in a mixture of ethanol–acetic acid–H2O (50:10:40, v/v/v).
2.4. ELISA Inhibition Experiments
Three 9-mer peptides that broadly correspond to the predicted epitope #1 (peptide #1: 9PGGGKRLDQ27), epitope #2 (peptide #2: 45RFWGRTGIT53), and epitope #3 (peptide #3: 161TNYSKIFKN169) of Cup s 3, were synthesized using the Fmoc (Fluorenylmethyloxycarbonyl) chemistry (JPT Peptide Technologies GmbH, Berlin, Germany). The capacity of synthetic peptides to inhibit the IgE-cypress extract interaction was checked by ELISA (Enzyme-Linked Inhibition Sorbent Assay). Briefly, the wells were coated with cypress pollen extract in phosphate buffer saline (PBS, pH 7.5), containing 1 μg.mL-1 protein, and after overnight incubation (4 °C), the wells were washed 3 times with PBS and then incubated (2 h, room temperature) with PBS containing 0.1% tween 20 (v/v) and 1% BSA (Bovine serum albumin) (w/v) (PBSTB). Then, 50 μL of 1:30 diluted CPAP serum in PBSTB, previously incubated with 1 mM or 2 mM of synthetic peptide, were added and the plates were incubated for 1 h 30, at room temperature, under constant stirring. Then, mAb anti human-IgE-AP (Sigma-Aldrich, St. Louis, MO, USA) was added (diluted 1:500) for 1 h incubation followed by BCIP (45 min in the dark). Between each incubation step, 3 washes with PBST were performed. The absorbance at 405 nm was recorded on a Titertek Multiscan spectrophotometer (Labsystems, Thermo Fisher Scientific, Villebon-sur-Yvette, France). Each value is the mean of three separate experiments and appropriate controls were performed under the same conditions.
2.5. Bioinformatics
Multiple amino acid sequence alignment of Cupressaceae PR-5 thaumatin-like proteins (TLP) was carried out with CLUSTAL-X [33]. An unrooted phylogenetic tree was built from the multiple amino acid sequence alignment of PR-5 TLP, using the neighbor-joining method.
Homology modeling of Cup s 3 from Cupressus sempervirens was performed with the YASARA Structure program [34]. Different models of Cup s 3 were built from the X-ray coordinates of the banana (Musa acuminata) thaumatin-like protein allergen Mus a 4 (PDB code 1Z3Q) [35], the kiwi (Actinidia deliciosa) fruit allergen Act d 2 (PDB code 4BCT), the grape (Vitis vinifera) thaumatin-like protein (PDB code 4JRU) [36], the Sodom apple (Calotropis procera) laticifer osmotin (PDB code 4L2J), and the antifungal protein zeamatin of maize (Zea mays) (PDB code 1DU5) [37], used as templates. Finally, a hybrid model of Cup s 3 was built up from the different previous models. PROCHECK [38], ANOLEA [39], and the calculated qualitative model energy analysis (QMEAN) scores [40,41], were used to assess the geometric and thermodynamic qualities of the three-dimensional model. Only three residues (Asn101, Cys118, Gln173) over 199, occurred in the non-allowed regions in the Ramachandran plot. Using ANOLEA to evaluate the model, only 6 residues (over 199) of the Cup s 3 model exhibited an energy over the threshold value. Both residues are mainly located in the loop regions connecting the β-sheets to the α-helices in the model. The calculated QMEAN score of the model gave a value of 0.664. Other thaumatin-like proteins from other Cupressaceae species including TLP-Ca from Hesperocyparis/Cupressus arizonica, TLP-Cj from Cryptomeria japonica, Jun a 3 from Juniperus ashei, TLP-Jr from Juniperus rigida, TLP-To from Thuja occidentalis and TLP-Ss from the closely related Taxodiaceae species Sequoia sempervirens, and TLP-Txm from the Taxaceae species Taxus × media, were similarly modeled using the same PDB code proteins as templates. Reliable values of 0.749 (TLP-Ca), 0.768 (TLP-Cj), 0.642 (Jun a 3), 0.699 (TLP-Jr), 0.717 (TLP-To), 0.731 (TLP-Ss) and 0.704 (TLP-Txm), were obtained for the QMEAN score of the different modeled allergens, respectively.
The surface electrostatic potentials were calculated and rendered with YASARA using the Amber96 forcefield with dielectric constants applied to the protein and the solvent fixed at 4.0 and 80.0, respectively. Electrostatic potentials were displayed on the molecular surface as red (electro-negatively charged) and blue (electro-positively charged) patches. Neutral surfaces are white. Assuming the putative N-glycosylation sites at Asn/N residues of Cup s 3 is glycosylated by a high-mannose bi-antennary glycan chain with a tri-mannoside core (Man)5-(GlcNAc)2, the glycan chains was built on the allergen using the GlyProt server (http://www.glycosciences.de/modeling/glyprot/php/main.php, accessed on 4 September 2022) and further represented in CPK on the molecular surface. Molecular cartoons were drawn with YASARA. The surface occupied by the SPOT identified sequential IgE-binding epitopic stretches of the modeled Cup s 3 and other modeled allergens were differently colored and displayed on the molecular surface of the PR-5 TLP allergen models with Chimera [42]. The root-mean-square deviation of atomic positions (rmsd, in Å) between the superposable Cα of pairwise superposed PR-5 TLP allergens was calculated at the SuperPose web server (http://www.wishart.biology.ualberta.ca, accessed on 4 September 2022).
3. Results
The modeled Cup s 3 displayed the canonical three-domain structure of PR-5 thaumatin-like proteins, made of a β-sandwich central domain (I) flanked on both sides by an α-helical domain (II) and a short β-hairpin domain (III) (Figure 1A,B). Both domains I and II delineate a strongly electronegative central groove responsible for the endo-1,3-β-glucanase activity reported for PR-5 thaumatin-like proteins [43] (Figure 1C,D). The face opposite to the groove displays a more pronounced electropositive character. A very similar three-dimensional conformation was observed for other modeled PR-5 allergens of Arizona cypress (TLP-Ca), Japanese cedar (TLP-Cj), mountain cedar (Jun a 3), and Northern white cedar pollen (TLP-To) (results not shown). All these allergens share a high degree of both amino acid sequence identity (from 74 to 98% with reference to Cup s 3) and similarity as shown on multiple amino acid sequence alignment (Table 2).
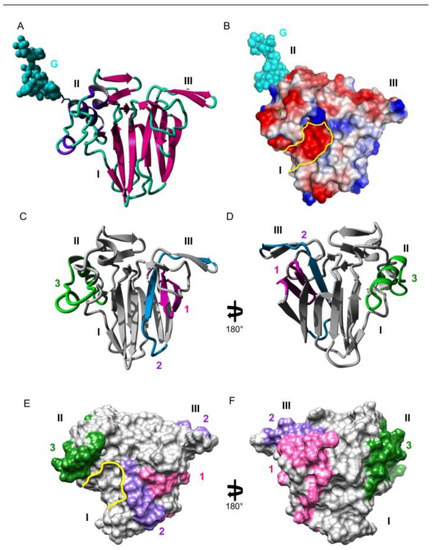
Figure 1.
(A). Ribbon diagram of the Cup s 3 model showing the overall organization of the allergen in three distinct domains labeled I, II, and III, and the N- glycan chain (G) attached to residue Asn162 in domain II, represented in CPK colored cyan. (B). Molecular surface of Cup s 3 showing the electrostatic potentials patches. The electronegatively charged groove is outlined in yellow. (C,D). Mapping of the sequential IgE-binding epitopes 1 (colored pink), 2 (colored blue), and 3 (colored green) on the front face (C) and the back face (D) of the ribbon diagram of Cup s 3. (E,F). Mapping of the sequential IgE-binding epitopes 1 (colored pink), #2 (colored blue) and 3 (colored green) on the front face (E) and the back face (F) of the molecular surface of Cup s 3. Epitope 2 consists of two separate patches on the molecular surface of the TLP allergen. Note the coalescence of epitope 1 with both patches of epitope 2.
Table 2.
Multiple amino-acid sequence alignments of seven thaumatin-like proteins from Cupressaceae pollen. The sequence of Jun v 3 is incomplete in the NCBI databank. Grey boxes correspond to the absence of insertion which are only present in TLP-Cj and TLP-To. Cystein residues are in red and red dotted lines indicate the eight disulfide bridges.
Three main IgE-binding epitopic stretches, that strongly interacted with all the tested patient’s sera, were identified using the SPOT technique (Figure 2A,B). They correspond to IgE-binding epitopic regions 1, 2, and 3, respectively. Two additional IgE-binding spots constantly interacted with all of the IgE-containing CPAP sera but the interaction was weaker and, therefore, they were not retained as major IgE-binding epitopes for Cup s 3. A few other spots, especially at the C-terminal end of the sequence, were more scarcely and faintly stained by a restricted number of patient sera and were not considered as relevant epitopic regions.
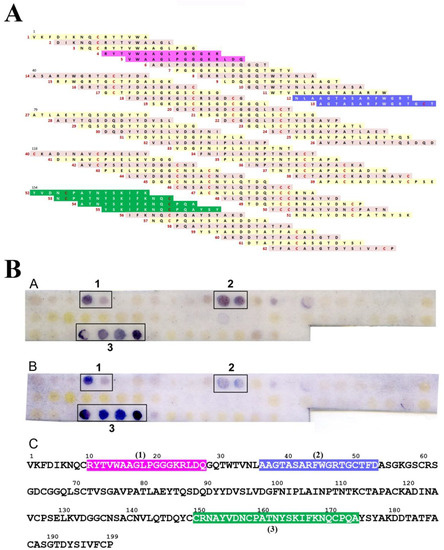
Figure 2.
(A). Amino acid sequences of the 62 overlapping peptides used in the SPOT technique. Epitope 1 (pink background), 2 (purple background), and 3 (green background) are indicated. Cystein residues are in red. Peptide numbering in red. (BA,BB). Examples of SPOT results obtained with two different sera from cypress allergic patients used as a probe. (BC). Sequential IgE-binding epitopic stretches 1, 2, and 3 deduced from the stained spots are indicated in colored boxed white letters along the amino acid sequence of Cup s 3.
Mapping of the major IgE-binding amino-acid stretches previously identified by the SPOT technique on the ribbon diagram (Figure 1C,D) and the molecular surface of Cup s 3 (Figure 1E,F), allowed us to accurately define those amino acid residues sufficiently exposed on the molecular surface of Cup s 3 to readily interact with the corresponding IgE. Exposed residues in each of the three IgE-binding stretches were thus considered as forming the major IgE-binding epitopic regions of Cup s 3 (Figure 1E,F). They correspond to more restricted areas on the surface of Cup s 3 than the whole IgE-binding peptide because some amino-acids are buried in the molecule and not accessible to antibodies. Epitope 1 (stretch 10RYTVWAAGLPGGGKRLDQ27) and part of epitope 2 (stretch 34NLAAGTASAR43) are distributed on domain I whereas the other part of epitope 2 (stretch 49RTGCTFD53) occurs only in the hairpin-containing domain III.
Epitope 3 (stretch 150CRNAYVDNCPATNYSKIFKNQCPQA174) coincides with the α-helix-containing domain II. These major IgE-binding epitopes are arrayed on both faces of Cup s 3 but do not interfere with the catalytic groove located in the front face of the molecule. Some of these epitopic regions, epitopes 1 and 2, are coalescent and generate an extended epitopic region on the molecular surface of Cup s 3 that could correspond to discontinuous epitopes. Epitope 3 contains a putative N-glycosylation site 162NYS that is predicted to be glycosylated (Figure 1B,D), and should therefore consists of a glycotope undetectable in the SPOT technique since all the assayed peptides are not glycosylated. This N-glycosylation site is conserved in other thaumatin-like allergens (TLP-Jr, TLP-Ca, Jun a 3, and TLP-To) (except TLC-Cj) from the Cupressaceae species (Table 2).
Synthetic peptides corresponding to the three main epitopic regions of Cup s 3 (epitopes 1, 2, and 3) were tested for ELISA inhibition of the interaction of IgE-containing sera from CPAPs with a total protein from cypress pollen extract. An inhibition in the range between 16–28% of the interaction occurred with all of the tested peptides, and up to 58% inhibition was obtained with an equimolar mixture of the three synthetic peptides (Figure 3). The synthetic peptide corresponding to epitope 3 was the most potent inhibitor. These results suggest that Cup s 3 participates in the allergenicity of Cupressaceae pollen.
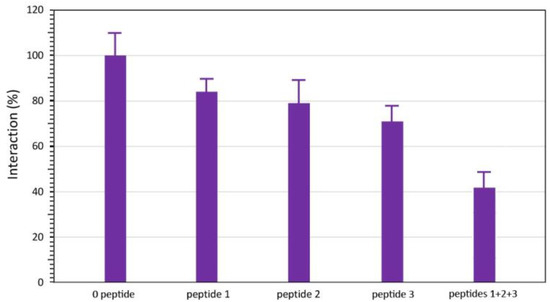
Figure 3.
ELISA inhibition of the interaction between cypress pollen extract and IgE from CPAPs, in the presence of 2 mM of synthetic peptides corresponding to epitope 1 (peptide 1: 9PGGGKRLDQ27), epitope 2 (peptide 2: 45RFWGRTGIT53), epitope 3 (peptide 3: 161TNYSKIFKN169), and an equimolar mixture of the three peptides 1, 2, and 3 (peptides 1 + 2 + 3). All values are the mean s.d. of three separate measurements performed in the same experimental conditions.
All of the three-dimensional models built up for other TLP pollen allergens consist of exhibit a very similar three-domain organization and are readily superimposed with a mean rms < 1.0-Å (Figure 4A,B). Accordingly, all the IgE-binding epitopic regions identified on the molecular surface of Cup s 3 consist of conserved residues and exhibit a high degree of conformational similarity (similar shape and extent) with the corresponding regions of TLP allergens from other Cupressaceae pollens. Despite amino acid changes that have occurred along the amino acid sequence stretch corresponding to epitope #2, the overall conformation of this epitopic region looks very similar in other Cupressaceae allergens (Figure 4C–H). However, the introduction of a bulky lysine residue in the sequence stretch TLP-Jr from Temple juniper and TLP-To from Northern whitecedar, induces some distortion into the overall conformation of epitope #2 (Figure 4E,H).
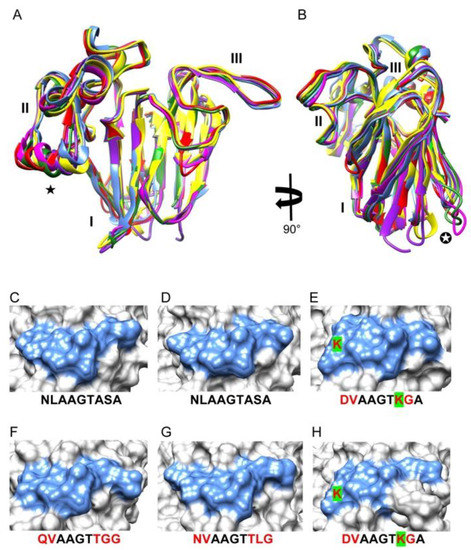
Figure 4.
Front view (A) and lateral view (B) of the superimposed ribbon diagrams of Cup s 3 (red), TLP-Ca (blue), TLP-Jr (yellow), TLP-Cj (magenta), TLP-Ss (green) and TLP-To (purple). The three domains I, II and III are labeled. Stars ★ and ✪ indicate the poorly superimposed regions in the ribbon diagrams of the TLP allergens, respectively. Overall conformation of epitope 2 at the molecular surface of Cup s 3 (C), TLP-Ca (D), TLP-Jr (E), TLP-Cj (F), TLP-Ss (G) and TLP-To 3 (H). The amino acid sequence stretches corresponding to epitope 2 are indicated and amino acid changes are shown in red letters. The K residue change responsible for the distortion of the overall conformation of epitope 2 observed in TLP-Jr (E) and TLP-To (H), is shown in green highlighted red letter.
These close conformational similarities most probably account for the currently reported IgE-binding cross-reactivity between Cupressaceae pollen species. In this respect, the unrooted phylogenetic tree built from the multiple alignment of the PR-5 TLP belonging to different species of higher plants, shows that pollen allergens from Cupressaceae, Taxodiaceae and Taxaceae, consist of a homogeneous clade of extremely closely related species, poorly related to other higher plant family clades (Figure 5).
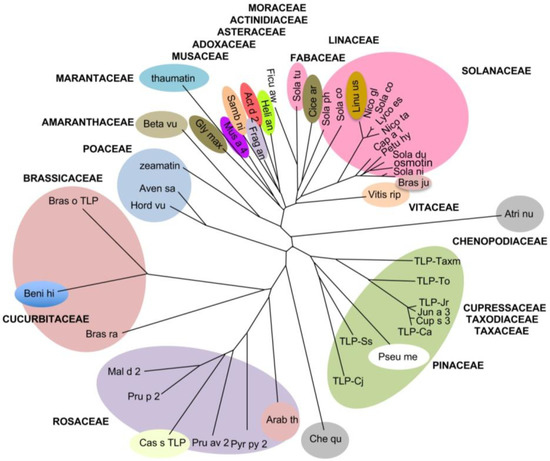
Figure 5.
Unrooted phylogenetic tree built up from the multiple alignment of thaumatin-like proteins of Actinidia deliciosa (Act d 2), Arabidopsis thaliana (Arab th), Atriplex numularia (Atri nu), Avena sativa (Aven sa), Benincasa hispida (Beni hi), Beta vulgaris (Beta vu), Brassica juncea (Bras ju), Brassica oleracea (Bras ol), Brassica rapa (Bras ra), Capsicum annuum (Cap a 1), Castanea sativa (Cas s TLP), Chenopodium quinoa (Che qu), Cicer arietinum (Cice ar), Cryptomeria japonica (TLP-Cj), Cupressus arizonica (TLP-Ca), Cupressus sempervirens (Cup s 3), Fragaria × ananassa (Frag an), Ficus awkeotsang (Ficu aw), Glycine max (Glyc ma), Helianthus annuus (Heli an), Hordeum vulgare (Hord vu), Juniperus ashei (Jun a 3), Juniperus rigida (TLP-Jr), Linum usitatissimum (Linu us), Lycopersicum esculentum (Lyco es), Malus domestica (Mal d 2), Musa acuminata (Mus a 4), Nicotiana glauca (Nico gl), Nicotiana tabacum (osmotin), Petunia hybrida (Petu hy), Prunus avium (Pru av 2), Prunus persica (Pru p 2), Pseudotsuga menziesii (Pseu me), Pyrus pyriformis (Pyr py 2), Sambucus nigra (Samb ni), Sequoia sempervirens (TLP-Ss), Solanum commersonii (Sola co), Solanum dulcamara (Sola du), Solanum nigrum (Sola ni), Solanum phyreja (Sola ph), Solanum tuberosum (Sola tu), Taxodium distichum (TLP-Txm), Thaumatococcus daniellii (thaumatin), Thuja occidentalis (TLP-To), Vitis riparia (Viti ri), Vitis vinifera (Vit v TLP), Zea mays (zeamatin). Thaumatin-like proteins belonging to the same plant family are boxed and the different family boxes are labeled and differently colored.
4. Discussion
As a typical PR-5 thaumatin-like protein, the modeled Cup s 3 allergen exhibits the canonical three-dimensional conformation made of three domains delineating a central electronegative catalytic groove responsible for the β-glucanase activity (Figure 1A,B). Three distinct sequential IgE-binding epitopic peptides distributed in four epitopic regions were identified on the molecular surface of Cup s 3. Epitope #2, named according to the results of SPOT techniques, splits into two exposed epitopic regions because one part is buried in the molecule most likely due to the eight disulfide bridges. In this respect, although the SPOT technique used herein clearly identified three epitopic regions, the existence of undetected additional discontinuous epitopes cannot be ruled out because of the very folded nature of these allergens. Epitopes are essentially distributed on the face opposite to that containing the catalytic groove. Two of these epitopes (1 and the larger part of 2) occur in domain I and epitope 3 in domain II. Epitope 3 possesses a N-glycosylation site, that could possibly correspond to a glycotope that does not prevent the induction of IgE specific to the underlying peptide. Alternatively, the TLP allergen might consist of two isoforms, glycosylated and not glycosylated. As could be expected, the protruding hairpin domain III is an epitope (smaller part of epitope 2). The identified peptides in the SPOT techniques were confirmed to be accessible on the intact allergen since they were able to significantly inhibit the binding of the specific serum IgE to the TLP allergen in its native form in pollen extract.
Conformational analysis of the IgE-binding peptides identified on Cup s 3 revealed striking similarities with the corresponding regions of the modeled regions from other closely related Cupressaceae TLP, from Cryptomeria japonica and from Thuja occidentalis (Figure 4). These conformational mimicries most probably account for the IgE-binding cross-reactivity reported between different Cupressaceae allergens [26,30,44,45]. In agreement with these conformational similarities, some sequence similarity was observed among the epitopic regions of the thaumatin-like allergens from different Cupressaceae. In this respect, four positively IgE-reacting peptides corresponding to sequence stretches 120–131, 132–145, 155–145, and 169–179 have been previously identified on the molecular surface of Jun a 3 from the mountain cedar pollen [46]. Epitope 3 (sequence strech 156–173) of Cup s 3 overlaps with two of the major IgE-binding epitopic regions (sequence stretches 155–165 and 169–173) characterized on Jun a 3. The Jun a 3 epitope is located in the α helix-containing domain II [46]. Cross-reactivity of Cup s 3 and Jun a 3 was demonstrated by ELISA inhibition assays in which the binding of IgE to Cup s 3 was inhibited by a preincubation in the presence of purified Jun a 3 [22]. Cross reactivities between TLPs from various plant species were also studied in TLP sensitized patients from restricted areas in Spain [24]. However, sensitization to Cup a 3 was barely represented and poorly significant “co-sensitization links” were observed between Cup a 3 and Pru av 2 or Mald 2, TLPs from cherry and apple, respectively. A similar value of “co-sensitization link” was also reported with Ana c 2, a cystein protease from pineapple. Whether these “co-sensitization links” might be attributed to cross-reactive oligosaccharide moieties present on TLPs and/or cystein protease was not studied.
The possible role of N-glycans with respect to the allergenicity and IgE-binding cross-reactivity of Cupressaceae allergens was deeply investigated by Italian authors using different allergens as N-glycosylated probes [47,48,49,50]. Their results indicated that a significant proportion of IgE from CPAP specifically recognizes the N-glycan moieties of the different glyco-allergens, pectate lyase (group 1), and polygalacturonase (group 2) [51]. However, the cross-reactivities observed for group 1 and group 2 allergens of Cupressaceae pollen do not rely only on oligosaccharides moieties since AA sequences are closely related, generating similar peptidic epitopic regions. This is particularly exemplified in the Cup s 3 results reported herein where epitopic region 3 of Cup s 3, including a glycosylation site, exhibited the most intense IgE-binding reactivity.
In agreement with the close structural similarities of their epitopic regions, PR-5 TLP allergens from the Cupressaceae, Taxaceae, and Taxodiaceae form a very homogeneous clade distinct from other PR-5 TLP family clades, as shown in the phylogenetic tree built up for the plant PR5 TLP (Figure 5). The lack of close relationship with other families of PR-5 TLP within the phylogenetic tree suggests that peptidic cross-reactivities between Cupressaceae TLPs and TLPs from others plant species are unlikely to occur.
5. Conclusions
The identification of putative IgE-binding cross-reactive epitopes of Cup s 3 and other closely related PR-5 TLP allergens from the Cupressaceae, Taxaceae, and Taxodiaceae pollen has been completed. The position in the phylogenetic tree of these pollen allergens, which form a distinct clade separated from other PR-5 TLP family clades, suggests that their IgE-binding cross-reactivity (1) is limited to the Cupressaceae and the closely related Taxaceae and Taxodiaceae families, irrespective of their sequential and structural similarities, and (2) should exclude cross-reactivity to plant-derived foods and food products.
Author Contributions
Conceptualization, P.R.; methodology, A.B.; software, C.N., C.G. and P.R.; validation, A.B. and P.R.; investigation, C.N. and C.G.; resources, A.B., H.S. and P.P.; data curation, H.S and P.P.; writing—original draft preparation, P.R.; writing—review and editing, P.R.; supervision, P.R.; project administration, A.B. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Informed consent was obtained from all subjects involved in the study.
Acknowledgments
The authors gratefully thank Beatrice Saggio (Laboratoire de Biologie, Hôpital Lenval, Nice, France) who provided us with the sera from cypress allergic patients, used as probes in SPOT experiments.
Conflicts of Interest
The authors declare no conflict of interest.
References
- D’Amato, G.; Cecchi, L.; Bonini, S.; Nunes, C.; Annesi-Maesano, I.; Behrendt, H.; Liccardi, G.; Popov, T.; Van Cauwenberge, P. Allergenic pollen and pollen allergy in Europe. Allergy 2007, 62, 976–990. [Google Scholar] [CrossRef]
- Bibi, H.; Shoseyov, D.; Feigenbaum, D.; Nir, P.; Shiachi, R.; Scharff, S.; Peled, R. Comparison of positive allergy skin tests among asthmatic children from rural and urban areas living within small geographic area. Ann. Allergy Asthma Immunol. 2002, 88, 416–420. [Google Scholar] [CrossRef]
- Gonianakis, M.I.; Baritaki, M.A.; Neonakis, I.K.; Gonianakis, I.M.; Kypriotakis, Z.; Darivianaki, E.; Bouros, D.; Kontou-Filli, K. A 10-year aerobiological study (1994–2003) in the Mediterranean island of Crete, Greece: Trees, aerobiologic data, and botanical and clinical correlations. Allergy Asthma Proc. 2006, 27, 371–377. [Google Scholar] [CrossRef]
- Pérez-Badia, R.; Rapp, A.; Vaquero, C.; Fernández-González, F. Aerobiological study in east-central Iberian Peninsula: Pollen diversity and dynamics for major taxa. Ann. Agric. Environ. Med. 2011, 18, 99–111. [Google Scholar]
- Lamy, E.; Savournin, C.; Balansard, G. Ten years of cypress pollen counts in Marseille. Allergol. Immunopathol. 2001, 33, 103–104. [Google Scholar]
- Kishikawa, R.; Kotoh, E.; Oshikawa, C.; Soh, N.; Shimoda, T.; Saito, A.; Sahashi, N.; Enomoto, T.; Usami, A.; Teranishi, H.; et al. Longitudinal monitoring of tree airborne pollen in japan. Arerugi 2017, 66, 97–111. [Google Scholar] [PubMed]
- Pham, N.H.; Baldo, B.A.; Bass, D.J. Cypress pollen allergy. Identification of allergens and crossreactivity between divergent species. Clin. Exp. Allergy 1994, 24, 558–565. [Google Scholar] [CrossRef]
- Mohanty, R.P.; Buchheim, M.A.; Levetin, E. Molecular approaches for the analysis of airborne pollen: A case study of Juniperus pollen. Ann. Allergy Asthma Immunol. 2017, 118, 204–211.e202. [Google Scholar] [CrossRef] [PubMed]
- Guerin, B.; Kanny, G.; Terrasse, G.; Guyot, J.; Moneret-Vautrin, D. Allergic Rhinitis to Thuja Pollen. Int. Arch. Allergy Immunol. 1996, 110, 91–94. [Google Scholar] [CrossRef] [PubMed]
- Iacovacci, P.; Afferni, C.; Barletta, B.; Tinghino, R.; Di Felice, G.; Pini, C.; Mari, A. Juniperus oxycedrus: A new allergenic pollen from the Cupressaceae family. J. Allergy Clin. Immunol. 1998, 101, 755–761. [Google Scholar] [CrossRef] [PubMed]
- Sneller, M.R.; Hayes, H.D.; Pinnas, J.L. Pollen changes during five decades of urbanization in Tucson, Arizona. Ann. Allergy 1993, 71, 519–524. [Google Scholar] [PubMed]
- Charpin, D.; Calleja, M.; Pichot, C.; Penel, V.; Hugues, B.; Poncet, P. Cypress pollen allergy. Rev. Mal. Respir. 2013, 30, 868–878. [Google Scholar] [CrossRef]
- Charpin, D.; Pichot, C.; Belmonte, J.; Sutra, J.-P.; Zidkova, J.; Chanez, P.; Shahali, Y.; Sénéchal, H.; Poncet, P. Cypress Pollinosis: From Tree to Clinic. Clin. Rev. Allergy Immunol. 2019, 56, 174–195. [Google Scholar] [CrossRef]
- Cortegano, I.; Civantos, E.; Aceituno, E.; Del Moral, A.; López, E.; Lombardero, M.; Del Pozo, V.; Lahoz, C. Cloning and expression of a major allergen from Cupressus arizonica pollen, Cup a 3, a PR-5 protein expressed under polluted environment. Allergy 2004, 59, 485–490. [Google Scholar] [CrossRef]
- Sénéchal, H.; Šantrůček, J.; Melčová, M.; Svoboda, P.; Zídková, J.; Charpin, D.; Guilloux, L.; Shahali, Y.; Selva, M.-A.; Couderc, R.; et al. A new allergen family involved in pollen food-associated syndrome: Snakin/gibberellin-regulated proteins. J. Allergy Clin. Immunol. 2018, 141, 411–414.e4. [Google Scholar] [CrossRef]
- Iizuka, T.; Takei, M.; Saito, Y.; Rumi, F.; Zheng, J.; Lu, X.; Chafey, P.; Broussard, C.; Guilloux-Assalet, L.; Charpin, D.; et al. Gibberellin-regulated protein sensitization in Japanese cedar (Cryptomeria japonica) pollen allergic Japanese cohorts. Allergy 2021, 76, 2297–2302. [Google Scholar] [CrossRef]
- Ehrenberg, A.E.; Klingebiel, C.; Östling, J.; Larsson, H.; Mattsson, L.; Vitte, J.; Lidholm, J. Characterization of a 7 kDa pollen allergen belonging to the gibberellin-regulated protein family from three Cupressaceae species. Clin. Exp. Allergy 2020, 50, 964–972. [Google Scholar] [CrossRef]
- Sénéchal, H.; Keykhosravi, S.; Couderc, R.; Selva, M.-A.; Shahali, Y.; Aizawa, T.; Busnel, J.-M.; Arif, R.; Mercier, I.; Pham-Thi, N.; et al. Pollen/Fruit Syndrome: Clinical Relevance of the Cypress Pollen Allergenic Gibberellin-Regulated Protein. Allergy Asthma Immunol. Res. 2019, 11, 143–151. [Google Scholar] [CrossRef]
- Takei, M.; Nin, C.; Iizuka, T.; Pawlikowski, M.; Selva, M.-A.; Chantran, Y.; Nakajima, Y.; Zheng, J.; Aizawa, T.; Ebisawa, M.; et al. Capsicum Allergy: Involvement of Cap a 7, a New Clinically Relevant Gibberellin-Regulated Protein Cross-Reactive With Cry j 7, the Gibberellin-Regulated Protein From Japanese Cedar Pollen. Allergy Asthma Immunol. Res. 2022, 14, 328–338. [Google Scholar] [CrossRef]
- Panzani, R.; Yasueda, H.; Shimizu, T.; Shida, T. Cross-reactivity between the pollens of Cupressus sempervirens (common cypress) and of Cryptomeria japonica (Japanese cedar). Ann. Allergy 1986, 57, 26–30. [Google Scholar]
- Breiteneder, H. Thaumatin-like proteins—A new family of pollen and fruit allergens. Allergy 2004, 59, 479–481. [Google Scholar] [CrossRef]
- Ivanciuc, O.; Mathura, V.; Midoro-Horiuti, T.; Braun, W.; Goldblum, R.M.; Schein, C.H. Detecting Potential IgE-Reactive Sites on Food Proteins Using a Sequence and Structure Database, SDAP-Food. J. Agric. Food Chem. 2003, 51, 4830–4837. [Google Scholar] [CrossRef]
- Fujimura, T.; Futamura, N.; Midoro-Horiuti, T.; Togawa, A.; Goldblum, R.M.; Yasueda, H.; Saito, A.; Shinohara, K.; Masuda, K.; Kurata, K.; et al. Isolation and characterization of native Cry j 3 from Japanese cedar (Cryptomeria japonica) pollen. Allergy 2007, 62, 547–553. [Google Scholar] [CrossRef]
- Palacin, A.; Rivas, L.A.; Gómez-Casado, C.; Aguirre, J.; Tordesillas, L.; Bartra, J.; Blanco, C.; Carrillo, T.; Cuesta-Herranz, J.; Bonny, J.A.C.; et al. The Involvement of Thaumatin-Like Proteins in Plant Food Cross-Reactivity: A Multicenter Study Using a Specific Protein Microarray. PLoS ONE 2012, 7, e44088. [Google Scholar] [CrossRef]
- Midoro-Horiuti, T.; Goldblum, R.M.; Kurosky, A.; Wood, T.G.; Brooks, E.G. Variable Expression of Pathogenesis-Related Protein Allergen in Mountain Cedar (Juniperus ashei) Pollen. J. Immunol. 2000, 164, 2188–2192. [Google Scholar] [CrossRef]
- Togawa, A.; Panzani, R.C.; Garza, M.A.; Kishikawa, R.; Goldblum, R.M.; Midoro-Horiuti, T. Identification of Italian cypress (Cupressus sempervirens) pollen allergen Cup s 3 using homology and cross-reactivity. Ann. Allergy Asthma Immunol. 2006, 97, 336–342. [Google Scholar] [CrossRef]
- Smith, P.K.; Krohn, R.I.; Hermanson, G.T.; Mallia, A.K.; Gartner, F.H.; Provenzano, M.D.; Fujimoto, E.K.; Goeke, N.M.; Olson, B.J.; Klenk, D.C. Measurement of protein using bicinchoninic acid. Anal. Biochem. 1987, 150, 76–85. [Google Scholar] [CrossRef]
- Barre, A.; Peumans, W.J.; Menu-Bouaouiche, L.; Van Damme, E.J.M.; May, G.D.; Herrera, A.F.; Van Leuven, F.; Rougé, P. Purification and structural analysis of an abundant thaumatin-like protein from ripe banana fruit. Planta 2000, 211, 791–799. [Google Scholar] [CrossRef]
- Tastet, C.; Lescuyer, P.; Diemer, H.; Luche, S.; Van Dorsselaer, A.; Rabilloud, T. A versatile electrophoresis system for the analysis of high- and low-molecular-weight proteins. Electrophoresis 2003, 24, 1787–1794. [Google Scholar] [CrossRef]
- Barre, A.; Sénéchal, H.; Nguyen, C.; Granier, C.; Rougé, P.; Poncet, P. Identification of Potential IgE-Binding Epitopes Contributing to the Cross-Reactivity of the Major Cupressaceae Pectate-Lyase Pollen Allergens (Group 1). Allergies 2022, 2, 106–118. [Google Scholar] [CrossRef]
- Frank, R. Spot-synthesis: An easy technique for the positionally addressable paralel chemical synthesis on a membrane support. Tetrehedron 1992, 48, 9217–9232. [Google Scholar] [CrossRef]
- Laune, D.; Molina, F.; Ferrières, G.; Villard, S.; Bès, C.; Rieunier, F.; Chardès, T.; Granier, C. Application of the Spot method to the identification of peptides and amino acids from the antibody paratope that contribute to antigen binding. J. Immunol. Methods 2002, 267, 53–70. [Google Scholar] [CrossRef]
- Thompson, J.D.; Gibson, T.J.; Plewniak, F.; Jeanmougin, F.; Higgins, D.G. The CLUSTAL_X windows interface: Flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acids Res. 1997, 25, 4876–4882. [Google Scholar] [CrossRef] [PubMed]
- Krieger, E.; Koraimann, G.; Vriend, G. Increasing the precision of comparative models with YASARA NOVA—A self-parameterizing force field. Proteins 2002, 47, 393–402. [Google Scholar] [CrossRef]
- Leone, P.; Menu-Bouaouiche, L.; Peumans, W.J.; Payan, F.; Barre, A.; Roussel, A.; Van Damme, E.; Rougé, P. Resolution of the structure of the allergenic and antifungal banana fruit thaumatin-like protein at 1.7-Å. Biochimie 2006, 88, 45–52. [Google Scholar] [CrossRef]
- Marangon, M.; Van Sluyter, S.C.; Waters, E.J.; Menz, R.I. Structure of Haze Forming Proteins in White Wines: Vitis vinifera Thaumatin-Like Proteins. PLoS ONE 2014, 9, e113757. [Google Scholar] [CrossRef] [PubMed]
- Batalia, M.A.; Monzingo, A.F.; Ernst, S.; Roberts, W.; Robertus, J.D. The crystal structure of the antifungal protein zeamatin, a member of the thaumatin-like, PR-5 protein family. Nat. Struct. Biol. 1996, 3, 19–23. [Google Scholar] [CrossRef]
- Laskowski, R.A.; MacArthur, M.W.; Moss, D.S.; Thornton, J.M. PROCHECK: A program to check the stereochemical quality of protein structures. J. Appl. Cryst. 1993, 26, 283–291. [Google Scholar] [CrossRef]
- Melo, F.; Feytmans, E. Assessing protein structures with a non-local atomic interaction energy. J. Mol. Biol. 1998, 277, 1141–1152. [Google Scholar] [CrossRef] [PubMed]
- Arnold, K.; Bordoli, L.; Kopp, J.; Schwede, T. The SWISS-MODEL workspace: A web-based environment for protein structure homology modelling. Bioinformatics 2006, 22, 195–201. [Google Scholar] [CrossRef] [PubMed]
- Benkert, P.; Biasini, M.; Schwede, T. Toward the estimation of the absolute quality of individual protein structure models. Bioinformatics 2011, 27, 343–350. [Google Scholar] [CrossRef] [PubMed]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Couch, G.S.; Greenblatt, D.M.; Meng, E.C.; Ferrin, T.E. UCSF Chimera—A visualization system for exploratory research and analysis. J. Comput. Chem. 2004, 25, 1605–1612. [Google Scholar] [CrossRef] [PubMed]
- Menu-Bouaouiche, L.; Vriet, C.; Peumans, W.J.; Barre, A.; Van Damme, E.J.M.; Rougé, P. A molecular basis for the endo-beta 1,3-glucanase activity of the thaumatin-like proteins from edible fruits. Biochimie 2003, 85, 123–131. [Google Scholar] [CrossRef] [PubMed]
- Bistoni, O.; Emiliani, C.; Agea, E.; Russano, A.M.; Mencarelli, S.; Orlacchio, A.; Spinozzi, F. Biochemical and immunological characterization of pollen-derived beta-galactosidase reveals a new cross-reactive class of allergens among Mediterranean trees. Int. Arch. Allergy Immunol. 2005, 136, 123–133. [Google Scholar] [CrossRef] [PubMed]
- Shahali, Y.; Sutra, J.-P.; Charpin, D.; Mari, A.; Guilloux, L.; Sénéchal, H.; Poncet, P. Differential IgE sensitization to cypress pollen associated to a basic allergen of 14 kDa. FEBS J. 2012, 279, 1445–1455. [Google Scholar] [CrossRef]
- Soman, K.V.; Midoro-Horiuti, T.; Ferreon, J.; Goldblum, R.M.; Brooks, E.G.; Kurosky, A.; Braun, W.; Schein, C.H. Homology Modeling and Characterization of IgE Binding Epitopes of Mountain Cedar Allergen Jun a 3. Biophys. J. 2000, 79, 161–169. [Google Scholar] [CrossRef]
- Afferni, C.; Iacovacci, P.; Barletta, B.; Di Felice, G.; Tinghino, R.; Mari, A.; Pini, C. Role of carbohydrate moieties in IgE binding to allergenic components of Cupressus arizonica pollen extract. Clin. Exp. Allergy 1999, 29, 1087–1094. [Google Scholar] [CrossRef]
- Barletta, B.; Tinghino, R.; Corinti, S.; Afferni, C.; Iacovacci, P.; Mari, A.; Pini, C.; Di Felice, G. Arizona cypress (Cupressus arizonica) pollen allergens. Identification of crossreactive periodate-resistant and sensitive epitopes with monoclonal antibodies. Allergy 1998, 53, 586–593. [Google Scholar] [CrossRef]
- Canini, A.; Giovinazzi, J.; Iacovacci, P.; Pini, C.; Caiola, M.G. Localisation of a carbohydrate epitope recognised by human IgE in pollen of Cupressaceae. J. Plant Res. 2004, 117, 147–153. [Google Scholar] [CrossRef]
- Iacovacci, P.; Afferni, C.; Butteroni, C.; Pironi, L.; Puggioni, E.M.R.; Orlandi, A.; Barletta, B.; Tinghino, R.; Ariano, R.; Panzani, R.C.; et al. Comparison between the native glycosylated and the recombinant Cup a1 allergen: Role of carbohydrates in the histamine release from basophils. Clin. Exp. Allergy 2002, 32, 1620–1627. [Google Scholar] [CrossRef]
- Shahali, Y.; Sutra, J.; Hilger, C.; Swiontek, K.; Haddad, I.; Vinh, J.; Guilloux, L.; Charpin, D.; Sénéchal, H.; Poncet, P. Identification of a polygalacturonase (Cup s 2) as the major CCD-bearing allergen in Cupressus sempervirens pollen. Allergy 2017, 72, 1806–1810. [Google Scholar] [CrossRef] [PubMed]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).