
Evaluation of Laminaria Digitata Hydrolysate for the Production of Bioethanol and Butanol by Fermentation
 ,
,  ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Hydrolysate
2.2. Ethanol Fermentation
2.2.1. Fermentation by Saccharomyces cerevisiae
2.2.2. Fermentation by Thermoanaerobacterium Strain AK17
2.3. ABE and IBE Fermentation
2.4. Analysis of Free Organic Acids and Monosaccharides by HPLC
2.5. Total Monosaccharides Analysis by HPAEC-PAD
2.6. Yields Calculation Formulas
2.7. Determination of Extracellular and Intracellular Enzymatic Activity in the Thermoanaerobacterium Strain AK17 Culture
3. Results
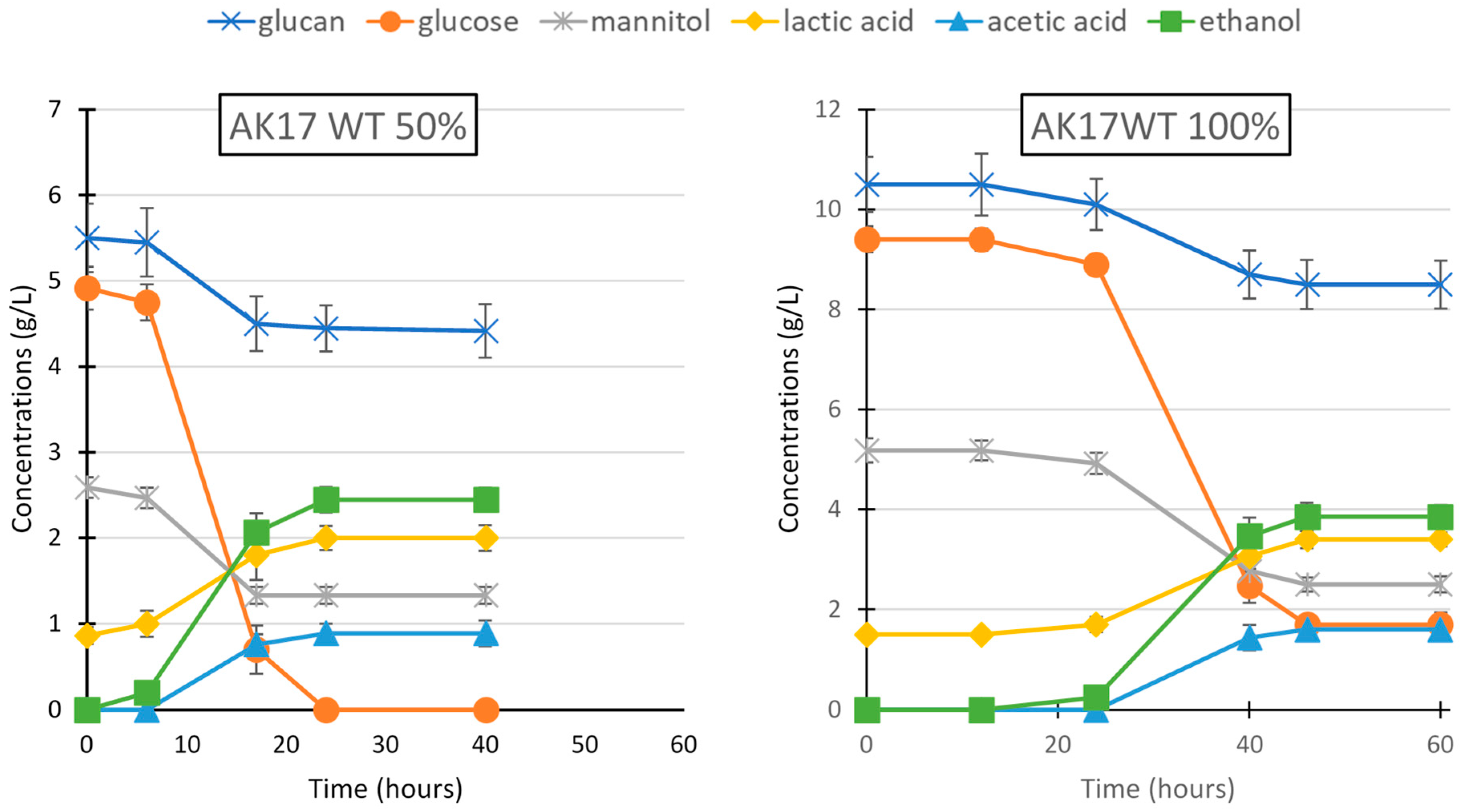
3.1. L. digitata Hydrolysate Utilization by S. cerevisiae and Thermoanaerobacterium Strain AK17 for Production of Ethanol
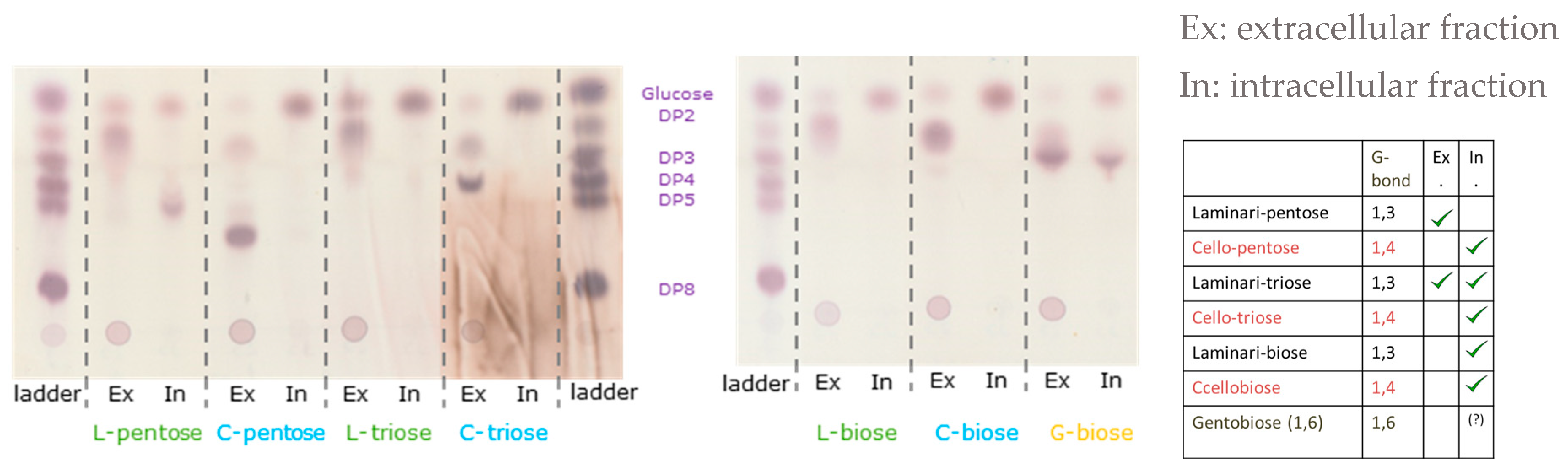
3.2. Oligosaccharide Degradation and Uptake by Thermoanaerobacterium Strain AK17
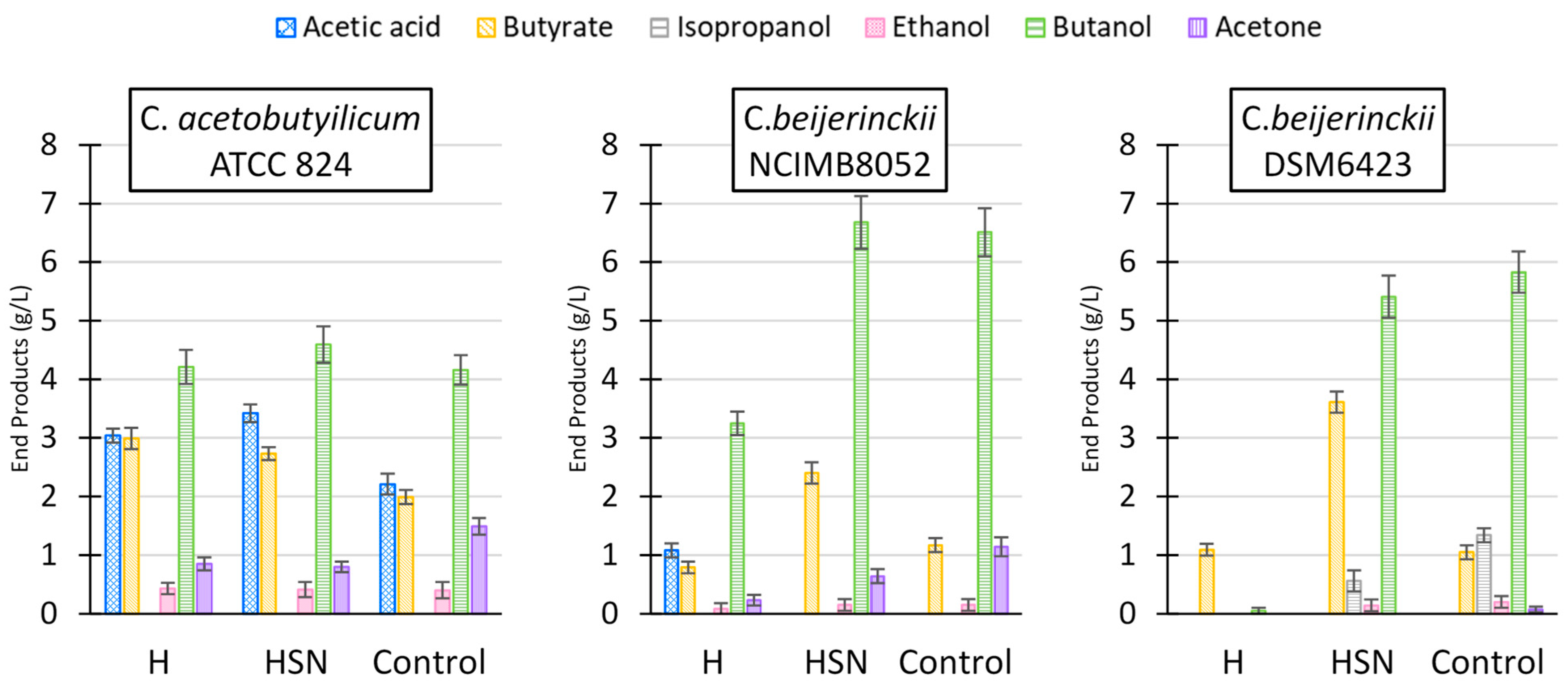
3.3. L. digitata Hydrolysate Utilization by Clostridial Strains for Production of A/IBE
4. Discussion
4.1. Strain AK17 and Ethanol Production
4.2. Butanol Production by Mesophilic Clostridia
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- BP Statistical Review of World Energy Globally Consistent Data on World Energy Markets and Authoritative Publications in the Field of Energy The Statistical Review World of World Energy and Data on World Energy Markets from Is The Review Has Been Providing. 2020, 66. Available online: https://www.bp.com/content/dam/bp/business-sites/en/global/corporate/pdfs/energy-economics/statistical-review/bp-stats-review-2020-full-report.pdf (accessed on 1 November 2022).
- Bentsen, N.S.; Felby, C. Biomass for energy in the European Union—A review of bioenergy resource assessments. Biotechnol. Biofuels 2012, 5, 25. [Google Scholar] [CrossRef]
- Kelly, M.S.; Dworjanyn, S. The Potential of Marine Biomass for Anaerobic Biogas Production; Crown Estate: London, UK, 2008; ISBN 978-1-906410-05-6. [Google Scholar]
- Tedesco, S.; Stokes, J. Valorisation to biogas of macroalgal waste streams: A circular approach to bioproducts and bioenergy in Ireland. Chem. Pap. 2017, 71, 721–728. [Google Scholar] [CrossRef]
- Zabed, H.; Sahu, J.N.; Boyce, A.N.; Faruq, G. Fuel ethanol production from lignocellulosic biomass: An overview on feedstocks and technological approaches. Renew. Sustain. Energy Rev. 2016, 66, 751–774. [Google Scholar] [CrossRef]
- Chung, I.K.; Beardall, J.; Mehta, S.; Sahoo, D.; Stojkovic, S. Using marine macroalgae for carbon sequestration: A critical appraisal. J. Appl. Phycol. 2011, 23, 877–886. [Google Scholar] [CrossRef]
- Adams, J.M.; Gallagher, J.A.; Donnison, I. Fermentation study on Saccharina latissima for bioethanol production considering variable pre-treatments. J. Appl. Phycol. 2009, 21, 569–574. [Google Scholar] [CrossRef]
- Minh, T.H.; Hanh, V. Bioethanol production from marine algae biomass: Prospect and troubles. J. Vietnam. Environ. 2012, 3, 25–29. [Google Scholar] [CrossRef]
- Kawai, S.; Murata, K. Biofuel Production Based on Carbohydrates from Both Brown and Red Macroalgae: Recent Developments in Key Biotechnologies. Int. J. Mol. Sci. 2016, 17, 145. [Google Scholar] [CrossRef]
- Manns, D.; Deutschle, A.L.; Saake, B.; Meyer, A.S. Methodology for quantitative determination of the carbohydrate composition of brown seaweeds (Laminariaceae). RSC Adv. 2014, 4, 25736–25746. [Google Scholar] [CrossRef]
- Manns, D.; Nielsen, M.M.; Bruhn, A.; Saake, B.; Meyer, A.S. Compositional variations of brown seaweeds Laminaria digitata and Saccharina latissima in Danish waters. J. Appl. Phycol. 2017, 29, 1493–1506. [Google Scholar] [CrossRef]
- Ferdouse, F.; Løvstad Holdt, S.; Smith, R.; Murúa, P.; Yang, Z. The Global Status of Seaweed Production, Trade and Utilization. FAO Globefish Res. Program. 2018, 124, 120. [Google Scholar]
- Radulovich, R.; Neori, A.; Valderrama, D.; Reddy, C.R.K.; Cronin, H.; Forster, J. Farming of Seaweeds; Elsevier Inc.: Amsterdam, The Netherlands, 2015; ISBN 9780124199583. [Google Scholar]
- Kostas, E.T.; Adams, J.M.M.; Ruiz, H.A.; Durán-Jiménez, G.; Lye, G.J. Macroalgal biorefinery concepts for the circular bioeconomy: A review on biotechnological developments and future perspectives. Renew. Sustain. Energy Rev. 2021, 151, 111553. [Google Scholar] [CrossRef]
- Montingelli, M.E.; Tedesco, S.; Olabi, A.G. Biogas production from algal biomass: A review. Renew. Sustain. Energy Rev. 2015, 43, 961–972. [Google Scholar] [CrossRef]
- Yan, L.; Wang, Y.; Li, J.; Zhang, Y.; Ma, L.; Fu, F.; Chen, B.; Liu, H. Hydrothermal liquefaction of Ulva prolifera macroalgae and the influence of base catalysts on products. Bioresour. Technol. 2019, 292, 121286. [Google Scholar] [CrossRef]
- Yuan, C.; Wang, S.; Cao, B.; Hu, Y.; Abomohra, A.E.-F.; Wang, Q.; Qian, L.; Liu, L.; Liu, X.; He, Z.; et al. Optimization of hydrothermal co-liquefaction of seaweeds with lignocellulosic biomass: Merging 2nd and 3rd generation feedstocks for enhanced bio-oil production. Energy 2019, 173, 413–422. [Google Scholar] [CrossRef]
- Maneein, S.; Milledge, J.J.; Nielsen, B.V.; Harvey, P.J. A Review of Seaweed Pre-Treatment Methods for Enhanced Biofuel Production by Anaerobic Digestion or Fermentation. Fermentation 2018, 4, 100. [Google Scholar] [CrossRef]
- Li, J.; He, Z.; Liang, Y.; Peng, T.; Hu, Z. Insights into Algal Polysaccharides: A Review of Their Structure, Depolymerases, and Metabolic Pathways. J. Agric. Food Chem. 2022, 70, 1749–1765. [Google Scholar] [CrossRef]
- Hreggviðsson, G.; Nordberg-Karlsson, E.M.; Tøndervik, A.; Aachmann, F.L.; Dobruchowska, J.M.; Linares-Pastén, J.; Daugbjerg-Christensen, M.; Moenaert, A.; Kristjansdottir, T.; Sletta, H.; et al. Biocatalytic refining of polysaccharides from brown seaweeds. Sustain. Seaweed Technol. 2020, 2020, 447–504. [Google Scholar] [CrossRef]
- Lee, O.K.; Lee, E.Y. Sustainable production of bioethanol from renewable brown algae biomass. Biomass Bioenergy 2016, 92, 70–75. [Google Scholar] [CrossRef]
- Jiang, R.; Ingle, K.N.; Golberg, A. Macroalgae (seaweed) for liquid transportation biofuel production: What is next? Algal Res. 2016, 14, 48–57. [Google Scholar] [CrossRef]
- Ji, S.-Q.; Wang, B.; Lu, M.; Li, F.-L. Direct bioconversion of brown algae into ethanol by thermophilic bacterium Defluviitalea phaphyphila. Biotechnol. Biofuels 2016, 9, 81. [Google Scholar] [CrossRef]
- Kim, N.-J.; Li, H.; Jung, K.; Chang, H.N.; Lee, P.C. Ethanol production from marine algal hydrolysates using Escherichia coli KO11. Bioresour. Technol. 2011, 102, 7466–7469. [Google Scholar] [CrossRef]
- Enquist-Newman, M.; Faust, A.M.E.; Bravo, D.D.; Santos, C.N.S.; Raisner, R.M.; Hanel, A.; Sarvabhowman, P.; Le, C.; Regitsky, D.D.; Cooper, S.R.; et al. Efficient ethanol production from brown macroalgae sugars by a synthetic yeast platform. Nature 2013, 505, 239–243. [Google Scholar] [CrossRef]
- Hou, X.; From, N.; Angelidaki, I.; Huijgen, W.J.J.; Bjerre, A.-B. Butanol fermentation of the brown seaweed Laminaria digitata by Clostridium beijerinckii DSM-6422. Bioresour. Technol. 2017, 238, 16–21. [Google Scholar] [CrossRef]
- Xue, C.; Cheng, C. Butanol production by Clostridium. Adv. Bioenergy 2019, 4, 35–77. [Google Scholar] [CrossRef]
- Contreras, A.M.L.; Kuit, W.; Siemerink, M.A.J.; Kengen, S.W.M.; Springer, J.; Claassen, P.A.M. Production of longer-chain alcohols from lignocellulosic biomass: Butanol, isopropanol and 2,3-butanediol. Bioalcohol. Prod. Biochem. Convers. Lignocellul. Biomass 2010, 415–460. [Google Scholar] [CrossRef]
- Hon, S.; Olson, D.G.; Holwerda, E.K.; Lanahan, A.A.; Murphy, S.J.L.; Maloney, M.I.; Zheng, T.; Papanek, B.; Guss, A.M.; Lynd, L.R. The ethanol pathway from Thermoanaerobacterium saccharolyticum improves ethanol production in Clostridium thermocellum. Metab. Eng. 2017, 42, 175–184. [Google Scholar] [CrossRef]
- Schultze-Jena, A.; Vroon, R.C.; Macleod, A.K.A.; Hreggviðsson, G.; Adalsteinsson, B.T.; Engelen-Smit, N.P.E.; de Vrije, T.; Budde, M.A.W.; van der Wal, H.; López-Contreras, A.M.; et al. Production of acetone, butanol, and ethanol by fermentation of Saccharina latissima: Cultivation, enzymatic hydrolysis, inhibitor removal, and fermentation. Algal Res. 2022, 62, 102618. [Google Scholar] [CrossRef]
- Koskinen, P.E.P.; Beck, S.R.; Örlygsson, J.; Puhakka, J.A. Ethanol and hydrogen production by two thermophilic, anaerobic bacteria isolated from Icelandic geothermal areas. Biotechnol. Bioeng. 2008, 101, 679–690. [Google Scholar] [CrossRef]
- Sveinsdottir, M.; Beck, S.R.; Orlygsson, J. Ethanol Production from Monosugars and Lignocellulosic Biomass by Thermophilic Bacteria Isolated from Icelandic Hot Springs. Icel. Agric. Sci. 2009, 22, 45–58. [Google Scholar]
- Van Wychen, S.; Laurens, L.M.L. Determination of Total Carbohydrates in Algal Biomass: Laboratory Analytical Procedure (LAP); National Renewable Energy Laboratory: Golden, CO, USA, 2015; p. 18.
- Allahgholi, L.; Sardari, R.R.R.; Hakvåg, S.; Ara, K.Z.G.; Kristjansdottir, T.; Aasen, I.M.; Fridjonsson, O.H.; Brautaset, T.; Hreggvidsson, G.O.; Karlsson, E.N. Composition analysis and minimal treatments to solubilize polysaccharides from the brown seaweed Laminaria digitata for microbial growth of thermophiles. J. Appl. Phycol. 2020, 32, 1933–1947. [Google Scholar] [CrossRef]
- Aparicio, E.; Rodríguez-Jasso, R.M.; Pinales-Márquez, C.D.; Loredo-Treviño, A.; Robledo-Olivo, A.; Aguilar, C.N.; Kostas, E.T.; Ruiz, H.A. High-pressure technology for Sargassum spp biomass pretreatment and fractionation in the third generation of bioethanol production. Bioresour. Technol. 2021, 329, 124935. [Google Scholar] [CrossRef] [PubMed]
- González-Gloria, K.D.; Rodríguez-Jasso, R.M.; Rosero-Chasoy, G.; Shiva; Kostas, E.T.; Aparicio, E.; Sanchez, A.; López-Sandin, I.; Ruiz, H.A. Scale-up of hydrothermal processing: Liquid hot water and pilot-scale tubular steam explosion batch reactor for bioethanol production using macroalgae Sargassum spp biomass. Bioresour. Technol. 2023, 369, 128448. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.M.; Wi, S.G.; Jung, S.; Song, Y.; Bae, H.-J. Efficient approach for bioethanol production from red seaweed Gelidium amansii. Bioresour. Technol. 2015, 175, 128–134. [Google Scholar] [CrossRef]
- Almarsdottir, A.R.; Sigurbjornsdottir, M.A.; Orlygsson, J. Effect of various factors on ethanol yields from lignocellulosic biomass by Thermoanaerobacterium AK17. Biotechnol. Bioeng. 2012, 109, 686–694. [Google Scholar] [CrossRef] [PubMed]
- Di Donato, P.; Finore, I.; Poli, A.; Nicolaus, B.; Lama, L. The production of second generation bioethanol: The biotechnology potential of thermophilic bacteria. J. Clean. Prod. 2019, 233, 1410–1417. [Google Scholar] [CrossRef]
- Taylor, M.P.; Eley, K.L.; Martin, S.; Tuffin, M.I.; Burton, S.G.; Cowan, D.A. Thermophilic ethanologenesis: Future prospects for second-generation bioethanol production. Trends Biotechnol. 2009, 27, 398–405. [Google Scholar] [CrossRef]
- Qu, C.; Chen, L.; Li, Y.; Fu, H.; Wang, J. The redox-sensing transcriptional repressor Rex is important for regulating the products distribution in Thermoanaerobacterium aotearoense SCUT27. Appl. Microbiol. Biotechnol. 2020, 104, 5605–5617. [Google Scholar] [CrossRef]
- Chades, T.; Scully, S.M.; Ingvadottir, E.M.; Orlygsson, J. Fermentation of Mannitol Extracts From Brown Macro Algae by Thermophilic Clostridia. Front. Microbiol. 2018, 9, 1931. [Google Scholar] [CrossRef]
- Heyndrickx, M.; De Vos, P.; Ley, J. De Fermentation Characteristics of Clostridium Pasteurianum LMG 3285 Grown on Glucose and Mannitol. J. Appl. Microbiol. 1991, 70, 52–58. [Google Scholar]
- Fu, H.; Hu, J.; Guo, X.; Feng, J.; Zhang, Y.; Wang, J. High-Selectivity Butyric Acid Production from Saccharina japonica Hydrolysate by Clostridium tyrobutyricum. Ind. Eng. Chem. Res. 2020, 59, 17147–17155. [Google Scholar] [CrossRef]
- Oh, H.J.; Kim, K.-Y.; Lee, K.M.; Lee, S.-M.; Gong, G.; Oh, M.-K.; Um, Y. Enhanced butyric acid production using mixed biomass of brown algae and rice straw by Clostridium tyrobutyricum ATCC25755. Bioresour. Technol. 2019, 273, 446–453. [Google Scholar] [CrossRef] [PubMed]
- López-Contreras, A.M.; Harmsen, P.F.H.; Blaauw, R.; Houweling-Tan, B.; Van Der Wal, H.; Huijgen, W.J.J.; Van Hal, J.W. Biorefinery of the Brown Seaweed Saccharina Latissima for Fuels and Chemicals. In Proceedings of the Mie Bioforum on lignocellulose degradation and biorefinery, Mie Prefecture, Japan, 18–21 November 2014; pp. 7–8. [Google Scholar]
- Huesemann, M.H.; Kuo, L.-J.; Urquhart, L.; Gill, G.; Roesijadi, G. Acetone-butanol fermentation of marine macroalgae. Bioresour. Technol. 2012, 108, 305–309. [Google Scholar] [CrossRef]
- Mitchell, W.J.; Albasheri, K.A.; Yazdanian, M. Factors affecting utilization of carbohydrates by clostridia. FEMS Microbiol. Rev. 1995, 17, 317–329. [Google Scholar] [CrossRef]
- López-Contreras, A.M.; Claassen, P.A.M.; Mooibroek, H.; De Vos, W.M. Utilisation of Saccharides in Extruded Domestic Organic Waste by Clostridium Acetobutylicum ATCC 824 for Production of Acetone, Butanol and Ethanol. Appl. Microbiol. Biotechnol. 2000, 54, 162–167. [Google Scholar] [CrossRef]
- Madihah, M.S.; Ariff, A.B.; Sahaid, K.M.; Suraini, A.A.; Karim, M.I.A. Direct fermentation of gelatinized sago starch to acetone–butanol–ethanol by Clostridium acetobutylicum. World J. Microbiol. Biotechnol. 2001, 17, 567–576. [Google Scholar] [CrossRef]
- Yoshida, T.; Tashiro, Y.; Sonomoto, K. Novel high butanol production from lactic acid and pentose by Clostridium saccharoperbutylacetonicum. J. Biosci. Bioeng. 2012, 114, 526–530. [Google Scholar] [CrossRef]
- Heyndrickx, M.; De Vos, P.; Speybrouck, A.; De Ley, J. Fermentation of mannitol by Clostridium butyricum: Role of acetate as an external hydrogen acceptor. Appl. Microbiol. Biotechnol. 1989, 31, 323–328. [Google Scholar] [CrossRef]
- Crabbendam, P.M.; Neijssel, O.M.; Tempest, D.W. Metabolic and energetic aspects of the growth of Clostridium butyricum on glucose in chemostat culture. Arch. Microbiol. 1985, 142, 375–382. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Fermentation Products (g/L) | ||||||||
|---|---|---|---|---|---|---|---|---|
| Strains and Conditions | Glucose (g/L) | Mannitol (g/L) | Glucan (g/L) | Ethanol | Acetic Acid | Lactic Acid | ||
| AK17 | 50% hydrolysate | initial | 4.92 ± 0.25 | 2.59 ± 0.12 | 5.50 ± 0.4 | ND | ND | 0.86 ± 0.1 |
| after 24 h | ND | 1.33 ± 0.09 | 4.45 ± 0.27 | 2.45 ± 0.14 | 0.89 ± 0.11 | 2.11 ± 0.15 | ||
| 100% hydrolysate | initial | 9.42 ± 0.26 | 5.18 ± 0.21 | 10.5 ± 0.55 | ND | ND | 1.50 ± 0.1 | |
| after 48 h | 1.70 ± 0.18 | 2.51 ± 0.14 | 8.5 ± 0.49 | 3.85 ± 0.24 | 1.61 ± 0.17 | 3.44 ± 0.16 | ||
| S. cerevisiae | 100% hydrolysate | initial | 9.27 ± 0.13 | 4.58 ± 0.07 | 11.11 ± 0.73 | ND | ND | 1.31 ± 0.12 |
| after 60 h | ND | 4.7 ± 0.7 | 11.65 ± 0.65 | 3.47 ± 0.12 | ND | 1.17 ± 0.15 | ||
| Strains and Conditions | Free Glucose Consumption (%) | Sugar Consumption in g/L (% of Total Sugars) | Ethanol Yield Total (% of Theoretical) | Ethanol Yield Partial (% of Theoretical) | |
|---|---|---|---|---|---|
| AK17 | 50% hydrolysate | 100 | 7.58 ± 0.5 (56) | 0.16 ± 0.01 (32) | 0.29 ± 0.01 (57) |
| 100% hydrolysate | 82 ± 2 | 12.2 ± 0.9 (49) | 0.15 ± 0.01 (30) | 0.31 ± 0.01 (61) | |
| S. cerevisiae | 100% hydrolysate | 100 | 9.3 ± 0.3 (38) | 0.14 ± 0.01 (27) | 0.37 ± 0.01 (72) |
| Strains and Conditions | Sugar Consumption in g/L (% of Total Sugars) | ABE Yield Total | ABE Yield Partial | IBE Yield Total | IBE Yield Partial | |
|---|---|---|---|---|---|---|
| C. acetobutylicum ATCC 824 | H | 26.0 ± 1.2 (87) | 0.21 ± 0.01 | 0.22 ± 0.01 | ||
| HSN | 26.2 ± 1.5 (84) | 0.21 ± 0.01 | 0.23 ± 0.01 | |||
| Control | 25.9 ± 0.9 (88) | 0.22 ± 0.01 | 0.23 ± 0.01 | |||
| C. beijerinckii NCIMB 8052 | H | 12.3 ± 0.5 (45) | 0.14 ± 0.01 | 0.32 ± 0.02 | ||
| HSN | 23.7 ± 1.1 (82) | 0.29 ± 0.02 | 0.34 ± 0.02 | |||
| Control | 25.2 ± 1.3 (87) | 0.29 ± 0.02 | 0.33 ± 0.02 | |||
| C. beijerinckii DSM6423 | H | 3.8 ± 0.4 (12) | <0.01 | 0.02 ± 0.01 | ||
| HSN | 22.8 ± 1.1 (77) | 0.23 ± 0.01 | 0.29 ± 0.02 | |||
| Control | 23.6 ± 1.1 (82) | 0.28 ± 0.02 | 0.33 ± 0.02 | |||
| C. beijerinckii DSM6422 * | H | 16.2 ± 0.8 (64) | 0.31 ± 0.02 | 0.48 ± 0.02 | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moenaert, A.; López-Contreras, A.M.; Budde, M.; Allahgholi, L.; Hou, X.; Bjerre, A.-B.; Örlygsson, J.; Karlsson, E.N.; Friðjónsson, Ó.H.; Hreggviðsson, G.Ó. Evaluation of Laminaria Digitata Hydrolysate for the Production of Bioethanol and Butanol by Fermentation. Fermentation 2023, 9, 59. https://doi.org/10.3390/fermentation9010059
Moenaert A, López-Contreras AM, Budde M, Allahgholi L, Hou X, Bjerre A-B, Örlygsson J, Karlsson EN, Friðjónsson ÓH, Hreggviðsson GÓ. Evaluation of Laminaria Digitata Hydrolysate for the Production of Bioethanol and Butanol by Fermentation. Fermentation. 2023; 9(1):59. https://doi.org/10.3390/fermentation9010059
Chicago/Turabian StyleMoenaert, Antoine, Ana M. López-Contreras, Miriam Budde, Leila Allahgholi, Xiaoru Hou, Anne-Belinda Bjerre, Johann Örlygsson, Eva Nordberg Karlsson, Ólafur H. Friðjónsson, and Guðmundur Óli Hreggviðsson. 2023. "Evaluation of Laminaria Digitata Hydrolysate for the Production of Bioethanol and Butanol by Fermentation" Fermentation 9, no. 1: 59. https://doi.org/10.3390/fermentation9010059
APA StyleMoenaert, A., López-Contreras, A. M., Budde, M., Allahgholi, L., Hou, X., Bjerre, A.-B., Örlygsson, J., Karlsson, E. N., Friðjónsson, Ó. H., & Hreggviðsson, G. Ó. (2023). Evaluation of Laminaria Digitata Hydrolysate for the Production of Bioethanol and Butanol by Fermentation. Fermentation, 9(1), 59. https://doi.org/10.3390/fermentation9010059