
Additions to Rhytidhysteron (Hysteriales, Dothideomycetes) in China

 ,
,  , ,
, ,  ,
,  , ,
, ,  and
and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Sample Collection, Morphological Identification, and Single Spore Isolation
2.2. DNA Extraction, PCR Amplification, and Sequencing
2.3. Phylogenetic Analyses
3. Results
3.1. Phylogenetic Analyses
3.2. Taxonomy
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Schoch, C.L.; Crous, P.W.; Groenewald, J.Z.; Boehm, E.W.A.; Burgess, T.I.; de Gruyter, J.; de Hoog, G.S.; Dixon, L.J.; Grube, M.; Gueidan, C.; et al. A class-wide phylogenetic assessment of Dothideomycetes. Stud. Mycol. 2009, 64, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Hyde, K.D.; Jones, E.B.G.; Liu, J.K.; Ariyawansa, H.; Boehm, E.; Boonmee, S.; Braun, U.; Chomnunti, P.; Crous, P.W.; Dai, D.Q.; et al. Families of Dothideomycetes. Fungal Divers. 2013, 63, 1–313. [Google Scholar] [CrossRef]
- Chethana, K.W.; Manawasinghe, I.S.; Hurdeal, V.G.; Bhunjun, C.S.; Appadoo, M.A.; Gentekaki, E.; Raspé, O.; Promputtha, I.; Hyde, K.D. What are fungal species and how to delineate them? Fungal Divers. 2021, 109, 1–25. [Google Scholar] [CrossRef]
- Hongsanan, S.; Hyde, K.D.; Phookamsak, R.; Wanasinghe, D.N.; McKenzie, E.H.C.; Sarma, V.V.; Boonmee, S.; Lücking, R.; Pem, D.; Bhat, J.D.; et al. Refined families of Dothideomycetes: Dothideomycetidae and Pleosporomycetidae. Mycosphere 2020, 11, 1553–2107. [Google Scholar] [CrossRef]
- Wijayawardene, N.N.; Hyde, K.D.; Dai, D.Q.; Sánchez-García, M.; Goto, B.T.; Saxena, R.K.; Erdoğdu, M.; Selçuk, F.; Rajeshkumar, K.C.; Aptroot, A.; et al. Outline of fungi and fungus-like taxa—2021. Mycosphere 2022, 13, 53–453. [Google Scholar] [CrossRef]
- De Silva, N.I.; Tennakoon, D.S.; Thambugala, K.M.; Karunarathna, S.C.; Lumyong, S.; Hyde, K.D. Morphology and multigene phylogeny reveal a new species and a new record of Rhytidhysteron (Dothideomycetes, Ascomycota) from China. Asian J. Mycol. 2020, 3, 295–306. [Google Scholar] [CrossRef]
- Zogg, H. Die Hysteriaceae s. str. und Lophiaceae, unter besonderer berücksichtigung der mitteleuropäischen formen. Ber. Zur Kryptogamenflora Der Schweiz. 1962, 11, 1–190. [Google Scholar]
- Müller, E.; von Arx, J.A. Einige aspekte zur systematik pseudopharilaler ascomyceten. Ber. Schweiz. Bot. Ges. 1950, 60, 329–397. [Google Scholar]
- Luttrell, E.S. The ascostromatic Ascomycetes. Mycologia 1955, 47, 511–532. [Google Scholar] [CrossRef]
- Kirk, P.M.; Cannon, P.F.; David, J.C.; Stalpers, J.A. Ainsworth & Bisby’s Dictionary of the Fungi, 9th ed.; CABI: Wallingford, UK, 2001. [Google Scholar]
- Kirk, P.; Cannon, P.; Minter, D.; Stalpers, J.A. Dictionary of the fungi, 10th edn. CAB International, Wallingford fermentations. Appl. Microbiol. Biotechnol. 2008, 39, 36–41. [Google Scholar]
- Kumar, V.; Cheewangkoon, R.; Thambugala, K.M.; Jones, G.E.; Brahmanage, R.S.; Doilom, M.; Jeewon, R.; Hyde, K.D. Rhytidhysteron mangrovei (Hysteriaceae), a new species from mangroves in Phetchaburi Province, Thailand. Phytotaxa 2019, 401, 166–178. [Google Scholar] [CrossRef]
- Boehm, E.W.; Schoch, C.L.; Spatafora, J.W. On the evolution of the Hysteriaceae and Mytilinidiaceae (Pleosporomycetidae, Dothideomycetes, Ascomycota) using four nuclear genes. Mycol. Res. 2009, 113, 461–479. [Google Scholar] [CrossRef] [PubMed]
- Boehm, E.W.A.; Mugambi, G.; Miller, A.N.; Huhndorf, S.; Marincowitz, S.; Schoch, C.L.; Spatafora, J.W. A molecular phylogenetic reappraisal of the Hysteriaceae, Mytilinidiaceae and Gloniaceae (Pleosporomycetidae, Dothideomycetes) with keys to world species. Stud. Mycol. 2009, 64, 49–83. [Google Scholar] [CrossRef] [PubMed]
- Almeida, D.A.C. A new genus and three new species of hysteriaceous Ascomycetes from the semiarid region of Brazil. Phytotaxa 2014, 176, 298–308. [Google Scholar] [CrossRef]
- Thambugala, K.M.; Hyde, K.D.; Eungwanichayapant, P.D.; Romero, A.I.; Liu, Z.Y. Additions to the genus Rhytidhysteron in Hysteriaceae. Cryptogam. Mycol. 2016, 37, 99–116. [Google Scholar] [CrossRef]
- Tibpromma, S.; Hyde, K.D.; Jeewon, R.; Maharachchikumbura, S.S.N.; Liu, J.K.; Bhat, D.J.; Jones, E.B.G.; McKenzie, E.H.C.; Camporesi, E.; Bulgakov, T.S.; et al. Fungal diversity notes 491–602: Taxonomic and phylogenetic contributions to fungal taxa. Fungal Divers. 2017, 83, 1–261. [Google Scholar] [CrossRef]
- Ren, G.C.; Wanasinghe, D.N.; Jeewon, R.; Monkai, J.; Mortimer, P.E.; Hyde, K.D.; Xu, J.C.; Gui, H. Taxonomy and phylogeny of the novel Rhytidhysteron-like collections in the Greater Mekong Subregion. MycoKeys 2022, 86, 65. [Google Scholar] [CrossRef]
- Spegazzini, C. Fungi argentini additis nonnullis brasiliensibus montevideensibusque. Pugillus quartus (Continuacion). An. Soc. Cient. Argent. 1881, 12, 174–189. [Google Scholar]
- Clements, F.E.; Shear, C.L. The Genera of Fungi; Hafner Publishing Co.: New York, NY, USA, 1931; p. 632. [Google Scholar]
- Samuels, G.J.; Müller, E. Life-history studies of Brazilian Ascomycetes. 7. Rhytidhysteron rufulum and the genus Eutryblidiella. Sydowia 1979, 32, 277–292. [Google Scholar]
- Yacharoen, S.; Tian, Q.; Chomnunti, P.; Boonmee, S.; Chukeatirote, E.; Bhat, J.D.; Hyde, K.D. Patellariaceae revisited. Mycosphere 2015, 6, 290–326. [Google Scholar] [CrossRef]
- Index Fungorum. 2022. Available online: http://www.indexfungorum.org/Names/Names.asp (accessed on 7 October 2022).
- Cobos-Villagrán, A.; Valenzuela, R.; Hernández-Rodríguez, C.; Calvillo-Medina, R.P.; Villa-Tanaca, L.; Mateo-Cid, L.E.; Pérez-Valdespino, A.; Martínez-González, C.R.; Raymundo, T. Three new species of Rhytidhysteron (Dothideomycetes, Ascomycota) from Mexico. MycoKeys 2021, 83, 123. [Google Scholar] [CrossRef]
- Wanasinghe, D.N.; Mortimer, P.E.; Xu, J. Insight into the systematics of microfungi colonizing dead woody twigs of Dodonaea viscosa in Honghe (China). J. Fungi 2021, 7, e180. [Google Scholar] [CrossRef] [PubMed]
- Farr, D.F.; Rossman, A.Y. Fungal Databases, U.S. National Fungus Collections, ARS, USDA. 2022. Available online: https://nt.ars-grin.gov/fungaldatabases/ (accessed on 12 November 2022).
- Soto-Medina, E.; Lücking, R. A new species of Rhytidhysteron (Ascomycota: Patellariaceae) from Colombia, with a provisional working key to known species in the world Artículo original. Rev. De La Acad. Colomb. De Cienc. Exactas Físicas Y Nat. 2017, 41, 59–63. [Google Scholar] [CrossRef]
- Senanayake, I.C.; Rathnayaka, A.R.; Marasinghe, D.S.; Calabon, M.S.; Gentekaki, E.; Lee, H.B.; Hurdeal, V.G.; Pem, D.; Dissanayake, L.S.; Wijesinghe, S.N.; et al. Morphological approaches in studying fungi, collection examination isolation sporulation and preservation. Mycosphere 2020, 11, 2678–2754. [Google Scholar] [CrossRef]
- Jayasiri, S.C.; Hyde, K.D.; Ariyawansa, H.A.; Bhat, J.; Buyck, B.; Cai, L.; Dai, Y.C.; Abd-Elsalam, K.A.; Ertz, D.; Hidayat, I.; et al. The Faces of fungi database: Fungal names linked with morphology, phylogeny and human impacts. Fungal Divers. 2015, 74, 3–18. [Google Scholar] [CrossRef]
- Dissanayake, A.J.; Bhunjun, C.S.; Maharachchikumbura, S.S.N.; Liu, J.K. Applied aspects of methods to infer phylogenetic relationships amongst fungi. Mycosphere 2020, 11, 2652–2676. [Google Scholar] [CrossRef]
- Vilgalys, R.; Hester, M. Rapid genetic identification and mapping of enzymatically amplified ribosomal DNA from several Cryptococcus species. J. Bacteriol. 1990, 172, 4238–4246. [Google Scholar] [CrossRef] [PubMed]
- White, T.J.; Bruns, T.; Lee, S.J.W.T.; Taylor, J.L. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols, a Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: San Diego, CA, USA, 1990; Volume 18, pp. 315–322. [Google Scholar] [CrossRef]
- Rehner, S. Primers for Elongation Factor 1-Alpha (EF1-Alpha); Insect Biocontrol Laboratory, USDA, ARS, PSI: Beltsville, MD, USA, 2001. [Google Scholar]
- Du, T.Y.; Hyde, K.D.; Mapook, A.; Mortimer, P.E.; Xu, J.C.; Karunarathna, S.C.; Tibpromma, S. Morphology and phylogenetic analyses reveal Montagnula puerensis sp. nov. (Didymosphaeriaceae, Pleosporales) from southwest China. Phytotaxa 2021, 514, 001–025. [Google Scholar] [CrossRef]
- Hall, T.A. BioEdit, a user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar] [CrossRef]
- Katoh, K.; Rozewicki, J.; Yamada, K.D. MAFFT online service: Multiple sequence alignment, interactive sequence choice and visualization. Brief. Bioinform. 2019, 20, 1160–1166. [Google Scholar] [CrossRef]
- Capella-Gutiérrez, S.; Silla-Martínez, J.M.; Gabaldón, T. TrimAl: A tool for automated alignment trimming in large-scale phylogenetic analyses. Bioinformatics 2009, 25, 1972–1973. [Google Scholar] [CrossRef]
- Glez-Peña, D.; Gómez-Blanco, D.; Reboiro-Jato, M.; Fdez-Riverola, F.; Posada, D. ALTER, program-oriented conversion of DNA and protein alignments. Nucleic Acids Res. 2010, 38, 14–18. [Google Scholar] [CrossRef] [PubMed]
- Miller, M.A.; Pfeiffer, W.; Schwartz, T. Creating the CIPRES Science Gateway for inference of large phylogenetic trees. In 2010 Gateway Computing Environments Workshop (GCE); IEEE Computer Society: New Orleans, LA, USA, 2010; pp. 1–8. [Google Scholar] [CrossRef]
- Stamatakis, A. RAxML version 8, a tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef] [PubMed]
- Stamatakis, A.; Hoover, P.; Rougemont, J. A rapid bootstrap algorithm for the RAxML web servers. Syst. Biol. 2008, 57, 758–771. [Google Scholar] [CrossRef] [PubMed]
- Ronquist, F.; Teslenko, M.; Van Der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef]
- Rannala, B.; Yang, Z. Probability distribution of molecular evolutionary trees: A new method of phylogenetic inference. J. Mol. Evol. 1996, 43, 304–311. [Google Scholar] [CrossRef]
- Zhaxybayeva, O.; Gogarten, J.P. Bootstrap, Bayesian probability and maximum likelihood mapping: Exploring new tools for comparative genome analyses. BMC Genom. 2002, 3, 4. [Google Scholar] [CrossRef]
- Rambaut, A. 2012—FigTree version 1. 4. University of Edinburgh: Edinburgh, Scotland, 2012. [Google Scholar]
- Hyde, K.D.; Hongsanan, S.; Jeewon, R.; Bhat, D.J.; McKenzie, E.H.C.; Jones, E.B.G.; Phookamsak, R.; Ariyawansa, H.A.; Boonmee, S.; Zhao, Q.; et al. Fungal diversity notes 367–490: Taxonomic and phylogenetic contributions to fungal taxa. Fungal Divers. 2016, 80, 1–270. [Google Scholar] [CrossRef]
- Dayarathne, M.C.; Jones, E.B.G.; Maharachchikumbura, S.S.N.; Devadatha, B.; Sarma, V.V.; Khongphinitbunjong, K.; Chomnunti, P.; Hyde, K.D. Morpho-molecular characterization of microfungi associated with marine based habitats. Mycosphere 2020, 11, 1–188. [Google Scholar] [CrossRef]
- Mapook, A.; Hyde, K.D.; McKenzie, E.H.C.; Jones, E.B.G.; Bhat, D.J.; Jeewon, R.; Stadler, M.; Samarakoon, M.C.; Malaithong, M.; Tanunchai, B.; et al. Taxonomic and phylogenetic contributions to fungi associated with the invasive weed Chromolaena odorata (Siam weed). Fungal Divers. 2020, 101, 175. [Google Scholar] [CrossRef]
- Hyde, K.D.; Dong, Y.; Phookamsak, R.; Jeewon, R.; Bhat, D.J.; Jones, E.B.G.; Liu, N.G.; Abeywickrama, P.D.; Mapook, A.; Wei, D.P.; et al. Fungal diversity notes 1151–1276: Taxonomic and phylogenetic contributions on genera and species of fungal taxa. Fungal Divers. 2020, 100, 5–277. [Google Scholar] [CrossRef]
- Vu, D.; Groenewald, M.; De Vries, M.; Gehrmann, T.; Stielow, B.; Eberhardt, U.; Al-Hatmi, A.; Groenewald, J.Z.; Cardinali, G.; Houbraken, J.; et al. Large-scale generation and analysis of filamentous fungal DNA barcodes boosts coverage for kingdom fungi and reveals thresholds for fungal species and higher taxon delimitation. Stud. Mycol. 2018, 91, 23–36. [Google Scholar] [CrossRef]
- Cobos-Villagrán, A.; Raymundo, T.; Calvillo-Medina, R.P.; Valenzuela, R. Rhytidhysteron mexicanum sp. nov. (Dothideomycetes, Ascomycota) from the Sierra of Guadalupe, Trans Mexican Volcanic Belt. Phytotaxa 2021, 479, 275–286. [Google Scholar] [CrossRef]
- Mugambi, G.K.; Huhndorf, S.M. Parallel evolution of hysterothecial ascomata in ascolocularous fungi (Ascomycota, Fungi). Syst. Biodivers. 2009, 7, 453–464. [Google Scholar] [CrossRef]
- Doilom, M.; Dissanayake, A.J.; Wanasinghe, D.N.; Boonmee, S.; Liu, J.K.; Bhat, D.J.; Taylor, J.E.; Bahkali, A.H.; McKenzie, E.H.C.; Hyde, K.D. Microfungi on Tectona grandis (teak) in northern Thailand. Fungal Divers. 2016, 82, 107–182. [Google Scholar] [CrossRef]
- Hyde, K.D.; de Silva, N.I.; Jeewon, R.; Bhat, D.J.; Phookamsak, R.; Doilom, M.; Boonmee, S.; Jayawardena, R.S.; Maharachchikumbura, S.S.N.; Senanayake, I.C.; et al. AJOM new records and collections of fungi: 1–100. Asian J. Mycol. 2020, 3, 22–294. [Google Scholar] [CrossRef]
- Cobos-Villagrán, A.; Hernández-Rodríguez, C.; Valenzuela, R.; Villa-Tanaca, L.; Calvillo-Medina, R.P.; Mateo-Cid, L.E.; Martínez-Pineda, M.; Raymundo, T. El género Rhytidhysteron (Dothideomycetes, Ascomycota) en México. Acta Bot. Mex. 2020, 127, e1675. [Google Scholar] [CrossRef]
- Jayasiri, S.C.; Hyde, K.D.; Jones, E.B.G.; Perŝoh, D.D.; Camporesi, E.; Kang, J.C. Taxonomic novelties of hysteriform Dothideomycetes. Mycosphere 2018, 9, 803–837. [Google Scholar] [CrossRef]
- Murillo, C.; Federico, J.A.; Julieta, C.; Thorsten, L.; Giselle, T. Molecular data indicate that Rhytidhysteron rufulum (Ascomycetes, Patellariales) in Costa Rica consists of four distinct lineages corroborated by morphological and chemical characters. Mycol. Res. 2009, 113, 405–416. [Google Scholar] [CrossRef] [PubMed]
- Petrak, F. Ein beitrag zur pilzflora Floridas. Sydowia 1953, 7, 103–116. [Google Scholar]
- Sarbhoy, A.K.; Lal, G.; Varshney, J.L. Fungi of India (1967–71); Navyug Traders: New Delhi, India, 1971; p. 148. [Google Scholar]
- Dingley, J.M.; Fullerton, R.A.; McKenzie, E.H.C. Survey of agricultural pests and diseases. Records of fungi, bacteria, algae, and angiosperms pathogenic on plants in Cook Islands, Fiji, Kiribati, Niue, Tonga, Tuvalu, and Western Samoa. Tech. Rep. 1981, 2, 485. [Google Scholar]
- Yuan, Z.Q. Fungi and associated tree diseases in Melville Island, Northern Territory, Australia. Austral. Syst. Bot. 1996, 9, 337–360. [Google Scholar] [CrossRef]
- Pande, A.; Rao, V.G. A Compendium of Fungi on Legumes from India; Scientific Publishers: Rajasthan, India, 1998; p. 188. [Google Scholar]
- McKenzie, E.H.C.; Buchanan, P.K.; Johnston, P.R. Checklist of fungi on Nothofagus species in New Zealand. N. Z. J. Bot. 2000, 38, 635–720. [Google Scholar] [CrossRef]
- Minter, D.W.; Rodriguez Hernandez, M.; Mena Portales, J. Fungi of the Caribbean: An Annotated Checklist; PDMS Publishing: Surrey, UK, 2001; p. 946. [Google Scholar]
- Urtiaga, R. Host index of plant diseases and disorders from Venezuela—addendum. Unknown journal or publisher. 2004; 268. [Google Scholar]
- Urtiaga, R. Indice de enfermedades en plantas de Venezuela y Cuba, second edition. Unknown journal or publisher. 2004; 301. [Google Scholar]
- Sierra-López, D. Contribución al estudio de los ascomicetes bitunicados de Cataluña. Acta Bot. Barcinon. 2006, 50, 5–434. [Google Scholar]
- Pande, A. Ascomycetes of Peninsular India; Scientific Publishers: Rajasthan, India, 2008; p. 584. [Google Scholar]
- Niranjan, M.; Tiwari, S.; Baghela, A.; Sarma, V.V. New records of ascomycetous fungi from Andaman Islands, India and their molecular sequence data. Curr. Res. Environ. Appl. Mycol. 2018, 8, 331–350. [Google Scholar] [CrossRef]
- Camino-Vilaro, M.; Castro-Hernandez, L.; Abreu-Herrera, Y.; Mena-Portales, J.; Cantillo-Perez, T. Fungi associated with invasive plant species in Cuba. Phytotaxa 2019, 419, 239–267. [Google Scholar] [CrossRef]
- Reale Accademia delle scienze fisiche e matematiche di NapoliSocietà reale di Napoli. Atti della Reale accademia delle scienze fisiche e matematiche; Tip. della R. Accademia delle scienze fisiche et matematiche Napoli 1863–1935; 1879; Volume 8. [Google Scholar]
- Sociedad Científica Argentina and Congreso Científico Latino Americano and Congreso Científico Internacional Americano. Anales de la Sociedad Científica Argentina; Buenos Aires, [Sociedad Científica Argentina]: Buenos Aires, Argentina, 1879; t.8-9.
- Sociedad Científica Argentina and Congreso Científico Latino Americano and Congreso Científico Internacional Americano. Anales de la Sociedad Científica Argentina; Buenos Aires, [Sociedad Científica Argentina]: Buenos Aires, Argentina, 1888; Volume 25.
- Anahosur, K.H. Ascomycetes of Coorg (India) II. M. A. C. S. Research Institute, Poona-4 (India). Sydowia 1970, 24, 177–182. [Google Scholar]
- New York State Museum and University of the State of New York. Annual Report of the Regents; University of the State of New York: Albany, NY, USA, 1893. [Google Scholar]
- Desai, B.G.; Pathak, V.N. A new Tryblidiella from India. Sydowia 1970, 24, 198–200. [Google Scholar]
- Sociedad Científica Argentina and Congreso Científico Latino Americano and Congreso Científico Internacional Americano. Anales de la Sociedad Científica Argentina; Buenos Aires, [Sociedad Científica Argentina]: Buenos Aires, Argentina, 1881; t.12.






{kind=link}
{kind=link}
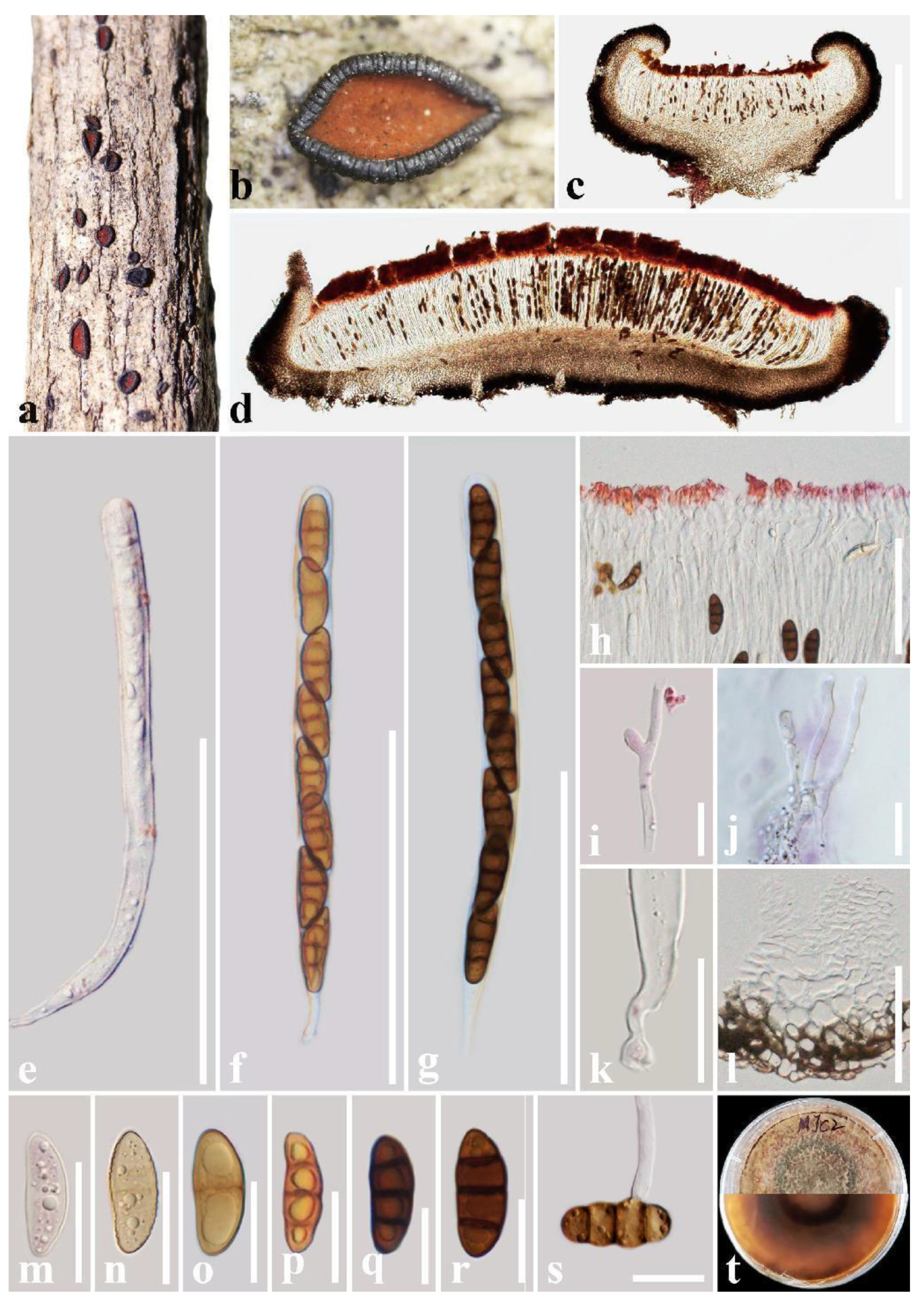{kind=link}
{kind=link}
{kind=link}
| Taxa Name | Strain Number | GenBank Accession Numbers | References | |||
|---|---|---|---|---|---|---|
| LSU | ITS | SSU | TEF | |||
| Gloniopsis calami | MFLUCC 15-0739 | NG059715 | KX669036 | KX669034 | KX671965 | [46] |
| G. praelonga | CBS 112415 | FJ161173 | — | FJ161134 | FJ161090 | [13] |
| Rhytidhysteron bannaense | KUMCC 21-0482 T | OP526408 | OP526398 | OP526395 | OP572199 | This study |
| R. bannaense | KUMCC 21-0483 | OP526409 | OP526399 | OP526396 | OP572200 | This study |
| R. bruguierae | KUMCC 21-0484 | OP442285 | OP494090 | OP482277 | OP572207 | This study |
| R. bruguierae | MFLU 18-0571 T | MN017833 | — | MN017901 | MN077056 | [47] |
| R. bruguierae | MFLUCC 17-1515 | MN632452 | MN632457 | MN632463 | MN635661 | [48] |
| R. bruguierae | MFLUCC 17-1511 | MN632454 | MN632459 | MN632465 | — | [48] |
| R. bruguierae | MFLUCC 17-1502 | MN632453 | MN632458 | MN632464 | MN635662 | [48] |
| R. bruguierae | MFLUCC 17-1509 | MN632455 | MN632460 | MN632466 | — | [48] |
| R. camporesii | KUMCC 21-0488 | OP482286 | OP494091 | OP482278 | OP572208 | This study |
| R. camporesii | KUN-HKAS 104277 T | MN429072 | MN429069 | — | MN442087 | [49] |
| R. chromolaenae | MFLUCC 17-1516 T | MN632456 | MN632461 | MN632467 | MN635663 | [48] |
| R. coffeae | KUMCC 21-0489 T | OP526406 | OP605963 | OP526412 | OP572201 | This study |
| R. coffeae | KUMCC 21-0492 | OP526407 | OP605964 | OP526413 | OP572202 | This study |
| R. cozumelense | A. Cobos-Villagrán 951 | MW939459 | MZ056797 | — | MZ457338 | [24] |
| R. cozumelense | T. Raymundo 7321 | MW939460 | MZ056798 | — | MZ457339 | [24] |
| R. erioi | MFLU 16-0584 T | MN429071 | MN429068 | — | MN442086 | [49] |
| R. esperanzae | T. Raymundo 6579 | MZ477203 | MZ056795 | — | MZ457336 | [24] |
| R. esperanzae | R. Valenzuela 17206 | MZ477204 | MZ056796 | — | MZ457337 | [24] |
| R. hongheense | KUMCC 21-0487 | OP482287 | OP494092 | OP482279 | OP572209 | This study |
| R. hongheense | KUMCC 20-0222 | MW264193 | MW264214 | MW264223 | MW256815 | [25] |
| R. hongheense | HKAS112348 | MW541820 | MW541824 | MW541831 | MW556132 | [25] |
| R. hongheense | HKAS112349 | MW541821 | MW541825 | MW541832 | MW556133 | [25] |
| R. hysterinum | EB 0351 | GU397350 | — | — | GU397340 | [13] |
| R. hysterinum | CBS 316.71 | MH871912 | MH860141 | — | — | [50] |
| R. magnoliae | KUMCC 21-0478 | OP482288 | OP494093 | OP482280 | OP572210 | This study |
| R. magnoliae | MFLUCC 18-0719 T | MN989384 | MN989383 | MN989382 | MN997309 | [6] |
| R. mangrovei | MFLU 18-1894 T | MK357777 | MK425188 | — | MK450030 | [12] |
| R. mengziense | KUMCC 21-0490 T | OP526396 | OP526402 | OP526414 | OP572203 | This study |
| R. mengziense | KUMCC 21-0491 | OP526397 | OP526403 | OP526415 | OP572204 | This study |
| R. mesophilum | A. Trejo 74 | MW939461 | MZ056799 | — | MZ457340 | [24] |
| R. mesophilum | A. Cobos-Villagrán 1800 | MW939462 | MZ056800 | — | MZ457341 | [24] |
| R. mexicanum | RV17107.1 T | MT626028 | MT626026 | — | — | [51] |
| R. mexicanum | RV17107.2 | MT626029 | MT626027 | — | — | [51] |
| R. neorufulum | KUMCC 21-0480 | OP482290 | OP494095 | OP482282 | OP572212 | This study |
| R. neorufulum | MFLUCC 21-0035 | MZ346015 | MZ346020 | MZ346025 | MZ356249 | [18] |
| R. neorufulum | MFLUCC 13-0216 T | KU377566 | KU377561 | KU377571 | KU510400 | [16] |
| R. neorufulum | GKM 361A | GQ221893 | — | GU296192 | — | [13] |
| R. neorufulum | HUEFS 192194 | KF914915 | — | — | — | [15] |
| R. neorufulum | MFLUCC 12-0528 | KJ418117 | KJ418118 | KJ418119 | — | [16] |
| R. neorufulum | CBS 306.38 | FJ469672 | — | GU296191 | GU349031 | [15] |
| R. neorufulum | MFLUCC 12-0011 | KJ418109 | KJ206287 | KJ418110 | — | [16] |
| R. neorufulum | MFLUCC 12-0567 | KJ526126 | KJ546124 | KJ546129 | — | [16] |
| R. neorufulum | MFLUCC 12-0569 | KJ526128 | KJ546126 | KJ546131 | — | [16] |
| R. neorufulum | EB 0381 | GU397351 | — | GU397366 | — | [13] |
| R. opuntiae | GKM 1190 | GQ221892 | — | — | GU397341 | [52] |
| R. rufulum | MFLUCC 14-0577 T | KU377565 | KU377560 | KU377570 | KU510399 | [16] |
| R. rufulum | EB 0384 | GU397354 | — | GU397368 | — | [13] |
| R. rufulum | EB 0382 | GU397352 | — | — | — | [13] |
| R. rufulum | EB 0383 | GU397353 | — | GU397367 | — | [13] |
| R. rufulum | MFLUCC 12-0013 | KJ418111 | KJ418112 | KJ418113 | — | [6] |
| R. tectonae | KUMCC 21-0479 | OP482291 | OP494096 | OP482283 | OP572213 | This study |
| R. tectonae | MFLUCC 21-0037 | MZ346013 | MZ346018 | MZ346023 | MZ356247 | [18] |
| R. tectonae | MFLUCC 21-0034 | MZ346014 | MZ346019 | MZ346024 | MZ356248 | [18] |
| R. tectonae | MFLUCC 13-0710 T | KU764698 | KU144936 | KU712457 | KU872760 | [53] |
| R. thailandicum | KUMCC 21-0493 | OP482292 | OP494097 | OP482284 | OP572214 | This study |
| R. thailandicum | MFLUCC 14-0503 T | KU377564 | KU377559 | KU377569 | KU497490 | [16] |
| R. thailandicum | MFLUCC 12-0530 | KJ526125 | KJ546123 | KJ546128 | — | [16] |
| R. thailandicum | MFLU17-0788 | MT093472 | MT093733 | MT093495 | — | [6] |
| R. thailandicum | MFLUCC 13-0051 | MN509434 | MN509433 | — | MN509435 | [54] |
| R. xiaokongense | KUMCC 20-0158 | MZ346011 | MZ346016 | MZ346021 | MZ356245 | [18] |
| R. xiaokongense | KUMCC 20-0160 T | MZ346012 | MZ346017 | MZ346022 | MZ356246 | [18] |
| R. yunnanense | KUMCC 21-0485 T | OP526404 | OP526410 | OP526400 | OP572205 | This study |
| R. yunnanense | KUMCC 21-0486 | OP526405 | OP526411 | OP526401 | OP572206 | This study |
| Closest Known Species | Strain Number | LSU Gene | ITS Gene | SSU Gene | TEF Gene | References |
|---|---|---|---|---|---|---|
| R. thailandicum | MFLUCC 14-0503 T | 0.00% | 1.79% (9/503 bp) | 0.00% | 2.26% (21/929 bp) | [16] |
| MFLUCC 12-0530 | 0.00% | 1.89% (10/527 bp) | 0.00% | — | [16] | |
| MFLU 17-0788 | 0.00% | 1.42% (7/494 bp) | 0.00% | — | [6] | |
| MFLUCC 13-0051 | 0.00% | 1.81% (9/496 bp) | — | 2.28% (21/923 bp) | [54] |
| Species | Ascomata/ Conidiomata | Exciples/Conidiomata Wall | Hamathecium/ Paraphysoids | Asci/ Conidiogenous Cells | Ascospores/ Conidia | Sequences | Hosts | Countries | References |
|---|---|---|---|---|---|---|---|---|---|
| Holomorph | |||||||||
| R. hysterinum | Sexual: ascomata 1000–3000 µm long × 500 µm wide × 500–1000 µm high, smooth-striated, erumpent, solitary or aggregated, sessile, deep longitudinal slit extending the entire length of the ascoma and with irregularly spaced, pseudoepithecium, orange or black when fresh and when dry | Exciples tightly compact, hyaline to light brown, becoming red in Melzer’s reagent, cells toward the interior less heavily pigmented | Paraphyses exceeding the ascus by ca. 25 µm, branching dichotomously just below the tip, tip cells globose to clavate, disintegrating and embedded in an amorphous substance to form the pseudoepithecium, becoming blue-green in Melzer’s reagent | Asci 185–220 µm × 15–17 µm, 4–8-spored, cylindrical, pedicellate | Ascospores 21–32 µm × 8–12 µm, 1-septate, septum median, fusiform with rounded to acute ends, slightly constricted at the septum, brown and translucent to nearly black, and opaque with septum obscured | ITS, LSU, TEF | Buxus spp., Diospyros sp., Ilex sp., Prosopis sp. | Australia, France, India, USA | [13,21] |
| Asexual (Aposphaeria-like): pycnidia produced abundantly in the aerialmycelium and on the surface of the agar on both MEA and PDA, non-stromatic, globose with a short papilla, 250–330 µm high × 220–250 µm wide, black | — | — | Phialides forming in a single layer over the entire inner surface of the pycnidial wall, hyaline, ampulliform to cylindrical, 7–8 µm long, × 2 µm wide basally, and 1.5 µm wide at the opening | Conidia globose, 2–3.5 µm diam., smooth, held in hyaline slime at the pycnidial opening | |||||
| Asexual (Diplodia-like): pycnidia form on MEA within one month, immersed, non-stromatic, subglobose, 350 µm high × 400 µm wide, non-papillate, black, smooth | Pycnidial wall 25–35 µm wide, consisting of pseudoparenchymatous cells, 5–7 µm × 3–4 µm, thin-walled, brown | Paraphyses arisingfrom among conidiogenous cells, up to 50 µm long × 3 µm wide, septate, unbranched, with rounded ends, hyaline | Conidiogenous cells forming in a single layer over the entire inner surface of the pycnidial wall, barely distinguishable from cells of the wall; consisting of a basal cell 6–7 µm across, and a 5–10 µm long elongation | Conidia arising holoblastically from the tip of the conidiogenous cell; at first hyaline and unicellular, becoming dark brown, opaque, minutely punctate, and 1-septate with a pore in the middle of the septum, oblong, with a truncate, non-cicatrized base, 22–26 µm × 9–11 µm | |||||
| R. rufulum | Sexual: ascomata 900–2350 µm long × 1134–1450 µm wide × 461–820 µm high, superficial, rough-striated, black or red at the center | Exciples 75–228 µm wide, two layers. Outer layer dark brown to black, cells of textura angularis or textura globosa. Inner layer hyaline cells of textura angularis to textura prismatica | Septate, branched pseudoparaphyses | Asci 150–250 µm × 11–16 µm, 8-spored | Ascospores 28–36 µm × 9–13 µm, 1–3-septate, reddish brown to brown when mature | ITS, LSU, SSU, TEF | Abrus precatorius, Abrus pulchellus, Acacia auriculiformis, Acacia cochliacantha, Acacia farnesiana, Acacia macracantha, Acacia spp., Adhatoda vasica, Albizia lebbeck, Albizia odoratissima, Alphitonia excelsa, Annona muricata, Bignonia unguis, Bougainvillea glabra, Capparis sepiaria, Casuarina sp., Celtis pallida, Citrus aurantifolia, Citrus aurantium, Codiaeum variegatum, Euterpe oleracea, Grevillea robusta, Guaiacum officinale, Helietta parvifolia, Juniperus lucayana, Nothofagus sp., Pisonia aculeata, Pithecellobium dulce, Prosopis juliflora, Torresia cearensis | Argentina, Australia, Brazil, China, Cook Islands, Costa Rica, Cuba, Dominica, France, Ghana, India, Jamaica, Japan, Kenya, Malaysia, Mexico, Micronesia, New Guinea, New Zealand, Philippine, Puerto, Rico, Spain, Tanzania, Thailand, Tonga, United States, Venezuela, West Indies | [15,16,21,55,58,59,60,61,62,63,64,65,66,67,68,69,70] |
| Asexual (Aposphaeria-like): pycnidia form abundantly in aerial mycelium, often associated with small tufts of red-brown hyphae, non-stromatic, globose to oblong, 100–150 µm high × 70–150 µm wide, non-papillate, black | — | — | Phialides forming in a single layer over the entire inner surface of the pycnidial wall, hyaline, ampulliform to cylindrical, 4.5–9.0 µm long × 1.5–3.0 µm diam. basally, and 1.5 µm wide at the opening | Conidia globose to elliptic, 2–3 µm diam. or 3.0 µm × 2.5 µm, smooth, held in a drop of hyaline slime at the pycnidial opening | |||||
| Asexual (Diplodia-like): pycnidia abundant to rare, immersed, non-stromatic, subglobose, 460 µm high × 400 µm wide, papillate or non-papillate, or seated on the surface of the agar, pyriform, 270 µm high × 130–180 µm wide and with a papilla 130–180 µm long × 70 µm wide, black, smooth | Pycnidial wall 45 µm wide, consisting of pseudoparenchymatous cells 8–20 µm × 8–10 µm, thin-walled, brown | Paraphyses arising from among conidiogenous cells, up to 50 µm long × 3 µm wide, septate, unbranched, with rounded ends, hyaline | Conidiogenous cells forming in a single layer over the entire inner surface of the pycnidial wall, consisting of a hyaline, globose cell 4–5 µm in diam. basally, and with a 5 µm long elongation | Conidia arising holoblastically from the tip of the elongation of the conidiogenous cell, at first hyaline and unicellular, becoming dark brown to opaque and 1-septate with a pore in the middle of the septum following discharge, oblong with a truncate, non-cicatrized base, 19.5–23.5 (–29.5) µm × (6.5–) 8–10 (–12) µm, smooth | |||||
| R. thailandicum | Sexual: ascomata 700–1200 µm long× 530–750 µm wide × 360–640 µm high, semi-immersed to superficial, rough without striations | Exciples 72–130 μm wide, brown to dark brown, thick-walled cells of textura angularis, becoming hyaline towards the inner layers and base | Septate, branched pseudoparaphyses, forming a yellow epithecium above asci when mounted in water | Asci 135–160 µm × 10.5–15 µm, (3–)6–8-spored | Ascospores 20–31 µm × 7.5–12 µm, (1–)3-septate, yellowish to brown when mature | ITS, LSU, SSU, TEF | Acacia sp., Aquilaria sinensis, Morus australis | China, Mexico, Thailand | [6,16,55], this study |
| Asexual (Aposphaeria-like): conidiomata 70–108 µm long × 63–110 µm wide, superficial on PDA, globose, black, appearing in a mycelium mass | Conidiomata wall thin, arranged in textura angularis | — | Conidiophores reduced to conidiogenous cells. Conidiogenous cells 5.9 µm × 3 µm, cylindrical to subcylindrical, truncate apex, short, smooth, hyaline | Conidia 2.9 µm × 2.2 µm, globose to subglobose, hyaline, smooth | |||||
| Asexual morph | |||||||||
| R. xiaokongense | Asexual (Diplodia-like): conidiomata 448–464 µm long × 324–422 µm wide, solitary, scattered, semi-immersed in the host, black, unilocular, subglobose to ampulliform. Ostioles 178–227 µm long × 166–234 µm wide, central, short papillate | Conidiomata wall 30–40 µm thick, 4–6 layers, reddish brown to dark brown cells of textura angularis | — | Conidiogenous cells 5–8 µm × 3–6 µm, subglobose or ellipsoidal, hyaline, smooth, discrete, producing a single conidium at the apex | Conidia 20–25 µm × 8–10 µm, 1-septate and brown to dark brown at maturity, oblong to ellipsoidal, straight to slightly curved, with granular appearance | ITS, LSU, SSU, TEF | Prunus sp. | China | [18] |
| Sexual morph | |||||||||
| R. bannaense | 1350 µm long × 750 µm wide × 670 µm high, rough, solitary to aggregated, semi-immersed to superficial, perpendicular striae, green at the center | 40–150 µm wide, composed of dark brown, thick-walled cells of textura angularis, outer layer brown to dark brown, inner layer pale brown to hyaline | Septate, branched, cellular pseudoparaphyses, forming an orange epithecium above asci when mounted in water | 166 µm × 14 µm, 8-spored, J- apical ring | 25 µm × 11.5 µm, 3-septate, brown to dark brown when mature | ITS, LSU, SSU, TEF | Buddleja officinalis | China | This study |
| R. beccarianum | 1000 µm long, erumpent, solitary, dark brown | — | — | 60 µm × 6 µm, 8-spored | 12–15 µm × 5–6 µm, 3-septate, constriction at the septa | — | — | Sri Lanka | [71] |
| R. brasiliense | Erumpent to nearly superficial | — | Septate, branched pseudoparaphyses | 230–250 µm × 20–30 µm, 8-spored | 40–45 µm × 15–20 µm, 3-septate | — | On rotten branches | Brazil, Thailand | [16,19] |
| R. bruguierae | 400–950 µm long × 548–570 µm wide × 410–520 µm high, superficial, striated | 148–162 µm wide, dark brown to black, thick-walled cells of textura angularis | Septate, branched pseudoparaphyses, forming a red epithecium above asci when mounted in water | 128–148 µm × 10–14 µm, 6–8-spored, J- apical ring | 14–26 µm × 6.2–9 µm, 1–3-septate, yellowish-brown to reddish brown when mature | ITS, LSU, SSU, TEF | Alnus nepalensis, Bruguiera sp., Chromolaena odorata | China, Thailand | [47,48], this study |
| R. camporesii | 800–1100 µm long × 500–650 µm high, erumpent, slightly dentate | Ectal excipulum 65–95 µm wide, blackish cells of textura globulosa to angularis. Medullary excipulum 19–22 µm wide, thin-walled, hyaline to brown cells of textura porrecta | Paraphyses septate, branched at the base, forming an orange-red epithecium above asci when mounted in water | 165–175 µm × 13–15 µm, 8-spored, J- apical ring | 25–28 µm × 9–11 µm, 3-septate, dark brown when mature | ITS, LSU, SSU, TEF | Cotoneaster franchetii | China | [49], this study |
| R. chromolaenae | 500–1000 µm long × 250–500 µm high, superficial, not perpendicular striae, scattered, dark brown to black with dark orange at the center | 45–60(–110) µm wide, hyaline or pale brown to brown cells arranged in textura globulosa to textura angularis | Septate, branched pseudoparaphyses | 120–140 µm × 10–15 µm, 8-spored | 18–22 µm × 7–9 µm, 3-septate, pale brown to brown when mature | ITS, LSU, SSU, TEF | Chromolaena odorata | Thailand | [48] |
| R. coffeae | 1520 µm long × 1120 µm wide × 450 µm high, rough, solitary to aggregated, mostly solitary, superficial, perpendicular striae, reddish brown at the center | 70–160 µm wide, composed of dark brown, thick-walled cells of textura angularis, outer layer brown to dark brown, inner layer pale brown to hyaline | Septate, branched, cellular pseudoparaphyses, forming a red to purple epithecium above asci when mounted in water | 179.5 µm × 13 µm, 8-spored, J- apical ring | 26 µm × 10 µm, 3-septate, reddish brown to brown when mature | ITS, LSU, SSU, TEF | Coffea sp. | China | This study |
| R. columbiense | 1500–3000 µm long × 1200–1800 µm wide × 600–700 µm high, superficial, striated, yellowish green on the margins | 60–90 µm wide, dark brown to black, thick-walled cells of textura angularis | Septate, branched pseudoparaphyses | 175–190 µm × 14–18 µm, 6–8-spored | 38–52 µm × 13–18 µm, (1–)3-septate, reddish brown when mature | — | Unidentified woody | Colombia | [27] |
| R. cozumelense | 2500–3500 µm long × 1100–1500 µm wide × 800–1900 µm high, erumpent, solitary, smooth to slightly striated, dark at the center | Two layers, the first carbonaceous, 45–100 µm wide thick cells of textura prismatica. The second cells hyaline, thin-walled | Septate pseudoparaphyses | 182–191 µm × 12–13 µm, 8-spored | 26–29 µm × 9–11 µm, 3-septate, dark brown when mature | ITS, LSU, TEF | Tabebuia rosea | Mexico | [24] |
| R. discolor | 1000–2000 µm long, cracking after maturity | Carbonaceous | Paraphyses filiform | 180–220 µm × 12–15 µm, 8-spored | 28–30 µm × 10–12 µm, 3-septate, elongated ellipse, guttules | — | Celtis tala | Argentina | [72] |
| R. erioi | 600–1200 µm long × 270–360 µm high, superficial or slightly erumpent, dentate | Ectal excipulum 55–75 µm wide, thin-walled, dark brown cells of textura angularis to textura globulosa. Medullary excipulum 14–20 µm wide, hyaline cells of textura porrecta | Paraphyses septate, slightly branched at the base | 140–200 µm × 9–16 µm, 8-spored, J- apical ring | 22–28 µm × 9–11 µm, 3-septate, dark brown when mature | ITS, LSU, TEF | Unidentified wood | Thailand | [49] |
| R. esperanzae | 2000–4500 µm long × 1200–3000 µm wide × 1000–2400 µm high, superficial, solitary, rarely gregarious, margin greyish green, striated, dark green to black at the center | Exciple in two layers, the first carbonaceous, 60–220 µm wide cells of textura globulosa-angularis. The second slightly pigmented to hyaline, thin-walled | Septate pseudoparaphyses | 265–270 µm × 19–20 µm, 8-spored | 45–47 µm × 17–19 µm, 3-septate, reddish brown to brown when mature | ITS, LSU, TEF | Oreomunnea mexicana | Mexico | [24] |
| R. guaraniticum | 1000–4000 µm long × 700–100 µm wide, superficial | — | — | 200 µm × 12–14 µm | 30–31 µm × 10–12 µm, 3-septate | — | On bark, rotten branches | Jawa, Paraguay | [73] |
| R. hongheense | 1200–2000 µm long × 600–1000 µm wide × 350–500 µm high, slightly erumpent, slightly dentate | Ectal excipulum 70–100 µm wide, thick-walled, with black cells of textura globulosa to textura angularis. Medullary excipulum is composed of narrow, long, thin-walled, hyaline to brown cells of textura angularis | Septate, branched pseudoparaphyses, forming a red epithecium above asci when mounted in water | 140–180 µm × 12–16 µm, 8-spored | 20–33 µm × 9–13 µm, 3-septate, dark brown when mature, rarely muriform, with one longitudinal septum | ITS, LSU, SSU, TEF, RPB2 | Dodonaea sp., Phyllanthus emblica | China | [25], this study |
| R. indicum | 1800–3000 µm long, black, carbonaceous, scattered, erumpent, uniloculate, discoid to elongated | — | Paraphyses filiform, septate, clavate expansion | 200–220 µm × 18–20 µm, 8-spored | 30–32 µm × 12–14 µm, dark brown, 3-septate, end cells slightly tapering, constricted at septa, uniseriate | — | Scutia indica | India | [74] |
| R. magnoliae | 1200–2300 µm long × 540–600 µm wide × 430–550 µm high µm semi-immersed to superficial, striated, dark brown at the center | 80–100 µm wide, two layers. Outer layer black to dark brown, thick-walled cells of textura angularis. Inner layer hyaline, thin-walled cells of textura angularis to textura prismatica | Septate, branched pseudoparaphyses, forming an orange epithecium above asci when mounted in water | 160–200 µm × 14 µm, 8-spored | 25–32 µm × 8–12 µm, 1–3-septate, pale brown to dark brown when mature | ITS, LSU, SSU, TEF | Hevea brasiliensis,Magnolia grandiflora | China | [6], this study |
| R. mangrovei | 930–1980 µm long × 780–910 µm wide × 500–520 µm high, crowded to aggregate, semi-immersed to superficial, rough-striated | 65–90 µm wide, dark brown to black, thin-walled cells of textura angularis | Septate, unbranched pseudoparaphyses | 110–150 µm × 9.4–10 µm, (2–6–) 8-spored | 21–28 µm × 7.5–8.5 µm, 1–3-septate, reddish brown when mature | ITS, LSU, TEF | Mangrove sp. | Thailand | [12] |
| R. mengziense | 1400 µm long × 910 µm wide × 640 µm high, smooth, solitary to aggregated, mostly solitary, semi-immersed to superficial, perpendicular striae, reddish brown at the center | 60–135 µm wide, composed of outer layer brown to black, thick-walled cells of textura angularis and inner layer light brown, thin-walled cells of textura prismatica | Septate, branched, cellular pseudoparaphyses, forming a reddish brown to brown epithecium above asci when mounted in water | 164.5 µm × 13 µm, 8-spored, J- apical ring | 27 µm × 12 µm, 3-septate, reddish brown to brown when mature | ITS, LSU, SSU, TEF | Crataegus scabrifolia | China | This study |
| R. mesophilum | 2500–4000 µm long × 1000–1500 µm wide × 1400–1700 µm high, superficial or erumpent, gregarious, rarely solitary, margin yellowish green, striated, orange at the center | Two layers, the first carbonaceous, 62.5–75 µm wide, green yellowish cells of textura prismatica. The second hyaline, thin-walled | Aseptate, branched pseudoparaphyses | 267–282 µm × 15.5–16 µm, 8-spored | 40–44 µm × 12–14 µm, 3-septate, light brown when mature | ITS, LSU, TEF | On decayed wood | Mexico | [24] |
| R. mexicanum | 2000–4000 µm long × 1500–2500 µm wide × 1500 µm high, superficial or erumpent, gregarious, rarely solitary, striated | Two layers, the first carbonaceous, 104.5–114 µm wide in the medium cells of textura globulosa to textura angularis, thick-walled. The second composed of cells of textura prismatica, hyaline, thin-walled | Aseptate, bifurcated to branched pseudoparaphyses | 285–297 µm × 16–17 µm, 8-spored | 34–40 µm × 10–12 µm, 3-septate, reddish brown when mature | ITS, LSU | — | Mexico | [51] |
| R. neohysterinum | 1500–2500 µm long × 700–2200 µm wide × 700–1100 µm high, superficial, solitary, rarely gregarious, striated, orange at the center | 52–68 µm wide, dark brown to black, thick-walled cells of textura prismatica | Septate pseudoparaphyses | 160–185 µm × 12–13 µm, 8-spored | 24.8–29 µm × 8.8–10 µm, 1-septate, brown when mature | — | Acacia sp. | Mexico | [55] |
| R. neorufulum | 835–2100 µm long × 350–1320 µm wide × 430–1000 µm high, superficial, elliptic or irregular, without striations, black or yellow at the center | 64–160 µm wide, dark brown to black, thick-walled cells of textura angularis | Septate, branched pseudoparaphyses, forming a yellow epithecium above asci when mounted in water | 185–260 µm × 9.5–18 µm, 8-spored | 27–44 µm × 6.5–17 µm, 1–3-septate, reddish brown to brown when mature | ITS, LSU, SSU, TEF | Bursera sp., Elaeagnus sarmentosa, Hevea brasiliensis, Tectona grandis | China, Mexico, Thailand | [16,18,55], this study |
| R. opuntiae | 640–1700 µm long | — | — | 85–160 µm × 12.5–16 µm, 3–8-spored | 17–33 µm × 13 µm, 3–5-septate | LSU, TEF | Opuntia fulgida | USA | [52] |
| R. prosopidis | Superficial, very hard when dry, elliptical or triangular, black, with very obtuse, thick, yellowish green disc | — | — | — | 3-septate, unineriate, oblong, sometimes slightly curved | — | Prosopis juliflora | USA | [75] |
| R. quercinum | 1000–3000 µm in diam., leathery apothecia, scattered, superficial, erumpent, pedicellate (short pedicel) | Excipulum black with low seated, reddish | — | Asci cylindrical, slender, stalked hyaline with inconspicuous wall | 19.0–24.7 µm × 7.6–11.4 µm, 1–3-septate more commonly 3 | — | Quercus sp. | India | [76] |
| R. tectonae | 550–3365 µm long × 325–728 µm wide × 370−835 µm high, semi-immersed to superficial, smooth without striation, yellow at the center | 80–135 µm wide, two layers. Outer layer black to dark reddish, thick-walled cells of textura angularis. Inner layer hyaline, thin-walled cells of textura angularis | Septate, branched pseudoparaphyses, forming an orange epithecium above asci when mounted in water | 150–200 µm × 10–15 µm, 8-spored | 19–31 µm × 8–13 µm, 1–3-septate, pale brown to dark brown when mature | ITS, LSU, SSU, TEF | Betula sp., Fabaceae sp., Magnolia delavayi, Tectona grandis | China, Thailand | [18,53], this study |
| R. viride | 1000–1500 µm long × 500–600 µm wide, erumpent | — | Filiform, hyaline | 200–250 µm × 10–12 µm | 20–30 µm × 7–9 µm, 3-septate | — | On bark, associated with lichens | Brazil | [77] |
| R. yunnanense | 2510 µm × 625 µm × 455 µm, solitary to aggregated, mostly aggregated, semi-immersed, each hysterothecia has two parallel striae parallel to the longitudinal slit and slight perpendicular striae, longitudinal slit, no opening | 60–180 µm wide, composed of dark brown, thick-walled cells of textura globulosa, outer layer brown to dark brown, inner layer pale brown to hyaline | Septate, branched, cellular pseudoparaphyses, forming a yellow epithecium above asci when mounted in water | 230 µm × 14 µm, 8-spored, J- apical ring | 32.5 µm × 13 µm, 3-septate, reddish brown to brown when mature | ITS, LSU, SSU, TEF | Rhus chinensis | China | This study |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Du, T.-Y.; Dai, D.-Q.; Mapook, A.; Lu, L.; Stephenson, S.L.; Suwannarach, N.; Elgorban, A.M.; Al-Rejaie, S.; Karunarathna, S.C.; Tibpromma, S. Additions to Rhytidhysteron (Hysteriales, Dothideomycetes) in China. J. Fungi 2023, 9, 148. https://doi.org/10.3390/jof9020148
Du T-Y, Dai D-Q, Mapook A, Lu L, Stephenson SL, Suwannarach N, Elgorban AM, Al-Rejaie S, Karunarathna SC, Tibpromma S. Additions to Rhytidhysteron (Hysteriales, Dothideomycetes) in China. Journal of Fungi. 2023; 9(2):148. https://doi.org/10.3390/jof9020148
Chicago/Turabian StyleDu, Tian-Ye, Dong-Qin Dai, Ausana Mapook, Li Lu, Steven L. Stephenson, Nakarin Suwannarach, Abdallah M. Elgorban, Salim Al-Rejaie, Samantha C. Karunarathna, and Saowaluck Tibpromma. 2023. "Additions to Rhytidhysteron (Hysteriales, Dothideomycetes) in China" Journal of Fungi 9, no. 2: 148. https://doi.org/10.3390/jof9020148
APA StyleDu, T.-Y., Dai, D.-Q., Mapook, A., Lu, L., Stephenson, S. L., Suwannarach, N., Elgorban, A. M., Al-Rejaie, S., Karunarathna, S. C., & Tibpromma, S. (2023). Additions to Rhytidhysteron (Hysteriales, Dothideomycetes) in China. Journal of Fungi, 9(2), 148. https://doi.org/10.3390/jof9020148