
Pilot-Scale Production of Traditional Galotyri PDO Cheese from Boiled Ewes’ Milk Fermented with the Aid of Greek Indigenous Lactococcus lactis subsp. cremoris Starter and Lactiplantibacillus plantarum Adjunct Strains
 ,
,  ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Bacterial Strains, Starter/Adjunct LAB Strain Combinations, and Culture Conditions
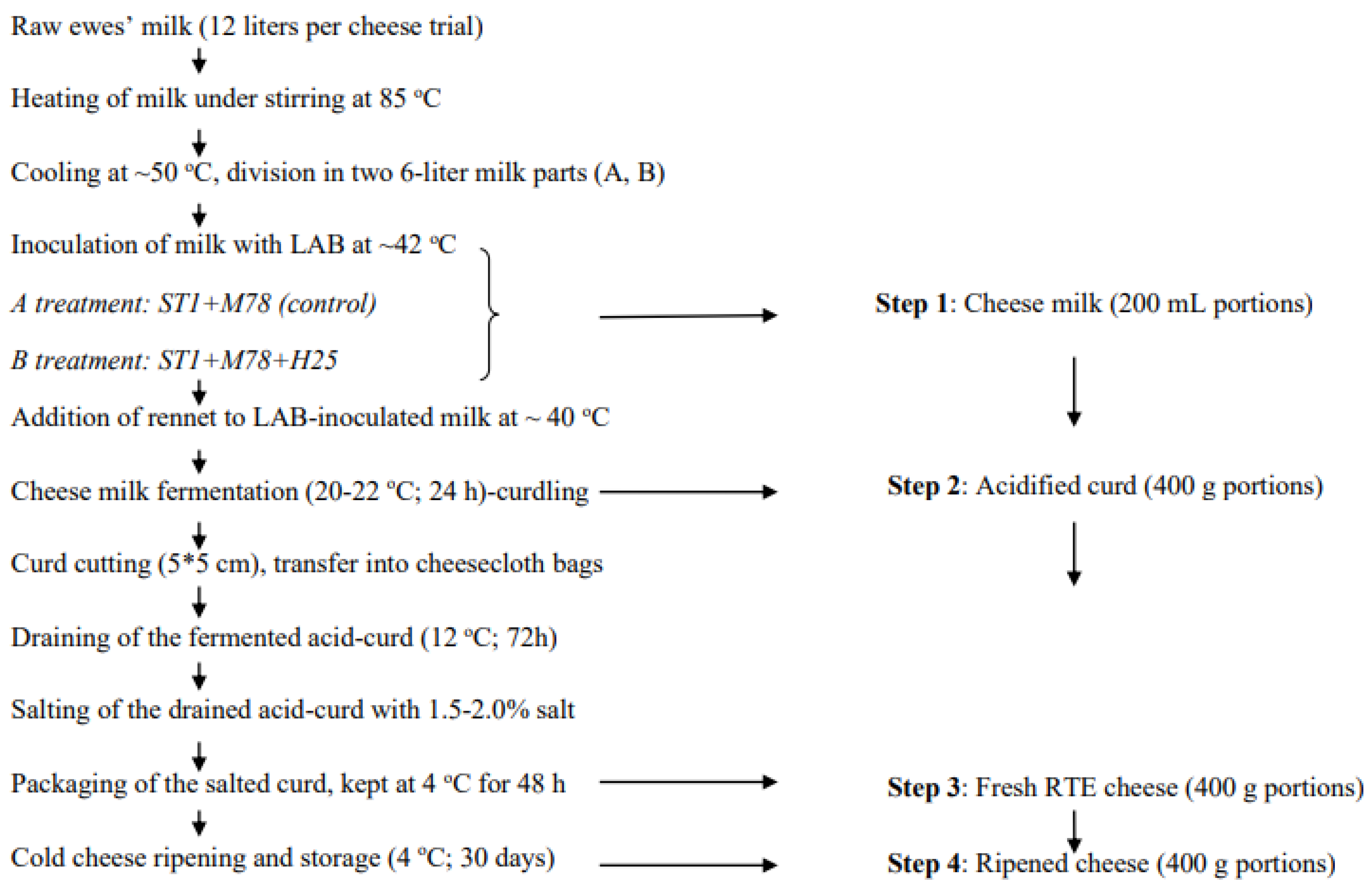
2.2. Artisanal Galotyri Cheese Preparation and Sampling
2.3. Microbiological Analyses of the Artisanal Galotyri Cheese Samples
2.4. Measurement of pH and Gross Composition of the Artisanal Galotyri Cheese Samples
2.5. Assessment of Proteolysis in the Artisanal Galotyri Cheese Samples
2.6. Determination of Sugar and Organic Acid Concentrations
2.7. Sensory Evaluation
2.8. Statistical Analysis
3. Results
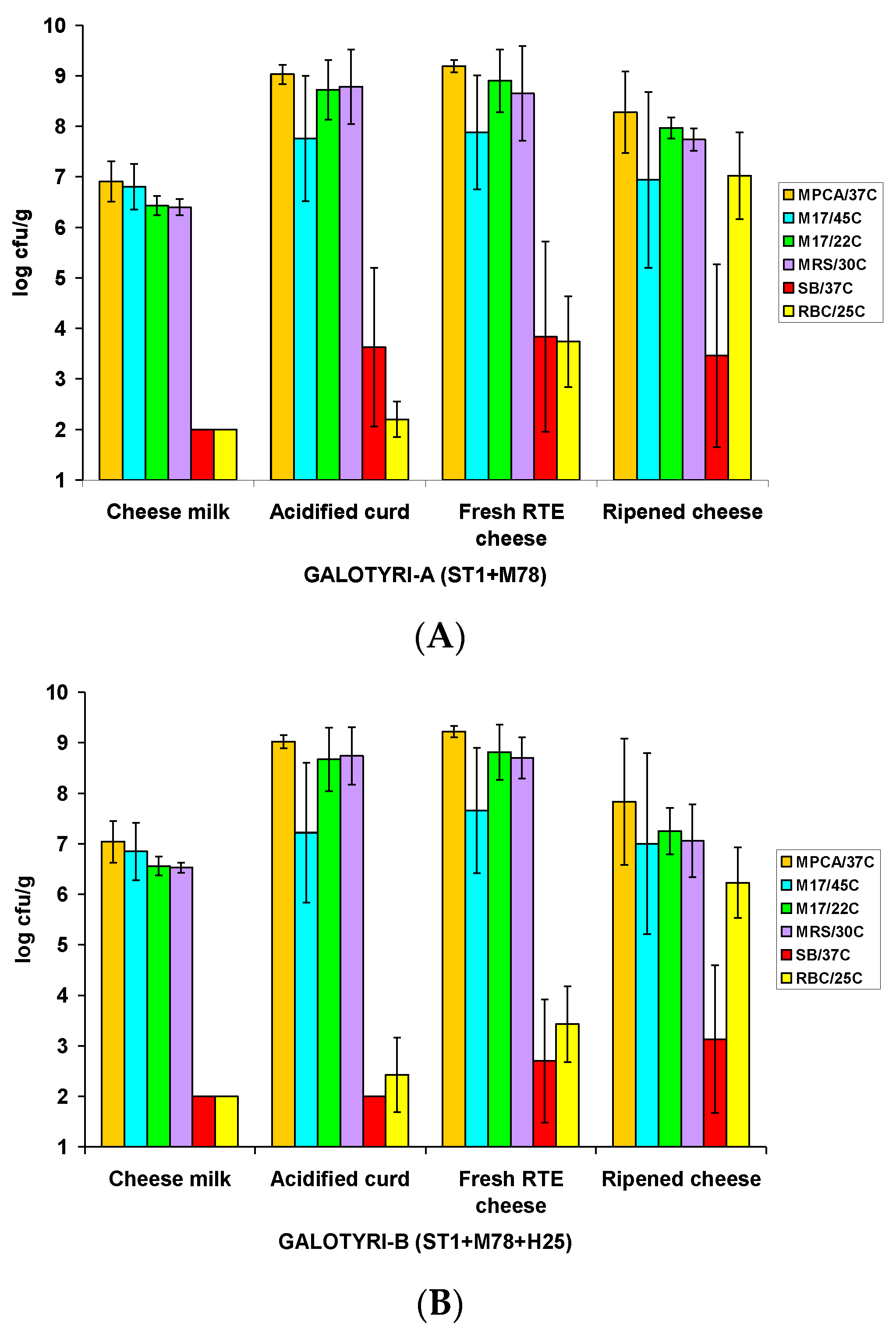
3.1. Growth of Starter LAB and Native Microbiota in the Artisanal Galotyri Cheeses
3.2. pH Values and Gross Composition of the Artisanal Galotyri Cheeses
3.3. Sugar and Organic Acid Concentrations in the Artisanal Galotyri Cheeses
3.4. Proteolysis—Nitrogen Fractions of the Artisanal Galotyri Cheeses
3.5. Sensory Evaluation of the Artisanal Galotyri Cheeses
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Samelis, J.; Lianou, A.; Kakouri, A.; Delbès, C.; Rogelj, I.; Matijašic, B.B.; Montel, M.C. Changes in the microbial composition of raw milk induced by thermization treatments applied prior to traditional Greek hard cheese processing. J. Food Prot. 2009, 72, 783–790. [Google Scholar] [CrossRef] [PubMed]
- Litopoulou-Tzanetaki, E.; Tzanetakis, N. Microbiological characteristics of Greek traditional cheeses. Small Rum. Res. 2011, 101, 17–32. [Google Scholar] [CrossRef]
- Panagou, E.Z.; Nychas, G.J.E.; Sofos, J.N. Types of traditional Greek foods and their safety. Food Cont. 2013, 29, 32–41. [Google Scholar] [CrossRef]
- Rhoades, J.; Anastasiou, I.; Michailidou, S.; Koinidis, A.; Doulgerakis, C.; Alexa, E.A.; Alvarez-Ordonez, A.; Argiriou, A.; Likotrafiti, E. Microbiological analysis of Greek Protected Designation of Origin cheeses and characterization of the isolated lactic acid bacteria. Int. Dairy J. 2021, 123, 105183. [Google Scholar] [CrossRef]
- Beresford, T.P.; Fitzsimons, N.A.; Brennan, N.L.; Cogan, T.M. Recent advances in cheese microbiology. Int. Dairy. J. 2001, 11, 259–274. [Google Scholar] [CrossRef]
- Leroy, F.; De Vuyst, L. Lactic acid bacteria as functional starter cultures for food fermentation industry. Trends Food Sci. Technol. 2004, 15, 67–78. [Google Scholar] [CrossRef]
- Bintsis, T.; Athanasoulsas, A. Dairy Starter Cultures. In Dairy Microbiology, a Practical Approach; Papademas, P., Ed.; CRC Press: Boca Raton, FL, USA, 2015; pp. 114–154. [Google Scholar]
- Bintsis, T. Lactic acid bacteria: Their applications in foods. J. Bacteriol. Mycol. 2018, 5, 1065. [Google Scholar]
- Wouters, J.T.M.; Ayad, E.H.E.; Hugenholtz, J.; Smit, G. Microbes from raw milk for fermented dairy products. Int. Dairy J. 2002, 12, 91–109. [Google Scholar] [CrossRef]
- Johansen, E. Use of natural selection and evolution to develop new starter cultures for fermented foods. Annu. Rev. Food. Sci. Technol. 2018, 9, 411–428. [Google Scholar] [CrossRef]
- Bintsis, T. Lactic acid bacteria as starter cultures: An update of their metabolism and genetics. AIMS Microbiol. 2018, 4, 665–684. [Google Scholar] [CrossRef]
- Anifantakis, E.M. Greek Cheeses: A Tradition of Centuries; National Dairy Committee of Greece: Athens, Greece, 1991. [Google Scholar]
- Samelis, J.; Kakouri, A.; Pappa, E.C.; Matijašic, B.B.; Georgalaki, M.D.; Tsakalidou, E.; Rogelj, I. Microbial stability and safety of traditional Greek Graviera cheese: Characterization of the lactic acid bacterial flora and culture-independent detection of bacteriocin genes in the ripened cheeses and their microbial consortia. J. Food Prot. 2010, 73, 1294–1303. [Google Scholar] [CrossRef] [PubMed]
- Papadopoulou, O.S.; Argyri, A.A.; Varzakis, E.E.; Tassou, C.C.; Chorianopoulos, N.G. Greek functional Feta cheese: Enhancing quality and safety using a Lactobacillus plantarum strain with probiotic potential. Food Microbiol. 2018, 74, 21–33. [Google Scholar] [CrossRef] [PubMed]
- Smid, E.J.; Erkus, O.; Spus, M.; Wolkers-Rooijackers, J.C.M.; Alexeeva, S.; Kleerebezem, M. Functional implications of the microbial community structure of undefined mesophilic starter cultures. Microbial Cell Fact. 2014, 13 (Suppl. S1), S2. [Google Scholar] [CrossRef] [PubMed]
- Samelis, J.; Kakouri, A. Major technological differences between an industrial-type and five artisan-type Greek PDO Galotyri market cheeses as revealed by great variations in their lactic acid microbiota. AIMS Agric. Food 2019, 4, 685–710. [Google Scholar] [CrossRef]
- Cogan, T.M.; Beresford, T.P.; Steele, J.; Broadbent, J.; Shah, N.P.; Ustunol, Z. Invited review: Advances in starter cultures and cultured foods. J. Dairy Sci. 2007, 90, 4005–4021. [Google Scholar] [CrossRef]
- Settanni, L.; Moschetti, G. Non-starter lactic acid bacteria used to improve cheese quality and provide health benefits. Food Microbiol. 2010, 27, 691–697. [Google Scholar] [CrossRef]
- Gobbetti, M.; De Angelis, M.; Di Cagno, R.; Mancini, L.; Fox, P.F. Pros and cons for using non-starter lactic acid bacteria (NSLAB) as secondary/adjunct starters for cheese ripening. Trends Food Sci. Technol. 2015, 45, 167–178. [Google Scholar] [CrossRef]
- Cheeses of protected denomination of origin. In Hellenic Code of Food and Beverages, 3rd ed.; Ministry of Finance, National Publishing Office: Athens, Greece, 2014; pp. 14–59.
- Danezis, G.P.; Tsiplakou, E.; Pappa, E.C.; Pappas, A.C.; Mavrommatis, A.; Sotirakoglou, K.; Georgiou, C.A.; Zervas, G. Fatty acid profile and physicochemical properties of Greek protected designation of origin cheeses, implications for authentication. Eur. Food Res. Technol. 2020, 246, 1741–1753. [Google Scholar] [CrossRef]
- Asteri, I.A.; Robertson, N.; Kagkli, D.M.; Andrews, P.; Nychas, G.; Coolbear, T.; Holland, R.; Crow, V.; Tsakalidou, E. Technological and flavor potential of cultures isolated from traditional Greek cheeses—A pool of novel species and starters. Int. Dairy J. 2009, 19, 595–604. [Google Scholar] [CrossRef]
- Samelis, J.; Argyri, A.; Doulgeraki, A.; Koukkou, A.I.; Skandamis, P.; Athanasoulas, A.; Pappas, D.; Chorianopoulos, N. Development of Novel Biofunctional Foods and Total Quality Enhancement of Traditional Dairy Products by Suitable Management of Their Microbial Ecology—BIO TRUST. In Proceedings of the 26th International ICFMH Conference FoodMicro, Freie Universität, Berlin, Germany, 3–6 September 2018; P1.62. p. 152. [Google Scholar]
- Tsanasidou, C.; Asimakoula, S.; Sameli, N.; Fanitsios, C.; Vandera, E.; Bosnea, L.; Koukou, A.I.; Samelis, J. Safety evaluation, biogenic amine formation, and enzymatic activity profiles of autochthonous enterocin-producing Greek cheese isolates of the Enterococcus faecium/durans group. Microorganisms 2021, 9, 777. [Google Scholar] [CrossRef]
- Kamarinou, C.S.; Papadopoulou, O.S.; Doulgeraki, A.I.; Tassou, C.C.; Galanis, A.; Chorianopoulos, N.G.; Argyri, A.A. Mapping the key technological and functional characteristics of indigenous lactic acid bacteria isolated from Greek traditional dairy products. Microorganisms 2022, 10, 246. [Google Scholar] [CrossRef]
- Samelis, J.; Kakouri, A. Cell growth density and nisin A activity of the indigenous Lactococcus lactis subsp. cremoris M78 costarter depend strongly on inoculation levels of a commercial Streptococcus thermophilus starter in milk: Practical aspects for traditional Greek cheese processors. J. Food Prot. 2020, 83, 542–551. [Google Scholar] [PubMed]
- Asimakoula, S.; Giaka, K.; Fanitsios, C.; Kakouri, A.; Vandera, E.; Samelis, J.; Koukkou, A.-I. Monitoring growth compatibility and bacteriocin gene transcription of adjunct and starter lactic acid bacterial strains in milk. J. Food Prot. 2021, 84, 509–520. [Google Scholar] [CrossRef]
- Sameli, N.; Skandamis, P.N.; Samelis, J. Application of Enterococcus faecium KE82, an enterocin A-B-P-producing strain, as an adjunct culture enhances inactivation of Listeria monocytogenes during traditional protected designation of origin Galotyri processing. J. Food Prot. 2021, 84, 87–98. [Google Scholar] [CrossRef]
- Samelis, J.; Kakouri, A. Microbiological characterization of Greek Glotyri cheese PDO products relative to whether they are marketed fresh or ripened. Fermentation 2022, 8, 492. [Google Scholar] [CrossRef]
- Samelis, J.; Doulgeraki, A.I.; Bikouli, V.; Pappas, D.; Kakouri, A. Microbiological and metagenomic characterization of a retail delicatessen Galotyri-like fresh acid-curd cheese product. Fermentation 2021, 7, 67. [Google Scholar] [CrossRef]
- Trmčić, A.; Samelis, J.; Monnet, C.; Rogelj, I.; Matijašić, B.B. Complete nisin A gene cluster from Lactococcus lactis M78 (HM219853): Obtaining the nucleic acid sequence and comparing it to other published nisin sequences. Genes Genom. 2011, 33, 217–221. [Google Scholar] [CrossRef]
- Parapouli, M.; Delbés-Paus, C.; Kakouri, A.; Koukkou, A.-I.; Montel, M.C.; Samelis, J. Characterization of a wild, novel nisin A-producing Lactococcus strain with an L. lactis subsp. cremoris genotype and an L. lactis subsp. lactis phenotype isolated from Greek raw milk. Appl. Environ. Microbiol. 2013, 79, 3476–3484. [Google Scholar]
- Vandera, E.; Parapouli, M.; Koukkou, A.-I.; Hatziloukas, E.; Samelis, J. Selection of Potential Probiotic Lactic Acid Bacteria Isolated from Traditional Dairy Products by in Vitro Tests. In Proceedings of the Book of Abstracts of the IAFP’s European Symposium on Food Safety, Athens, Greece, 11–13 May 2016; P2–24. pp. 115–116. [Google Scholar]
- Vandera, E.; Kakouri, A.; Koukkou, A.I.; Samelis, J. Major ecological shifts within the dominant nonstarter lactic acid bacteria in mature Greek Graviera cheese as affected by the starter culture type. Int. J. Food Microbiol. 2019, 290, 15–26. [Google Scholar] [CrossRef]
- Noutsopoulos, D.; Kakouri, A.; Kartezini, E.; Pappas, D.; Hatziloukas, E.; Samelis, J. Growth, nisA gene expression and in situ activity of novel Lactococcus lactis subsp. cremoris costarter culture in commercial hard cheese production. J. Food Prot. 2017, 80, 2137–2146. [Google Scholar]
- IDF. Cheese and Processed Cheese, Determination of the Total Solids Content (Reference Method); IDF Standard 4A; International Dairy Federation: Brussels, Belgium, 1982. [Google Scholar]
- BSI. Gerber Method for the Determination of Fat in Milk and Milk Products; British Standard 696; British Standards Institution: London, UK, 1955. [Google Scholar]
- IDF. Milk. Determination of Nitrogen Content; IDF Standard 20B; International Dairy Federation: Brussels, Belgium, 1993. [Google Scholar]
- Kosikowski, F.V. Cheese and Fermented Milk Foods; Edwards Brothers: Ann Arbor, MI, USA, 1982. [Google Scholar]
- AOAC. Ash of Cheese 935.42 Method. In Official Methods of Analysis EI, 16th ed.; Association of Official Analytical Chemists: Arlington, VA, USA, 1995; p. 59. [Google Scholar]
- Ling, R. A Text Book of Dairy Chemistry, 3rd ed.; Chapman & Hall: London, UK, 1963. [Google Scholar]
- Mallatou, H.; Pappa, E.C.; Boumba, V.A. Proteolysis in Telemecheese made from sheep’s, goats’ or a mixture of sheep’s and goats’ milk. Int. Dairy J. 2004, 14, 977–987. [Google Scholar] [CrossRef]
- Kuchroo, C.N.; Fox, P.F. Soluble nitrogen in Cheddar cheese. Comparison of the extraction procedures. Milchwissenschaft 1982, 37, 331–335. [Google Scholar]
- Stadhouders, J. The hydrolysis of proteins during the ripening of Dutch cheese.The enzymes and the bacteria involved. Neth. Milks Dairy J. 1960, 2, 67–76. [Google Scholar]
- Sousa, M.J.; Ardo, Y.; McSweeney, P.L.H. Advances in the study of proteolysis during cheese ripening. Int. Dairy J. 2001, 11, 327–345. [Google Scholar] [CrossRef]
- Pereira, C.I.; Graça, J.A.; Ogando, N.S.; Gomes, A.M.P.; Malcata, F.X. Influence of bacterial dynamics upon the final characteristics of model Portuguese traditional cheeses. Food Microbiol. 2010, 27, 339–346. [Google Scholar] [CrossRef]
- Mortera, P.; Zuljan, F.A.; Magni, C.; Bortolato, S.A.; Alarcón, S.H. Multivariate analysis of organic acids in fermented food from reversed-phase high-performance liquid chromatography data. Talanta 2018, 178, 15–23. [Google Scholar] [CrossRef] [PubMed]
- Kondyli, E.; Katsiari, M.C.; Voutsinas, L.P. Chemical and sensory characteristics of Galotyri-type cheese made using different procedures. Food Cont. 2008, 19, 301–307. [Google Scholar] [CrossRef]
- IDF. Sensory Evaluation of Dairy Products; IDF Standard 99A; International Dairy Federation: Brussels, Belgium, 1987. [Google Scholar]
- Recognition of a Protected Denomination of Origin (P.D.O.) for Galotyri cheese. In Newspaper of the Government of the Republic of Greece; Ministry of Finance, National Publishing Office: Athens, Greece, 1994; pp. 51–62.
- Michailidou, S.; Pavlou, E.; Pasentsis, K.; Rhoades, J.; Likotrafiti, E.; Agririou, A. Microbial profiles of Greek PDO cheeses assessed with amplicon metabarcoding. Food Microbiol. 2021, 99, 103836. [Google Scholar] [CrossRef]
- Zygouris, N.P. The Milk Industry, 2nd ed.; Ministry of Agriculture: Athens, Greece, 1952. (In Greek) [Google Scholar]
- Rogga, K.J.; Samelis, J.; Kakouri, A.; Katsiari, M.C.; Savvaidis, I.N.; Kontominas, M.G. Survival of Listeria monocytogenes in Galotyri, a traditional Greek soft acid-curd cheese, stored aerobically at 4 and 12 °C. Int. Dairy J. 2005, 15, 59–67. [Google Scholar] [CrossRef]
- Joishy, T.K.; Dehingia, M.; Khan, M.R. Bacterial diversity and metabolite profiles of curd prepared by natural fermentation of raw milk and back sloping of boiled milk. World J. Microbiol. Biotechnol. 2019, 35, 102. [Google Scholar] [CrossRef]
- Spyrelli, E.D.; Stamatiou, A.; Tassou, C.C.; Nychas, G.-J.E.; Doulgeraki, A.I. Microbiological and metagenomic analysis to assess the effect of container material on the microbiota of Feta cheese during ripening. Fermentation 2020, 6, 12. [Google Scholar] [CrossRef]
- Katsiari, M.C.; Kondyli, E.; Voutsinas, L.P. The quality of Galotyri-type cheese made with different starter cultures. Food Cont. 2009, 20, 113–118. [Google Scholar] [CrossRef]
- Samelis, J.; Kakouri, A. Microbial and safety qualities of PDO Galotyri cheese manufactured at the industrial or artisan scale in Epirus, Greece. Ital. J. Food Sci. 2007, 19, 91–99. [Google Scholar]
- Pexara, A.; Solomakos, N.; Sergelidis, D.; Govaris, A. Fate of enterotoxigenic Staphylococcus aureus and staphylococcal enterotoxins in Feta and Galotyri cheeses. J. Dairy Res. 2012, 79, 405–413. [Google Scholar] [CrossRef] [PubMed]
- Lekkas, C.; Kakouri, A.; Paleologos, E.; Voutsinas, L.P.; Kontominas, M.G.; Samelis, J. Survival of Escherichia coli O157:H7 in Galotyri cheese stored at 4 and 12 °C. Food Microbiol. 2006, 23, 268–276. [Google Scholar] [CrossRef] [PubMed]
- Kondyli, E.; Massouras, T.; Katsiari, M.C.; Voutsinas, L.P. Lipolysis and volatile compounds of Galotyri-type cheese made using different procedures. Small Rum. Res. 2013, 113, 432–436. [Google Scholar] [CrossRef]
- Salas, M.L.; Thierry, A.; Lemaitre, M.; Garric, G.; Harel-Oger, M.; Chatel, M.; Le, S.; Mounier, J.; Valence, F.; Coton, E. Antifungal activity of lactic acid bacteria combinations in dairy mimicking models and their potential as bioprotective cultures in pilot scale applications. Front. Microbiol. 2018, 9, 1787. [Google Scholar] [CrossRef]
- Kavkova, M.; Cihlar, J.; Drab, V.; Bazalova, O.; Dlouha, Z. The interactions among isolates of Lactiplantibacillus plantarum and dairy yeast contaminants: Towards biocontrol applications. Fermentation 2022, 8, 14. [Google Scholar] [CrossRef]
- Pisano, M.B.; Fadda, M.E.; Viale, S.; Deplano, M.; Mereu, F.; Blazic, M.; Cosentino, S. Inhibitory effect of Lactiplantibacillusplantarum and Lactococcus lactis autochthonous strains against Listeria monocytogenes in a laboratory cheese model. Foods 2022, 11, 715. [Google Scholar] [CrossRef]
- Xanthopoulos, V.; Hatzikamari, M.; Adamidis, T.; Tsakalidou, E.; Tzanetakis, N.; Litopoulou-Tzanetaki, E. Heterogeneity of Lactobacillus plantarum isolates from Feta cheese throughout ripening. J. Appl. Microbiol. 2000, 88, 1056–1064. [Google Scholar] [CrossRef]
- Manolopoulou, E.; Sarantinopoulos, P.; Zoidou, E.; Aktypis, A.; Moschopoulou, E.; Kandarakis, I.G.; Anifantakis, M.E. Evolution of microbial populations during traditional Feta cheese manufacture and ripening. Int. J. Food Microbiol. 2003, 82, 153–161. [Google Scholar] [CrossRef] [PubMed]
- Zoumpopoulou, G.; Tzouvanou, A.; Mavrogonatou, E.; Alexandraki, V.; Georgalaki, M.; Anastasiou, R.; Papadelli, M.; Manolopoulou, E.; Kazou, M.; Kletsas, D.; et al. Probiotic features of lactic acid bacteria isolated from a diverse pool of traditional Greek dairy products regarding specific strain-host interactions. Prob. Antimicrob. Prot. 2018, 10, 313–322. [Google Scholar] [CrossRef] [PubMed]
- Papadimitriou, K.; Anastasiou, R.; Georgalaki, M.; Bounenni, R.; Paximadaki, A.; Charmpi, C.; Alexandraki, V.; Kazou, M.; Tsakalidou, E. Comparison of the microbiome of artisanal homemade and industrial Feta cheese through amplicon sequencing and shortgun metagenomics. Microorganisms 2022, 10, 1073. [Google Scholar] [CrossRef] [PubMed]
- Maragkoudakis, P.A.; Miaris, C.; Rojez, P.; Manalis, N.; Magkanari, F.; Kalantzopulos, G.; Tsakalidou, E. Production of traditional Greek yogurt using Lactobacillus strains with probiotic potential as starter adjuncts. Int. Dairy J. 2006, 16, 52–60. [Google Scholar] [CrossRef]
- Saxami, G.; Papadopoulou, O.; Chorianopoulos, N.; Kourkoutas, Y.; Tassou, C.; Galanis, A. Molecular detection of two potential probiotic lactobacilli strains and evaluation of their performance as starter adjuncts in yogurt production. Int. J. Mol. Sci. 2016, 17, 668. [Google Scholar] [CrossRef]
- Binda, S.; Hill, C.; Johansen, E.; Obis, D.; Pot, B.; Sanders, M.E.; Tremblay, A.; Ouwehand, A.C. Criteria to qualify microorganisms as “probiotic” in foods and dietary supplements. Front. Microbiol. 2020, 11, 1662. [Google Scholar] [CrossRef]
- Kallinteri, L.D.; Kostoula, O.K.; Savvaidis, I.N. Efficacy of nisin and/or natamycin to improve the shelf-life of Galotyri cheese. Food Microbiol. 2013, 36, 176–181. [Google Scholar] [CrossRef]
- Pappa, E.C.; Kondyli, E. Descriptive characteristics and cheesemaking technology of Greek cheeses not listed in the EU geographical indications registers. Dairy 2023, 4, 43–67. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Starter or Adjunct LAB Strain | Galotyri Cheese A (Basic Starter Mix: ST1+M78) 2 | Galotyri Cheese B (Starter Mix: ST1+M78+H25) 2 | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Trial/ Batch | Cheese Milk | Acidified Curd | Fresh RTE Cheese | Ripened Cheese | Trial/ Batch | Cheese Milk | Acidified Curd | Fresh RTE Cheese | Ripened Cheese | |
| Streptococcus thermophilus ST1 | ||||||||||
| A1 | 7.19 ± 0.06 cA | <6.00 bA | 7.67 ± 0.10 cB | 5.87 ± 0.02 bA | B1 | 7.39 ± 0.04 cB | <6.00 bA | <5.00 aA | 5.41 ± 0.15 bA | |
| A2 | 6.36 ± 0.06 aA | 9.17 ± 0.00 bC | 9.09 ± 0.00 bC | 8.95 ± 0.07 bB | B2 | 6.27 ± 0.01 aA | 8.72 ± 0.00 bC | 9.09 ± 0.00 bC | 9.01 ± 0.03 bC | |
| A3 | 6.77 ± 0.10 aA | 6.83 ± 0.11 aB | 6.86 ± 0.01 aA | 5.90 ± 0.62 bA | B3 | 6.88 ± 0.04 aAB | 6.95 ± 0.08 aB | 7.10 ± 0.18 aB | 6.41 ± 0.01 abB | |
| Lactococcus lactis sub. cremoris M78 | ||||||||||
| A1 | 6.52 ± 0.02 aA | <7.00 aA | 7.57 ± 0.24 bA | 6.30 ± 0.00 aB | B1 | 6.59 ± 0.08 aA | 8.97 ± 0.16 cB | 9.19 ± 0.01 cB | 7.08 ± 0.32 abB | |
| A2 | 6.21 ± 0.03 cA | 8.06 ± 0.13 dB | 8.20 ± 0.03 dB | <4.00 aA | B2 | 6.22 ± 0.03 cA | 7.96 ± 0.04 dA | 7.98 ± 0.10 dA | 5.30 ± 0.00 bA | |
| A3 | 6.56 ± 0.06 aA | 9.19 ± 0.01 cC | 9.34 ± 0.01 cC | 7.82 ± 0.45 bC | B3 | 6.57 ± 0.00 aA | 9.18 ± 0.00 cB | 9.00 ± 0.02 cB | 6.71 ± 0.01 aB | |
| Lactiplantibacillus plantarum H25 | ||||||||||
| A1 | NA | NA | NA | NA | B1 | 5.63 ± 0.32 aA | 7.24 ± 0.06 bA | 7.30 ± 0.42 bA | 6.22 ± 0.37 aA | |
| A2 | NA | NA | NA | NA | B2 | 6.08 ± 0.21 aA | 7.08 ± 0.00 bA | 7.74 ± 0.06 cAB | 7.77 ± 0.05 cC | |
| A3 | NA | NA | NA | NA | B3 | 6.15 ± 0.08 aA | 8.92 ± 0.00 cB | 8.21 ± 0.13 cB | 6.98 ± 0.07 bB | |
| Type of Microbial Contamination | Galotyri Cheese A (Basic Starter Mix: ST1+M78) 2 | Galotyri Cheese B (Starter Mix: ST1+M78+H25) 2 | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Trial/ Batch | Cheese Milk | Acidified Curd | Fresh RTE Cheese | Ripened Cheese | Trial/ Batch | Cheese Milk | Acidified Curd | Fresh RTE Cheese | Ripened Cheese | |
| Enterococci | A1 | <2.00 aA | 5.09 ± 0.27 cC | 5.96 ± 0.28 dC | 5.48 ± 0.10 cdC | B1 | <2.00 aA | <2.00 aA | 4.04 ± 0.37 bB | 4.78 ± 0.10 cC |
| A2 | <2.00 aA | <2.00 aA | <2.00 aA | <2.00 aA | B2 | <2.00 aA | <2.00 aA | <2.00 aA | <2.00 aA | |
| A3 | <2.00 aA | 3.77 ± 0.12 cB | 3.76 ± 0.04 cB | 2.90 ± 0.00 bB | B3 | <2.00 aA | <2.00 aA | <2.00 aA | 2.60 ± 0.00 bB | |
| Coliform bacteria | A1 | <1.00 aA | 7.80 ± 0.04 dC | 6.25 ± 0.07 cC | 4.38 ± 0.00 bC | B1 | <1.00 aA | 4.25 ± 0.07 bB | 3.97 ± 0.15 bB | <1.00 aA |
| A2 | <1.00 aA | <1.00 aA | <1.00 aA | <1.00 aA | B2 | <1.00 aA | <1.00 aA | <1.00 aA | <1.00 aA | |
| A3 | <1.00 aA | 3.31 ± 0.10 cB | 2.85 ± 0.35 bcB | 2.30 ± 0.01 bB | B3 | <1.00 aA | <1.00 aA | <1.00 aA | <1.00 aA | |
| Pseudomonad-like bacteria | A1 | <2.00 aA | 7.51 ± 0.10 cB | 7.05 ± 0.10 cB | 5.82 ± 0.05 * | B1 | <2.00 aA | 4.01 ± 0.15 bB | 3.63 ± 0.31 bB | 5.14 ± 0.10 * |
| A2 | <2.00 aA | <2.00 aA | <2.00 aA | <2.00 aA | B2 | <2.00 aA | <2.00 aA | <2.00 aA | <2.00 aA | |
| A3 | <2.00 aA | <2.00 aA | <2.00 aA | <2.00 aA | B3 | <2.00 aA | <2.00 aA | <2.00 aA | <2.00 aA | |
| Total staphylococci | A1 | <2.00 aA | 4.91 ± 0.08 cC | <2.00 aA | 2.54 ± 0.34 bB | B1 | <2.00 aA | <2.00 aA | <2.00 aA | <2.00 aA |
| A2 | <2.00 aA | <2.00 aA | <2.00 aA | <2.00 aA | B2 | <2.00 aA | <2.00 aA | <2.00 aA | <2.00 aA | |
| A3 | <2.00 aA | 4.06 ± 0.04 cB | <2.00 aA | 2.47 ± 0.00 bB | B3 | <2.00 aA | 4.33 ± 0.01 cB | 2.70 ± 0.01 bB | <2.00 aA | |
| Yeasts | A1 | <2.00 aA | <2.00 aA | 3.48 ± 0.00 cB | 6.40 ± 0.01 eA | B1 | <2.00 aA | <2.00 aA | <3.00 bA | 5.52 ± 0.13 dA |
| A2 | <2.00 aA | <2.00 aA | 4.74 ± 0.17 bC | 7.98 ± 0.18 dB | B2 | <2.00 aA | <2.00 aA | 4.30 ± 0.01 bB | 6.92 ± 0.12 cC | |
| A3 | <2.00 aA | 2.60 ± 0.01 bB | <3.00 bA | 6.65 ± 0.02 dA | B3 | <2.00 aA | 3.28 ± 0.03 cB | <3.00 bA | 6.25 ± 0.02 dB | |
| Parameter | Galotyri Cheese A (Basic Starter Mix: ST1+M78) 2 | Galotyri Cheese B (Starter Mix: ST1+M78+H25) 2 | ||
|---|---|---|---|---|
| Fresh RTE Cheese | Ripened Cheese | Fresh RTE Cheese | Ripened Cheese | |
| pH | 4.57 ± 0.21 a | 4.61 ± 0.19 a | 4.52 ± 0.15 a | 4.55 ± 0.20 a |
| Moisture (%) | 66.32 ± 5.46 a | 65.59 ± 6.44 a | 67.81 ± 4.17 a | 66.59 ± 3.29 a |
| Fat (%) | 12.25 ± 0.66 ab | 13.00 ± 1.80 b | 11.88 ± 0.82 a | 12.25 ± 0.43 ab |
| Protein (%) | 12.15 ± 0.63 b | 11.77 ± 0.27 ab | 11.44 ± 1.50 ab | 10.62 ± 1.08 a |
| Salt (NaCl) (%) | 2.23 ± 0.80 a | 2.29 ± 0.92 a | 2.05 ± 0.49 a | 1.98 ± 0.52 a |
| Ash (%) | 2.85 ± 0.76 a | 2.73 ± 0.79 a | 2.51 ± 0.30 a | 2.51 ± 0.45 a |
| Acidity (%) | 1.19 ± 0.42 a | 1.25 ± 0.36 a | 1.18 ± 0.47 a | 1.27 ± 0.38 a |
| Biochemical Parameter | Galotyri Cheese A (Basic Starter Mix: ST1+M78) 2 | Galotyri Cheese B (Starter Mix: ST1+M78+H25) 2 | ||
|---|---|---|---|---|
| Fresh RTE Cheese | Ripened RTE Cheese | Fresh RTE Cheese | Ripened Cheese | |
| Lactose | 2.69 ± 0.86 a | 2.55 ± 0.90 a | 3.07 ± 1.01 a | 2.44 ± 0.70 a |
| D-glucose | n.d. | n.d | n.d. | n.d. |
| D-galactose | 0.54 ± 0.17 b | 0.25 ± 0.15 a | 0.55 ± 0.14 b | 0.43 ± 0.25 ab |
| Lactic acid | 2.47 ± 1.17 a | 2.50 ± 0.63 a | 2.62 ± 1.15 a | 2.34 ± 1.26 a |
| Citric acid | 0.22 ± 0.14 a | 0.21 ± 0.07 a | 0.27 ± 0.12 a | 0.23 ± 0.14 a |
| Acetic acid | n.d. | 1.04/n.d./n.d. | 1.68/n.d./n.d. | 1.63/n.d./n.d. |
| Propionic acid | 0.46/0.58/n.d. | 0.36/0.56/n.d. | 0.28/0.55/n.d. | 0.16/0.54/n.d. |
| Protein Fraction | Galotyri Cheese A (Basic Starter Mix: ST1+M78) 2 | Galotyri Cheese B (Starter Mix: ST1+M78+H25) 2 | ||||
|---|---|---|---|---|---|---|
| Trial/Batch | Fresh RTE Cheese | Ripened Cheese | Trial/Batch | Fresh RTE Cheese | Ripened Cheese | |
| Total nitrogen (TN%) | A1 | 2.01 | 1.89 | B1 | 1.55 | 1.48 |
| A2 | 1.89 | 1.81 | B2 | 1.81 | 1.82 | |
| A3 | 1.81 | 1.84 | B3 | 2.02 | 2.01 | |
| Mean ± SD | 1.90 ± 0.10 a | 1.85 ± 0.04 a | Mean ± SD | 1.79 ± 0.24 a | 1.77 ± 0.27 a | |
| WSN a (%TN) | A1 | 9.37 | 9.97 | B1 | 10.75 | 11.17 |
| A2 | 9.15 | 7.66 | B2 | 6.93 | 6.23 | |
| A3 | 5.18 | 7.12 | B3 | 4.13 | 6.93 | |
| Mean ± SD | 7.90 ± 1.93 a | 8.25 ± 1.24 a | Mean ± SD | 7.27 ± 2.71 a | 8.11 ± 2.18 a | |
| TCA b (%TN) | A1 | 8.57 | 9.15 | B1 | 10.84 | 11.07 |
| A2 | 5.56 | 6.42 | B2 | 6.27 | 5.89 | |
| A3 | 4.32 | 5.89 | B3 | 5.69 | 6.38 | |
| Mean ± SD | 6.15 ± 1.78 a | 7.15 ± 1.43 a | Mean ± SD | 7.16 ± 2.3 a | 7.78 ± 2.33 a | |
| PTA c (%TN) | A1 | 5.78 | 6.00 | B1 | 6.37 | 6.91 |
| A2 | 4.11 | 4.18 | B2 | 4.22 | 4.89 | |
| A3 | 5.72 | 6.67 | B3 | 5.57 | 6.10 | |
| Mean ± SD | 5.20 ±0.77 a | 5.62 ± 1.05 a | Mean ± SD | 5.39 ± 0.89 a | 5.97 ± 0.83 a | |
| REI d | A1 | 4.66 | 5.27 | B1 | 6.94 | 7.55 |
| A2 | 4.84 | 4.23 | B2 | 3.83 | 3.42 | |
| A3 | 2.86 | 3.87 | B3 | 2.04 | 3.45 | |
| Mean ± SD | 4.12 ± 0.89 a | 4.46 ± 0.59 a | Mean ± SD | 4.27 ± 2.02 a | 4.80 ± 1.94 a | |
| RDI e | A1 | 4.27 | 4.84 | B1 | 6.99 | 7.48 |
| A2 | 2.94 | 3.55 | B2 | 3.46 | 3.24 | |
| A3 | 3.16 | 3.20 | B3 | 2.82 | 3.17 | |
| Mean ± SD | 3.46 ± 0.58 a | 3.86 ± 0.71 a | Mean ± SD | 4.42 ± 1.83 a | 4.63 ± 2.02 a | |
| FAAI f | A1 | 2.88 | 3.18 | B1 | 4.11 | 4.67 |
| A2 | 2.18 | 2.31 | B2 | 2.33 | 2.69 | |
| A3 | 3.16 | 3.62 | B3 | 2.76 | 3.03 | |
| Mean ± SD | 2.74 ± 0.41 a | 3.04 ± 0.54 a | Mean ± SD | 3.07 ± 0.76 a | 3.46 ± 0.86 a | |
| Cheese Trial/Batch | Appearance (10) 2 | Body/Texture (40) | Flavor (50) | Total Quality (100) |
|---|---|---|---|---|
| A1 (ST1+M78) 3 | 8.0 ± 1.2 a | 33.6 ± 2.2 a | 29.0 ± 2.2 a | 70.6 ± 2.6 a |
| B1 (ST1+M78+H25) | 9.0 ± 0.7 a | 33.6 ± 1.7 a | 42.5 ± 2.5 b | 85.1 ± 4.6 b |
| A2 (ST1+M78) | 9.2 ± 0.4 a | 35.2 ± 1.8 a | 43.0 ± 4.5 b | 87.4 ± 5.1 b |
| B2 (ST1+M78+H25) | 9.2 ± 0.4 a | 36.5 ± 3.0 a | 44.8 ± 4.8 b | 90.5 ± 7.8 b |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Samelis, J.; Tsanasidou, C.; Bosnea, L.; Ntziadima, C.; Gatzias, I.; Kakouri, A.; Pappas, D. Pilot-Scale Production of Traditional Galotyri PDO Cheese from Boiled Ewes’ Milk Fermented with the Aid of Greek Indigenous Lactococcus lactis subsp. cremoris Starter and Lactiplantibacillus plantarum Adjunct Strains. Fermentation 2023, 9, 345. https://doi.org/10.3390/fermentation9040345
Samelis J, Tsanasidou C, Bosnea L, Ntziadima C, Gatzias I, Kakouri A, Pappas D. Pilot-Scale Production of Traditional Galotyri PDO Cheese from Boiled Ewes’ Milk Fermented with the Aid of Greek Indigenous Lactococcus lactis subsp. cremoris Starter and Lactiplantibacillus plantarum Adjunct Strains. Fermentation. 2023; 9(4):345. https://doi.org/10.3390/fermentation9040345
Chicago/Turabian StyleSamelis, John, Charikleia Tsanasidou, Loulouda Bosnea, Charikleia Ntziadima, Ilias Gatzias, Athanasia Kakouri, and Dimitrios Pappas. 2023. "Pilot-Scale Production of Traditional Galotyri PDO Cheese from Boiled Ewes’ Milk Fermented with the Aid of Greek Indigenous Lactococcus lactis subsp. cremoris Starter and Lactiplantibacillus plantarum Adjunct Strains" Fermentation 9, no. 4: 345. https://doi.org/10.3390/fermentation9040345
APA StyleSamelis, J., Tsanasidou, C., Bosnea, L., Ntziadima, C., Gatzias, I., Kakouri, A., & Pappas, D. (2023). Pilot-Scale Production of Traditional Galotyri PDO Cheese from Boiled Ewes’ Milk Fermented with the Aid of Greek Indigenous Lactococcus lactis subsp. cremoris Starter and Lactiplantibacillus plantarum Adjunct Strains. Fermentation, 9(4), 345. https://doi.org/10.3390/fermentation9040345