
The C2H2 Zinc Finger Protein MaNCP1 Contributes to Conidiation through Governing the Nitrate Assimilation Pathway in the Entomopathogenic Fungus Metarhizium acridum


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains and Culture Conditions
2.2. Conidial Yield and Fungal Growth Assays
2.3. Bioinformatics Analysis and Yeast One-Hybrid Assay
2.4. Electrophoretic Mobility Shift Assay (EMSA)
2.5. Quantitative Reverse Transcription PCR (qRT-PCR) Analyses
2.6. RNA-Seq Analysis
2.7. Statistical Analysis
3. Results
3.1. MaNCP1 Regulates the Conidiation
3.2. MaNCP1 Regulates the Nitrogen Utilization
3.3. MaNCP1 Regulates the Expression of Nitrate Metabolism Genes
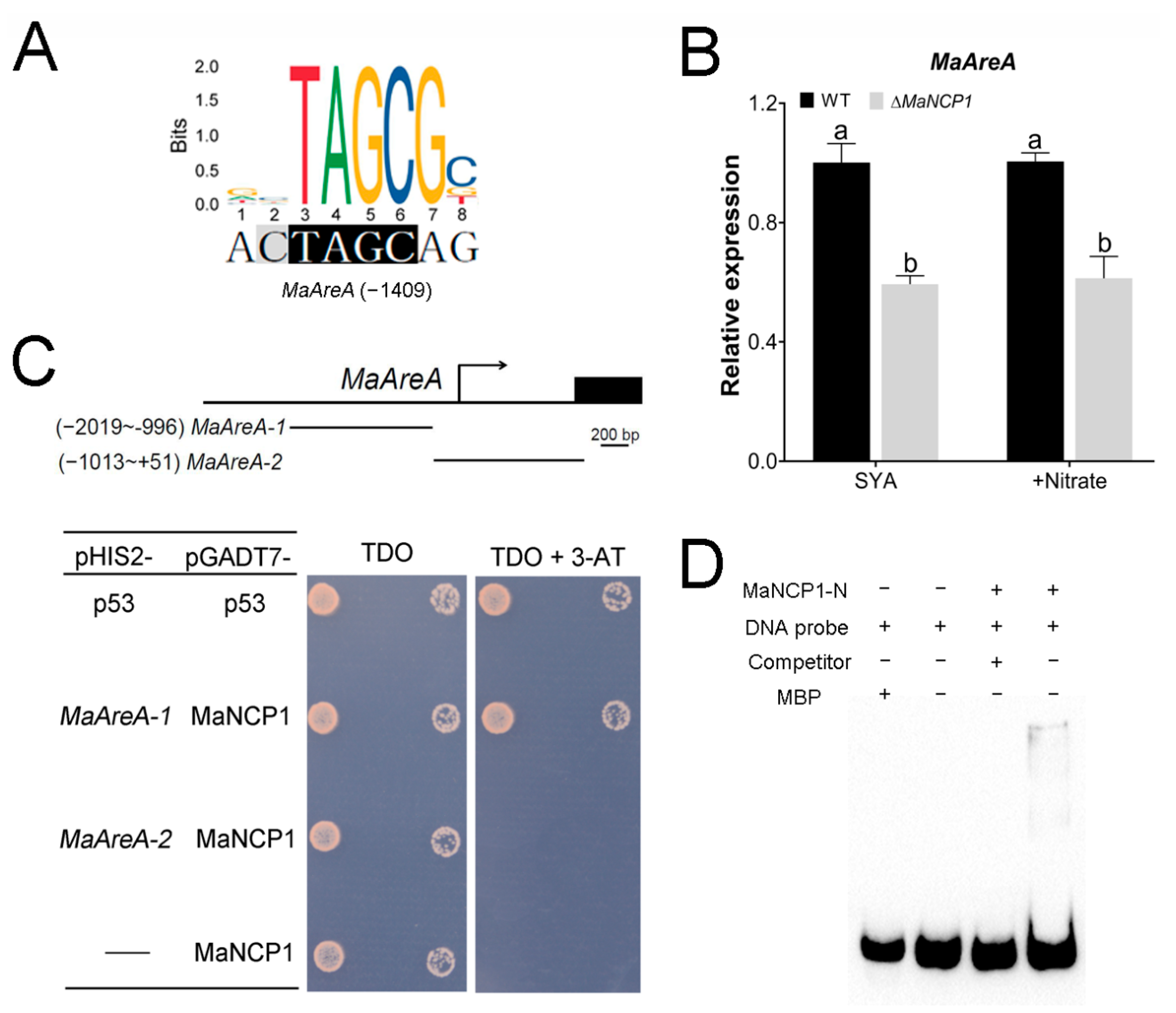
3.4. MaAreA Is a Target Gene of MaNCP1
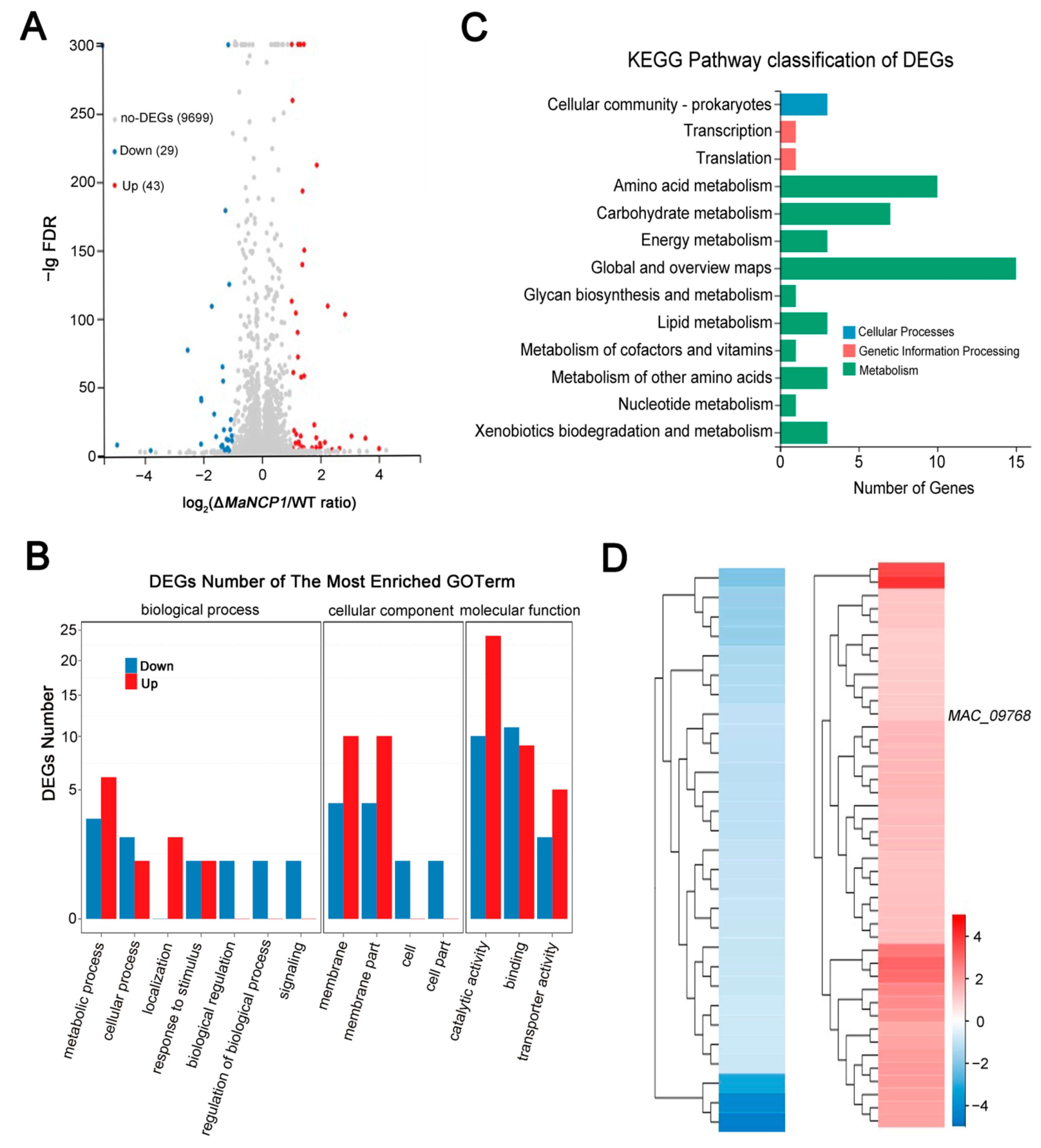
3.5. Transcriptomic Insights into Pleiotropic Effects of MaNCP1
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Savary, S.; Bregaglio, S.; Willocquet, L.; Gustafson, D.; D’Croz, D.M.; Sparks, A.; Castilla, N.; Djurle, A.; Allinne, C.; Sharma, M.; et al. Crop health and its global impacts on the components of food security. Food Secur. 2017, 9, 311–327. [Google Scholar] [CrossRef]
- Savary, S.; Willocquet, L.; Pethybridge, S.J.; Esker, P.; McRoberts, N.; Nelson, A. The global burden of pathogens and pests on major food crops. Nat. Ecol. Evol. 2019, 3, 430–439. [Google Scholar] [CrossRef] [PubMed]
- Savary, S.; Teng, P.S.; Willocquet, L.; Nutter, F.W. Quantification and modeling of crop losses: A review of purposes. Annu. Rev. Phytopathol. 2006, 44, 89–112. [Google Scholar] [CrossRef] [PubMed]
- Esker, P.; Savary, S.; McRoberts, N. Crop loss analysis and global food supply: Focusing now on required harvests. CAB Rev. 2012, 7, 1–14. [Google Scholar] [CrossRef]
- Wang, C.S.; Feng, M.G. Advances in fundamental and applied studies in China of fungal biocontrol agents for use against arthropod pests. Biol. Control 2014, 68, 129–135. [Google Scholar] [CrossRef]
- Wang, C.S.; Wang, S.B. Insect pathogenic fungi: Genomics, molecular interactions, and genetic improvements. Annu. Rev. Entomol. 2017, 62, 73–90. [Google Scholar] [CrossRef]
- Jackson, M.A.; Cliquet, S.; Iten, L.B. Media and fermentation processes for the rapid production of high concentrations of stable blastospores of the bioinsecticidal fungus Paecilomyces fumosoroseus. Biocontrol Sci. Techn. 2003, 13, 23–33. [Google Scholar] [CrossRef]
- Mascarin, G.M.; Alves, S.B.; Lopes, R.B. Culture media selection for mass production of Isaria fumosorosea and Isaria farinosa. Braz. Arch. Biol. Techn. 2010, 53, 753–761. [Google Scholar] [CrossRef]
- Mirabito, P.M.; Adams, T.H.; Timberlake, W.E. Interactions of three sequentially expressed genes control temporal and spatial specificity in Aspergillus development. Cell 1989, 57, 859–868. [Google Scholar] [CrossRef]
- Adams, T.H.; Wieser, J.K.; Yu, J.H. Asexual sporulation in Aspergillus nidulans. Microbiol. Mol. Biol. Rev. 1998, 62, 35–54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, H.S.; Yu, J.H. Genetic control of asexual sporulation in filamentous fungi. Curr. Opin. Microbiol. 2012, 15, 669–677. [Google Scholar] [CrossRef] [PubMed]
- Wieser, J.; Lee, B.N.; Fondon, J.W.; Adams, T.H. Genetic requirements for initiating asexual development in Aspergillus nidulans. Curr. Genet. 1994, 27, 62–69. [Google Scholar] [CrossRef]
- Seo, J.A.; Guan, Y.; Yu, J.H. FluG-dependent asexual development in Aspergillus nidulans occurs via derepression. Genetics 2006, 172, 1535–1544. [Google Scholar] [CrossRef]
- Lee, B.N.; Adams, T.H. The Aspergillus nidulans fluG gene is required for production of an extracellular developmental signal and is related to prokaryotic glutamine synthetase I. Gene Dev. 1994, 8, 641–651. [Google Scholar] [CrossRef]
- Sekiguchi, J.; Gaucher, G.M.; Costerton, J.W. Microcycle conidiation in Penicillium urticae: An ultrastructural investigation of conidiogenesis. Can. J. Microbiol. 1975, 21, 2069–2083. [Google Scholar] [CrossRef]
- Bosch, A.; Yantorno, O. Microcycle conidiation in the entomopathogenic fungus Beauveria bassiana bals. (vuill.). Process Biochem. 1999, 34, 707–716. [Google Scholar] [CrossRef]
- Ni, M.; Gao, N.; Kwon, N.J.; Shin, K.S.; Yu, J.H. Regulation of Aspergillus conidiation. In Cellular and Molecular Biology of Filamentous Fungi; Borkovich, K., Ebbole, D., Eds.; ACM Press: Washington, DC, USA, 2010; pp. 559–576. [Google Scholar]
- Jung, B.; Kim, S.; Lee, J. Microcyle conidiation in filamentous fungi. Mycobiology 2014, 42, 1–5. [Google Scholar] [CrossRef]
- Zhang, S.; Xia, Y. Identification of genes preferentially expressed during microcycle conidiation of Metarhizium anisopliae using suppression subtractive hybridization. FEMS Microbiol. Lett. 2008, 286, 71–77. [Google Scholar] [CrossRef]
- Zhang, S.; Peng, G.; Xia, Y. Microcycle conidiation and the conidial properties in the entomopathogenic fungus Metarhizium anisopliae on agar medium. Biocontrol Sci. Techn. 2010, 20, 809–819. [Google Scholar] [CrossRef]
- Wang, Z.L.; Jin, K.; Xia, Y.X. Transcriptional analysis of the conidiation pattern shift of the entomopathogenic fungus Metarhizium acridum in response to different nutrients. BMC Genom. 2016, 17, 586. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Todd, R.B. Regulation of fungal nitrogen metabolism. In The Mycota III: Biochemistry and Molecular Biology; Hoffmeister, D., Ed.; Springer International Publishing: Cham, Switzerland, 2016; pp. 281–303. [Google Scholar]
- Tudzynski, B. Nitrogen regulation of fungal secondary metabolism in fungi. Front. Microbiol. 2014, 5, 656. [Google Scholar] [CrossRef] [PubMed]
- Langdon, T.; Sheerins, A.; Ravagnani, A.; Gielkens, M.; Caddick, M.X.; Arst, H.N., Jr. Mutational analysis reveals dispensability of the N-terminal region of the Aspergillus transcription factor mediating nitrogen metabolite repression. Mol. Microbiol. 1995, 17, 877–888. [Google Scholar] [CrossRef] [PubMed]
- Morozov, I.Y.; Galbis-Martinez, M.; Jones, M.G.; Caddick, M.X. Characterization of nitrogen metabolite signalling in Aspergillus via the regulated degradation of areA mRNA. Mol. Microbiol. 2001, 42, 269–277. [Google Scholar] [CrossRef] [PubMed]
- Caddick, M.X.; Jones, M.G.; van Tonder, J.M.; Le Cordier, H.; Narendja, F.; Strauss, J.; Morozov, I.Y. Opposing signals differentially regulate transcript stability in Aspergillus nidulans. Mol. Microbiol. 2006, 62, 509–519. [Google Scholar] [CrossRef]
- Todd, R.B.; Fraser, J.A.; Wong, K.H.; Davis, M.A.; Hynes, M.J. Nuclear accumulation of the GATA factor AreA in response to complete nitrogen starvation by regulation of nuclear export. Eukaryot Cell 2005, 4, 1646–1653. [Google Scholar] [CrossRef]
- Andrianopoulos, A.; Kourambas, S.; Sharp, J.A.; Davis, M.A.; Hynes, M.J. Characterization of the Aspergillus nidulans nmrA gene involved in nitrogen metabolite repression. J. Bacteriol. 1998, 180, 1973–1977. [Google Scholar] [CrossRef]
- Wong, K.H.; Hynes, M.J.; Todd, R.B.; Davis, M.A. Transcriptional control of nmrA by the bZIP transcription factor MeaB reveals a new level of nitrogen regulation in Aspergillus nidulans. Mol. Microbiol. 2007, 66, 534–551. [Google Scholar] [CrossRef]
- Li, C.C.; Xu, D.X.; Hu, M.W.; Zhang, Q.P.; Xia, Y.X.; Jin, K. MaNCP1, a C2H2 zinc finger protein, governs the conidiation pattern shift through regulating the reductive pathway for nitric oxide synthesis in the filamentous fungus Metarhizium acridum. Microbiol. Spectr. 2022, 10, e00538-22. [Google Scholar] [CrossRef]
- Li, C.C.; Xia, Y.X.; Jin, K. N-terminal zinc fingers of MaNCP1 contribute to growth, stress tolerance, and virulence in Metarhizium acridum. Int. J. Biol. Macromol. 2022, 216, 426–436. [Google Scholar] [CrossRef]
- Li, C.C.; Zhang, Q.P.; Xia, Y.X.; Jin, K. MaNmrA, a negative transcription regulator in nitrogen catabolite repression pathway, contributes to nutrient utilization, stress resistance and virulence in entomopathogenic fungus Metarhizium acridum. Biology 2021, 10, 1167. [Google Scholar] [CrossRef]
- Fornes, O.; Castro-Mondragon, J.A.; Khan, A.; van der Lee, R.; Zhang, X.; Richmond, P.A.; Modi, B.P.; Correard, S.; Gheorghe, M.; Baranasic, D.; et al. JASPAR 2020: Update of the open-access database of transcription factor binding profiles. Nucleic Acids Res. 2020, 48, D87–D92. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCt method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Chen, E.J.; Kaiser, C.A. Amino acids regulate the intracellular trafficking of the general amino acid permease of Saccharomyces cerevisiae. Proc. Natl. Acad. Sci. USA 2002, 99, 14837–14842. [Google Scholar] [CrossRef] [PubMed]
- Scherens, B.; Feller, A.; Vierendeels, F.; Messenguy, F.; Dubois, E. Identification of direct and indirect targets of the Gln3 and Gat1 activators by transcriptional profiling in response to nitrogen availability in the short and long term. FEMS Yeast Res. 2006, 6, 777–791. [Google Scholar] [CrossRef]
- Cruz-Leite, V.R.M.; Salem-Izacc, S.M.; Novaes, E.; Neves, B.; Brito, W.D.; Silva, L.O.S.; Paccez, J.D.; Parente-Rocha, J.A.; Pereira, M.; Soares, C.M.D.; et al. Nitrogen Catabolite Repression in members of Paracoccidioides complex. Microb. Pathog. 2020, 149, 104281. [Google Scholar] [CrossRef] [PubMed]
- Murphy, R.L.; Andrianopoulos, A.; Davis, M.A.; Hynes, M.J. Identification of amdx, a new Cys2His2 (C2H2) zinc-finger gene involved in the regulation of the amds gene of Aspergillus nidulans. Mol. Microbiol. 1997, 23, 591–602. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.R.; Chae, K.S.; Hant, K.H.; Han, D.M. The nsdC gene encoding a putative C2H2-type transcription factor is a key activator of sexual development in Aspergillus nidulans. Genetics 2009, 182, 771–783. [Google Scholar] [CrossRef] [PubMed]
- Hu, P.; Wang, Y.; Zhou, J.; Pan, Y.; Liu, G. AcstuA, which encodes an APSES transcription regulator, is involved in conidiation, cephalosporin biosynthesis and cell wall integrity of Acremonium chrysogenum. Fungal Genet. Biol. 2015, 83, 26–40. [Google Scholar] [CrossRef]
- Zhang, M.; Wei, Q.; Xia, Y.; Jin, K. MaPacC, a pH-responsive transcription factor, negatively regulates thermotolerance and contributes to conidiation and virulence in Metarhizium acridum. Curr. Genet. 2020, 66, 397–408. [Google Scholar] [CrossRef]
- Song, D.X.; Cao, Y.Q.; Xia, Y.X. Transcription factor MaMsn2 regulates conidiation pattern shift under the control of MaH1 through homeobox domain in Metarhizium acridum. J. Fungi 2021, 7, 840. [Google Scholar] [CrossRef]
- Cao, H.; Huang, P.; Zhang, L.; Shi, Y.; Sun, D.; Yan, Y.; Liu, X.; Dong, B.; Chen, G.; Snyder, J.H.; et al. Characterization of 47 Cys2-His2 zinc finger proteins required for the development and pathogenicity of the rice blast fungus Magnaporthe oryzae. New Phytol. 2016, 211, 1035–1051. [Google Scholar] [CrossRef]
- Stanbrough, M.; Magasanik, B. Two transcription factors, Gln3p and Nil1p, use the same GATAAG sites to activate the expression of GAP1 of Saccharomyces cerevisiae. J. Bacteriol 1996, 178, 2465–2468. [Google Scholar] [CrossRef] [PubMed]
- Narendja, F.; Goller, S.P.; Wolschuk, M.; Strauss, J. Nitrate and the GATA factor AreA are necessary for in vivo binding of NirA, the pathway-specific transcriptiona lactivator of Aspergillus nidulans. Mol. Microbiol. 2002, 44, 573–583. [Google Scholar] [CrossRef] [PubMed]
- Muro-Pastor, M.I.; Strauss, J.; Ramon, A.; Scazzocchio, C. A paradoxical mutant GATA factor. Eukaryot Cell 2004, 3, 393–405. [Google Scholar] [CrossRef]
- Berger, H.; Basheer, A.; Bock, S.; Reyes-Dominguez, Y.; Dalik, T.; Altmann, F.; Strauss, J. Dissecting individual steps of nitrogen transcription factor cooperation in the Aspergillus nidulans nitrate cluster. Mol. Microbiol. 2008, 69, 1385–1398. [Google Scholar] [CrossRef]
- Wong, K.H.; Hynes, M.J.; Todd, R.B.; Davis, M.A. Deletion and overexpression of the Aspergillus nidulans GATA factor AreB reveals unexpected pleiotropy. Microbiology 2009, 155, 3868–3880. [Google Scholar] [CrossRef] [PubMed]
- Wiemann, P.; Sieber, C.M.K.; vonBargen, K.W.; Studt., L.; Niehaus, E.M.; Espino, J.J.; Huß, K.; Michielse, C.B.; Albermann, S.; Wagner, D.; et al. Deciphering the cryptic genome: Genome-wide analyses of the rice pathogen Fusarium fujikuroi reveal complex regulation of secondary metabolism and novel metabolites. PLoS Pathog. 2013, 9, e1003475. [Google Scholar] [CrossRef] [PubMed]
- Wilson, R.A.; Arst, H.N. Mutational analysis of AREA, a transcriptional activator mediating nitrogen metabolite repression in Aspergillus nidulans and a member of the “streetwise” GATA family of transcription factors. Microbiol. Mol. Biol. Rev. 1998, 62, 586–596. [Google Scholar] [CrossRef] [PubMed]
- Ninnemann, H.; Maier, J. Indications for the occurrence of nitric oxide synthases in fungi and plants and the involvement in photoconidiation of Neurospora crassa. Photochem. Photobiol. 1996, 64, 393–398. [Google Scholar] [CrossRef]
- Gong, X.; Fu, Y.; Jiang, D.; Li, G.; Yi, X.; Peng, Y. L-Arginine is essential for conidiation in the filamentous fungus Coniothyrium minitans. Fungal Genet. Biol. 2007, 44, 368–1379. [Google Scholar] [CrossRef]
- Zweier, J.L.; Li, H.; Samouilov, A.; Liu, X. Mechanisms of nitrite reduction to nitric oxide in the heart and vessel wall. Nitric Oxide 2010, 22, 83–90. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Lim, J.; Xu, J.; Yu, J.H.; Zheng, W. Nitric oxide as a developmental and metabolic signal in filamentous fungi. Mol. Microbiol. 2020, 113, 872–882. [Google Scholar] [CrossRef] [PubMed]
- Badaruddin, M.; Holcombe, L.J.; Wilson, R.A.; Wang, Z.Y.; Kershaw, M.J.; Talbot, N.J. Glycogen metabolic genes are involved in trehalose-6-phosphate synthase-mediated regulation of pathogenicity by the rice blast fungus. Magnaporthe oryzae. PLoS Pathog. 2013, 9, e1003604. [Google Scholar] [CrossRef] [PubMed]
- Zhou, T.; Qin, L.; Zhu, X.; Shen, W.; Zou, J.; Wang, Z.; Wei, Y. The D-lactate dehydrogenase MoDLD1 is essential for growth and infection-related development in Magnaporthe oryzae. Environ. Microbiol. 2017, 19, 3938–3958. [Google Scholar] [CrossRef]
- Chen, W.; Wei, L.; Zhang, Y.; Shi, D.; Ren, W.; Zhang, Z.; Wang, J.; Shao, W.; Liu, X.; Chen, C.; et al. Involvement of the two L-lactate dehydrogenase in development and pathogenicity in Fusarium graminearum. Curr. Genet. 2019, 65, 591–605. [Google Scholar] [CrossRef]
- Storts, D.R.; Bhattacharjee, J.K. Properties of revertants of lys2 and lys5 mutants as well as alpha-aminoadipate-semialdehyde dehydrogenase from Saccharomyces cerevisiae. Biochem. Bioph. Res. Co. 1989, 161, 182–186. [Google Scholar] [CrossRef]
- Vander Beken, S.; de Vries, J.C.; Meier-Schiesser, B.; Meyer, P.; Jiang, D.; Sindrilaru, A.; Ferreira, F.F.; Hainzl, A.; Schatz, S.; Muschhammer, J.; et al. Newly defined ATP-binding cassette subfamily b member 5 positive dermal mesenchymal stem cells promote healing of chronic iron-overload wounds via secretion of interleukin-1 receptor antagonist. Stem. Cells 2019, 37, 1057–1074. [Google Scholar] [CrossRef]
- Koivistoinen, O.M.; Richard, P.; Penttilä, M.; Ruohonen, L.; Mojzita, D. Sorbitol dehydrogenase of Aspergillus niger, SdhA, is part of the oxido-reductive D-galactose pathway and essential for D-sorbitol catabolism. FEBS Lett. 2012, 586, 378–383. [Google Scholar] [CrossRef]
- Chang, T.S. An updated review of tyrosinase inhibitors. Int. J. Mol. Sci. 2009, 10, 2440–2475. [Google Scholar] [CrossRef]
- Hayaishi, O.; Katagiri, M.; Rothberg, S. Mechanism of the pyrocatechase reaction. J. Am. chem. Soc. 1955, 77, 5450–5451. [Google Scholar] [CrossRef]
- Ozawa, K.; Iwasa, H.; Sasaki, N.; Kinoshita, N.; Hiratsuka, A.; Yokoyama, K. Identification and characterization of thermostable glucose dehydrogenases from thermophilic filamentous fungi. Appl. Microbiol. Biot. 2017, 101, 173–183. [Google Scholar] [CrossRef] [PubMed]
- Deng, J.J.; Huang, W.Q.; Li, Z.W.; Lu, D.L.; Zhang, Y.; Luo, X.C. Biocontrol activity of recombinant aspartic protease from Trichoderma harzianum against pathogenic fungi. Enzyme Microb. Tech. 2018, 12, 35–42. [Google Scholar] [CrossRef] [PubMed]
- Gorlatova, N.; Tchorzewski, M.; Kurihara, T.; Soda, K.; Esaki, N. Purification, characterization, and mechanism of a flavin mononucleotidedependent 2-nitropropane dioxygenase from Neurospora crassa. Appl. Environ. Microb. 1998, 64, 1029–1033. [Google Scholar] [CrossRef] [PubMed]
- Francis, K.; Russell, B.; Gadda, G. Involvement of a flavosemiquinone in the enzymatic oxidation of nitroalkanes catalyzed by 2-nitropropane dioxygenase. J. Biol. Chem. 2005, 280, 5195–5204. [Google Scholar] [CrossRef]
- Thakur, S.; Chakrabarti, A. Saccharomyces cerevisiae Hsp30 is necessary for homeostasis of a set of thermal stress response functions. J. Microbiol. Biotechn. 2010, 20, 403–409. [Google Scholar] [CrossRef]
- Wamelink, M.; Grüning, N.M.; Jansen, E.E.; Bluemlein, K.; Lehrach, H.; Jakobs, C.; Ralser, M. The difference between rare and exceptionally rare: Molecular characterization of ribose 5-phosphate isomerase deficiency. J. Mol. Med. 2010, 88, 931–939. [Google Scholar] [CrossRef]
- Jauniaux, J.C.; Grenson, M. GAP1, the general amino acid permease gene of Saccharomyces cerevisiae. FEBS J. 1990, 190, 39–44. [Google Scholar]
- Kim, G.J.; Lee, D.E.; Kim, H.S. Construction and evaluation of a novel bifunctional N-carbamylase–D-hydantoinase fusion enzyme. Appl. Environ. Microb. 2000, 66, 2133–2138. [Google Scholar] [CrossRef]
- Etxebeste, O.; Herrero-Garcia, E.; Cortese, M.S.; Garzia, A.; Oiartzabal-Arano, E.; de los Ríos, V.; Ugalde, U.; Espeso, E.A. GmcA is a putative glucose-methanol-choline oxidoreductase required for the induction of asexual development in Aspergillus nidulans. PloS ONE 2012, 7, e40292. [Google Scholar] [CrossRef]
- Tomme, P.; Warren, R.A.J.; Gilkes, N.R. Cellulose hydrolysis by bacteria and fungi. Adv. Microb. Physiol. 1995, 37, 1–81. [Google Scholar]
- Sun, W.; Shen, Y.H.; Yang, W.J.; Cao, Y.F.; Xiang, Z.H.; Zhang, Z. Expansion of the silkworm GMC oxidoreductase genes is associated with immunity. Insect Biochem. Molec. 2012, 42, 935–945. [Google Scholar] [CrossRef] [PubMed]
- Shin, J.Y.; Bui, D.C.; Lee, Y.; Nam, H.; Jung, S.; Fang, M.; Kim, J.C.; Lee, T.; Kim, H.; Choi, G.J.; et al. Functional characterization of cytochrome P450 monooxygenases in the cereal head blight fungus Fusarium graminearum. Environ. Microbiol. 2017, 19, 2053–2067. [Google Scholar] [CrossRef] [PubMed]
- Johnson, G. The α/β hydrolase fold proteins of mycobacterium tuberculosis, with reference to their contribution to virulence. Curr. Protein Pept. Sc. 2017, 18, 190–210. [Google Scholar] [CrossRef] [PubMed]
- Tisi, A.; Angelini, R.; Cona, A. Wound healing in plants cooperation of copper amine oxidase and flavin-containing polyamine oxidase. Plant Signal. Behav. 2008, 3, 204–206. [Google Scholar] [CrossRef]
- Piscitelli, A.; Del Vecchio, C.; Faraco, V.; Giardina, P.; Macellaro, G.; Miele, A.; Pezzella, C.; Sannia, G. Fungal laccases: Versatile tools for lignocellulose transformation. Comptes Rendus Biol. 2011, 334, 789–794. [Google Scholar] [CrossRef]
- Langfelder, K.; Jahn, B.; Gehringer, H.; Schmidt, A.; Wanner, G.; Brakhage, A.A. Identification of a polyketide synthase gene (pksP) of Aspergillus fumigatus involved in conidial pigment biosynthesis and virulence. Med. Microbiol. Immunol. 1998, 187, 79–89. [Google Scholar] [CrossRef]
- Jeon, J.H.; Kim, S.-J.; Lee, H.S.; Cha, S.-S.; Lee, J.H.; Yoon, S.-H.; Koo, B.-S.; Lee, C.-M.; Choi, S.H.; Lee, S.H.; et al. Novel metagenome-derived carboxylesterase that hydrolyzes β-lactam antibiotics. Appl. Environ. Microbiol. 2011, 77, 7830–7836. [Google Scholar] [CrossRef] [Green Version]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, C.; Xia, Y.; Jin, K. The C2H2 Zinc Finger Protein MaNCP1 Contributes to Conidiation through Governing the Nitrate Assimilation Pathway in the Entomopathogenic Fungus Metarhizium acridum. J. Fungi 2022, 8, 942. https://doi.org/10.3390/jof8090942
Li C, Xia Y, Jin K. The C2H2 Zinc Finger Protein MaNCP1 Contributes to Conidiation through Governing the Nitrate Assimilation Pathway in the Entomopathogenic Fungus Metarhizium acridum. Journal of Fungi. 2022; 8(9):942. https://doi.org/10.3390/jof8090942
Chicago/Turabian StyleLi, Chaochuang, Yuxian Xia, and Kai Jin. 2022. "The C2H2 Zinc Finger Protein MaNCP1 Contributes to Conidiation through Governing the Nitrate Assimilation Pathway in the Entomopathogenic Fungus Metarhizium acridum" Journal of Fungi 8, no. 9: 942. https://doi.org/10.3390/jof8090942
APA StyleLi, C., Xia, Y., & Jin, K. (2022). The C2H2 Zinc Finger Protein MaNCP1 Contributes to Conidiation through Governing the Nitrate Assimilation Pathway in the Entomopathogenic Fungus Metarhizium acridum. Journal of Fungi, 8(9), 942. https://doi.org/10.3390/jof8090942