
Physiological and Transcriptional Analysis Provides Insights into Tea Saponin Biosynthesis and Regulation in Response to SA in Camellia vietnamensis Huang

 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials and SA Treatment
2.2. Determination of Secondary Metabolites
2.3. Determination of Physiological Enzyme Activity Index
2.4. Transcriptomic Analysis
2.5. Verification of RT-qPCR Analysis
2.6. Statistical Analysis
3. Results
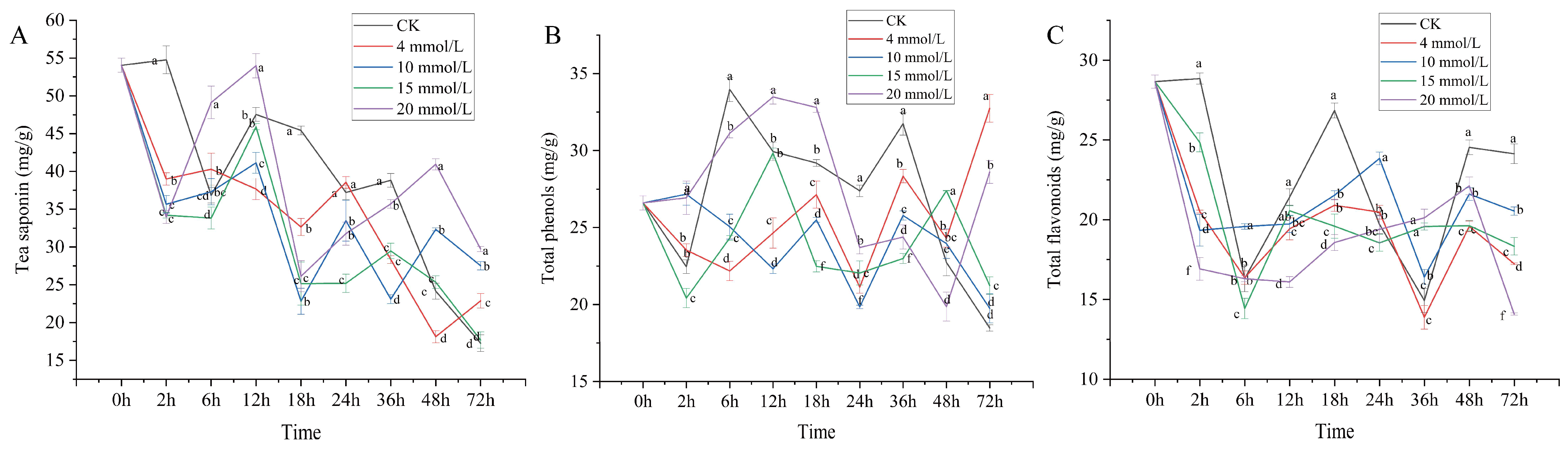
3.1. Effect of SA on Secondary Metabolic Components of C. vietnamensis
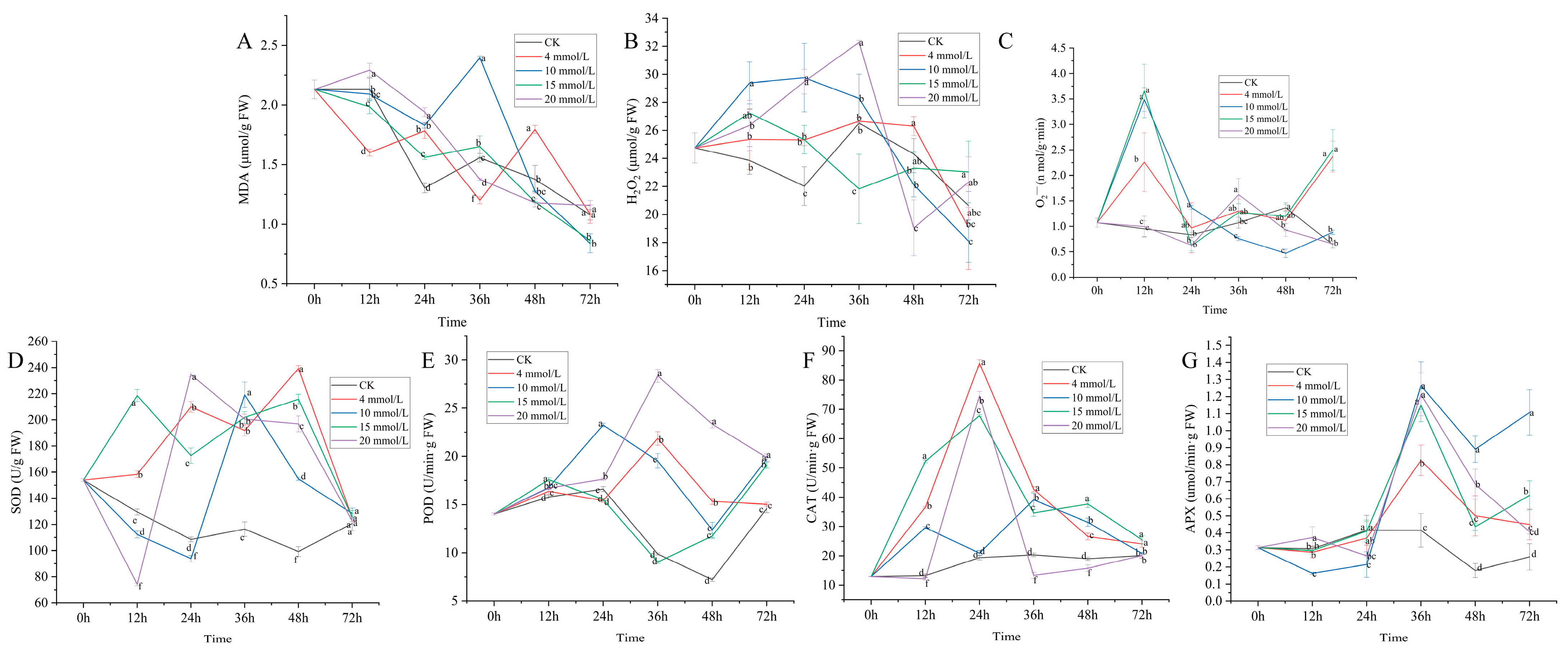
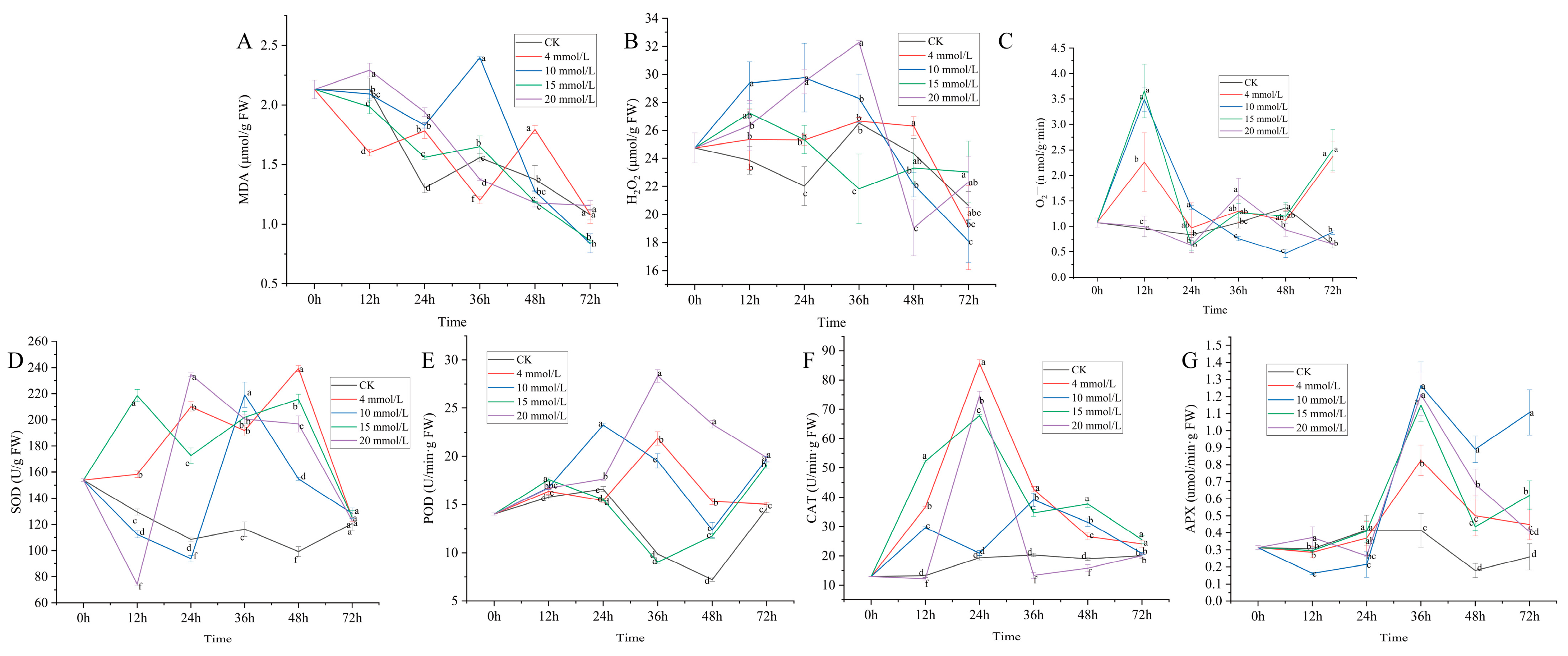
3.2. Effect of SA on Physiological Activity of C. vietnamensis
3.3. Correlation Analysis of Secondary Metabolic Components and Physiological Activity Indexes
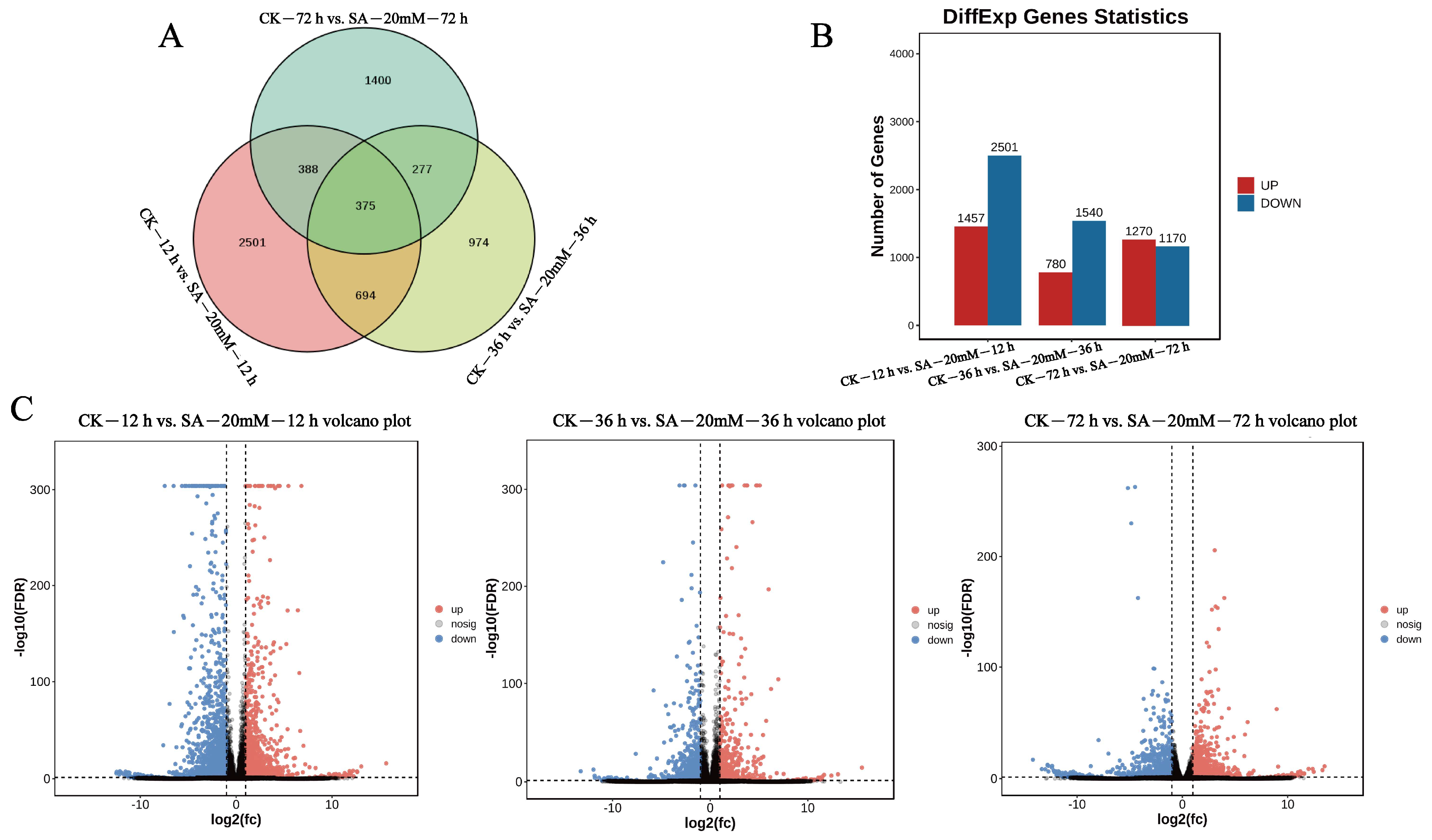
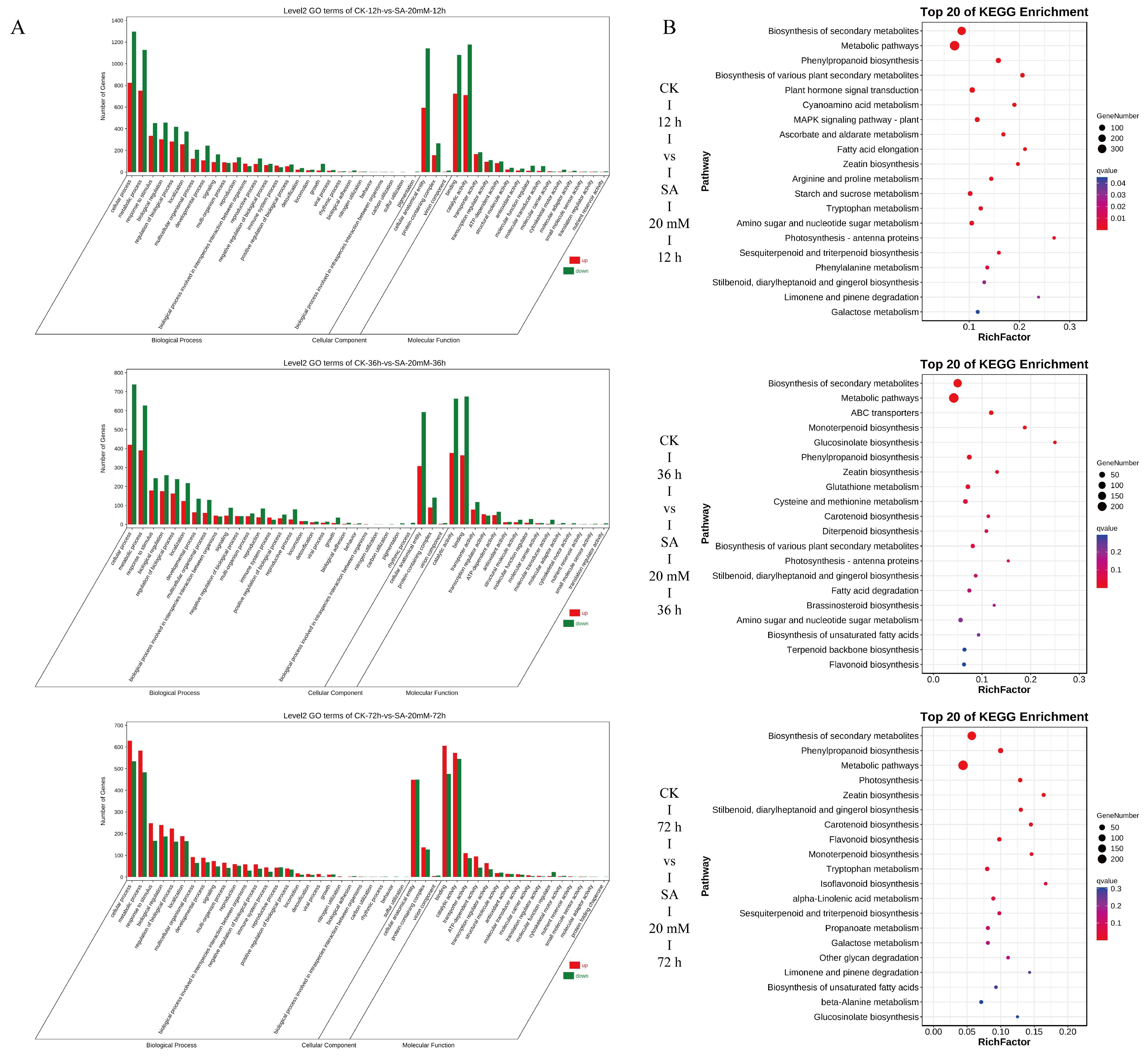
3.4. Differential Gene Identification, GO, and KEGG Enrichment Analysis
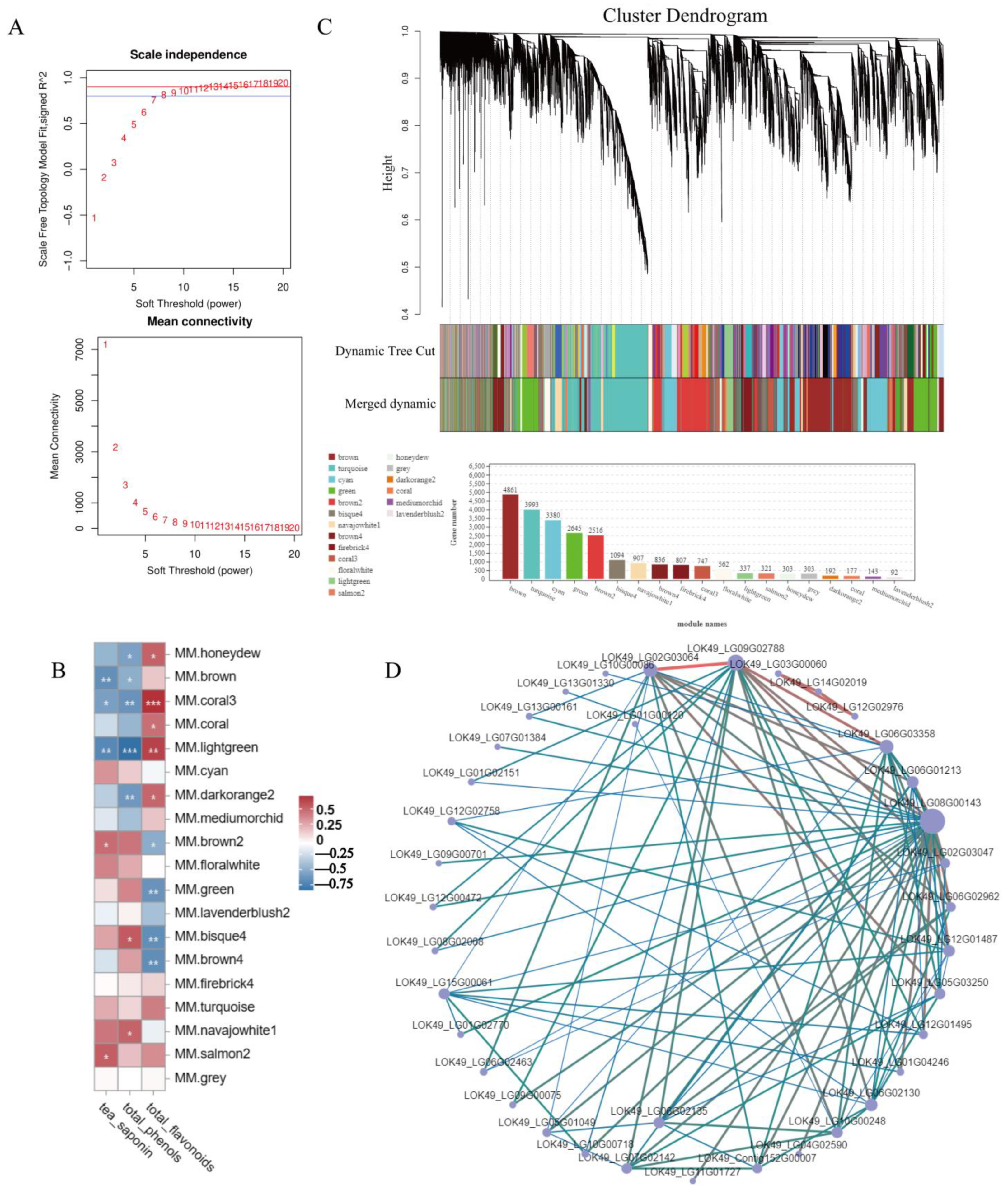
3.5. WGCNA of Transcriptome and Secondary Metabolites
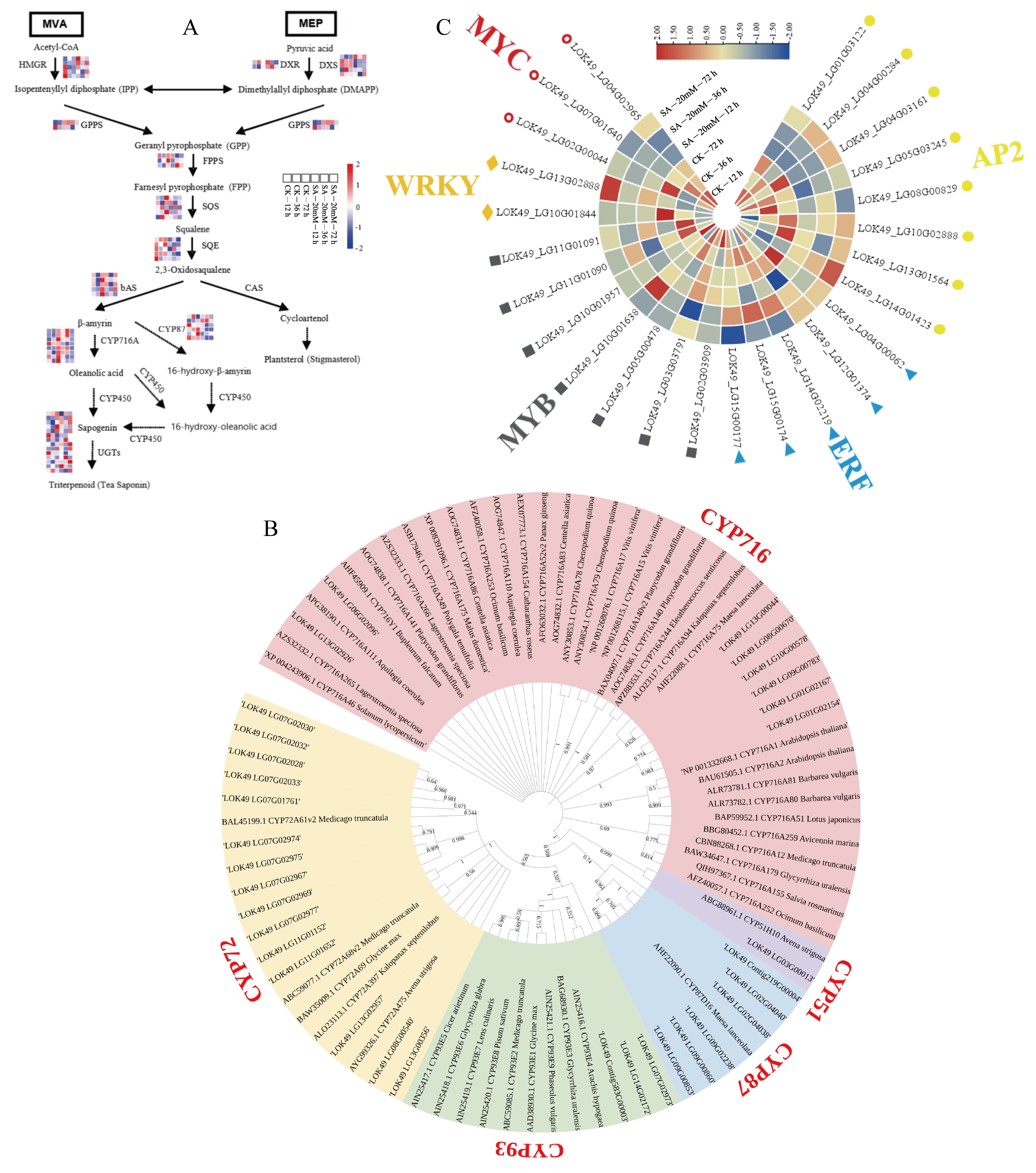
3.6. Analysis of the Synthesis Pathway of Tea Saponin from C. vietnamensis and Identification of Related CYP450
3.7. Screening of Transcription Factor (TF) Associated with Tea Saponin Synthesis in C. vietnamensis
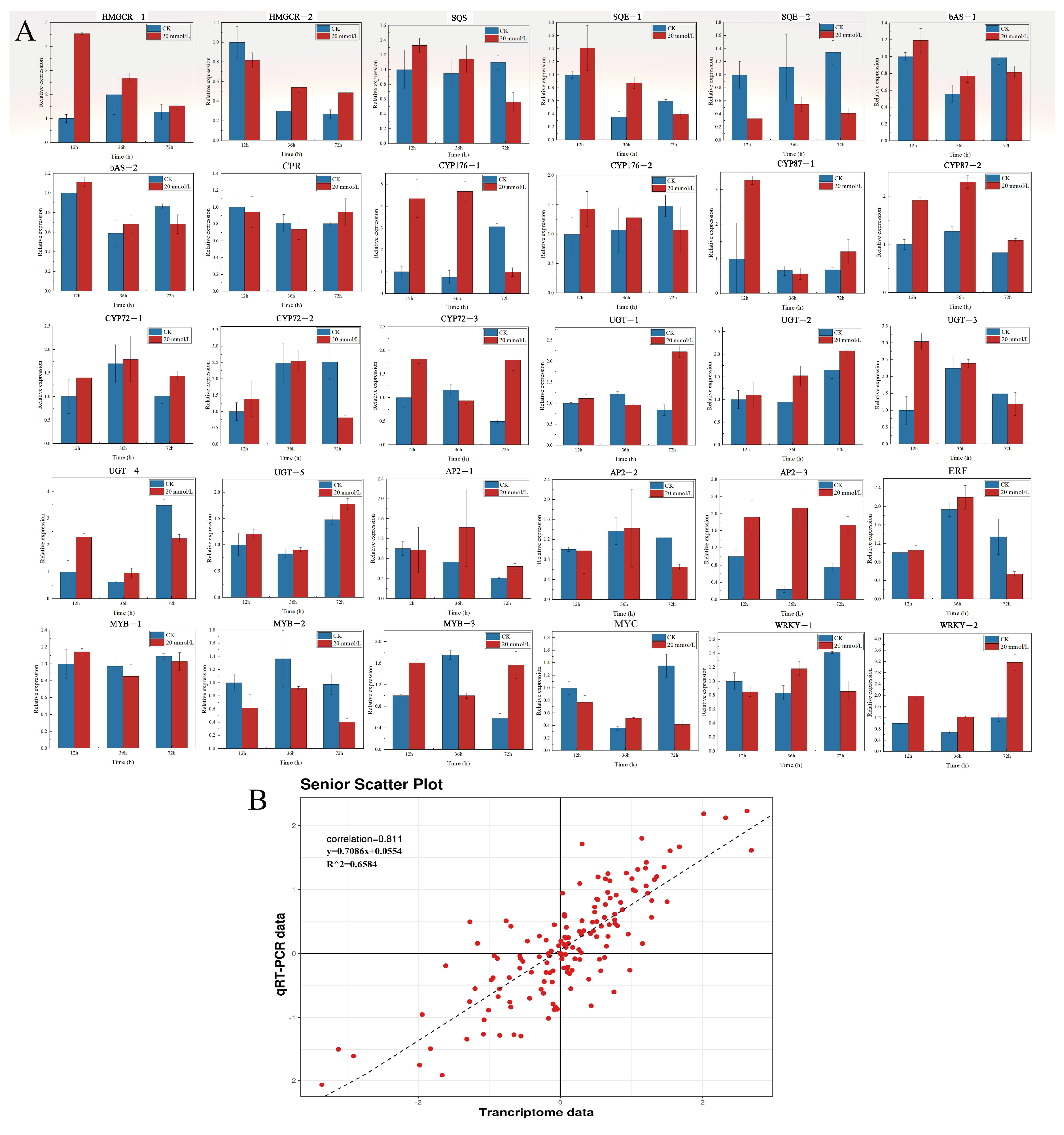
3.8. Correlation Analysis of RT-qPCR and Transcriptome
4. Discussion
4.1. Analysis of Secondary Metabolic Components and Physiological Activity Response to SA in C. vietnamensis
4.2. Candidate Structural Genes Involved in Theasaponins Pathway in C. vietnamensis
4.3. TFs Involved in Theasaponins Biosynthesis of C. vietnamensis
5. Conclusions
6. Patents
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dai, J.; Zheng, W.; Yu, J.; Yan, H.; Wang, Y.; Wu, Y.; Hu, X.; Lai, H. cDNA cloning, prokaryotic expression, and functional analysis of squalene synthase (SQS) in Camellia vietnamensis Huang. Protein Expr. Purif. 2022, 194, 106078. [Google Scholar] [CrossRef]
- Zhang, W.; Zhao, Y.; Yang, G.; Peng, J.; Chen, S.; Xu, Z. Determination of the evolutionary pressure on Camellia Oleifera on Hainan Island using the complete chloroplast genome sequence. PeerJ 2019, 7, e7210. [Google Scholar] [CrossRef] [PubMed]
- Ye, Z.; Wu, Y.; Ul Haq Muhammad, Z.; Yan, W.; Yu, J.; Zhang, J.; Yao, G.; Hu, X. Complementary transcriptome and proteome profiling in the mature seeds of Camellia oleifera from Hainan Island. PLoS ONE 2020, 15, e0226888. [Google Scholar] [CrossRef]
- Chen, J.; Guo, Y.; Hu, X.; Zhou, K. Comparison of the chloroplast genome sequences of 13 oil-tea Camellia samples and identification of an undetermined oil-tea Camellia species from Hainan province. Front. Plant Sci. 2022, 12, 798581. [Google Scholar] [CrossRef]
- He, L.; Guoying, Z.; Huaiyun, Z.; Yuanhao, H. Chemical constituents and biological activities of saponin from the seed of Camellia oleifera. Sci. Res. Essays 2010, 5, 4088–4092. [Google Scholar]
- Guo, N.; Tong, T.; Ren, N.; Tu, Y.; Li, B. Saponins from seeds of genus Camellia: Phytochemistry and bioactivity. Phytochemistry 2018, 149, 42–55. [Google Scholar] [CrossRef] [PubMed]
- Żwawiak, J.; Pawełczyk, A.; Olender, D.; Zaprutko, L. Structure and activity of pentacyclic triterpenes codrugs. A review. Mini-Rev. Med. Chem. 2021, 21, 1509–1526. [Google Scholar] [CrossRef]
- Li, Y.; Wang, J.; Li, L.; Song, W.; Li, M.; Hua, X.; Wang, Y.; Yuan, J.; Xue, Z. Natural products of pentacyclic triterpenoids: From discovery to heterologous biosynthesis. Nat. Prod. Rep. 2023, 40, 1303–1353. [Google Scholar] [CrossRef]
- Su, L.; Li, S.; Qiu, H.; Wang, H.; Wang, C.; He, C.; Xu, M.; Zhang, Z. Full-length transcriptome analyses of genes involved in triterpenoid saponin biosynthesis of Psammosilene tunicoides hairy root cultures with exogenous salicylic acid. Front. Genet. 2021, 12, 657060. [Google Scholar] [CrossRef]
- Yang, L.; Qiao, L.; Su, X.; Ji, B.; Dong, C. Drought stress stimulates the terpenoid backbone and triterpenoid biosynthesis pathway to promote the synthesis of saikosaponin in Bupleurum chinense DC. roots. Molecules 2022, 27, 5470. [Google Scholar] [CrossRef]
- Sallaud, C.; Rontein, D.; Onillon, S.; Jabès, F.; Duffé, P.; Giacalone, C.; Thoraval, S.; Escoffier, C.; Herbette, G.; Leonhardt, N.; et al. A novel pathway for sesquiterpene biosynthesis from Z,Z-farnesyl pyrophosphate in the wild tomato Solanum habrochaites. Plant Cell 2009, 21, 301–317. [Google Scholar] [CrossRef] [PubMed]
- Lin, P.; Wang, K.; Wang, Y.; Hu, Z.; Yan, C.; Huang, H.; Ma, X.; Cao, Y.; Long, W.; Liu, W.; et al. The genome of oil-Camellia and population genomics analysis provide insights into seed oil domestication. Genome Biol. 2022, 23, 14. [Google Scholar] [CrossRef] [PubMed]
- Yang, R.; Yan, Y.; Zeng, Y.; Jin, S.; Jiang, Q.; Yu, L.; Lai, R. Correlation between squalene synthase promoter and WRKY transcription factor in Camellia oleifera. J. Hortic. Sci. Biotechnol. 2021, 96, 34–43. [Google Scholar] [CrossRef]
- Nielsen, N.J.; Nielsen, J.; Staerk, D. New resistance-correlated saponins from the insect-resistant Crucifer Barbarea vulgaris. J. Agric. Food Chem. 2010, 58, 5509–5514. [Google Scholar] [CrossRef] [PubMed]
- Augustin, J.M.; Kuzina, V.; Andersen, S.B.; Bak, S. Molecular activities, biosynthesis and evolution of triterpenoid saponins. Phytochemistry 2011, 72, 435–457. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.; Wang, J.; Mao, S.; Xu, H.; Wu, Q.; Liang, M.; Yuan, Y.; Liu, M.; Huang, K. Comparative transcriptome analyses of genes involved in sulforaphane metabolism at different treatment in Chinese kale using full-length transcriptome sequencing. BMC Genom. 2019, 20, 377. [Google Scholar] [CrossRef]
- Lu, H.; Greenberg, J.T.; Holuigue, L. Editorial: Salicylic acid signaling networks. Front. Plant Sci. 2016, 7, 238. [Google Scholar] [CrossRef]
- Sawai, S.; Saito, K. Triterpenoid biosynthesis and engineering in plants. Front. Plant Sci. 2011, 2, 25. [Google Scholar] [CrossRef]
- Yendo, A.C.A.; De Costa, F.; Gosmann, G.; Fett-Neto, A.G. Production of plant bioactive triterpenoid saponins: Elicitation strategies and target genes to improve yields. Mol. Biotechnol. 2010, 46, 94–104. [Google Scholar] [CrossRef]
- Ramirez-Estrada, K.; Vidal-Limon, H.; Hidalgo, D.; Moyano, E.; Golenioswki, M.; Cusidó, R.; Palazon, J. Elicitation, an effective strategy for the biotechnological production of bioactive high-added value compounds in plant cell factories. Molecules 2016, 21, 182. [Google Scholar] [CrossRef]
- Gutierrez-Valdes, N.; Häkkinen, S.T.; Lemasson, C.; Guillet, M.; Oksman-Caldentey, K.-M.; Ritala, A.; Cardon, F. Hairy root cultures—A versatile tool with multiple applications. Front. Plant Sci. 2020, 11, 33. [Google Scholar] [CrossRef] [PubMed]
- Tewari, R.K.; Paek, K.-Y. Salicylic acid-induced nitric oxide and ROS generation stimulate ginsenoside accumulation in Panax ginseng roots. J. Plant Growth Regul. 2011, 30, 396–404. [Google Scholar] [CrossRef]
- Teng, H.; Xie, X.; Wang, L.; Hu, Z. Effects of exogenous substances on saponin content in root of Polygala tenuifolia Willd. South. J. Agric. 2014, 45, 2143–2147. [Google Scholar]
- Ye, J.; Mao, D.; Cheng, S.; Zhang, X.; Tan, J.; Zheng, J.; Xu, F. Comparative transcriptome analysis reveals the potential stimulatory mechanism of terpene trilactone biosynthesis by exogenous salicylic acid in Ginkgo biloba. Ind. Crops Prod. 2020, 145, 112104. [Google Scholar] [CrossRef]
- Kim, J.D.; Khan, M.I.; Shin, J.H.; Lee, M.G.; Seo, H.J.; Shin, T.S.; Kim, M.Y. HPLC fractionation and pharmacological assessment of green tea seed saponins for antimicrobial, anti-angiogenic and hemolytic activities. Biotechnol. Bioprocess Eng. 2015, 20, 1035–1043. [Google Scholar] [CrossRef]
- Maulana, T.I.; Falah, S.; Andrianto, D. Total phenolic content, total flavonoid content, and antioxidant activity of water and ethanol extract from Surian (Toona sinensis) leaves. IOP Conf. Ser. Earth Environ. Sci. 2019, 299, 012021. [Google Scholar] [CrossRef]
- Zhao, Y.; Su, R.; Zhang, W.; Yao, G.-L.; Chen, J. Antibacterial activity of tea saponin from Camellia oleifera Shell by novel extraction method. Ind. Crops Prod. 2020, 153, 112604. [Google Scholar] [CrossRef]
- Yu, J.; Yan, H.; Wu, Y.; Wang, Y.; Xia, P. Quality evaluation of the oil of Camellia spp. Foods 2022, 11, 2221. [Google Scholar] [CrossRef]
- El Amine, B.; Mosseddaq, F.; Naciri, R.; Oukarroum, A. Interactive effect of Fe and Mn deficiencies on physiological, biochemical, nutritional and growth status of Soybean. Plant Physiol. Biochem. 2023, 199, 107718. [Google Scholar] [CrossRef]
- Saidi, I.; Ayouni, M.; Dhieb, A.; Chtourou, Y.; Chaïbi, W.; Djebali, W. Oxidative damages induced by short-term exposure to cadmium in bean plants: Protective role of salicylic acid. S. Afr. J. Bot. 2013, 85, 32–38. [Google Scholar] [CrossRef]
- Zhang, B.; Horvath, S. A general framework for weighted gene coexpression network analysis. Stat. Appl. Genet. Mol. Biol. 2005, 4, 17. [Google Scholar] [CrossRef] [PubMed]
- Naz, S.; Bilal, A.; Saddiq, B.; Ejaz, S.; Ali, S.; Ain Haider, S.T.; Sardar, H.; Nasir, B.; Ahmad, I.; Tiwari, R.K.; et al. Foliar application of salicylic acid improved growth, yield, quality and photosynthesis of Pea (Pisum sativum L.) by improving antioxidant defense mechanism under saline conditions. Sustainability 2022, 14, 14180. [Google Scholar] [CrossRef]
- Ni, J.; Dong, L.; Jiang, Z.; Yang, X.; Sun, Z.; Li, J.; Wu, Y.; Xu, M. Salicylic acid-induced flavonoid accumulation in ginkgo biloba leaves is dependent on red and far-red light. Ind. Crops Prod. 2018, 118, 102–110. [Google Scholar] [CrossRef]
- Biswas, T.; Mathur, A.; Gupta, V.; Singh, M.; Mathur, A.K. Salicylic acid and ultrasonic stress modulated gene expression and ginsenoside production in differentially affected Panax quinquefolius (L.) and Panax sikkimensis (Ban.) cell suspensions. Plant Cell Tissue Organ Cult. PCTOC 2019, 136, 575–588. [Google Scholar] [CrossRef]
- Tajik, S.; Zarinkamar, F.; Soltani, B.M.; Nazari, M. Induction of phenolic and flavonoid compounds in leaves of Saffron (Crocus sativus L.) by salicylic acid. Sci. Hortic. 2019, 257, 108751. [Google Scholar] [CrossRef]
- Ghasemzadeh, A.; Jaafar, H. Interactive effect of salicylic acid on some physiological features and antioxidant enzymes activity in ginger (Zingiber officinale Roscoe). Molecules 2013, 18, 5965–5979. [Google Scholar] [CrossRef] [PubMed]
- Syed, B.A.; Patel, M.; Patel, A.; Gami, B.; Patel, B. Regulation of antioxidant enzymes and osmo-protectant molecules by salt and drought responsive genes in Bambusa balcooa. J. Plant Res. 2021, 134, 165–175. [Google Scholar] [CrossRef] [PubMed]
- Tyagi, S.; Shumayla; Verma, P.C.; Singh, K.; Upadhyay, S.K. Molecular characterization of ascorbate peroxidase (APX) and APX-related (APX-R) genes in Triticum aestivum L. Genomics 2020, 112, 4208–4223. [Google Scholar] [CrossRef]
- Wang, L.-J.; Li, S.-H. Thermotolerance and related antioxidant enzyme activities induced by heat acclimation and salicylic acid in grape (Vitis vinifera L.) leaves. Plant Growth Regul. 2006, 48, 137–144. [Google Scholar] [CrossRef]
- Xu, Y.; Zhao, G.; Ji, X.; Liu, J.; Zhao, T.; Gao, Y.; Gao, S.; Hao, Y.; Gao, Y.; Wang, L.; et al. Metabolome and transcriptome analysis reveals the transcriptional regulatory mechanism of triterpenoid saponin biosynthesis in Soapberry (Sapindus mukorossi Gaertn.). J. Agric. Food Chem. 2022, 70, 7095–7109. [Google Scholar] [CrossRef]
- Yao, L.; Wang, J.; Sun, J.; He, J.; Paek, K.-Y.; Park, S.-Y.; Huang, L.; Gao, W. A WRKY transcription factor, PgWRKY4X, positively regulates ginsenoside biosynthesis by activating squalene epoxidase transcription in Panax ginseng. Ind. Crops Prod. 2020, 154, 112671. [Google Scholar] [CrossRef]
- Yin, J.; Sun, L.; Li, Y.; Xiao, J.; Wang, S.; Yang, J.; Qu, Z.; Zhan, Y. Functional identification of BpMYB21 and BpMYB61 transcription factors responding to MeJA and SA in birch triterpenoid synthesis. BMC Plant Biol. 2020, 20, 374. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Tea Saponin | Total Phenols | Total Flavonoids | MDA | SOD | POD | CAT | APX | H2O2 | O2− | |
|---|---|---|---|---|---|---|---|---|---|---|
| Tea saponin | 1 | 0.413 * | 0.385 * | 0.666 ** | −0.241 | −0.238 | −0.242 | −0.318 | 0.203 | 0.11 |
| Total phenols | 0.413 * | 1 | −0.253 | 0.262 | −0.233 | −0.215 | −0.213 | −0.148 | 0.08 | 0.118 |
| Total flavonoids | 0.385 * | −0.253 | 1 | 0.663 ** | −0.234 | −0.573 ** | −0.352 | −0.34 | −0.093 | −0.065 |
| MDA | 0.666 ** | 0.262 | 0.663 ** | 1 | −0.131 | −0.567 ** | −0.206 | −0.316 | 0.311 | −0.017 |
| SOD | −0.241 | −0.233 | −0.234 | −0.131 | 1 | 0.278 | 0.615 ** | 0.404 * | 0.282 | −0.068 |
| POD | −0.238 | −0.215 | −0.573 ** | −0.567 ** | 0.278 | 1 | 0.171 | 0.388 * | 0.252 | 0.119 |
| CAT | −0.242 | −0.213 | −0.352 | −0.206 | 0.615 ** | 0.171 | 1 | −0.003 | 0.245 | 0.026 |
| APX | −0.318 | −0.148 | −0.34 | −0.316 | 0.404 * | 0.388 * | −0.003 | 1 | −0.075 | −0.201 |
| H2O2 | 0.203 | 0.08 | −0.093 | 0.311 | 0.282 | 0.252 | 0.245 | −0.075 | 1 | 0.238 |
| O2− | 0.11 | 0.118 | −0.065 | −0.017 | −0.068 | 0.119 | 0.026 | −0.201 | 0.238 | 1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Y.; Yan, H.; Zeeshan Ul Haq, M.; Liu, Y.; Wu, Y.; Yu, J.; Xia, P. Physiological and Transcriptional Analysis Provides Insights into Tea Saponin Biosynthesis and Regulation in Response to SA in Camellia vietnamensis Huang. Horticulturae 2024, 10, 8. https://doi.org/10.3390/horticulturae10010008
Li Y, Yan H, Zeeshan Ul Haq M, Liu Y, Wu Y, Yu J, Xia P. Physiological and Transcriptional Analysis Provides Insights into Tea Saponin Biosynthesis and Regulation in Response to SA in Camellia vietnamensis Huang. Horticulturae. 2024; 10(1):8. https://doi.org/10.3390/horticulturae10010008
Chicago/Turabian StyleLi, Yang, Heqin Yan, Muhammad Zeeshan Ul Haq, Ya Liu, Yougen Wu, Jing Yu, and Pengguo Xia. 2024. "Physiological and Transcriptional Analysis Provides Insights into Tea Saponin Biosynthesis and Regulation in Response to SA in Camellia vietnamensis Huang" Horticulturae 10, no. 1: 8. https://doi.org/10.3390/horticulturae10010008
APA StyleLi, Y., Yan, H., Zeeshan Ul Haq, M., Liu, Y., Wu, Y., Yu, J., & Xia, P. (2024). Physiological and Transcriptional Analysis Provides Insights into Tea Saponin Biosynthesis and Regulation in Response to SA in Camellia vietnamensis Huang. Horticulturae, 10(1), 8. https://doi.org/10.3390/horticulturae10010008