
Polypeptides Targeting Paracoccidioides brasiliensis Drk1

 , ,
, ,  , , ,
, , , 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Fungal Isolates and Growth Conditions
2.2. Phage-Display Library and Biopanning
2.3. Phage Amplification
2.4. Phage Titration
2.5. DNA Extraction for Sequencing and Peptides Sequence Deduction
2.6. Reactivity of Individual Peptides with P. brasiliensis and PbDrk1 Protein Using Enzyme-Linked Immunosorbent Assay (ELISA)
2.7. Antifungal Activity of Peptides
2.8. Peptides-Antifungal Combinations
2.9. Peptides Toxicity
2.9.1. In Pneumocytes
2.9.2. In G. mellonella
2.10. Paracoccidioides Brasiliensis-Pneumocytes and P. brasiliensis-Extracellular Matrix Adhesion after Peptides Treatment Using Enzyme-Linked Immunosorbent Assay
2.11. Therapeutic Use of Peptides in G. mellonella In Vivo Model of Infection
2.12. Effects of Peptides on Hemocyte Density and Phenoloxidase Activity
2.13. Cell Wall Modifications Mediated by Exposure of Fungi to Peptides
2.14. Mycelium to Yeast Alterations in P. brasiliensis following Peptides Treatment
2.15. Statistical Analysis
3. Results
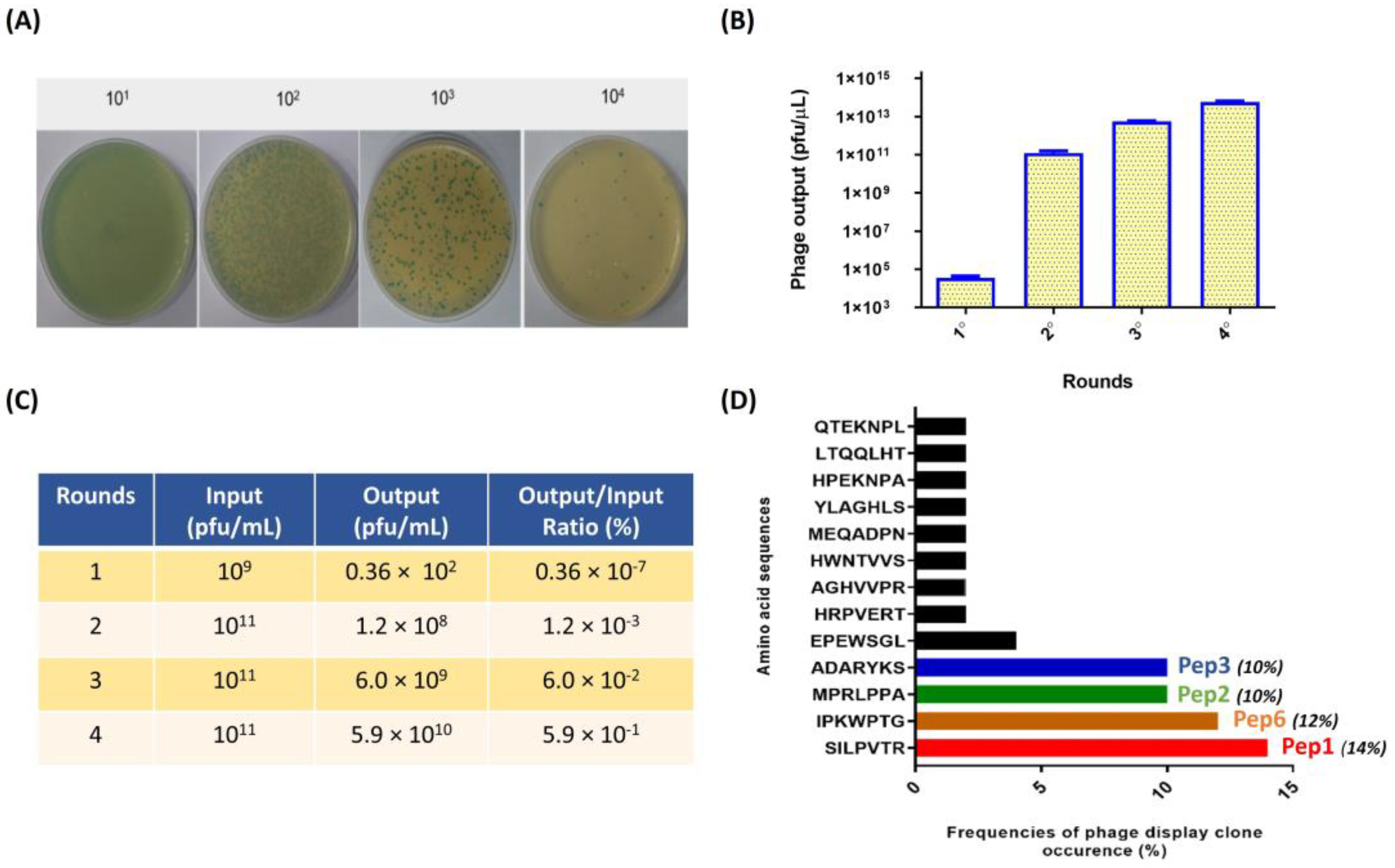
3.1. Enrichment of PbDrk1-Binding Phages and Identified Polypeptides
3.2. Binding of Polypeptides to PbDrk1 Protein and P. brasiliensis
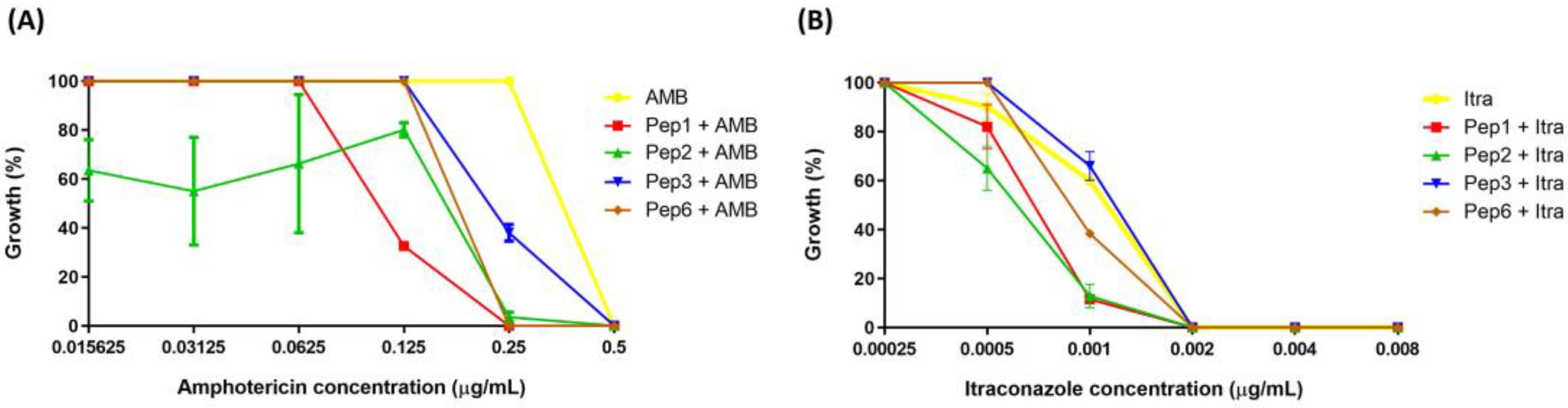
3.3. Antifungal Activity of Synthetic Polypeptide and Its Potentiation of Antifungals
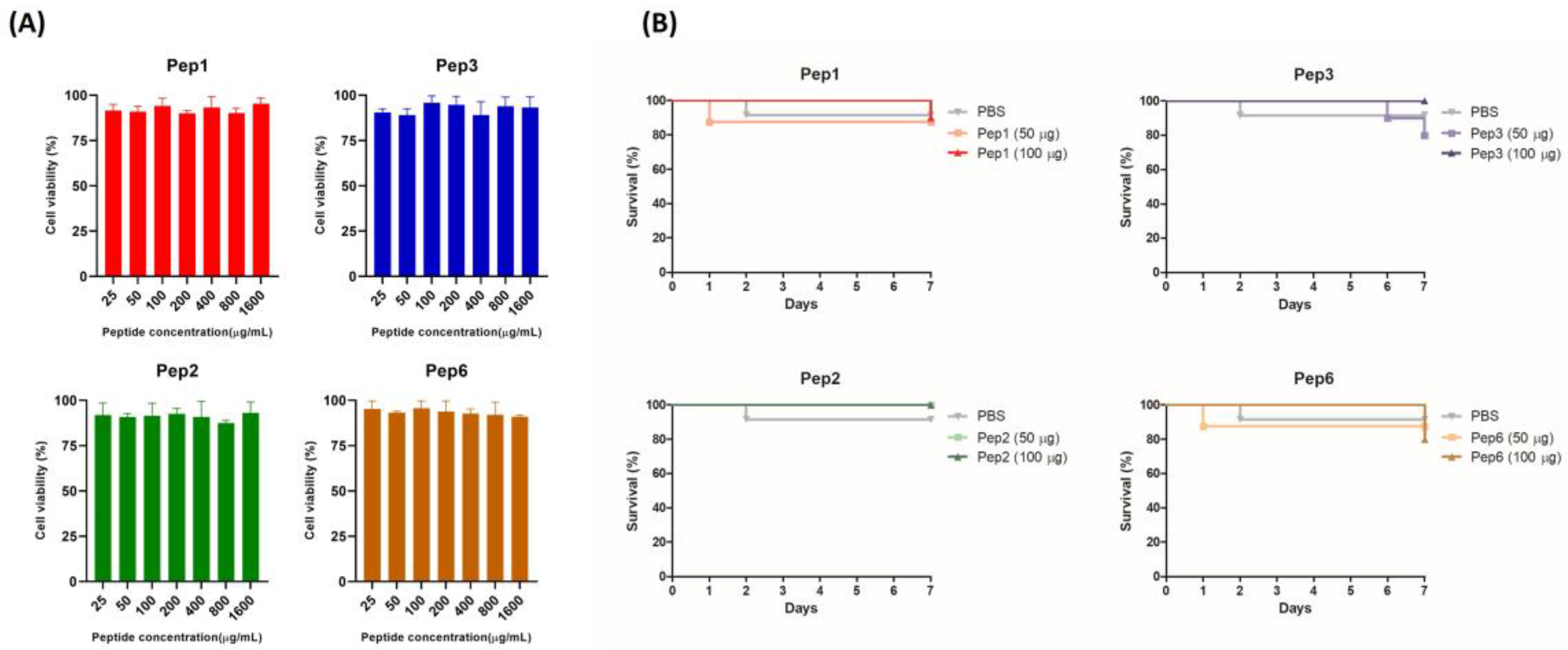
3.4. Cytotoxicity of Polypeptides in Pneumocytes A549 Cells and G. mellonella
3.5. Some Peptides against PbDrk1 Inhibited Adhesion Abilities to A549 Cells but Not to ECM
3.6. Paracoccidioides Brasiliensis-Cell Wall Modifications Mediated by Identified Peptides
3.7. Mycelium to Yeast Transition of P. brasiliensis Is Altered by Contact with Peptides
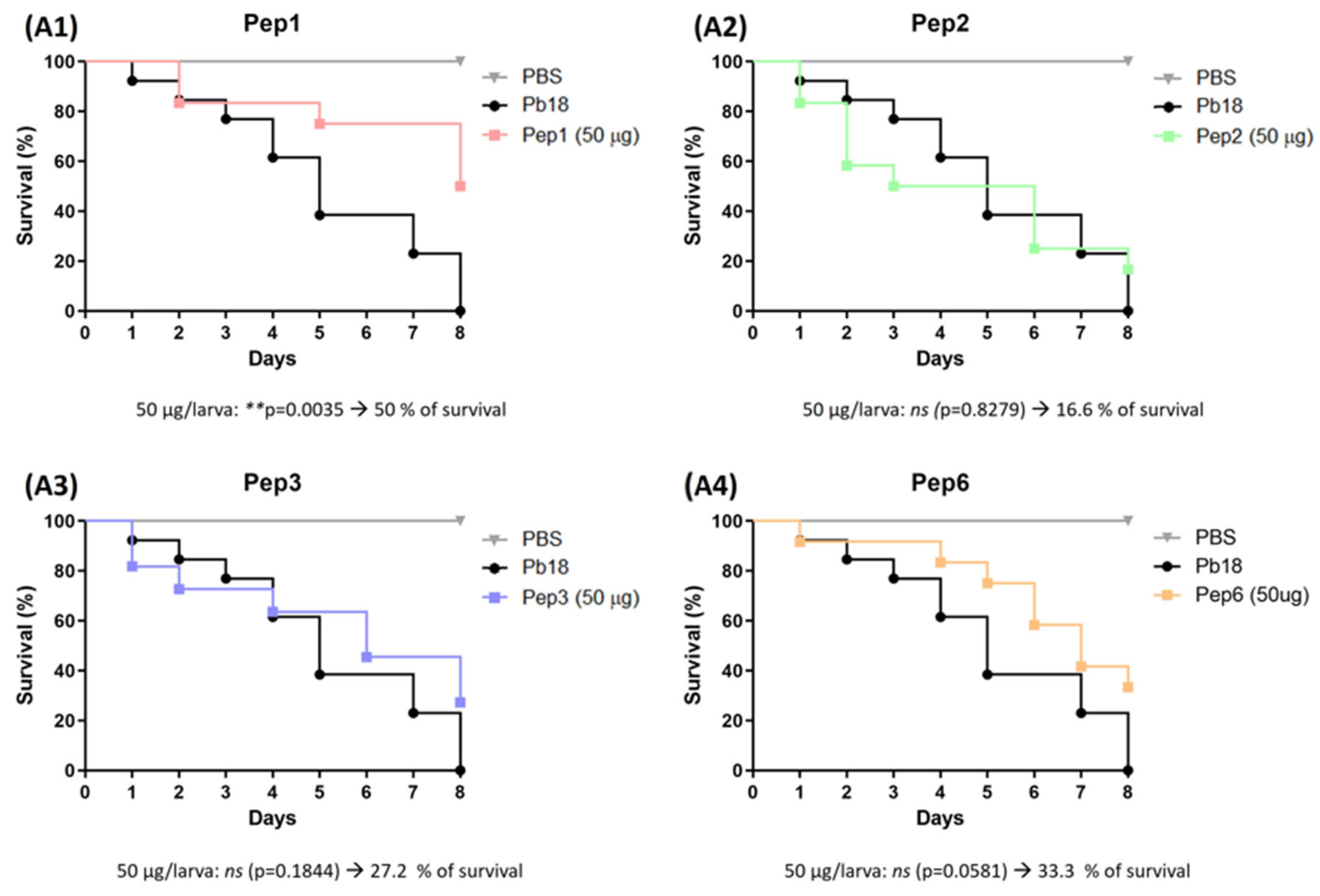
3.8. Prophylactic and Therapeutic Use of Drk1-Binding Peptides in G. mellonella Model of Infection
3.9. Effects of Peptides in Hemocyte Density and Phenoloxidase Activity of G. mellonella
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kim, J.H.; Cheng, L.W.; Land, K.M. Advances in Antifungal Development: Discovery of New Drugs and Drug Repurposing. Pharmaceuticals 2022, 15, 787. [Google Scholar] [CrossRef] [PubMed]
- De Oliveira, H.C.; Bezerra, B.T.; Rodrigues, M.L. Antifungal Development and the Urgency of Minimizing the Impact of Fungal Diseases on Public Health. ACS Bio Med. Chem. Au 2022, 3, 137–146. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Hube, B.; Kämper, J.; Meyer, V.; Krappmann, S. When green and red mycology meet: Impressions from an interdisciplinary forum on virulence mechanisms of phyto- and human-pathogenic fungi. Virulence 2017, 8, 1435–1444. [Google Scholar] [CrossRef] [PubMed]
- Borges-Walmsley, M.I.; Chen, D.; Shu, X.; Walmsley, A.R. The pathobiology of Paracoccidioides brasiliensis. Trends Microbiol. 2002, 10, 80–87. [Google Scholar] [CrossRef]
- San-Blas, G.; Niño-Vega, G.; Iturriaga, T. Paracoccidioides brasiliensis and paracoccidioidomycosis: Molecular approaches to morphogenesis, diagnosis, epidemiology, taxonomy and genetics. Med. Mycol. 2002, 40, 225–242. [Google Scholar] [CrossRef]
- Shikanai-Yasuda, M.A.; Mendes, R.P.; Colombo, A.L.; Queiroz-Telles, F.; Kono, A.S.G.; Paniago, A.M.M.; Nathan, A.; Valle, A.C.F.D.; Bagagli, E.; Benard, G.; et al. Brazilian guidelines for the clinical management of paracoccidioidomycosis. Rev. Soc. Bras. Med. Trop. 2017, 50, 715–740. [Google Scholar] [CrossRef]
- Scorzoni, L.; de Paula e Silva, A.C.A.; Marcos, C.M.; Assato, P.A.; de Melo, W.C.M.A.; de Oliveira, H.C.; Costa-Orlandi, C.B.; Mendes-Giannini, M.J.S.; Fusco-Almeida, A.M. Antifungal therapy: New advances in the understanding and treatment of mycosis. Front. Microbiol. 2017, 8, 36. [Google Scholar] [CrossRef]
- Kim, J.H.; Cheng, L.W.; Chan, K.L.; Tam, C.C.; Mahoney, N.; Friedman, M.; Shilman, M.M.; Land, K.M. Antifungal Drug Repurposing. Antibiotics 2020, 9, 812. [Google Scholar] [CrossRef]
- Ivanov, M.; Ćirić, A.; Stojković, D. Emerging Antifungal Targets and Strategies. Int. J. Mol. Sci. 2022, 23, 2756. [Google Scholar] [CrossRef]
- McCarthy, M.W.; Kontoyiannis, D.P.; Cornely, O.A.; Perfect, J.R.; Walsh, T.J. Novel Agents and Drug Targets to Meet the Challenges of Resistant Fungi. J. Infect. Dis. 2017, 216, S474–S483. [Google Scholar] [CrossRef]
- Nguyen, S.; Truong, J.Q.; Bruning, J.B. Targeting Unconventional Pathways in Pursuit of Novel Antifungals. Front. Mol. Biosci. 2020, 7, 621366. [Google Scholar] [CrossRef]
- Do Carmo Silva, L.; de Oliveira, A.A.; de Souza, D.R.; Barbosa, K.L.; Freitas e Silva, K.S.; Carvalho Júnior, M.A.; Rocha, O.B.; Lima, R.M.; Santos, T.G.; Soares, C.M.; et al. Overview of Antifungal Drugs against Paracoccidioidomycosis: How Do We Start, Where Are We, and Where Are We Going? J. Fungi 2020, 6, 300. [Google Scholar] [CrossRef] [PubMed]
- De Oliveira, H.C.; Assato, P.A.; Marcos, C.M.; Scorzoni, L.; de Paula, E.; Silva, A.C.A.; Da Silva, J.D.F.; Singulani, J.d.L.; Alarcon, K.M.; Fusco-Almeida, A.M.; et al. Paracoccidioides-host Interaction: An Overview on Recent Advances in the Paracoccidioidomycosis. Front. Microbiol. 2015, 6, 1319. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Hou, B.; Wu, Y.Z.; Wang, Y.; Liu, X.; Han, S. Two-component histidine kinase DRK1 is required for pathogenesis in Sporothrix schenckii. Mol. Med. Rep. 2018, 17, 721–728. [Google Scholar] [CrossRef]
- Hou, B.; Zhang, Z.; Zheng, F.; Liu, X. Molecular cloning, characterization and differential expression of DRK1 in Sporothrix schenckii. Int. J. Mol. Med. 2013, 31, 99–104. [Google Scholar] [CrossRef]
- Boyce, K.J.; Schreider, L.; Kirszenblat, L.; Andrianopoulos, A. The two-component histidine kinases DrkA and SlnA are required for in vivo growth in the human pathogen Penicillium marneffei. Mol. Microbiol. 2011, 82, 1164–1184. [Google Scholar] [CrossRef]
- Nemecek, J.C.; Wüthrich, M.; Klein, B.S. Global Control of Dimorphism and Virulence in Fungi. Science 2006, 312, 583–588. [Google Scholar] [CrossRef]
- Marcos, C.M.; de Oliveira, H.C.; Assato, P.A.; Castelli, R.F.; Fusco-Almeida, A.M.; Mendes-Giannini, M.J. Drk1, a Dimorphism Histidine Kinase, Contributes to Morphology, Virulence, and Stress Adaptation in Paracoccidioides brasiliensis. J. Fungi 2021, 7, 852. [Google Scholar] [CrossRef]
- Chaves, A.F.A.; Navarro, M.V.; Castilho, D.G.; Calado, J.C.P.; Conceição, P.M.; Batista, W.L. A conserved dimorphism-regulating histidine kinase controls the dimorphic switching in Paracoccidioides brasiliensis. FEMS Yeast Res. 2016, 16, fow047. [Google Scholar] [CrossRef]
- Navarro, M.V.; de Barros, Y.N.; Segura, W.D.; Chaves, A.F.; Jannuzzi, G.P.; Ferreira, K.S.; Xander, P.; Batista, W.L. The Role of Dimorphism Regulating Histidine Kinase (Drk1) in the Pathogenic Fungus Paracoccidioides brasiliensis Cell Wall. J. Fungi 2021, 7, 1014. [Google Scholar] [CrossRef]
- Capra, E.J.; Laub, M.T. Evolution of Two-Component Signal Transduction Systems. Annu. Rev. Microbiol. 2012, 66, 325–347. [Google Scholar] [CrossRef] [PubMed]
- Boyce, K.J.; Andrianopoulos, A. Fungal dimorphism: The switch from hyphae to yeast is a specialized morphogenetic adaptation allowing colonization of a host. FEMS Microbiol. Rev. 2015, 39, 797–811. [Google Scholar] [CrossRef] [PubMed]
- Yaakoub, H.; Sanchez, N.S.; Ongay-Larios, L.; Courdavault, V.; Calenda, A.; Bouchara, J.-P.; Coria, R.; Papon, N. The high osmolarity glycerol (HOG) pathway in fungi†. Crit. Rev. Microbiol. 2022, 48, 657–695. [Google Scholar] [CrossRef]
- Yaakoub, H.; Mina, S.; Calenda, A.; Bouchara, J.-P.; Papon, N. Oxidative stress response pathways in fungi. Cell. Mol. Life Sci. 2022, 79, 333. [Google Scholar] [CrossRef] [PubMed]
- Yaakoub, H.; Mina, S.; Marot, A.; Papon, N.; Calenda, A.; Bouchara, J.-P. A Stress Hub in Scedosporium apiospermum: The High Osmolarity Glycerol (HOG) Pathway. Kinases Phosphatases 2022, 1, 4–13. [Google Scholar] [CrossRef]
- Marshall, C.J. MAP kinase kinase kinase, MAP kinase kinase and MAP kinase. Curr. Opin. Genet. Dev. 1994, 4, 82–89. [Google Scholar] [CrossRef]
- Wuichet, K.; Cantwell, B.J.; Zhulin, I.B. Evolution and phyletic distribution of two-component signal transduction systems. Curr. Opin. Microbiol. 2010, 13, 219–225. [Google Scholar] [CrossRef]
- University of Wisconsin-Madison. New mechanism of action found for agricultural pesticide fludioxonil. Sci. Dly. 2019. Available online: https://www.sciencedaily.com/releases/2019/03/190325080431.htm (accessed on 8 May 2023).
- Fava Netto, C. Contribuição para o estudo imunológico da blastomicose de Lutz (Blastomicosesulamericana). Rev. Inst. A Lutz. 1961, 21, 1961. [Google Scholar]
- Strober, W. Trypan Blue Exclusion Test of Cell Viability. Curr. Protoc. Immunol. 2015, 111, A3.B.1–A3.B.3. [Google Scholar] [CrossRef]
- De Oliveira, H.C.; Michaloski, J.S.; da Silva, J.F.; Scorzoni, L.; de Paula e Silva, A.C.A.; Marcos, C.M.; Assato, P.A.; Yamazaki, D.S.; Fusco-Almeida, A.M.; Giordano, R.J.; et al. Peptides derived from a phage display library inhibit adhesion and protect the host against infection by Paracoccidioides brasiliensis and Paracoccidioides lutzii. Front. Pharmacol. 2016, 7, 509. [Google Scholar] [CrossRef]
- De Paula e Silva, A.C.A.; Oliveira, H.C.; Silva, J.F.; Sangalli-Leite, F.; Scorzoni, L.; Fusco-Almeida, A.M.; Mendes-Giannini, M.J.S. Microplate alamarBlue assay for Paracoccidioides susceptibility testing. J. Clin. Microbiol. 2013, 51, 1250–1252. [Google Scholar] [CrossRef] [PubMed]
- Scorzoni, L.; de Paula e Silva, A.C.A.; Singulani, J.d.L.; Leite, F.S.; de Oliveira, H.C.; da Silva, R.A.M.; Fusco-Almeida, A.M.; Mendes-Giannini, M.J.S. Comparison of virulence between Paracoccidioides brasiliensis and Paracoccidioides lutzii using Galleria mellonella as a host model. Virulence 2015, 6, 766–776. [Google Scholar] [CrossRef] [PubMed]
- Scorzoni, L.; Alves de Paula e Silva, A.C.; de Oliveira, H.C.; Dos Santos, C.T.; Singulani, J.L.; Assato, P.A.; Marcos, C.M.; Oliveira, L.T.; Fregonezi, N.F.; Rossi, D.C.P.; et al. In vitro and in vivo effect of peptides derived from 14-3-3 Paracoccidioides spp. Protein. J. Fungi 2021, 7, 52. [Google Scholar] [CrossRef] [PubMed]
- De Curcio, J.S.; Silva, M.G.; Silva Bailão, M.G.; Báo, S.N.; Casaletti, L.; Bailão, A.M.; de Almeida Soares, C.M. Identification of membrane proteome of Paracoccidioides lutzii and its regulation by zinc. Futur. Sci. OA 2017, 3, FSO232. [Google Scholar] [CrossRef]
- Guttentag, A.; Krishnakumar, K.; Cokcetin, N.; Hainsworth, S.; Harry, E.; Carter, D. Inhibition of Dermatophyte Fungi by Australian Jarrah Honey. Pathogens 2021, 10, 194. [Google Scholar] [CrossRef]
- De Paula, E.; Silva, A.C.A.; de Oliveira, H.C.; Scorzoni, L.; Marcos, C.M.; Dos Santos, C.T.; Fusco-Almeida, A.M.; Salina, A.C.G.; Medeiros, A.I.; Almeida, F.; et al. Decyl gallate as a possible inhibitor of N-glycosylation process in. Antimicrob. Agents Chemother. 2019, 63, 10–1128. [Google Scholar] [CrossRef]
- Nunes, L.R.; Costa de Oliveira, R.; Leite, D.B.; da Silva, V.S.; dos Reis Marques, E.; da Silva Ferreira, M.E.; Ribeiro, D.C.D.; de Souza Bernardes, L.A.; Goldman, M.H.S.; Puccia, R.; et al. Transcriptome analysis of Paracoccidioides brasiliensis cells undergoing mycelium-to-yeast transition. Eukaryot. Cell 2005, 4, 2115–2128. [Google Scholar] [CrossRef]
- Anandakumar, S.; Boosi, K.N.; Bugatha, H.; Padmanabhan, B.; Sadhale, P.P. Phage displayed short peptides against cells of Candida albicans demonstrate presence of species, morphology and region specific carbohydrate epitopes. PLoS ONE 2011, 6, e16868. [Google Scholar] [CrossRef]
- Shor, E.; Chauhan, N. A case for two-component signaling systems as antifungal drug targets. PLoS Pathog. 2015, 11, e1004632. [Google Scholar] [CrossRef]
- Chauhan, N.; Calderone, R. Two-component signal transduction proteins as potential drug targets in medically important fungi. Infect. Immun. 2008, 76, 4795–4803. [Google Scholar] [CrossRef]
- Lawry, S.M.; Tebbets, B.; Kean, I.; Stewart, D.; Hetelle, J.; Klein, B.S. Fludioxonil Induces Drk1, a Fungal Group III Hybrid Histidine Kinase, To Dephosphorylate Its Downstream Target, Ypd1. Antimicrob. Agents Chemother. 2017, 61, 10–1128. [Google Scholar] [CrossRef]
- Yoshimi, A.; Kojima, K.; Takano, Y.; Tanaka, C. Group III histidine kinase is a positive regulator of Hog1-type mitogen-activated protein kinase in filamentous fungi. Eukaryot. Cell 2005, 4, 1820–1828. [Google Scholar] [CrossRef] [PubMed]
- Jacob, S.; Foster, A.J.; Yemelin, A.; Thines, E. High osmolarity glycerol (HOG) signalling in Magnaporthe oryzae: Identification of MoYPD1 and its role in osmoregulation, fungicide action, and pathogenicity. Fungal Biol. 2015, 119, 580–594. [Google Scholar] [CrossRef] [PubMed]
- Brandhorst, T.T.; Kean, I.R.L.; Lawry, S.M.; Wiesner, D.L.; Klein, B.S. Phenylpyrrole fungicides act on triosephosphate isomerase to induce methylglyoxal stress and alter hybrid histidine kinase activity. Sci. Rep. 2019, 9, 5047. [Google Scholar] [CrossRef]
- Kim, J.H.; Chan, K.L.; Faria, N.C.G.; Martins, M.d.L.; Campbell, B.C. Targeting the oxidative stress response system of fungi with redox-potent chemosensitizing agents. Front. Microbiol. 2012, 3, 88. [Google Scholar] [CrossRef] [PubMed]
- Okamoto, Y.; Aoki, S.; Mataga, I. Enhancement of amphotericin B activity against Candida albicans by superoxide radical. Mycopathologia 2004, 158, 9–15. [Google Scholar] [CrossRef]
- González-Párraga, P.; Sánchez-Fresneda, R.; Zaragoza, O.; Argüelles, J.-C. Amphotericin B induces trehalose synthesis and simultaneously activates an antioxidant enzymatic response in Candida albicans. Biochim. Biophys. Acta 2011, 1810, 777–783. [Google Scholar] [CrossRef]
- Blum, G.; Perkhofer, S.; Haas, H.; Schrettl, M.; Würzner, R.; Dierich, M.P.; Lass-Flörl, C. Potential basis for amphotericin B resistance in Aspergillus terreus. Antimicrob. Agents Chemother. 2008, 52, 1553–1555. [Google Scholar] [CrossRef]
- Dib, L.; Hayek, P.; Sadek, H.; Beyrouthy, B.; Khalaf, R.A. The Candida albicans Ddr48 protein is essential for filamentation, stress response, and confers partial antifungal drug resistance. Med. Sci. Monit. Int. Med. J. Exp. Clin. Res. 2008, 14, BR113–BR121. [Google Scholar]
- An, M.; Shen, H.; Cao, Y.; Zhang, J.; Cai, Y.; Wang, R.; Jiang, Y. Allicin enhances the oxidative damage effect of amphotericin B against Candida albicans. Int. J. Antimicrob. Agents 2009, 33, 258–263. [Google Scholar] [CrossRef]
- Toone, W.M.; Jones, N. Stress-activated signalling pathways in yeast. Genes Cells 1998, 3, 485–498. [Google Scholar] [CrossRef] [PubMed]
- Miskei, M.; Karányi, Z.; Pócsi, I. Annotation of stress-response proteins in the aspergilli. Fungal Genet. Biol. 2009, 46 (Suppl. 1), S105–S120. [Google Scholar] [CrossRef] [PubMed]
- Brown, A.J.P. Fungal resilience and host-pathogen interactions: Future perspectives and opportunities. Parasite Immunol. 2023, 45, e12946. [Google Scholar] [CrossRef]
- Alonso-Monge, R.; Navarro-García, F.; Román, E.; Negredo, A.I.; Eisman, B.; Nombela, C.; Pla, J. The Hog1 mitogen-activated protein kinase is essential in the oxidative stress response and chlamydospore formation in Candida albicans. Eukaryot. Cell 2003, 2, 351–361. [Google Scholar] [CrossRef]
- Bernhardt, J.; Herman, D.; Sheridan, M.; Calderone, R. Adherence and invasion studies of Candida albicans strains, using in vitro models of esophageal candidiasis. J. Infect. Dis. 2001, 184, 1170–1175. [Google Scholar] [CrossRef]
- Hohmann, S. Osmotic stress signaling and osmoadaptation in yeasts. Microbiol. Mol. Biol. Rev. 2002, 66, 300–372. [Google Scholar] [CrossRef]
- Calera, J.A.; Zhao, X.J.; Calderone, R. Defective hyphal development and avirulence caused by a deletion of the SSK1 response regulator gene in Candida albicans. Infect. Immun. 2000, 68, 518–525. [Google Scholar] [CrossRef]
- Nagahashi, S.; Mio, T.; Ono, N.; Yamada-Okabe, T.; Arisawa, M.; Bussey, H.; Yamada-Okabe, H. Isolation of CaSLN1 and CaNIK1, the genes for osmosensing histidine kinase homologues, from the pathogenic fungus Candida albicans. Microbiology 1998, 144 Pt 2, 425–432. [Google Scholar] [CrossRef]
- Wurgler-Murphy, S.M.; Saito, H. Two-component signal transducers and MAPK cascades. Trends Biochem. Sci. 1997, 22, 172–176. [Google Scholar] [CrossRef]
- Liao, B.; Ye, X.; Chen, X.; Zhou, Y.; Cheng, L.; Zhou, X.; Ren, B. The two-component signal transduction system and its regulation in Candida albicans. Virulence 2021, 12, 1884–1899. [Google Scholar] [CrossRef]
- Davies, S.P.; Reddy, H.; Caivano, M.; Cohen, P. Specificity and mechanism of action of some commonly used protein kinase inhibitors. Biochem. J. 2000, 351, 95–105. [Google Scholar] [CrossRef] [PubMed]
- Bain, J.; Plater, L.; Elliott, M.; Shpiro, N.; Hastie, C.J.; Mclauchlan, H.; Klevernic, I.; Arthur, J.S.C.; Alessi, D.R.; Cohen, P. The selectivity of protein kinase inhibitors: A further update. Biochem. J. 2007, 408, 297–315. [Google Scholar] [CrossRef]
- Sestari, S.J.; Brito, W.A.; Neves, B.J.; Soares, C.M.A.; Salem-Izacc, S.M. Inhibition of protein kinase A affects Paracoccidioides lutzii dimorphism. Int. J. Biol. Macromol. 2018, 113, 1214–1220. [Google Scholar] [CrossRef] [PubMed]
- Pereira, T.C.; de Barros, P.P.; Fugisaki, L.R.d.O.; Rossoni, R.D.; Ribeiro, F.d.C.; de Menezes, R.T.; Junqueira, J.C.; Scorzoni, L. Recent Advances in the Use of Galleria mellonella Model to Study Immune Responses against Human Pathogens. J. Fungi 2018, 4, 128. [Google Scholar] [CrossRef]
- Sheehan, G.; Kavanagh, K. Analysis of the early cellular and humoral responses of Galleria mellonella larvae to infection by Candida albicans. Virulence 2018, 9, 163–172. [Google Scholar] [CrossRef]
- Browne, N.; Heelan, M.; Kavanagh, K. An analysis of the structural and functional similarities of insect hemocytes and mammalian phagocytes. Virulence 2013, 4, 597–603. [Google Scholar] [CrossRef]
- Wojda, I. Immunity of the greater wax moth Galleria mellonella. Insect Sci. 2017, 24, 342–357. [Google Scholar] [CrossRef]
- Semenova, A.D.; Glazachev, Y.I.; Slepneva, I.A.; Glupov, V. Quantitative determination of nitric oxide production in haemocytes: Nitrite reduction activity as a potential pathway of NO formation in haemolymph of Galleria mellonella larvae. Nitric Oxide 2014, 37, 46–52. [Google Scholar] [CrossRef]
- Brown, S.E.; Howard, A.; Kasprzak, A.B.; Gordon, K.H.; East, P.D. A peptidomics study reveals the impressive antimicrobial peptide arsenal of the wax moth Galleria mellonella. Insect Biochem. Mol. Biol. 2009, 39, 792–800. [Google Scholar] [CrossRef]
- De Moraes Borba, C.; Schäffer, G.M. Paracoccidioides brasiliensis: Virulence and an attempt to induce the dimorphic process with fetal calf serum. Mycoses 2002, 45, 174–179. [Google Scholar] [CrossRef]
- Rooney, P.J.; Klein, B.S. Linking fungal morphogenesis with virulence. Cell. Microbiol. 2002, 4, 127–137. [Google Scholar] [CrossRef]










Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Marcos, C.M.; de Oliveira, H.C.; Assato, P.A.; de Oliveira, L.T.; Fregonezi, N.; dos Santos, K.S.; Costa-Orlandi, C.B.; Fusco-Almeida, A.M.; Mendes-Giannini, M.J.S. Polypeptides Targeting Paracoccidioides brasiliensis Drk1. J. Fungi 2023, 9, 980. https://doi.org/10.3390/jof9100980
Marcos CM, de Oliveira HC, Assato PA, de Oliveira LT, Fregonezi N, dos Santos KS, Costa-Orlandi CB, Fusco-Almeida AM, Mendes-Giannini MJS. Polypeptides Targeting Paracoccidioides brasiliensis Drk1. Journal of Fungi. 2023; 9(10):980. https://doi.org/10.3390/jof9100980
Chicago/Turabian StyleMarcos, Caroline Maria, Haroldo Cesar de Oliveira, Patricia Akemi Assato, Lariane Teodoro de Oliveira, Nathália Fregonezi, Kelvin Sousa dos Santos, Caroline Barcelos Costa-Orlandi, Ana Marisa Fusco-Almeida, and Maria José Soares Mendes-Giannini. 2023. "Polypeptides Targeting Paracoccidioides brasiliensis Drk1" Journal of Fungi 9, no. 10: 980. https://doi.org/10.3390/jof9100980
APA StyleMarcos, C. M., de Oliveira, H. C., Assato, P. A., de Oliveira, L. T., Fregonezi, N., dos Santos, K. S., Costa-Orlandi, C. B., Fusco-Almeida, A. M., & Mendes-Giannini, M. J. S. (2023). Polypeptides Targeting Paracoccidioides brasiliensis Drk1. Journal of Fungi, 9(10), 980. https://doi.org/10.3390/jof9100980