
Morphological and Phylogenetic Characterisations Reveal Four New Species in Leptosphaeriaceae (Pleosporales, Dothideomycetes)

 ,
,  , ,
, ,  and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Sample Collection, Isolation, and Identification
2.2. DNA Extraction, PCR Amplification, and DNA Sequencing
2.3. Phylogenetic Analyses
3. Results
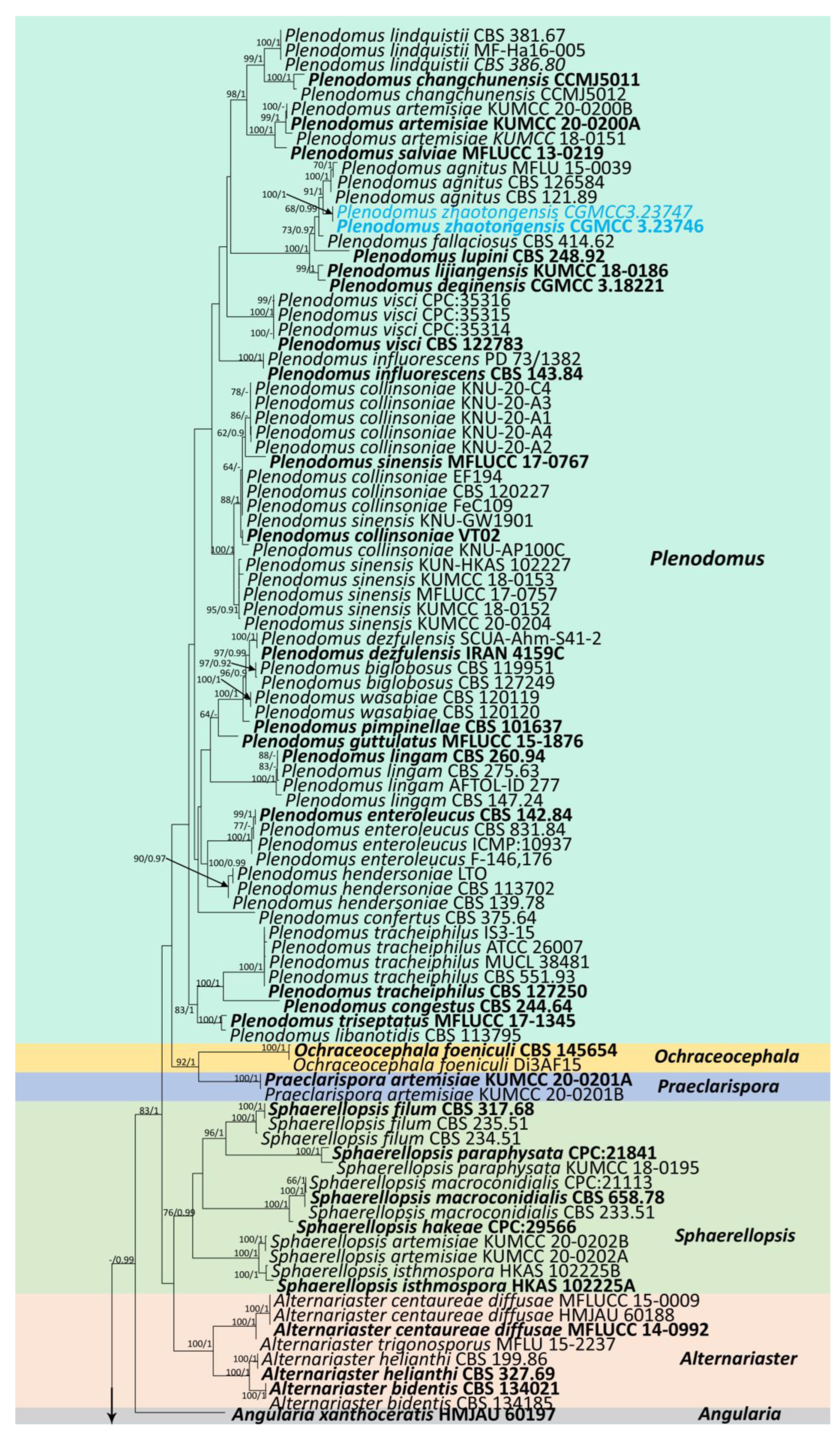
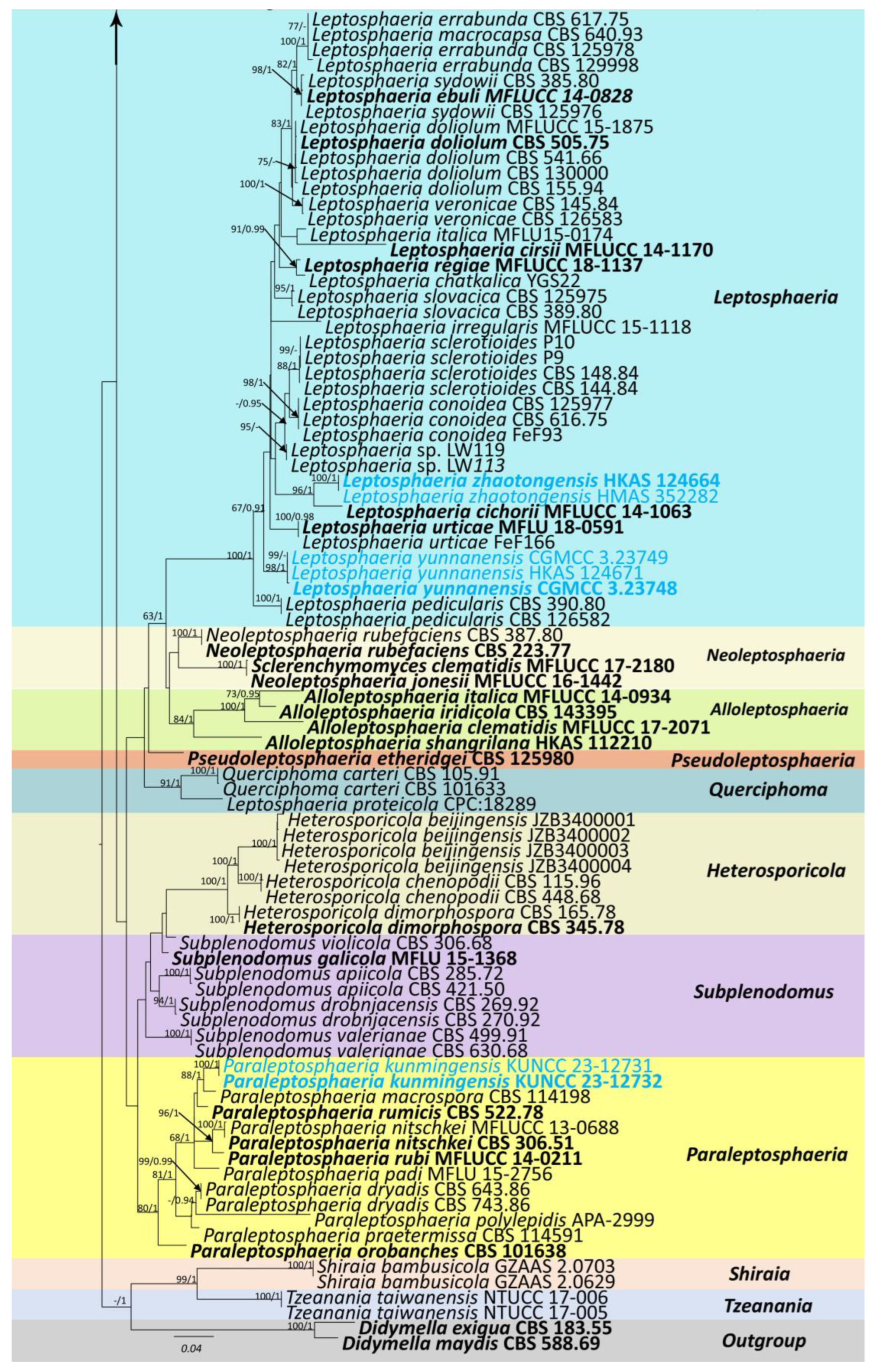
3.1. Phylogenetic Analyses
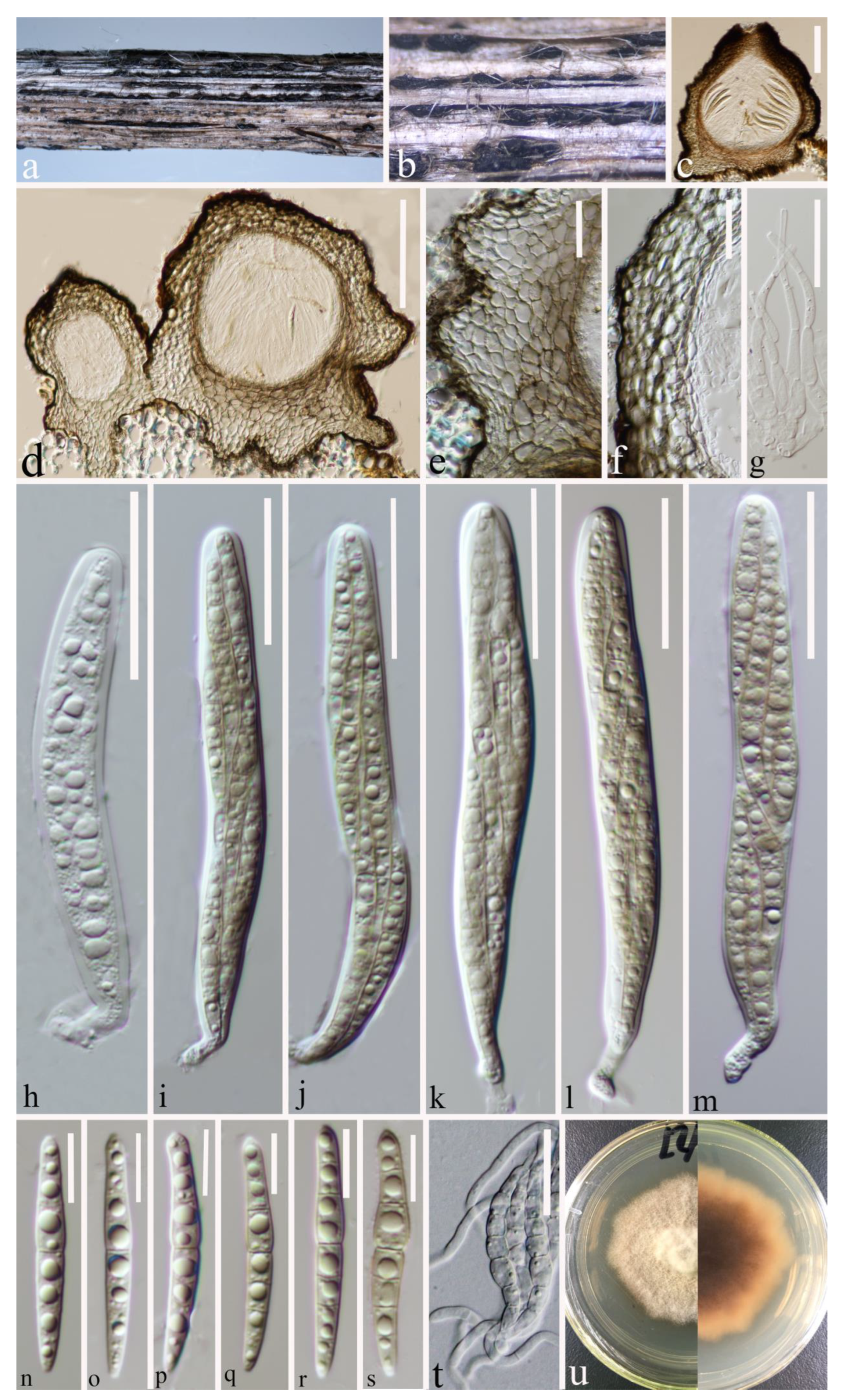
3.2. Taxonomy
3.2.1. Leptosphaeria yunnanensis Y. Gao and H. Gui, sp. nov.
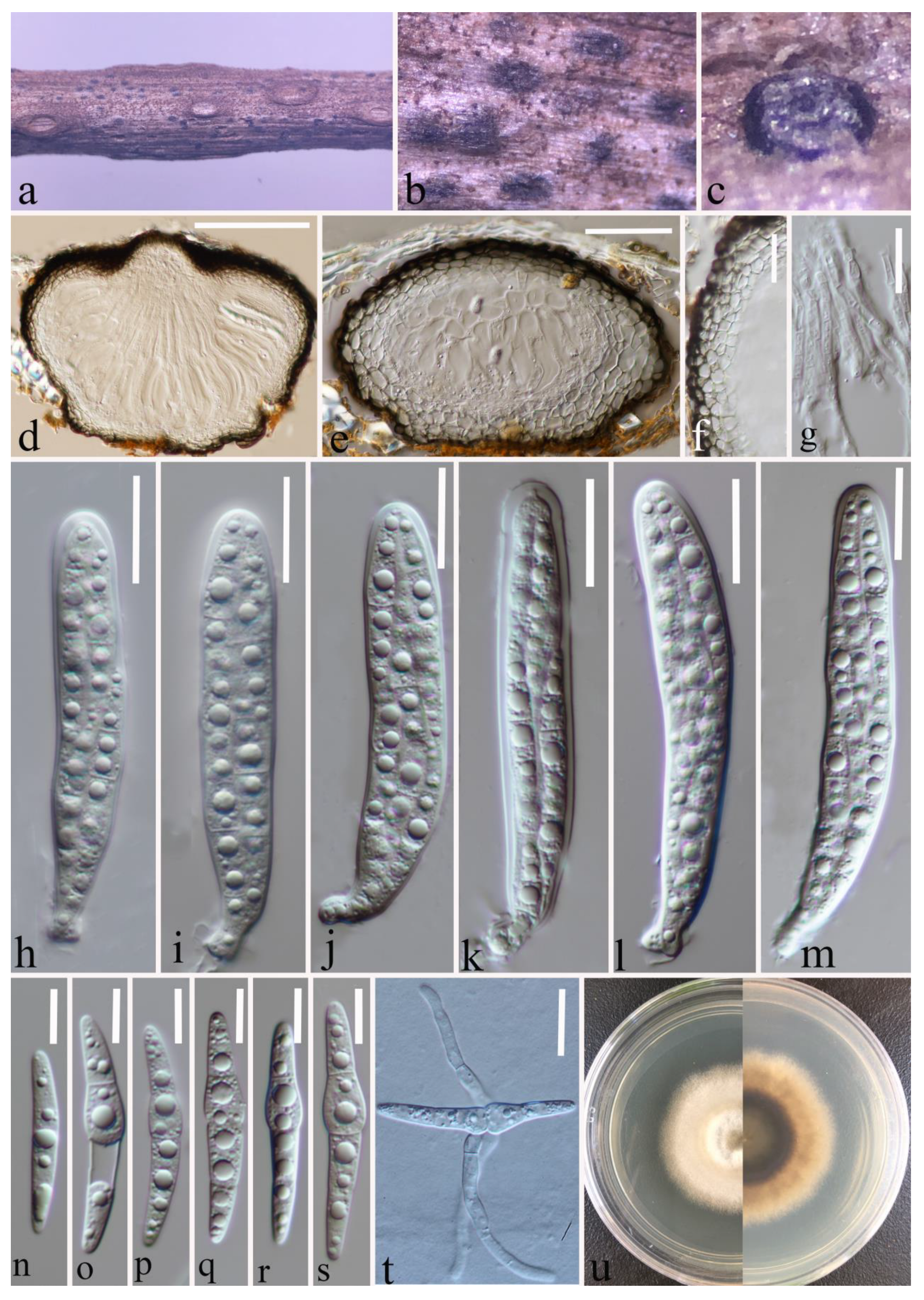
3.2.2. Leptosphaeria zhaotongensis Y. Gao and H. Gui, sp. nov.
3.2.3. Paraleptosphaeria kunmingensis Y. Gao and H. Gui, sp. nov.
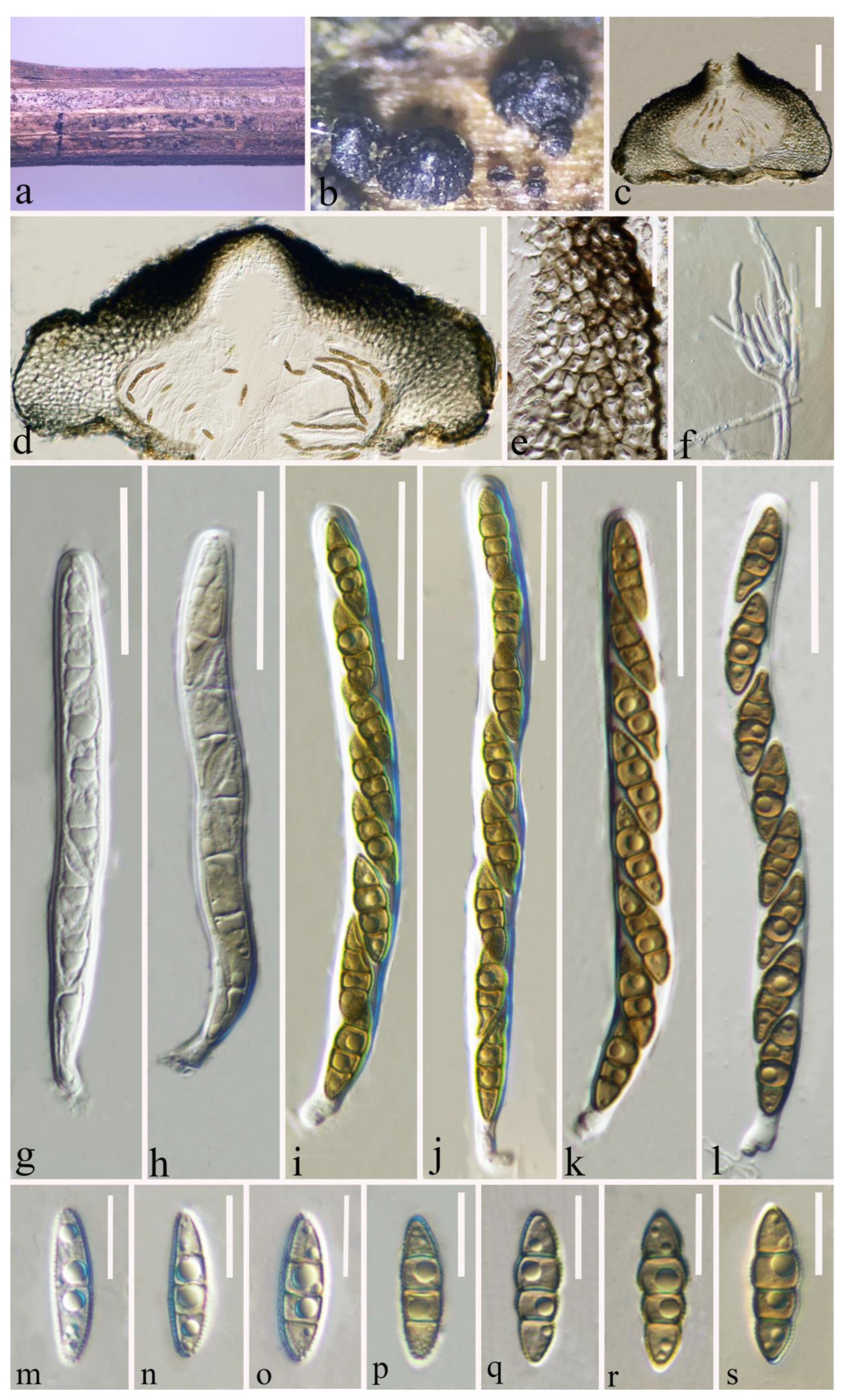
3.2.4. Plenodomus zhaotongensis Y. Gao and H. Gui, sp. nov.
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Barr, M.E. New taxa and combinations in the Loculoascomycetes. Mycotaxon 1987, 29, 501–505. [Google Scholar]
- Ariyawansa, H.A.; Phukhamsakda, C.; Thambugala, K.M.; Bulgakov, T.S.; Wanasinghe, D.N.; Perera, R.H.; Mapook, A.; Camporesi, E.; Kang, J.C.; Gareth Jones, E.B.G.; et al. Revision and phylogeny of Leptosphaeriaceae. Fungal Divers. 2015, 74, 19–51. [Google Scholar] [CrossRef]
- Phookamsak, R.; Hyde, K.D.; Jeewon, R.; Bhat, D.J.; Jones, E.B.G.; Maharachchikumbura, S.S.N.; Raspé, O.; Karunarathna, S.C.; Wanasinghe, D.N.; Hongsanan, S.; et al. Fungal diversity notes 929–1035: Taxonomic and phylogenetic contributions on genera and species of fungi. Fungal Divers. 2019, 95, 1–273. [Google Scholar] [CrossRef]
- Sun, J.Z.; Liu, X.Z.; McKenzie, E.H.C.; Jeewon, R.; Liu, J.K.; Zhang, X.L.; Zhao, Q.; Hyde, K.D. Fungicolous fungi: Terminology, diversity, distribution, evolution and species checklist. Fungal Divers. 2019, 95, 1–94. [Google Scholar] [CrossRef]
- Lestari, A.S.; Wanasinghe, D.N.; Gafforov, Y.; Tennakoon, D.S.; Chethana, K.W.T.; Aburazakov, A.; Promputtha, I.; Hyde, K.D. Taxonomy and phylogenetic appraisal of Leptosphaeria chatkalica sp. nov. (Leptosphaeriaceae, Pleosporales) from Uzbekistan. Phytotaxa 2021, 520, 155–168. [Google Scholar] [CrossRef]
- Xu, R.; Su, W.X.; Tian, S.Q.; Bhunjun, C.S.; Tibpromma, S.; Hyde, K.D.; Li, Y.; Phukhamsakda, C. Synopsis of Leptosphaeriaceae and introduction of three new taxa and one new record from China. J. Fungi 2022, 8, 416. [Google Scholar] [CrossRef]
- Howlett, B.J.; Idnurm, A.; Soledade, M.; Pedras, C. Leptosphaeria maculans, the causal agent of blackleg disease of Brassicas. Fungal Genet. Biol. 2001, 33, 1–14. [Google Scholar] [CrossRef]
- Zhang, X.; White, R.P.; Demir, E.; Jedryczka, M.; Lange, R.M.; Islam, M.; Li, Z.Q.; Huang, Y.J.; Hall, A.M.; Zhou, G.; et al. Leptosphaeria spp., phoma stem canker and potential spread of L. maculans on oilseed rape crops in China. Plant Pathol. 2014, 63, 598–612. [Google Scholar] [CrossRef]
- Wanasinghe, D.N.; Camporesi, E.; Hu, D.M. Neoleptosphaeria jonesii sp. nov., a novel saprobic sexual species, in Leptosphaeriaceae. Mycosphere 2016, 7, 1368–1377. [Google Scholar] [CrossRef]
- Hyde, K.D.; De Silva, N.; Jeewon, R.; Bhat, D.J.; Phookamsak, R.; Doilom, M.; Boonmee, S.; Jayawardena, R.S.; Maharachchikumbura, S.S.N.; Senanayake, I.C.; et al. AJOM new records and collections of fungi: 1–100. Asian J. Mycol. 2020, 3, 22–294. [Google Scholar] [CrossRef]
- Pem, D.; Hongsanan, S.; Doilom, M.; Tibpromma, S.; Wanasinghe, D.N.; Dong, W.; Liu, N.G.; Phookamsak, R.; Phillips, A.J.L.; Jeewon, R.; et al. https://www.dothideomycetes.org: An online taxonomic resource for the classification, identification, and nomenclature of Dothideomycetes. Asian J. Mycol. 2020, 3, 287–297. [Google Scholar] [CrossRef]
- Hyde, K.D.; Jones, E.B.G.; Liu, J.K.; Ariyawansa, H.; Boehm, E.; Boonmee, S.; Braun, U.; Chomnunti, P.; Crous, P.W.; Dai, D.Q.; et al. Families of Dothideomycetes. Fungal Divers. 2013, 63, 1–313. [Google Scholar] [CrossRef]
- Gruyter, J.de.; Woudenberg, J.H.C.; Aveskamp, M.M.; Verkley, G.J.M.; Groenewald, J.Z.; Crous, P.W. Redisposition of Phoma-like anamorphs in Pleosporales. Stud. Mycol. 2013, 75, 1–36. [Google Scholar] [CrossRef] [PubMed]
- Phukhamsakda, C.; McKenzie, E.H.C.; Phillips, A.J.L.; Gareth Jones, E.B.; Jayarama Bhat, D.; Stadler, M.; Bhunjun, C.S.; Wanasinghe, D.N.; Thongbai, B.; Camporesi, E.; et al. Microfungi associated with Clematis (Ranunculaceae) with an integrated approach to delimiting species boundaries. Fungal Divers. 2020, 102, 1–203. [Google Scholar] [CrossRef]
- Thiyagaraja, V.; Wanasinghe, D.N.; Karunarathna, S.C.; Tennakoon, D.S.; Hyde, K.D.; To-Anun, C.; Cheewangkoon, R. Alloleptosphaeria shangrilana sp. nov. and first report of the genus (Leptosphaeriaceae, Dothideomycetes) from China. Phytotaxa 2021, 491, 12–22. [Google Scholar] [CrossRef]
- Alves, J.L.; Woudenberg, J.H.C.; Duarte, L.L.; Crous, P.W.; Barreto, R.W. Reappraisal of the genus Alternariaster (Dothideomycetes). Persoonia 2013, 31, 77–85. [Google Scholar] [CrossRef]
- Aiello, D.; Vitale, A.; Polizzi, G.; Voglmayr, H. Ochraceocephala foeniculi gen. et sp. nov., a new pathogen causing crown rot of fennel in Italy. MycoKeys 2020, 66, 1–22. [Google Scholar] [CrossRef]
- Liu, J.K.; Hyde, K.D.; Jones, E.B.G.; Ariyawansa, H.A.; Bhat, D.J.; Boonmee, S.; Maharachchikumbura, S.S.N.; McKenzie, E.H.C.; Phookamsak, R.; Phukhamsakda, C.; et al. Fungal Diversity Notes 1-100: Taxonomic and phylogenetic contributions to fungal species. Fungal Divers. 2015, 72, 1–197. [Google Scholar] [CrossRef]
- Tibpromma, S.; Hyde, K.D.; Jeewon, R.; Maharachchikumbura, S.S.N.; Liu, J.K.; Bhat, D.J.; Jones, E.B.G.; McKenzie, E.H.C.; Camporesi, E.; Bulgakov, T.S. Fungal diversity notes 491–602: Taxonomic and phylogenetic contributions to fungal taxa. Fungal Divers. 2017, 83, 1–261. [Google Scholar] [CrossRef]
- Piątek, M.; Rodriguez-Flakus, P.; Domic, A.; Palabral-Aguilera, A.N.; Gómez, M.I.; Flakus, A. Phylogenetic placement of Leptosphaeria polylepidis, a pathogen of Andean endemic Polylepis tarapacana, and its newly discovered mycoparasite Sajamaea mycophila gen. et sp. nov. Mycol. Prog. 2020, 19, 1–14. [Google Scholar] [CrossRef]
- Hongsanan, S.; Hyde, K.D.; Phookamsak, R.; Wanasinghe, D.N.; Mckenzie, E.H.C.; Sarma, V.V.; Boonmee, S.; Lücking, R.; Bhat, D.J.; Liu, N.G.; et al. Refined families of Dothideomycetes: Dothideomycetidae and Pleosporomycetidae. Mycosphere 2020, 11, 1553–2107. [Google Scholar] [CrossRef]
- Wijayawardene, N.N.; Hyde, K.D.; Dai, D.Q.; Sánchez-García, M.; Goto, B.T.; Saxena, R.K.; Erdoğdu, M.; Selçuk, F.; Rajeshkumar, K.C.; Aptroot, A.; et al. Outline of Fungi and fungus-like taxa—2021. Mycosphere 2022, 13, 53–453. [Google Scholar] [CrossRef]
- Doilom, M.; Hyde, K.D.; Dong, W.; Liao, C.F.; Suwannarach, N.; Lumyong, S. The plant family Asteraceae is a cache for novel fungal diversity: Novel species and genera with remarkable ascospores in Leptosphaeriaceae. Front. Microbiol. 2021, 12, 660261. [Google Scholar] [CrossRef] [PubMed]
- Index Fungorum. 2023. Available online: http://www.indexfungorum.org/Names/Names.asp (accessed on 29 March 2023).
- Marin-Felix, Y.; Groenewald, J.Z.; Cai, L.; Chen, Q.; Marincowitz, S.; Barnes, I.; Bensch, K.; Braun, U.; Camporesi, E.; Damm, U.; et al. Genera of phytopathogenic fungi: GOPHY 1. Stud. Mycol. 2017, 86, 99–216. [Google Scholar] [CrossRef] [PubMed]
- Zhao, P.; Crous, P.W.; Hou, L.W.; Duan, W.J.; Cai, L.; Ma, Z.Y.; Liu, F. Fungi of quarantine concern for China I: Dothideomycetes. Pers. Mol. Phylogeny Evol. Fungi 2021, 47, 45–105. [Google Scholar] [CrossRef]
- Risser, P.G. Diversity in and Among Grasslands. In Biodiversity; Wilson, E., Peter, F., Eds.; National Academics Press: Washington, DC, USA, 1988; pp. 176–180. [Google Scholar]
- Karunarathna, A.; Tibpromma, S.; Jayawardena, R.S.; Nanayakkara, C.; Asad, S.; Xu, J.-C.; Hyde, K.D.; Karunarathna, S.C.; Stephenson, S.L.; Lumyong, S.; et al. Fungal pathogens in Grasslands. Front. Cell. Infect. Microbiol 2021, 11, 695087. [Google Scholar] [CrossRef]
- Karunarathna, A.; Withee, P.; Pakdeeniti, P.; Haituk, S.; Tanakaew, N.; Senwanna, C.; Działak, P.; Karunarathna, S.C.; Tibpromma, S.; Promthep, T.; et al. Worldwide checklist on Grass Fungi: What do we know so far in Ascomycota. Chiang Mai J. Sci. 2022, 49, 742–984. [Google Scholar] [CrossRef]
- Thambugala, K.M.; Wanasinghe, D.N.; Phillips, A.J.L.; Camporesi, E.; Bulgakov, T.S.; Phukhamsakda, C.; Ariyawansa, H.A.; Goonasekara, I.D.; Phookamsak, R.; Dissanayake, A.; et al. Mycosphere notes 1-50: Grass (Poaceae) inhabiting Dothideomycetes. Mycosphere 2017, 8, 697–796. [Google Scholar] [CrossRef]
- Goonasekara, I.D.; Jayawardene, R.S.; Saichana, N.; Hyde, K.D. Checklist of microfungi on grasses in Thailand (excluding bambusicolous fungi). Asian J. Mycol. 2018, 1, 88–105. [Google Scholar] [CrossRef]
- Brahmanage, R.S.; Dayarathne, M.C.; Wanasinghe, D.N.; Thambugala, K.M.; Jeewon, R.; Chethana, K.W.T.; Samarakoon, M.C.; Tennakoon, D.S.; De Silva, N.I.; Camporesi, E.; et al. Taxonomic novelties of saprobic Pleosporales from selected dicotyledons and grasses. Mycosphere 2020, 11, 2481–2541. [Google Scholar] [CrossRef]
- Senanayake, I.C.; Rathnayake, A.R.; Marasinghe, D.S.; Calabon, M.S.; Gentekaki, E.; Lee, H.B. Morphological approaches in studying fungi: Collection, examination, isolation, sporulation and preservation. Mycosphere 2020, 11, 2678–2754. [Google Scholar] [CrossRef]
- Jayasiri, S.C.; Hyde, K.D.; Ariyawansa, H.A.; Bhat, J.; Buyck, B.; Cai, L.; Dai, Y.C.; Abd-Elsalam, K.A.; Ertz, D.; Hidayat, I.; et al. The Faces of Fungi database: Fungal names linked with morphology, phylogeny and human impacts. Fungal Divers. 2015, 74, 3–18. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. 38 - Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: San Diego, CA, USA, 1990; pp. 315–322. [Google Scholar] [CrossRef]
- Vilgalys, R.; Hester, M. Rapid genetic identification and mapping of enzymatically amplified ribosomal DNA from several Cryptococcus species. J. Bacteriol. 1990, 172, 4238–4246. [Google Scholar] [CrossRef] [PubMed]
- O’Donnell, K.; Cigelnik, E. Two divergent intragenomic rDNA ITS2 types within a monophyletic lineage of the fungusfusariumare nonorthologous. Mol. Phylogenet. Evol. 1997, 7, 103–116. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.J.; Whelen, S.; Hall, B.D. Phylogenetic relationships amongascomycetes evidence from an RNA polymerse II subunit. Mol. Biol. Evol 1999, 16, 1799–1808. [Google Scholar] [CrossRef]
- Crous, P.W.; Wingfield, M.J.; Schumacher, R.K.; Akulov, A.; Bulgakov, T.S.; Carnegie, A.J.; Jurjević, Ž.; Decock, C.; Denman, S.; Lombard, L.; et al. New and Interesting Fungi. 3. Fungal Syst. Evol. 2020, 6, 157–231. [Google Scholar] [CrossRef]
- Hyde, K.D.; Dong, Y.; Phookamsak, R.; Jeewon, R.; Bhat, D.J.; Jones, E.B.G.; Liu, N.G.; Abeywickrama, P.D.; Mapook, A.; Wei, D.P.; et al. Fungal diversity notes 1151–1276: Taxonomic and phylogenetic contributions on genera and species of fungal taxa. Fungal Divers. 2020, 100, 5–277. [Google Scholar] [CrossRef]
- Pem, D.; Jeewon, R.; Selcuk, F.; Ulukapi, M.; Bhat, J.; Doilom, M.; Lumyong, S.; Hyde, K.D. Ribosomal and protein gene phylogeny reveals novel saprobic fungal species from Juglans regia and Urtica dioica. Front. Microbiol. 2020, 11, 1303. [Google Scholar] [CrossRef]
- Katoh, K.; Rozewicki, J.; Yamada, K.D. MAFFT online service: Multiple sequence alignment, interactive sequence choice and visualization. Brief. Bioinform. 2019, 20, 1160–1166. [Google Scholar] [CrossRef]
- Capella-Gutiérrez, S.; Silla-Martínez, J.M.; Gabaldón, T. trimAl: A tool for automated alignment trimming in large-scale phylogenetic analyses. Bioinformatics 2009, 25, 1972–1973. [Google Scholar] [CrossRef]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef] [PubMed]
- Miller, M.A.; Pfeiffer, W.; Schwartz, T. Creating the CIPRES Science Gateway for inference of large phylogenetic trees. In Proceedings of the 2010 Gateway Computing Environments Workshop (GCE), New Orleans, LA, USA, 14 November 2010; pp. 1–8. [Google Scholar] [CrossRef]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [PubMed]
- Nylander, J.A.A.; Wilgenbusch, J.C.; Warren, D.L.; Swofford, D.L. AWTY: A system for graphical exploration of MCMC convergence in Bayesian phylogenetics. Bioinformatics 2008, 24, 581–583. [Google Scholar] [CrossRef]
- Ronquist, F.; Huelsenbeck, J.P. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef]
- Rambaut, A. FigTree Version 1.4.0. 2012. Available online: http://tree.bio.ed.ac.uk/software/figtree/ (accessed on 18 February 2023).
- Chethana, K.T.; Manawasinghe, I.S.; Hurdeal, V.G.; Bhunjun, C.S.; Appadoo, M.A.; Gentekaki, E.; Raspé, O.; Promputtha, I.; Hyde, K.D. What are fungal species and how to delineate them? Fungal Divers. 2021, 109, 1–25. [Google Scholar] [CrossRef]
- Maharachchikumbura, S.S.; Chen, Y.; Ariyawansa, H.A.; Hyde, K.D.; Haelewaters, D.; Perera, R.H.; Samarakoon, M.C.; Wanasinghe, D.N.; Bustamante, D.E.; Liu, J.K.; et al. Integrative approaches for species delimitation in Ascomycota. Fungal Divers. 2021, 109, 155–179. [Google Scholar] [CrossRef]
- Niranjan, M.; Sarma, V.V. Twelve new species of ascomycetous from Andaman Islands, India. Kavaka 2018, 50, 84–97. [Google Scholar]
- Saccardo, P.A. Sylloge Pyrenomycetum, In Sylloge Fungorum; Sumptibus Auctoris: Padua, Italy, 1883; Volume 2, pp. 1–813. [Google Scholar]
- Boerema, G.H.; de Gruyter, J.; van Kesteren, H.A. Contributions towards a monograph of Phoma (Coelomycetes)–III-1. Section Plenodomus: Taxa often with a Leptosphaeria teleomorph. Pers. Mol. Phylogeny Evol. Fungi 1994, 15, 431–487. [Google Scholar]
- Jayawardena, R.S.; Hyde, K.D.; de Farias, A.R.; Bhunjun, C.S.; Ferdinandez, H.S.; Manamgoda, D.S.; Udayanga, D.; Herath, I.S.; Thambugala, K.M.; Manawasinghe, I.S.; et al. What is a species in fungal plant pathogens? Fungal Divers. 2021, 109, 239–266. [Google Scholar] [CrossRef]
- Manawasinghe, I.S.; Phillips, A.J.; Xu, J.; Balasuriya, A.; Hyde, K.D.; Stępień, Ł.; Harischandra, D.L.; Karunarathna, A.; Yan, J.; Weerasinghe, J.; et al. Defining a species in fungal plant pathology: Beyond the species level. Fungal Divers. 2021, 109, 267–282. [Google Scholar] [CrossRef]
- Bhunjun, C.S.; Niskanen, T.; Suwannarach, N.; Wannathes, N.; Chen, Y.J.; McKenzie, E.H.C.; Maharachchikumbura, S.S.N.; Buyck, B.; Zhao, C.L.; Fan, Y.G.; et al. The numbers of fungi: Are the most speciose genera truly diverse? Fungal Divers. 2022, 114, 387–462. [Google Scholar] [CrossRef]
- Kirk, P.M.; Cannon, P.F.; Minter, D.W.; Stalpers, J.A. Dictionary of the Fungi, 10th ed.; CABI Bioscience: Wallingford, UK, 2008. [Google Scholar]
- CABI. Leptosphaeria maculans (stem canker). Distribution Maps of Plant Diseases; CAB International: Wallingford, UK, 2019; Available online: https://www.cabi.org/isc/datasheet/31468 (accessed on 20 February 2023).
- Farr, D.F.; Rossman, A.Y.; Fungal Databases. U.S. National Fungus Collections, ARS, USDA. 2021. Available online: https://nt.ars-grin.gov/fungaldatabases (accessed on 15 February 2023).
- Petrak, F. Mykologische Notizen. III. Annales Mycologici 1921, 19, 176–223. [Google Scholar]
- Petrak, F. List of New Species and Varieties of Fungi, New Combinations and New Names Published 1932-1935; Commonwealth Mycological Institute: Egham, UK, 1944; Volume 7, p. 983. [Google Scholar]
- Tennakoon, D.S.; Gentekaki, E.; Jeewon, R.; Kuo, C.H.; Promputtha, I.; Hyde, K.D. Life in leaf litter: Fungal community succession during decomposition. Mycosphere 2021, 12, 406–429. [Google Scholar] [CrossRef]
- Tennakoon, D.S.; Kuo, C.H.; Maharachchikumbura, S.S.; Thambugala, K.M.; Gentekaki, E.; Phillips, A.J.; Bhat, D.J.; Wanasinghe, D.N.; de Silva, N.I.; Promputtha, I.; et al. Taxonomic and phylogenetic contributions to Celtis formosana, Ficus ampelas, F. septica, Macaranga tanarius and Morus australis leaf litter inhabiting microfungi. Fungal Divers. 2021, 108, 1–215. [Google Scholar] [CrossRef]
- Hyde, K.D.; Norphanphoun, C.; Chen, J.; Dissanayake, A.J.; Doilom, M.; Hongsanan, S.; Jayawardena, R.S.; Jeewon, R.; Perera, R.H.; Thongbai, B.; et al. Thailand’s amazing diversity: Up to 96% of fungi in northern Thailand may be novel. Fungal Divers. 2018, 93, 215–239. [Google Scholar] [CrossRef]
- Hyde, K.D.; Jeewon, R.; Chen, Y.J.; Bhunjun, C.S.; Calabon, M.S.; Jiang, H.B.; Lin, C.G.; Norphanphoun, C.; Sysouphanthong, P.; Pem, D.; et al. The numbers of fungi: Is the descriptive curve flattening? Fungal Divers. 2020, 103, 219–271. [Google Scholar] [CrossRef]
- Liu, S.Q. Preliminary study on the development path of Zhaotong literature. Yunnan People’s Publ. House China 2015, 12, 178. [Google Scholar]
- Pei, Y. Analysis of temperature variation characteristics in Zhaotong City in recent 50 years. J. Agric. Catastrophol. 2022, 12, 3. [Google Scholar]
- Wang, L.; Shi, Z.; Ye, L.; Su, B. Analysis on the characteristics of extreme weather events in Kunming City during recent 20 Years. IOP Conf. Ser. Earth Environ. Sci. 2019, 252, 042124. [Google Scholar] [CrossRef]
- Wanasinghe, D.N.; Wijayawardene, N.N.; Xu, J.; Cheewangkoon, R.; Mortimer, P.E. Taxonomic novelties in Magnolia-associated pleosporalean fungi in the Kunming Botanical Gardens (Yunnan, China). PLoS ONE 2020, 15, e0235855. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Chen, S.L.; Dai, Y.C.; Jia, Z.F.; Li, T.H.; Liu, T.Z.; Phurbu, D.; Mamut, R.; Sun, G.Y.; Bau, T.; et al. Overview of China’s nomenclature novelties of fungi in the new century (2000–2020). Mycosystema 2021, 40, 822–833. [Google Scholar]
- Shoemaker, R.A. Canadian and some extralimital Leptosphaeria species. Can. J. Bot. 1984, 62, 2688–2729. [Google Scholar] [CrossRef]
- Hyde, K.D.; Hongsanan, S.; Jeewon, R.; Bhat, J.D.; McKenzie, E.H.C.; Jones, E.B.G.; Phookamsak, R.; Ariyawansa, H.A.; Boonmee, S.; Zhao, Q.; et al. Fungal diversity notes 367–490: Taxonomic and phylogenetic contributions to fungal taxa. Fungal Divers. 2016, 80, 1–270. [Google Scholar] [CrossRef]
- Dayarathne, M.C.; Phookamsak, R.; Ariyawansa, H.A.; Jones, E.B.G.; Camporesi, E.; Hyde, K.D. Phylogenetic and morphological appraisal of Leptosphaeria italica sp. nov. (Leptosphaeriaceae, Pleosporales) from Italy. Mycosphere 2015, 6, 634–642. [Google Scholar] [CrossRef]
- Huhndorf, S.M. Systematics of Leptosphaeria species found on the Rosaceae. Ill. Nat. Hist. Surv. Bull. 1992, 34, 479–533. [Google Scholar] [CrossRef]
- Müller, E. Die schweizerischen arten der gattung Leptosphaeria und ihrer verwandten. Sydowia Ann. Mycol. Ed. Notitiam Sci. Mycol. Univers. 1950, 4, 185–319. [Google Scholar]
- Macia, M.J.; Palm, M.E.; Martin, M.P. A new species of Leptosphaeria (Ascomycotina, Pleosporales) on Rosaceae from Bolivia. Mycotaxon 2005, 93, 401–406. [Google Scholar]
- Boerema, G.H.; de Gruyter, J. Contributions towards a monograph of Phoma (Coelomycetes)–III-Supplement: Additional species of section Plenodomus. Pers. Mol. Phylogeny Evol. Fungi 1999, 17, 273–280. [Google Scholar]
- Quaedvlieg, W.; Verkley, G.J.; Shin, H.D.; Barreto, R.W.; Alfenas, A.C.; Swart, W.J.; Groenewald, J.Z.; Crous, P.W. Sizing up Septoria. Stud. Mycol. 2013, 75, 307–390. [Google Scholar] [CrossRef]
- Shoemaker, R.A.; Brun, H. The teleomorph of the weakly aggressive segregate of Leptosphaeria maculans. Can. J. Bot. 2001, 79, 412–419. [Google Scholar] [CrossRef]
- Dearness, J.; House, H.D. New or noteworthy species of fungi. II. Bull. New York State Mus. 1921, 233–234, 32–43. [Google Scholar]
- Lucas, M.T. Culture studies on Portuguese species of Leptosphaeria I. Trans. Br. Mycol. Soc. 1963, 46, 361–367. [Google Scholar] [CrossRef]
- Saccardo, P.A. Fungi nonnulli extra-Italici novi ex herbariis C.C. Gillet, P. Morthier, et G. Winter. Michelia 1878, 1, 357–360. [Google Scholar]
- Berlese, A.N. Excursion mycologique dans le Frioul. Bull. De La Société Mycol. De Fr. 1889, 5, 36–59. [Google Scholar]
- Fuckel, L. Symbolae mycologicae. Beiträge zur Kenntniss der Rheinischen Pilze. Jahrbücher des Nassauischen Vereins für Naturkunde 1870, 23–24, 1–459. [Google Scholar]
- Fuckel, L. Symbolae mycologicae. Beiträge zur Kenntniss der rheinischen Pilze. Zweiter Nachtrag; Julius Neidner: Wiesbaden, Germany, 1873; pp. 1–99. [Google Scholar]
- Lowen, R.; Sivanesan, A. Leptosphaeria pimpinellae and its Phoma anamorph. Mycotaxon 1989, 35, 205–210. [Google Scholar]
- Boerema, G.H.; Loerakker, W.M. Notes on Phoma. 2. Trans. Br. Mycol. Soc. 1985, 84, 289–302. [Google Scholar] [CrossRef]
- Ellis, J.B.; Everhart, B.M. New species of fungi from Washington Territory. Collected by W.N. Suksdorf during the summer and fall of 1883. Bull. Washburn Lab. Nat. Hist. 1884, 1, 3–6. [Google Scholar]
- Punithalingam, E.; Holliday, P. Deuterophoma tracheiphila. In Descriptions of Pathogenic Fungi and Bacteria; CABI International: Wallingford, UK, 1973; p. 399. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genes/Loci | PCR Primers (Forward/Reverse) | PCR Thermal Cycle Protocols | References |
|---|---|---|---|
| ITS | ITS5/ITS4 | a Annealing at 55 °C for 15 s c | [35] |
| LSU | LR0R/LR5 | [36] | |
| SSU | NS1/NS4 | a Annealing at 55 °C for 30 s c | [35] |
| tub2 | T1/Bt2b | [37] | |
| rpb2 | fRPB2-5F/fRPB2-7cR | b Annealing at 57 °C for 50 s c | [38] |
| Species Name | Strain Numbers | GenBank Accession Numbers | ||||
|---|---|---|---|---|---|---|
| ITS | LSU | SSU | tub2 | rpb2 | ||
| Alloleptosphaeria clematidis | MFLUCC 17-2071 | MT310604 | MT214557 | MT226674 | NA | NA |
| All. iridicola | CBS 143395 | MH107919 | MH107965 | NA | NA | NA |
| All. italica | MFLUCC 14-0934 | KT454722 | KT454714 | NA | NA | NA |
| All. shangrilana | HKAS 112210 | MW431059 | MW431315 | MW431058 | NA | NA |
| Alternariaster bidentis | CBS 134021 | KC609333 | KC609341 | NA | NA | KC609347 |
| Alt. bidentis | CBS 134185 | KC609334 | KC609342 | NA | NA | KC609348 |
| Alt. centaureae-diffusae | MFLUCC 14-0992 | KT454723 | KT454715 | KT454730 | NA | NA |
| Alt. centaureae-diffusae | MFLUCC 15-0009 | KT454724 | KT454716 | KT454731 | NA | NA |
| Alt. centaureae-diffusae | HMJAU 60188 | OL996125 | OL897175 | OL891810 | OL898721 | NA |
| Alt. helianthi | CBS 199.86 | KC609336 | KC609343 | NA | NA | KC609349 |
| Alt. helianthi | CBS 327.69 | KC609335 | KC584369 | KC584627 | NA | KC584494 |
| Alt. trigonosporus | MFLU 15-2237 | KY674857 | KY674858 | NA | NA | NA |
| Angularia xanthoceratis | HMJAU 60197 | OM295683 | OM295682 | OM295681 | OM304358 | NA |
| Didymella exigua | CBS 183.55 | EF192139 | EU754155 | EU754056 | GU237525 | EU874850 |
| D. maydis | CBS 588.69 | FJ427086 | MH871149 | EU754093 | FJ427190 | GU371782 |
| Heterosporicola beijingensis | JZB3400001 | MN733734 | MN737597 | MN733738 | NA | NA |
| H. beijingensis | JZB3400002 | MN733735 | MN737598 | MN733739 | NA | NA |
| H. beijingensis | JZB3400003 | MN733736 | MN737599 | MN733740 | NA | NA |
| H. beijingensis | JZB3400004 | MN733737 | MN737600 | MN733741 | NA | NA |
| H. chenopodii | CBS 448.68 | FJ427023 | EU754187 | EU754088 | NA | NA |
| H. chenopodii | CBS 115.96 | JF740227 | EU754188 | EU754089 | NA | GU371775 |
| H. dimorphospora | CBS 345.78 | JF740203 | GU238069 | GU238213 | NA | NA |
| H. dimorphospora | CBS 165.78 | JF740204 | JF740281 | JF740098 | NA | NA |
| Leptosphaeria chatkalica | YGS22 | MW886101 | MW886099 | MW886100 | NA | NA |
| L. cichorii | MFLUCC 14-1063 | KT454720 | KT454712 | KT454728 | NA | NA |
| L. cirsii | MFLUCC 14-1170 | NR155328 | NG059725 | NA | NA | NA |
| L. conoidea | CBS 125977 | JF740202 | JF740280 | NA | NA | NA |
| L. conoidea | CBS 616.75 | JF740201 | JF740279 | JF740099 | KT389804 | KT389639 |
| L. conoidea | FeF93 | MZ492958 | NA | NA | NA | NA |
| L. doliolum | MFLUCC 15-1875 | KT454727 | KT454719 | KT454734 | NA | NA |
| L. doliolum | CBS 155.94 | JF740207 | JF740282 | NA | JF740146 | NA |
| L. doliolum | CBS 505.75 | JF740205 | GQ387576 | GQ387515 | JF740144 | KY064035 |
| L. doliolum | CBS 541.66 | JF740206 | JF740284 | NA | JF740145 | NA |
| L. doliolum | CBS 130000 | JF740210 | NA | NA | JF740149 | NA |
| L. ebuli | MFLUCC 14-0828 | KP744446 | KP744488 | KP753954 | NA | NA |
| L. errabunda | CBS 617.75 | JF740216 | JF740289 | NA | JF740150 | NA |
| L. errabunda | CBS 125978 | JF740217 | JF740290 | NA | JF740151 | NA |
| L. errabunda | CBS 129998 | JF740219 | MH877027 | NA | JF740153 | NA |
| L. italica | MFLU15-0174 | NA | KT783670 | NA | NA | NA |
| L. irregularis | MFLUCC 15-1118 | KX856056 | KX856055 | NA | NA | NA |
| L. macrocapsa | CBS 640.93 | JF740237 | JF740304 | NA | JF740156 | NA |
| L. pedicularis | CBS 390.80 | JF740224 | JF740294 | NA | JF740155 | NA |
| L. pedicularis | CBS 126582 | JF740223 | JF740293 | NA | NA | NA |
| L. proteicola | CPC:18289 | JQ044439 | JQ044458 | NA | NA | NA |
| L. regiae | MFLUCC 18-1137 | MN244201 | MN244171 | MN244177 | NA | NA |
| L. sclerotioides | CBS 144.84 | JF740192 | JF740269 | NA | NA | NA |
| L. sclerotioides | CBS 148.84 | JF740193 | JF740270 | NA | NA | NA |
| L. sclerotioides | FeF422 | MZ492959 | NA | NA | NA | NA |
| L. sclerotioides | P10 | MT996500 | MT996501 | NA | MT996502 | MT996503 |
| L. sclerotioides | P9 | MT996499 | MT992704 | NA | MT989358 | MT992705 |
| Leptosphaeria sp. | LW119 | MH128282 | NA | NA | NA | NA |
| Leptosphaeria sp. | LW113 | MH128276 | NA | NA | NA | NA |
| L. slovacica | CBS 389.80 | JF740247 | JF740315 | JF740101 | NA | NA |
| L. slovacica | CBS 125975 | JF740248 | JF740316 | NA | NA | NA |
| L. sydowii | CBS 385.80 | JF740244 | JF740313 | NA | JF740157 | NA |
| L. sydowii | CBS 125976 | JF740245 | JF740314 | NA | JF740158 | NA |
| L. urticae | MFLU 18-0591 | MK123333 | MK123332 | MK123329 | NA | NA |
| L. urticae | FeF166 | MZ492960 | NA | NA | NA | NA |
| L. veronicae | CBS 126583 | JF740255 | JF740321 | NA | JF740161 | NA |
| L. veronicae | CBS 145.84 | JF740254 | JF740320 | NA | JF740160 | NA |
| L. yunnanensis | CGMCC 3.23748 | OP494319 | OP494327 | OP494333 | OP476696 | NA |
| L. yunnanensis | CGMCC 3.23749 | OP494320 | OP494328 | OP494334 | OP476697 | NA |
| L. yunnanensis | HKAS 124671 | OP494321 | OP494329 | OP494335 | OP476698 | NA |
| L. zhaotongensis | HKAS 124664 | OP494318 | OP494326 | OP494332 | OP476695 | NA |
| L. zhaotongensis | HMAS 352282 | OQ446062 | OQ446132 | OQ448836 | OQ511597 | |
| Neoleptosphaeria jonesii | MFLUCC 16-1442 | KY211869 | KY211870 | KY211871 | NA | NA |
| N. rubefaciens | CBS 223.77 | JF740243 | JF740312 | NA | NA | NA |
| N. rubefaciens | CBS 387.80 | JF740242 | JF740311 | NA | NA | NA |
| Ochraceocephala foeniculi | CBS 145654 | MN516753 | MN516774 | MN516743 | MN520147 | MN520145 |
| O. foeniculi | Di3AF15 | MN516766 | MN516783 | MN516752 | NA | NA |
| Paraleptosphaeria dryadis | CBS 643.86 | JF740213 | GU301828 | KC584632 | NA | GU371733 |
| Pa. dryadis | CBS 743.86 | AF439461 | NA | NA | NA | NA |
| Pa. kunmingensis | KUNCC 23-12732 | OP494316 | OP494324 | OP494330 | OP476693 | OP476691 |
| Pa. kunmingensis | KUNCC 23-12731 | OQ446060 | OQ446130 | OQ448834 | OQ511598 | OQ455053 |
| Pa. macrospora | CBS 114198 | JF740238 | JF740305 | NA | NA | NA |
| Pa. nitschkei | CBS 306.51 | JF740239 | JF740308 | NA | KT389833 | KT389660 |
| Pa. nitschkei | MFLUCC 13-0688 | KR025860 | KR025864 | NA | NA | NA |
| Pa. orobanches | CBS 101638 | JF740230 | JF740299 | A | NA | NA |
| Pa. padi | MFLU 15-2756 | KY554203 | KY554198 | KY554201 | NA | NA |
| Pa. polylepidis | APA-2999 | MK795714 | MK795717 | NA | NA | NA |
| Pa. praetermissa | CBS 114591 | JF740241 | JF740310 | NA | NA | NA |
| Pa. rubi | MFLUCC 14-0211 | KT454726 | KT454718 | KT454733 | NA | NA |
| Pa. rumicis | CBS 522.78 | KF251144 | KF251648 | NA | NA | NA |
| Plenodomus agnitus | CBS 121.89 | JF740194 | JF740271 | NA | KY064053 | KY064036 |
| Pl. agnitus | CBS 126584 | JF740195 | JF740272 | NA | NA | NA |
| Pl. agnitus | MFLU 15-0039 | KP744459 | KP744504 | NA | NA | NA |
| Pl. artemisiae | KUMCC 18-0151 | MK387920 | MK387958 | MK387928 | NA | MK435607 |
| Pl. artemisiae | KUMCC 20-0200A | MT957062 | MT957055 | MT957048 | NA | NA |
| Pl. artemisiae | KUMCC 20-0200B | MT957063 | MT957056 | MT957049 | NA | NA |
| Pl. biglobosus | CBS 119951 | JF740198 | JF740274 | JF740102 | KY064054 | KY064037 |
| Pl. biglobosus | CBS 127249 | JF740199 | JF740275 | NA | NA | NA |
| Pl. changchunensis | CCMJ5011 | OL996123 | OL897174 | OL984031 | NA | NA |
| Pl. changchunensis | CCMJ5012 | OL996124 | OL966928 | OL984032 | OL898716 | OL944508 |
| Pl. collinsoniae | VT02 | MN653010 | MN982862 | MN652269 | NA | NA |
| Pl. collinsoniae | CBS 120227 | JF740200 | JF740276 | NA | KY064056 | KY064039 |
| Pl. collinsoniae | KNU-AP100C | LC550566 | LC550568 | NA | NA | NA |
| Pl. collinsoniae | KNU-20-A1 | LC591836 | NA | NA | LC591846 | LC591841 |
| Pl. collinsoniae | KNU-20-A2 | LC591837 | NA | NA | LC591847 | LC591842 |
| Pl. collinsoniae | KNU-20-A3 | LC591838 | NA | NA | LC591848 | LC591843 |
| Pl. collinsoniae | KNU-20-A4 | LC591839 | NA | NA | LC591849 | LC591844 |
| Pl. collinsoniae | KNU-20-C4 | LC591840 | NA | NA | LC591850 | LC591845 |
| Pl. collinsoniae | FeC109 | MW446975 | NA | NA | NA | NA |
| Pl. collinsoniae | EF194 | MK842112 | NA | NA | NA | NA |
| Pl. confertus | CBS 375.64 | AF439459 | JF740277 | NA | KY064057 | KY064040 |
| Pl. congestus | CBS 244.64 | AF439460 | JF740278 | NA | KY064058 | KY064041 |
| Pl. deqinensis | CGMCC 3.18221 | KY064027 | KY064031 | NA | KY064052 | KY064034 |
| Pl. dezfulensis | IRAN 4159C | MZ048609 | NA | NA | MZ043102 | MZ043104 |
| Pl. dezfulensis | SCUA-Ahm-S41-2 | MZ048610 | NA | NA | MZ043103 | MZ043105 |
| Pl. enteroleucus | CBS 142.84 | JF740214 | JF740287 | NA | KT266266 | KY064042 |
| Pl. enteroleucus | CBS 831.84 | JF740215 | JF740288 | NA | KT266270 | NA |
| Pl. enteroleucus | F-146,176 | MN910295 | MN910294 | NA | NA | NA |
| Pl. enteroleucus | ICMP:10937 | KT309810 | KT309635 | NA | KT309399 | NA |
| Pl. fallaciosus | CBS 414.62 | JF740222 | JF740292 | NA | NA | KY064043 |
| Pl. guttulatus | MFLUCC 15-1876 | KT454721 | KT454713 | KT454729 | NA | NA |
| Pl. hendersoniae | CBS 113702 | JF740225 | JF740295 | NA | KT266271 | KY064044 |
| Pl. hendersoniae | CBS 139.78 | JF740226 | JF740296 | NA | NA | NA |
| Pl. hendersoniae | LTO | MF795790 | NA | NA | NA | MF795832 |
| Pl. influorescens | CBS 143.84 | JF740228 | JF740297 | NA | KT266267 | KY064045 |
| Pl. influorescens | PD 73/1382 | JF740229 | JF740298 | NA | KT266273 | NA |
| Pl. libanotidis | CBS 113795 | JF740231 | JF740300 | NA | KY064059 | KY064046 |
| Pl. lijiangensis | KUMCC 18-0186 | MK387921 | MK387959 | MK387929 | NA | NA |
| Pl. lindquistii | CBS 386.80 | JF740232 | JF740301 | NA | NA | NA |
| Pl. lindquistii | CBS 381.67 | JF740233 | JF740302 | NA | NA | NA |
| Pl. lindquistii | MF-Ha 16-005 | MK495988 | NA | NA | MK501790 | NA |
| Pl. lingam | AFTOL-ID 277 | KT225526 | DQ470946 | DQ470993 | NA | DQ470894 |
| Pl. lingam | CBS 275.63 | JF740234 | JF740306 | JF740103 | KT389841 | KT389669 |
| Pl. lingam | CBS 260.94 | JF740235 | JF740307 | NA | MZ073915 | KY064047 |
| Pl. lingam | CBS 147.24 | MH854784 | MH866288 | NA | MZ073914 | NA |
| Pl. lupini | CBS 248.92 | JF740236 | JF740303 | NA | KY064061 | KY064048 |
| Pl. pimpinellae | CBS 101637 | JF740240 | JF740309 | NA | KY064062 | NA |
| Pl. salviae | MFLUCC 13-0219 | KT454725 | KT454717 | KT454732 | NA | NA |
| Pl. sinensis | KUMCC 18-0152 | MK387923 | MK387961 | MK387931 | NA | NA |
| Pl. sinensis | KUMCC 18-0153 | MK387922 | MK387960 | MK387930 | NA | MK435608 |
| Pl. sinensis | KUN-HKAS 102227 | MK387924 | MK387962 | MK387932 | NA | NA |
| Pl. sinensis | MFLUCC 17-0757 | MF072722 | MF072718 | MF072720 | NA | NA |
| Pl. sinensis | MFLUCC 17-0767 | MF072721 | MF072717 | MF072719 | NA | NA |
| Pl. sinensis | KNU-GW1901 | LC550567 | LC550569 | LC550570 | NA | NA |
| Pl. sinensis | KUMCC 20-0204 | MT957064 | MT957057 | MT957050 | NA | NA |
| Pl. tracheiphilus | CBS 551.93 | JF740249 | JF740317 | JF740104 | MZ073918 | KY064049 |
| Pl. tracheiphilus | CBS 127250 | JF740250 | JF740318 | NA | MZ073919 | NA |
| Pl. tracheiphilus | MUCL 38481 | MW810293 | MW715037 | NA | MZ073920 | NA |
| Pl. tracheiphilus | ATCC 26007 | MZ049614 | MW959165 | NA | MZ073908 | MZ073893 |
| Pl. tracheiphilus | IS3-15 | MK461058 | NA | NA | NA | NA |
| Pl. triseptatus | MFLUCC 17-1345 | MN648452 | MN648451 | MN648453 | NA | NA |
| Pl. visci | CBS 122783 | JF740256 | EU754195 | EU754096 | KY064063 | KY064050 |
| Pl. visci | CPC:35314 | MT223830 | MT223922 | NA | NA | MT223696 |
| Pl. visci | CPC:35315 | MT223831 | MT223923 | NA | NA | NA |
| Pl. visci | CPC:35316 | MT223832 | MT223924 | NA | NA | NA |
| Pl. wasabiae | CBS 120119 | JF740257 | JF740323 | NA | KT266272 | NA |
| Pl. wasabiae | CBS 120120 | JF740258 | JF740324 | NA | NA | NA |
| Pl. zhaotongensis | CGMCC 3.23746 | OP494317 | OP494325 | OP494331 | OP476694 | OP476692 |
| Pl. zhaotongensis | CGMCC 3.23747 | OQ446061 | OQ446131 | OQ448835 | OQ511599 | OQ455054 |
| Praeclarispora artemisiae | KUMCC 20-0201A | MT957060 | MT957053 | MT957046 | NA | NA |
| Pr. artemisiae | KUMCC 20-0201B | MT957061 | MT957054 | MT957047 | NA | NA |
| Pseudoleptosphaeria etheridgei | CBS 125980 | JF740221 | JF740291 | NA | NA | MT394686 |
| Querciphoma carteri | CBS 101633 | KF251210 | GQ387593 | GQ387532 | KF252701 | NA |
| Querciphoma carteri | CBS 105.91 | KF251209 | GQ387594 | GQ387533 | KF252700 | NA |
| Sclerenchymomyces clematidis | MFLUCC 17-2180 | MT310605 | MT214558 | MT226675 | NA | NA |
| Sphaerellopsis artemisiae | KUMCC 20-0202A | MT957065 | MT957058 | MT957051 | NA | NA |
| Sp. artemisiae | KUMCC 20-0202B | MT957066 | MT957059 | MT957052 | NA | NA |
| Sp. filum | CBS 234.51 | KP170655 | KP170723 | NA | KP170704 | NA |
| Sp. filum | CBS 235.51 | KP170656 | KP170724 | NA | KP170705 | NA |
| Sp. filum | CBS 317.68 | KP170657 | KP170725 | NA | KP170706 | NA |
| Sp. hakeae | CPC:29566 | KY173466 | KY173555 | NA | NA | NA |
| Sp. isthmospora | HKAS 102225A | MK387925 | MK387963 | NA | NA | NA |
| Sp. isthmospora | HKAS 102225B | MK387926 | MK387964 | MK387934 | NA | NA |
| Sp. macroconidialis | CBS 233.51 | KP170658 | KP170726 | NA | KP170707 | NA |
| Sp. macroconidialis | CBS 658.78 | KP170659 | KP170727 | NA | KP170708 | NA |
| Sp. macroconidialis | CPC:21113 | KP170660 | KP170728 | NA | KP170709 | NA |
| Sp. paraphysata | CPC:21841 | KP170662 | KP170729 | NA | KP170710 | NA |
| Sp. paraphysata | KUMCC 18-0195 | MK387927 | MK387965 | MK387935 | NA | NA |
| Subplenodomus apiicola | CBS 285.72 | JF740196 | GU238040 | GU238211 | NA | NA |
| Su. apiicola | CBS 421.50 | MH856699 | MH868215 | NA | NA | NA |
| Su. drobnjacensis | CBS 270.92 | JF740212 | JF740286 | NA | NA | NA |
| Su. drobnjacensis | CBS 269.92 | JF740211 | JF740285 | JF740100 | NA | NA |
| Su. galicola | MFLU 15-1368 | KY554204 | KY554199 | NA | NA | NA |
| Su. valerianae | CBS 630.68 | JF740251 | GU238150 | GU238229 | NA | NA |
| Su. valerianae | CBS 499.91 | JF740252 | JF740319 | NA | NA | NA |
| Su. violicola | CBS 306.68 | FJ427083 | GU238156 | GU238231 | KT389849 | NA |
| Shiraia bambusicola | GZAAS2.0703 | GQ845412 | KC460981 | NA | NA | NA |
| Shiraia bambusicola | GZAAS2.0629 | GQ845415 | KC460980 | NA | NA | NA |
| Tzeanania taiwanensis | NTUCC 17-005 | MH461123 | MH461120 | MH461126 | MH461132 | NA |
| Tzeanania taiwanensis | NTUCC 17-006 | MH461124 | MH461121 | MH461127 | MH461133 | NA |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gao, Y.; de Farias, A.R.G.; Jiang, H.-B.; Karunarathna, S.C.; Xu, J.-C.; Tibpromma, S.; Gui, H. Morphological and Phylogenetic Characterisations Reveal Four New Species in Leptosphaeriaceae (Pleosporales, Dothideomycetes). J. Fungi 2023, 9, 612. https://doi.org/10.3390/jof9060612
Gao Y, de Farias ARG, Jiang H-B, Karunarathna SC, Xu J-C, Tibpromma S, Gui H. Morphological and Phylogenetic Characterisations Reveal Four New Species in Leptosphaeriaceae (Pleosporales, Dothideomycetes). Journal of Fungi. 2023; 9(6):612. https://doi.org/10.3390/jof9060612
Chicago/Turabian StyleGao, Ying, Antonio Roberto Gomes de Farias, Hong-Bo Jiang, Samantha C. Karunarathna, Jian-Chu Xu, Saowaluck Tibpromma, and Heng Gui. 2023. "Morphological and Phylogenetic Characterisations Reveal Four New Species in Leptosphaeriaceae (Pleosporales, Dothideomycetes)" Journal of Fungi 9, no. 6: 612. https://doi.org/10.3390/jof9060612
APA StyleGao, Y., de Farias, A. R. G., Jiang, H.-B., Karunarathna, S. C., Xu, J.-C., Tibpromma, S., & Gui, H. (2023). Morphological and Phylogenetic Characterisations Reveal Four New Species in Leptosphaeriaceae (Pleosporales, Dothideomycetes). Journal of Fungi, 9(6), 612. https://doi.org/10.3390/jof9060612