
Screening and Selection for Herbicide Tolerance among Diverse Tomato Germplasms


Abstract
:1. Introduction
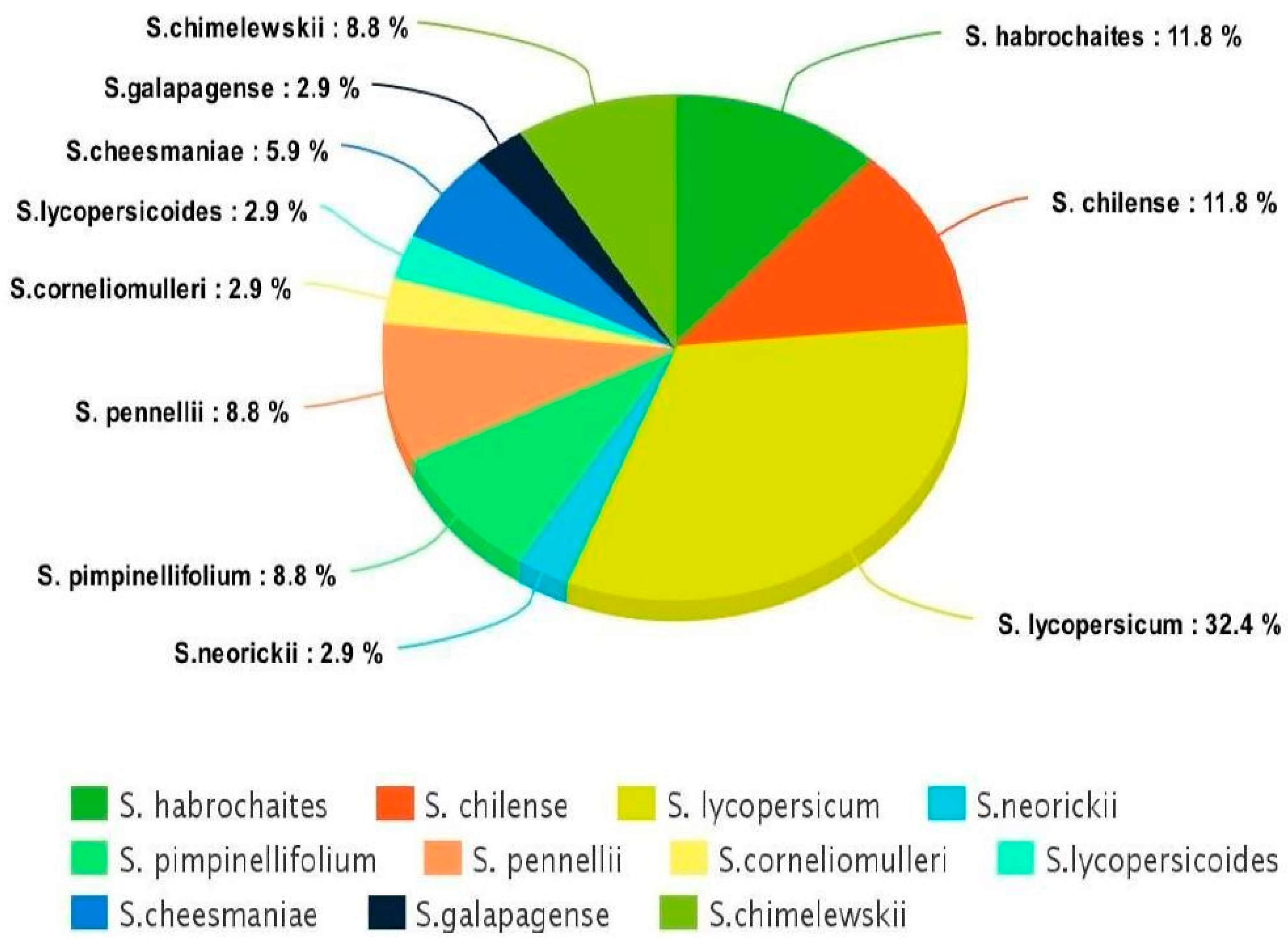
2. Materials and Methods
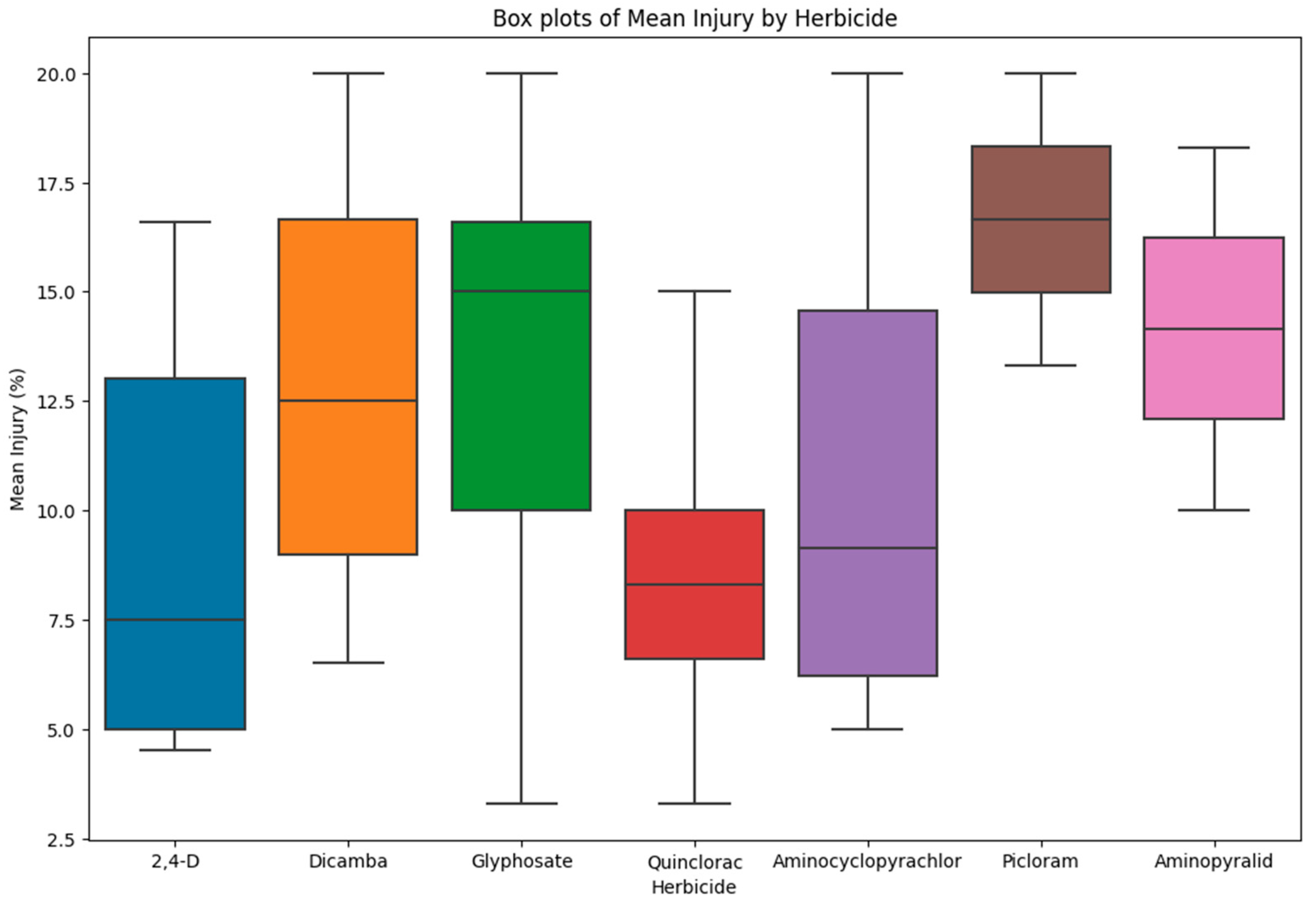
3. Results and Discussion
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Nomenclature
References
- USDA—National Agricultural Statistics Service Census of Agriculture (Year); USDA: Washington, DC, USA, 2012.
- Bramley, P.M. Is Lycopene Beneficial to Human Health? Phytochemistry 2000, 54, 233–236. [Google Scholar] [CrossRef]
- Vecchia, C.L. Mediterranean Epidemiological Evidence on Tomatoes and the Prevention of Digestive-Tract Cancers. Proc. Soc. Exp. Biol. Med. 1998, 218, 125–128. [Google Scholar] [CrossRef]
- Pan, H.Y.; Fisher, K.J.; Nichols, M.A. The Effect of Mulch and Row Covers on Yield of Process Tomatoes. In Proceedings of the VI International Symposium on Processing Tomato & Workshop on Irrigation & Fertigation of Processing Tomato 487, Pamplona, Spain, 25–29 May 1998; pp. 145–150. [Google Scholar]
- Webster, T.M. Weed Survey-Southern States. Proc. South. Weed Sci. Soc. 2002, 55, 237–254. [Google Scholar]
- Bangarwa, S.K.; Norsworthy, J.K.; Gbur, E.E. Integration of a Brassicaceae Cover Crop with Herbicides in Plasticulture Tomato. Weed Technol. 2009, 23, 280–286. [Google Scholar] [CrossRef]
- Scott, R.C.; Boyd, J.W.; Smith, K.L.; Selden, G.; Norsworthy, J.K. Recommended Chemicals for Weed and Brush Control-MP44; Cooperative Extension Service, University of Arkansas: Fayetteville, AR, USA, 2012. [Google Scholar]
- Lovelace, M.L.; Hoagland, R.E.; Talbert, R.E.; Scherder, E.F. Influence of Simulated Quinclorac Drift on the Accumulation and Movement of Herbicide in Tomato (Lycopersicon esculentum) Plants. J. Agric. Food Chem. 2009, 57, 6349–6355. [Google Scholar] [CrossRef]
- Clark, J. Tupelo Organic Farmer Loses Crop to Herbicide Drift. Dly. J. 2013. Available online: https://www.djournal.com/news/tupelo-organic-farmer-loses-crop-to-herbicide-drift/article_b0bf99b1-989a-51af-9148-747cb69332cf.html (accessed on 20 December 2017).
- Foresman, C.; Glasgow, L. US Grower Perceptions and Experiences with Glyphosate-Resistant Weeds. Pest Manag. Sci. Former. Pestic. Sci. 2008, 64, 388–391. [Google Scholar] [CrossRef]
- Benbrook, C.M. Impacts of Genetically Engineered Crops on Pesticide Use in the US–the First Sixteen Years. Environ. Sci. Eur. 2012, 24, 24. [Google Scholar] [CrossRef]
- Heap, I. The International Survey of Herbicide Resistant Weeds. Available online: www.weedscience.org (accessed on 18 June 2017).
- Caseley, J.C.; Coupland, D. Environmental and Plant Factors Affecting Glyphosate Uptake, Movement and Activity. In Herbicide Glyphosate; Grossbard, E., Atkinson, D., Eds.; Butterworth & Co.: London, UK, 1985; pp. 490–491. [Google Scholar]
- Ovidi, E.; Gambellini, G.; Taddei, A.R.; Cai, G.; Del Casino, C.; Ceci, M.; Rondini, S.; Tiezzi, A. Herbicides and the Microtubular Apparatus of Nicotiana Tabacum Pollen Tube: Immunofluorescence and Immunogold Labelling Studies. Toxicol. Vitr. 2001, 15, 143–151. [Google Scholar] [CrossRef]
- Jordan, T.N.; Romanowski, R.R. Comparison of Dicamba and 2, 4-D Injury to Field-Grown Tomatoes1. HortScience 1974, 9, 74–75. [Google Scholar] [CrossRef]
- Talbert, R.E.; Burgos, N.R. History and Management of Herbicide-Resistant Barnyardgrass (Echinochloa crus-galli) in Arkansas Rice. Weed Technol. 2007, 21, 324–331. [Google Scholar] [CrossRef]
- De Barreda, D.G.; Lorenzo, E.; Carbonell, E.A.; Cases, B.; Muñoz, N. Use of Tomato (Lycopersicon esculentum) Seedlings to Detect Bensulfuron and Quinclorac Residues in Water. Weed Technol. 1993, 7, 376–381. [Google Scholar] [CrossRef]
- Grossmann, K. Quinclorac Belongs to a New Class of Highly Selective Auxin Herbicides. Weed Sci. 1998, 46, 707–716. [Google Scholar] [CrossRef]
- Barrentine, W.L.; Street, J.E.; Eddie, J. Soybean Response to Quinclorac and Triclopyr; Department of Information Services, Division of Agriculture, Forestry, and Veterinary Medicine, Mississippi State University: Mississippi State, MS, USA, 1993. [Google Scholar]
- Flessner, M.L.; McElroy, J.S.; Cardoso, L.A.; Martins, D. Simulated Spray Drift of Aminocyclopyrachlor on Cantaloupe, Eggplant, and Cotton. Weed Technol. 2012, 26, 724–730. [Google Scholar] [CrossRef]
- Lewis, D.F.; Hoyle, S.T.; Fisher, L.R.; Yelverton, F.H.; Richardson, R.J. Effect of Simulated Aminocyclopyrachlor Drift on Flue-Cured Tobacco. Weed Technol. 2011, 25, 609–615. [Google Scholar] [CrossRef]
- Smith, L.L., Jr.; Geronimo, J. Response of Seven Crops to Foliar Applications of Six Auxin-like Herbicides. Down Earth A Rev. Agric. Chem. Prog. 1984, 40, 25–32. [Google Scholar]
- Fagliari, J.R.; de Oliveira, R.S., Jr.; Constantin, J. Impact of Sublethal Doses of 2, 4-D, Simulating Drift, on Tomato Yield. J. Environ. Sci. Health Part B 2005, 40, 201–206. [Google Scholar] [CrossRef]
- Kruger, G.R.; Johnson, W.G.; Doohan, D.J.; Weller, S.C. Dose Response of Glyphosate and Dicamba on Tomato (Lycopersicon esculentum) Injury. Weed Technol. 2012, 26, 256–260. [Google Scholar] [CrossRef]
- Lovelace, M.L.; Talbert, R.E.; Scherder, E.F.; Hoagland, R.E. Effects of Multiple Applications of Simulated Quinclorac Drift Rates on Tomato. Weed Sci. 2007, 55, 169–177. [Google Scholar] [CrossRef]
- Israel, T.D.; Rhodes, G.N.; Wszelaki, A. Diagnosing Suspected Off-Target Herbicide Damage to Tomato; University of Tennessee Knoxville: Knoxville, TN, USA, 2013. [Google Scholar]
- Wax, L.; Knuth, L.A.; Slife, F.W. Response of Soybeans to 2, 4-D, Dicamba, and Picloram. Weed Sci. 1969, 17, 388–393. [Google Scholar] [CrossRef]
- Robertson, L.D.; Labate, J.A. Genetic Resources of Tomato (Lycopersicon esculentum Mill.) and Wild Relatives. Genet. Improv. Solanaceous Crops 2006, 2, 25–75. [Google Scholar]
- Fobes, J.F.; Mudd, J.B.; Marsden, M.P. Epicuticular Lipid Accumulation on the Leaves of Lycopersicon pennellii (Corr.) D’Arcy and Lycopersicon esculentum Mill. Plant Physiol. 1985, 77, 567–570. [Google Scholar] [CrossRef]
- Ling, K.-S.; Scott, J.W. Sources of Resistance to Pepino Mosaic Virus in Tomato Accessions. Plant Dis. 2007, 91, 749–753. [Google Scholar] [CrossRef]
- Cassol, T.; St. Clair, D.A. Inheritance of Resistance to Blackmold (Alternaria alternata (Fr.) Keissler) in Two Interspecific Crosses of Tomato (Lycopersicon esculentum × L. cheesmanii f. typicum). Theor. Appl. Genet. 1994, 88, 581–588. [Google Scholar] [CrossRef]
- Atkinson, N.J.; Urwin, P.E. The Interaction of Plant Biotic and Abiotic Stresses: From Genes to the Field. J. Exp. Bot. 2012, 63, 3523–3543. [Google Scholar] [CrossRef]
- Pastori, G.M.; Foyer, C.H. Common Components, Networks, and Pathways of Cross-Tolerance to Stress. The Central Role of “Redox” and Abscisic Acid-Mediated Controls. Plant Physiol. 2002, 129, 460–468. [Google Scholar] [CrossRef]
- Ignatova, S.I.; Gorshkova, N.S.; Tereshonkova, T.A. Resistance of Tomato F1 Hybrids to Grey Mold. Acta Physiol. Plant 2000, 22, 326–328. [Google Scholar] [CrossRef]
- Bournival, B.L.; Scott, J.W.; Vallejos, C.E. An Isozyme Marker for Resistance to Race 3 of Fusarium oxysporum f. Sp. lycopersici in Tomato. Theoret. Appl. Genet. 1989, 78, 489–494. [Google Scholar] [CrossRef]
- Labate, J.A.; Grandillo, S.; Fulton, T.; Muños, S.; Caicedo, A.L.; Peralta, I.; Ji, Y.; Chetelat, R.T.; Scott, J.W.; Gonzalo, M.J. Tomato. In Vegetables; Springer: Berlin/Heidelberg, Germany, 2007; pp. 1–125. [Google Scholar]
- Francis, D.M.; Kabelka, E.; Bell, J.; Franchino, B.; St. Clair, D. Resistance to Bacterial Canker in Tomato (Lycopersicon hirsutum LA407) and Its Progeny Derived from Crosses to L. esculentum. Plant Dis. 2001, 85, 1171–1176. [Google Scholar] [CrossRef]
- Rakha, M.; Bouba, N.; Ramasamy, S.; Regnard, J.-L.; Hanson, P. Evaluation of Wild Tomato Accessions (Solanum spp.) for Resistance to Two-Spotted Spider Mite (Tetranychus urticae Koch) Based on Trichome Type and Acylsugar Content. Genet. Resour. Crop Evol. 2017, 64, 1011–1022. [Google Scholar] [CrossRef]
- O’Connell, M.A.; Medina, A.L.; Sanchez Pena, P.; Treviño, M.B. Molecular Genetics of Drought Resistance Response in Tomato and Related Species. Genet. Improv. Solanaceous Crops 2006, 2, 261–283. [Google Scholar]
- Ciccarese, F.; Amenduni, M.; Schiavone, D.; Cirulli, M. Occurrence and Inheritance of Resistance to Powdery Mildew (Oidium lycopersici) in Lycopersicon Species. Plant Pathol. 1998, 47, 417–419. [Google Scholar] [CrossRef]
- Cillo, F.; Pasciuto, M.M.; De Giovanni, C.; Finetti-Sialer, M.M.; Ricciardi, L.; Gallitelli, D. Response of Tomato and Its Wild Relatives in the Genus Solanum to Cucumber Mosaic Virus and Satellite RNA Combinations. J. Gen. Virol. 2007, 88, 3166–3176. [Google Scholar] [CrossRef]
- Lebeda, A.; Mieslerová, B.; Petřivalskỳ, M.; Luhová, L.; Špundová, M.; Sedlářová, M.; Nožková-Hlaváčková, V.; Pink, D.A. Resistance Mechanisms of Wild Tomato Germplasm to Infection of Oidium Neolycopersici. Eur. J. Plant Pathol. 2014, 138, 569–596. [Google Scholar] [CrossRef]
- Arms, E.M.; Bloom, A.J.; St. Clair, D.A. High-Resolution Mapping of a Major Effect QTL from Wild Tomato Solanum Habrochaites That Influences Water Relations under Root Chilling. Theor. Appl. Genet. 2015, 128, 1713–1724. [Google Scholar] [CrossRef]
- Levy, J.; Tamborindeguy, C. Solanum habrochaites, a Potential Source of Resistance against Bactericera cockerelli (Hemiptera: Triozidae) and “Candidatus Liberibacter Solanacearum”. J. Econ. Entomol. 2014, 107, 1187–1193. [Google Scholar] [CrossRef]
- Nosenko, T.; Böndel, K.B.; Kumpfmüller, G.; Stephan, W. Adaptation to Low Temperatures in the Wild Tomato Species Solanum Chilense. Mol. Ecol. 2016, 25, 2853–2869. [Google Scholar] [CrossRef]
- Bolger, A.; Scossa, F.; Bolger, M.E.; Lanz, C.; Maumus, F.; Tohge, T.; Quesneville, H.; Alseekh, S.; Sørensen, I.; Lichtenstein, G. The Genome of the Stress-Tolerant Wild Tomato Species Solanum Pennellii. Nat. Genet. 2014, 46, 1034–1038. [Google Scholar] [CrossRef]
- Sharma, A.; Zhang, L.; Nino-Liu, D.; Ashrafi, H.; Foolad, M.R. A Solanum lycopersicum × Solanum pimpinellifolium Linkage Map of Tomato Displaying Genomic Locations of R-Genes, RGAs, and Candidate Resistance/Defense-Response ESTs. Int. J. Plant Genom. 2008, 2008, 926090. [Google Scholar]


{kind=link}
{kind=link}
{kind=link}
| Herbicide | Trade Name | Rate Used | Drift Rates (g ae ha−1) |
|---|---|---|---|
| 2,4-D | Weedar-64® | 0.01X | 11.2 |
| Dicamba | Clarity® | 0.01X | 2.8 |
| Glyphosate | Roundup Powermax® | 0.01X | 8.4 |
| Quinclorac | Facet L® | 0.01X | 39.2 |
| Aminopyralid | Milestone® | 0.05X | 6.15 |
| Aminocyclopyrachlor | Streamline® | 0.05X | 15.65 |
| Picloram | Tordon® | 0.05X | 28.0 |
| Tomato Injury (%) | Tomato Symptomology |
|---|---|
| 0–10 | No symptoms to slight injury, no growth reduction |
| 10–30 | Slight-to-moderate injury. Slight growth reduction |
| 30–50 | Epinastic and twisting of leaves in auxin herbicides, slight-to-moderate growth reduction, white/yellow discoloration at the base |
| 50–70 | Moderate-to-severe injury, callusing on the stems in auxins, and growth reduction |
| 70–95 | Severe injury and no growth |
| 95–100 | Near to death or death |
| Herbicide | Sum of Squares | Mean Square | F Ratio | Prob > F |
|---|---|---|---|---|
| 2,4-D | 97,293.27 | 1201.15 | 7.5987 | <0.0001 |
| Aminocyclopyrachlor | 93,162.80 | 970.446 | 1.1362 | 0.2912 |
| Aminopyralid | 66,377.215 | 677.319 | 1.3343 | 0.1155 |
| Dicamba | 56,952.692 | 720.920 | 1.5689 | 0.0445 |
| Glyphosate | 39,430.523 | 788.610 | 2.4212 | 0.0036 |
| Picloram | 93,820.05 | 1054.16 | 2.6679 | <0.0001 |
| Quinclorac | 43,175.610 | 881.135 | 2.8772 | 0.0011 |
| Herbicide | Accession | Mean Injury (%) (28 DAT) |
|---|---|---|
| 2,4-D | TOM45 | 7.5J * |
| 2,4-D | TOM1 | 7.5J |
| 2,4-D | TOM56 | 5J |
| 2,4-D | TOM11 | 16.6HIJ |
| 2,4-D | TOM13 | 14.5HIJ |
| 2,4-D | TOM14 | 13IJ |
| 2,4-D | TOM22 | 11.6IJ |
| 2,4-D | TOM83 | 5J |
| 2,4-D | TOM17 | 4.5J |
| Dicamba | TOM1 | 8.5G |
| Dicamba | TOM3 | 9.5FG |
| Dicamba | TOM35 | 20EFG |
| Dicamba | TOM18 | 18.3EFG |
| Dicamba | TOM74 | 20EFG |
| Dicamba | TOM13 | 7.3FG |
| Dicamba | TOM14 | 15FG |
| Dicamba | TOM17 | 6.5G |
| Dicamba | TOM12 | 13.6FG |
| Dicamba | TOM262 | 12.5FG |
| Dicamba | TOM44 | 11.6FG |
| Glyphosate | TOM46 | 10.0IJ |
| Glyphosate | TOM60 | 3.3J |
| Glyphosate | TOM61 | 10.0IJ |
| Glyphosate | TOM64 | 11.6IJ |
| Glyphosate | TOM108 | 16.6HIJ |
| Glyphosate | TOM66 | 18.3GHIJ |
| Glyphosate | TOM18 | 16.6HIJ |
| Glyphosate | TOM102 | 20.0GHIJ |
| Glyphosate | TOM47 | 15.0HIJ |
| Quinclorac | TOM66 | 6.6H |
| Quinclorac | TOM129 | 3.3H |
| Quinclorac | TOM77 | 10.0EGH |
| Quinclorac | TOM410 | 15.0GH |
| Quinclorac | TOM63 | 8.3EGH |
| Aminocyclopyrachlor | TOM27 | 13.3FG |
| Aminocyclopyrachlor | TOM74 | 8.3FG |
| Aminocyclopyrachlor | TOM54 | 20.0EFG |
| Aminocyclopyrachlor | TOM78 | 10.0FG |
| Aminocyclopyrachlor | TOM29 | 6.6FG |
| Aminocyclopyrachlor | TOM44 | 5.0FG |
| Aminocyclopyrachlor | TOM129 | 5.0G |
| Aminocyclopyrachlor | TOM103 | 18.3EFG |
| Picloram | TOM17 | 20.0HIJ |
| Picloram | TOM47 | 13.3J |
| Aminopyralid | TOM76 | 10.0F |
| Aminopyralid | TOM84 | 18.3EF |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sharma, G.; Shrestha, S.; Tseng, T.-M.; Shrestha, S. Screening and Selection for Herbicide Tolerance among Diverse Tomato Germplasms. Horticulturae 2023, 9, 1354. https://doi.org/10.3390/horticulturae9121354
Sharma G, Shrestha S, Tseng T-M, Shrestha S. Screening and Selection for Herbicide Tolerance among Diverse Tomato Germplasms. Horticulturae. 2023; 9(12):1354. https://doi.org/10.3390/horticulturae9121354
Chicago/Turabian StyleSharma, Gourav, Swati Shrestha, Te-Ming Tseng, and Sanju Shrestha. 2023. "Screening and Selection for Herbicide Tolerance among Diverse Tomato Germplasms" Horticulturae 9, no. 12: 1354. https://doi.org/10.3390/horticulturae9121354
APA StyleSharma, G., Shrestha, S., Tseng, T.-M., & Shrestha, S. (2023). Screening and Selection for Herbicide Tolerance among Diverse Tomato Germplasms. Horticulturae, 9(12), 1354. https://doi.org/10.3390/horticulturae9121354