
Epidemiology and Genomic Characterization of Trichophyton mentagrophytes over a Period of 4 Years in Northern Italy



Abstract
1. Introduction
2. Material and Methods
2.1. Clinical Isolates
2.2. Evaluation of Terbinafine Sensitivity
2.3. Molecular Identification of ITS Region and SQLE Gene of Clinical Isolates
2.4. Genomic Characterization of Clinical Isolates
3. Results
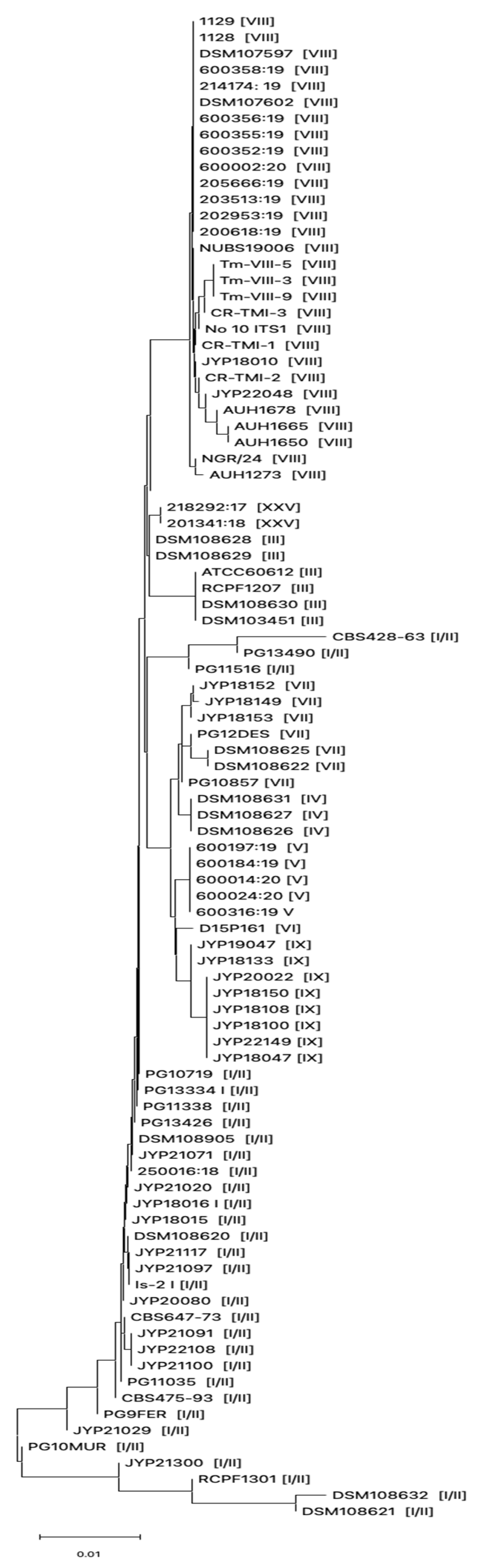
3.1. Phenotypic and Genotypic Characterization of Dermatophytes Clinical Strains
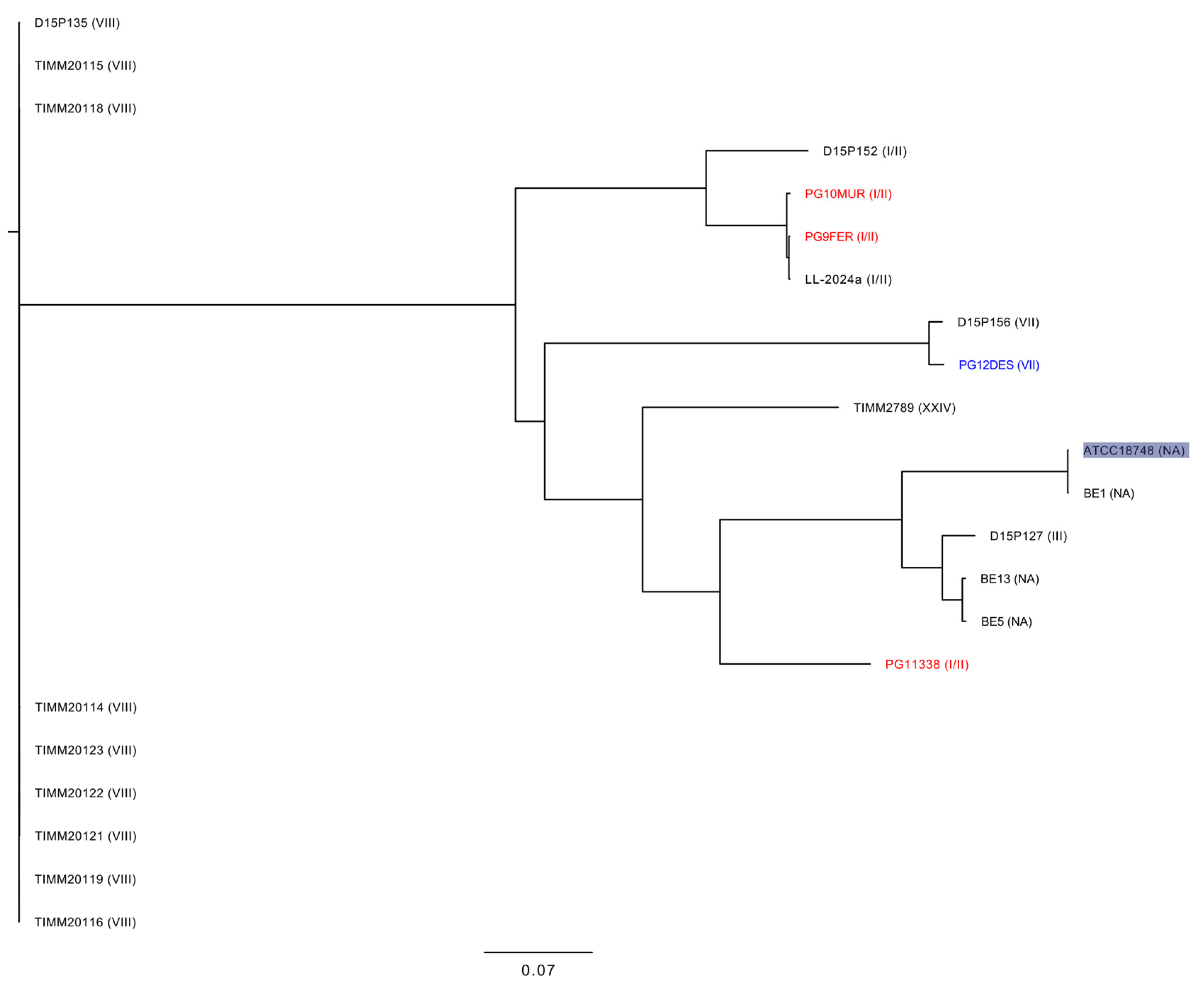
3.2. Genomic Characterization of T. mentagrophytes Genotype I/II
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lockhart, S.R.; Chowdhary, A.; Gold, J.A.W. The rapid emergence of antifungal-resistant human-pathogenic fungi. Nat. Rev. Microbiol. 2023, 21, 818–832. [Google Scholar] [CrossRef] [PubMed]
- Jabet, A.; Normand, A.C.; Brun, S.; Dannaoui, E.; Bachmeyer, C.; Piarroux, R.; Hennequin, C.; Moreno-Sabater, A. Trichophyton indotineae, from epidemiology to therapeutic. J. Mycol. Med. 2023, 33, 101383. [Google Scholar] [CrossRef] [PubMed]
- Deng, R.; Wang, X.; Li, R. Dermatophyte infection: From fungal pathogenicity to host immune responses. Front. Immunol. 2023, 14, 1285887. [Google Scholar] [CrossRef] [PubMed]
- Jia, S.; Long, X.; Hu, W.; Zhu, J.; Jiang, Y.; Ahmed, S.; de Hoog, G.S.; Liu, W.; Jiang, Y. The epidemic of the multiresistant dermatophyte Trichophyton indotineae has reached China. Front. Immunol. 2023, 13, 1113065. [Google Scholar] [CrossRef] [PubMed]
- Moreno-Sabater, A.; Normand, A.C.; Bidaud, A.L.; Cremer, G.; Foulet, F.; Brun, S.; Bonnal, C.; Aït-Ammar, N.; Jabet, A.; Ayachi, A.; et al. Terbinafine Resistance in Dermatophytes: A French Multicenter Prospective Study. J. Fungi 2022, 8, 220. [Google Scholar] [CrossRef] [PubMed]
- Havlickova, B.; Czaika, V.A.; Friedrich, M. Epidemiological trends in skin mycoses worldwide. Mycose 2008, 51, 2–15. [Google Scholar] [CrossRef] [PubMed]
- Uhrlaß, S.; Verma, S.B.; Gräser, Y.; Rezaei-Matehkolaei, A.; Hatami, M.; Schaller, M.; Nenoff, P. Trichophyton indotineae-An Emerging Pathogen Causing Recalcitrant Dermatophytoses in India and Worldwide-A Multidimensional Perspective. J. Fungi 2022, 8, 757. [Google Scholar] [CrossRef] [PubMed]
- Kano, R.; Kimura, U.; Kakurai, M.; Hiruma, J.; Kamata, H.; Suga, Y.; Harada, K. Trichophyton indotineae sp. nov.: A new highly terbinafine-resistant anthropophilic dermatophyte species. Mycopathologia 2020, 185, 947–958. [Google Scholar] [CrossRef] [PubMed]
- Sonego, B.; Corio, A.; Mazzoletti, V.; Zerbato, V.; Benini, A.; di Meo, N.; Zalaudek, I.; Stinco, G.; Errichetti, E.; Zelin, E. Trichophyton indotineae, an Emerging Drug-Resistant Dermatophyte: A Review of the Treatment Options. J. Clin. Med. 2024, 13, 3558. [Google Scholar] [CrossRef] [PubMed]
- Kruithoff, C.; Gamal, A.; McCormick, T.S.; Ghannoum, M.A. Dermatophyte Infections Worldwide: Increase in Incidence and Associated Antifungal Resistance. Life 2023, 14, 1. [Google Scholar] [CrossRef] [PubMed]
- Bidaud, A.L.; Normand, A.C.; Jabet, A.; Brun, S.; Delliere, S.; Cremer, G.; Foulet, F.; Ayachi, A.; Imbert, S.; Hennequin, C.; et al. Reliability of a terbinafine agar containing method for the screening of dermatophyte resistance. Med. Mycol. 2023, 61, myad043. [Google Scholar] [CrossRef] [PubMed]
- Taghipour, S.; Shamsizadeh, F.; Pchelin, I.M.; Rezaei-Matehhkolaei, A.; Zarei Mahmoudabadi, A.; Valadan, R.; Ansari, S.; Katiraee, F.; Pakshir, K.; Zomorodian, K.; et al. Emergence of Terbinafine Resistant Trichophyton mentagrophytes in Iran, Harboring Mutations in the Squalene Epoxidase (SQLE) Gene. Infect. Drug Resist. 2020, 13, 845–850. [Google Scholar] [CrossRef] [PubMed]
- Amadesi, S.; Amedeo, A.; Rinaldi, M.; Palombo, M.; Giannella, M.; Gaibani, P. In vivo emergence of cefiderocol and ceftazidime/avibactam cross-resistance in KPC- producing Klebsiella pneumoniae following ceftazidime/avibactam -based therapies. Diagn. Microbiol. Infect. Dis. 2024, 110, 116372. [Google Scholar] [CrossRef] [PubMed]
- Marangoni, A.; Amadesi, S.; Djusse, M.E.; Foschi, C.; Gaspari, V.; Lazzarotto, T.; Gaibani, P. Whole Genome Sequencing of a Chlamydia trachomatis Strain Responsible for a Case of Rectal Lymphogranuloma Venereum in Italy. Curr. Issues Mol. Biol. 2023, 45, 1852–1859. [Google Scholar] [CrossRef] [PubMed]
- Taghipour, S.; Pchelin, I.M.; Zarei Mahmoudabadi, A.; Ansari, S.; Katiraee, F.; Rafiei, A.; Shokohi, T.; Abastabar, M.; Taraskina, A.E.; Kermani, F.; et al. Trichophyton mentagrophytes and T interdigitale genotypes are associated with particular geographic areas and clinical manifestations. Mycoses 2019, 62, 1084–1091. [Google Scholar] [CrossRef] [PubMed]
- Kapranou, R.; Kotsafti, O.; Vrioni, G.; Giannoukos, A.; Papanikou, S.; Stratigos, A.; Nicolaidou, E. Trichophyton Mentagrophytes type VII (TMVII): An emerging sexually transmitted pathogen. QJM 2025, hcaf049. [Google Scholar] [CrossRef] [PubMed]
- Jabet, A.; Bérot, V.; Chiarabini, T.; Dellière, S.; Bosshard, P.P.; Siguier, M.; Tubiana, R.; Favier, M.; Canestri, A.; Makhloufi, S.; et al. Trichophyton mentagrophytes ITS genotype VII infections among men who have sex with men in France: An ongoing phenomenon. J. Eur. Acad. Dermatol. Venereol. 2025, 39, 407–415. [Google Scholar] [CrossRef] [PubMed]
- Kupsch, C.; Czaika, V.A.; Deutsch, C.; Gräser, Y. Trichophyton mentagrophytes—A new genotype of zoophilic dermatophyte causes sexually transmitted infections. J. Dtsch. Dermatol. Ges. 2019, 17, 493–501. [Google Scholar] [CrossRef] [PubMed]
- Caplan, A.S.; Sikora, M.; Strome, A.; Akoh, C.C.; Otto, C.; Chaturvedi, S.; Zampella, J.G. Potential Sexual Transmission of Tinea Pubogenitalis From TMVII. JAMA Dermatol. 2024, 160, 783–785. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
| Strain | Country | Host | Genotype | Acc. No. |
|---|---|---|---|---|
| JYP22149 | China | Human | IX | OP961423 |
| JYP18047 | China | Human | IX | OP961397 |
| JYP18108 | China | Human | IX | OP961399 |
| JYP18150 | China | Human | IX | OP961402 |
| JYP18100 | China | Human | IX | OP961398 |
| JYP20022 | China | Human | IX | OP961406 |
| JYP18133 | China | Human | IX | OP961400 |
| JYP19047 | China | Human | IX | OP961405 |
| D15P161 | Russia | Cat | VI | MK722518 |
| DSM 108631 | Germany | Human | IV | MK447607 |
| DSM 108627 | Germany | Human | IV | MK447608 |
| DSM 108626 | Germany | Human | IV | MK447609 |
| DSM 108625 | Germany | Human | VII | MK450323 |
| DSM 108622 | Germany | Human | VII | MK450322 |
| JYP18149 | China | Human | VII | OP961401 |
| JYP18153 | China | Human | VII | OP961404 |
| 600316:9 | Iraq | Human | V | MT374257 |
| 600024:20 | Iraq | Human | V | MT374269 |
| 600197:19 | Iraq | Human | V | MT374258 |
| 600184:19 | Iran | Human | V | MT374259 |
| ATCC60612 | N.A. | N.A. | III | KJ606099 |
| RCPF 1207 | Russia | Human | III | KT253559 |
| DSM 108630 | Swiss | Human | III | MK450325 |
| DSM 103451 | Swiss | Leopard | III | KX866689 |
| DSM 108632 | Germany | Human | I/II | MK447606 |
| 218292:17 | Cambodia | Human | XXV | MN886815 |
| 201341:18 | Cambodia | Human | XXV | MN886816 |
| DSM 108628 | Germany | Human | III | MK447605 |
| DSM 108629 | Germany | Human | III | MK447604 |
| CR-TMI-1 | France | Human | VIII | MW898018 |
| Tm-VIII-5 | France | Human | VIII | MW959757 |
| Tm-VIII-3 | France | Human | VIII | MW959755 |
| Tm-VIII-9 | France | Human | VIII | MW959756 |
| CR-TMI-3 | France | Human | VIII | MW898020 |
| No_10_ITS1 | Vietnam | Human | VIII | OM108103 |
| AUH1273 | Greece | Human | VIII | MW752105 |
| JYP22048 | India | Human | VIII | OP961421 |
| JYP18010 | India | Human | VIII | OP961393 |
| CR-TMI-2 | France | Human | VIII | MW898019 |
| AUH1678 | Greece | Human | VIII | MW752108 |
| AUH1665 | Greece | Human | VIII | MW752107 |
| AUH1650 | Greece | Human | VIII | MW752111 |
| NUBS19006 | Japan | Human | VIII | LC508024 |
| 200618:19 | Germany | Human | VIII | MT330285 |
| 202953:19 | Germany | Human | VIII | MT330284 |
| 203513:19 | Germany | Human | VIII | MT330280 |
| 600002:20 | Germany | Human | VIII | MT333227 |
| 600352:19 | Poland | Human | VIII | OM951136 |
| 600355:19 | Poland | Human | VIII | OM951134 |
| 600356:19 | Poland | Human | VIII | OM951138 |
| 214174:19 | Germany | Human | VIII | MT330289 |
| 600358:19 | Poland | Human | VIII | OM951141 |
| DSM 107597 | India | Human | VIII | MH791420 |
| DSM 107602 | India | Human | VIII | MH791425 |
| 1129 | France | Human | VIII | ON528187 |
| 1128 | France | Human | VIII | ON528186 |
| JYP20080 | China | Human | I/II | OP961407 |
| 250016:18 | Cambodia | Human | II | QKE51044 |
| DSM 108905 | Germany | Human | II | QFR36155 |
| JYP21071 | China | Human | I/II | OP961415 |
| JYP21117 | China | Human | I/II | OP961419 |
| JYP21097 | China | Human | I/II | OP961417 |
| Is-2 | Tunisia | N.A. | I | KC595991 |
| RCPF 1301 | Russia | Human | I/II | AKH90742 |
| DSM 108621 | Germany | Human | I/II | QBA85641 |
| JYP18016 | China | Human | I/II | OP961395 |
| DSM 108620 | Germany | Human | I | QBA85640 |
| JYP21020 | China | Human | I/II | OP961411 |
| JYP22108 | China | Human | I/II | OP961422 |
| JYP21100 | China | Human | I/II | OP961418 |
| JYP21091 | China | Human | I/II | OP961416 |
| JYP18015 | China | Human | I/II | OP961394 |
| JYP21029 | China | Human | I/II | OP961412 |
| JYP21300 | China | Human | I/II | OP961420 |
| CBS475.93 | N.A. | N.A. | I/II | MF926357 |
| CBS647.73 | Germany | Human | I/II | KT155955 |
| CBS428.63 | N.A. | N.A. | I/II | NR_144900 |
| Strain | Acc. No. | Country | Genotype |
|---|---|---|---|
| ATCC18748 | GCA_045862495.1 | Reference | NA |
| D15P127 | GCA_003664465.1 | Russia | III |
| D15P135 | GCA_003664455.1 | India | VIII |
| D15P152 | GCA_003664425.1 | Russia | I/II |
| D15P156 | GCA_003664385.1 | Moldova | VI |
| TIMM2789 | GCA_003118255.1 | Japan | XXIV |
| TIMM20114 | GCA_023065905.1 | India | VIII |
| TIMM20115 | GCA_023065845.1 | India | VIII |
| TIMM20116 | GCA_023065885.1 | India | VIII |
| TIMM20118 | GCA_023065865.1 | India | VIII |
| TIMM20119 | GCA_023065815.1 | India | VIII |
| TIMM20121 | JAUJAB010000000 | India | VIII |
| TIMM20122 | GCA_032157395.1 | India | VIII |
| TIMM20123 | GCA_032157405.1 | India | VIII |
| LL-2024a | SAMN44006357 | China | I/II |
| BE-1 | SAMN43365157 | Belgium | NA |
| BE-5 | SAMN43365161 | Belgium | NA |
| BE-13 | SAMN43365169 | Belgium | NA |
| PG12DES | JBKIZU000000000 | Italy | VII |
| Strain | Species | Clinical Sample | Source | Year of Isolation | SQLE Gene | Genotype |
|---|---|---|---|---|---|---|
| ARTRO | T. verrucosum | NA | NA | 2020 | NA | NA |
| PG10719 | T. mentagrophytes | Nail scales | Toenails | 2020 | WT | I/II |
| PG11516 | T. mentagrophytes | Nail scales | Big toe | 2020 | WT | I/II |
| PG10565 | Beauveria bassiana | Beard hair | Beard | 2020 | NA | NA |
| PG10857 | T. mentagrophytes | Skin scales | Face | 2020 | WT | VII |
| PG10MUR | T. mentagrophytes | Nail scales | Toenails | 2021 | WT | I/II |
| PG11338 | T. mentagrophytes | NA | Face | 2021 | Lys276Asn | I/II |
| PG11035 | T. mentagrophytes | Nail scales | Toenails | 2022 | WT | I/II |
| PG12DES | T. mentagrophytes | Skin scales | Periocular | 2022 | WT | VII |
| PG13426 | T. mentagrophytes | Skin scales | NA | 2023 | Lys276Asn | I/II |
| PG9FER | T. mentagrophytes | Nail scales | Big toe | 2023 | WT | I/II |
| PG13490 | T. mentagrophytes | Nail scales | Big toe | 2024 | WT | I/II |
| PG13334 | T. mentagrophytes | Nail scales | Thumb | 2024 | WT | I/II |
| Genome Characteristics | PG9FER | PG10MUR | PG11338 | PG12DES |
|---|---|---|---|---|
| Total Length (bp) | 22,823,954 | 22,793,562 | 23,205,410 | 23,117,013 |
| No. of Contigs | 615 | 679 | 712 | 306 |
| N50 | 91.543 | 91.906 | 100.681 | 160.843 |
| N90 | 24.082 | 22.100 | 24.509 | 41.955 |
| L50 | 79 | 79 | 71 | 47 |
| L90 | 257 | 275 | 241 | 156 |
| G + C (%) | 47.98 | 48 | 47.52 | 47.71 |
| No. of predicted genes | 7.124 | 7.140 | 7.140 | 7.187 |
| Virulence Factors | PG9FER | PG10MUR | PG11338 | PG12DES |
|---|---|---|---|---|
| MEP-1 | S62A | G153D | S62A | S62A, G153D |
| MEP-2 | Y334F | Y334F | Y334F | Y334F, N517S |
| MEP-3 | D122G | WT | D122G | I573L, R615K, P620A |
| MEP-4 | NC | NC | NC | WT |
| MEP-5 | N164S, WPI252-254RAL, Del255S, T265S, I510T | H39R, N164S, WPI252-254RAL, Del255S, T265S | N164S, WPI252-254RAL, Del255S, T265S, I510T | NC |
| ZafA | F111S, Del366-401, D402Y, Ins484N, Del485-505 | Del366-401, D402Y, Ins484T | F111S, Del366-401, D402Y, Ins484T | Ins484N |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rossi, L.; Sorrentino, A.; Signoretto, C.; Gaibani, P. Epidemiology and Genomic Characterization of Trichophyton mentagrophytes over a Period of 4 Years in Northern Italy. J. Fungi 2025, 11, 566. https://doi.org/10.3390/jof11080566
Rossi L, Sorrentino A, Signoretto C, Gaibani P. Epidemiology and Genomic Characterization of Trichophyton mentagrophytes over a Period of 4 Years in Northern Italy. Journal of Fungi. 2025; 11(8):566. https://doi.org/10.3390/jof11080566
Chicago/Turabian StyleRossi, Luca, Annarita Sorrentino, Caterina Signoretto, and Paolo Gaibani. 2025. "Epidemiology and Genomic Characterization of Trichophyton mentagrophytes over a Period of 4 Years in Northern Italy" Journal of Fungi 11, no. 8: 566. https://doi.org/10.3390/jof11080566
APA StyleRossi, L., Sorrentino, A., Signoretto, C., & Gaibani, P. (2025). Epidemiology and Genomic Characterization of Trichophyton mentagrophytes over a Period of 4 Years in Northern Italy. Journal of Fungi, 11(8), 566. https://doi.org/10.3390/jof11080566