
Effect of High-Temperature Stress on Plant Physiological Traits and Mycorrhizal Symbiosis in Maize Plants


 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material, AMF Inoculum
2.2. Growing Conditions and Experimental Design
2.3. Measurement of Total Chlorophyll Content
2.4. Chlorophyll a Fluorescence Measurement
2.5. Measurement of Malondialdehyde (MDA) Content
2.6. AMF Root Colonization and Spore Density
2.7. Quantification of Signature Fatty Acids in Soil
2.8. Statistical Analysis
3. Results
3.1. Total Chlorophyll Content Measurement
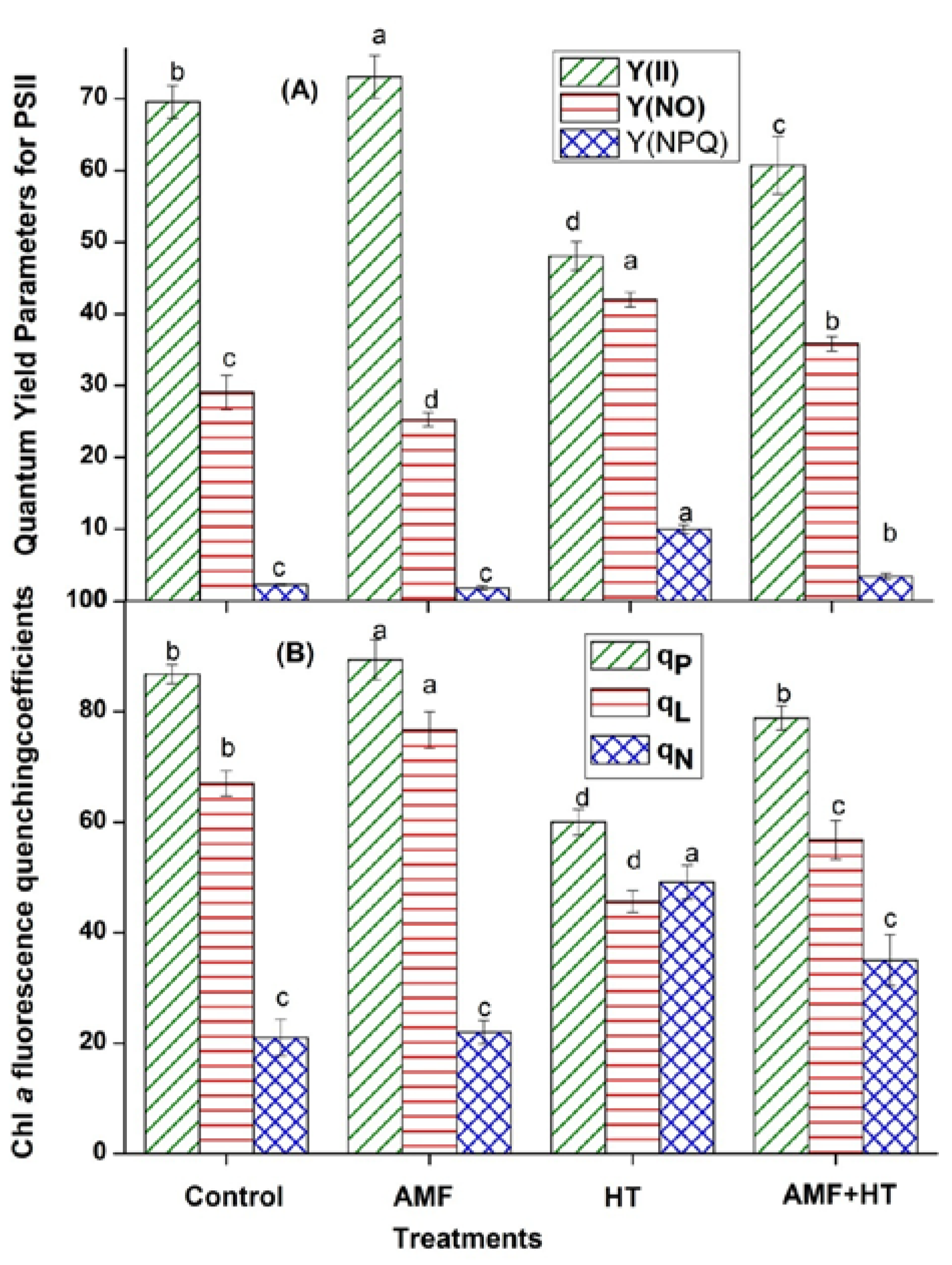
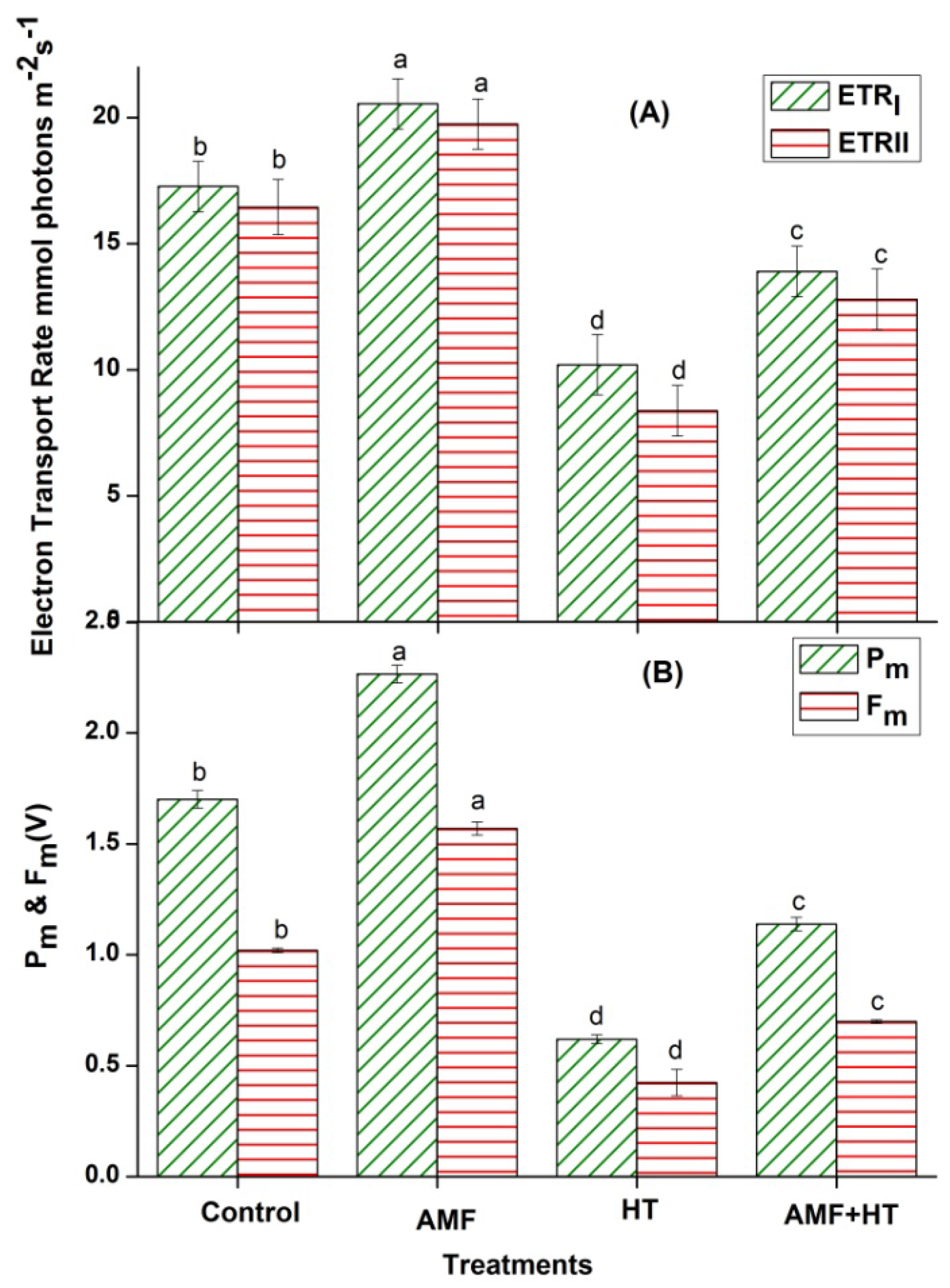
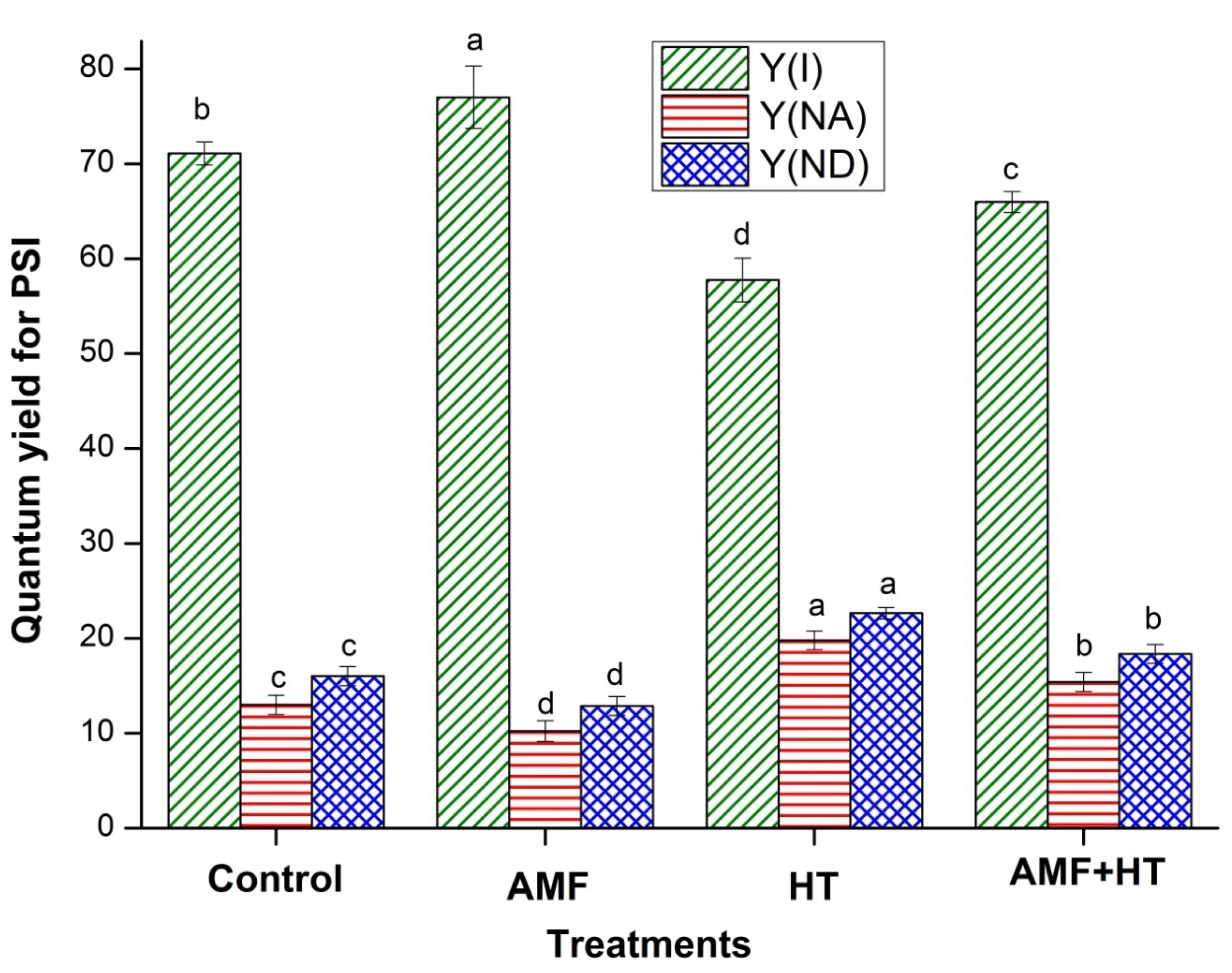
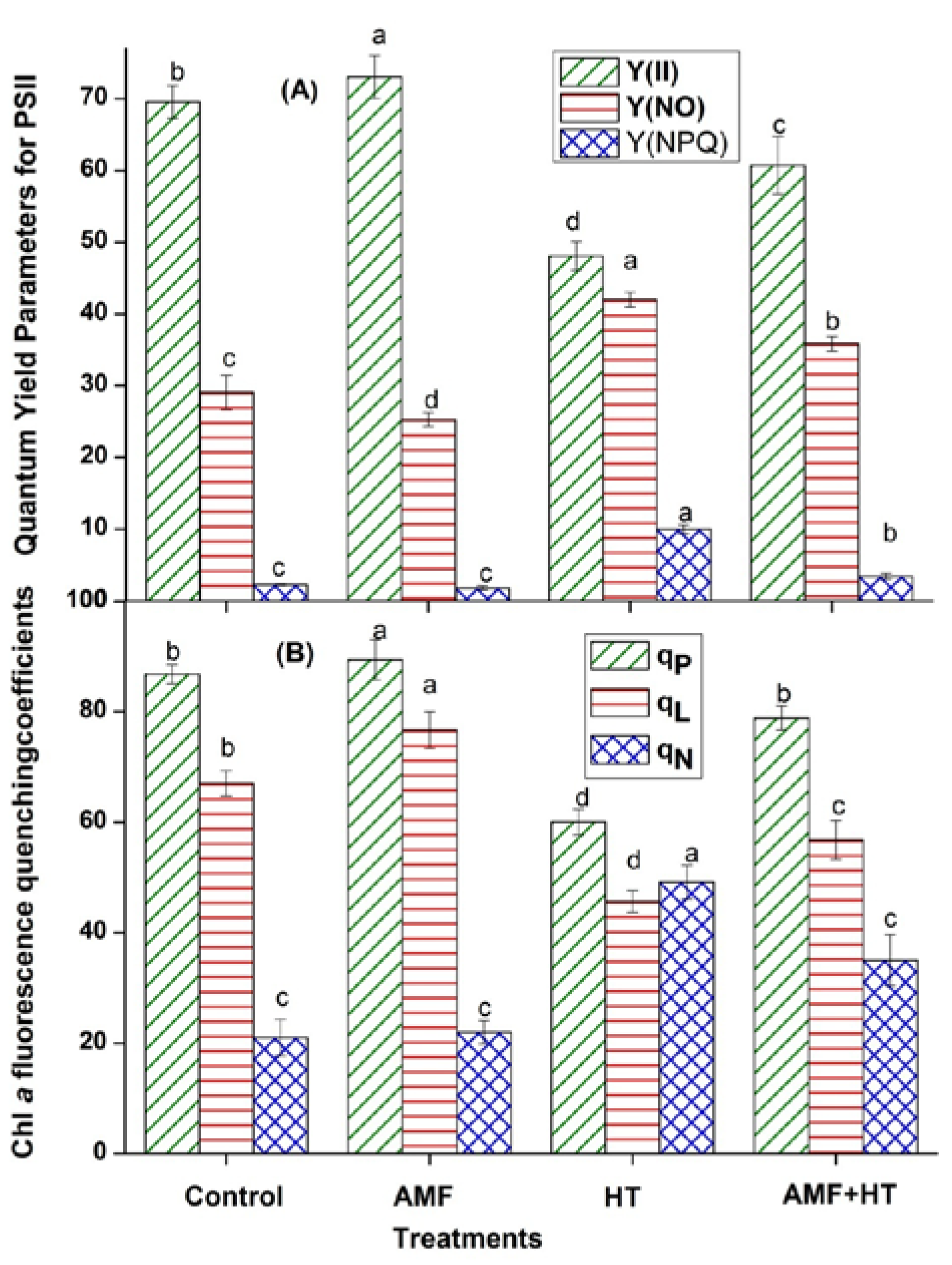
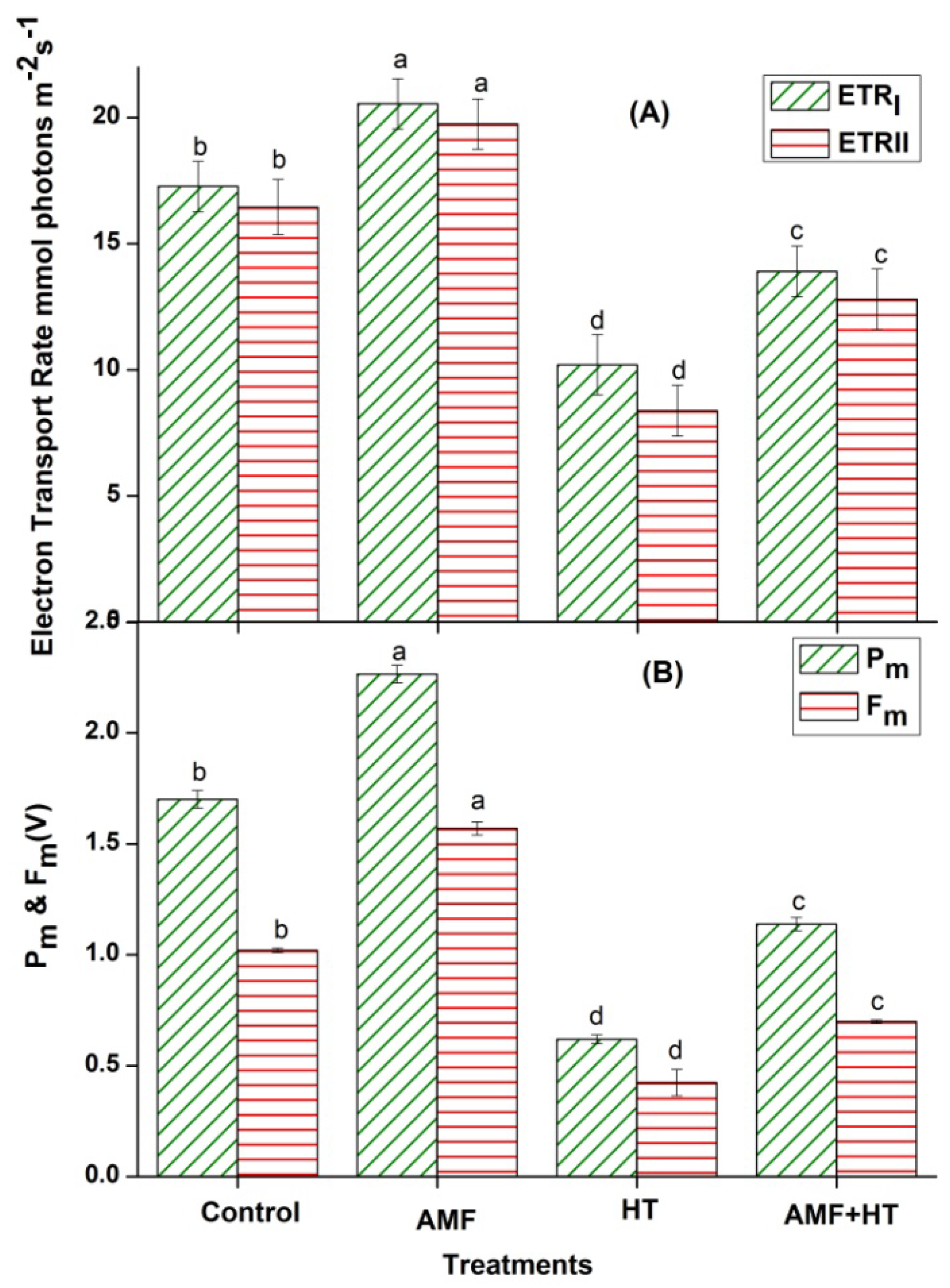
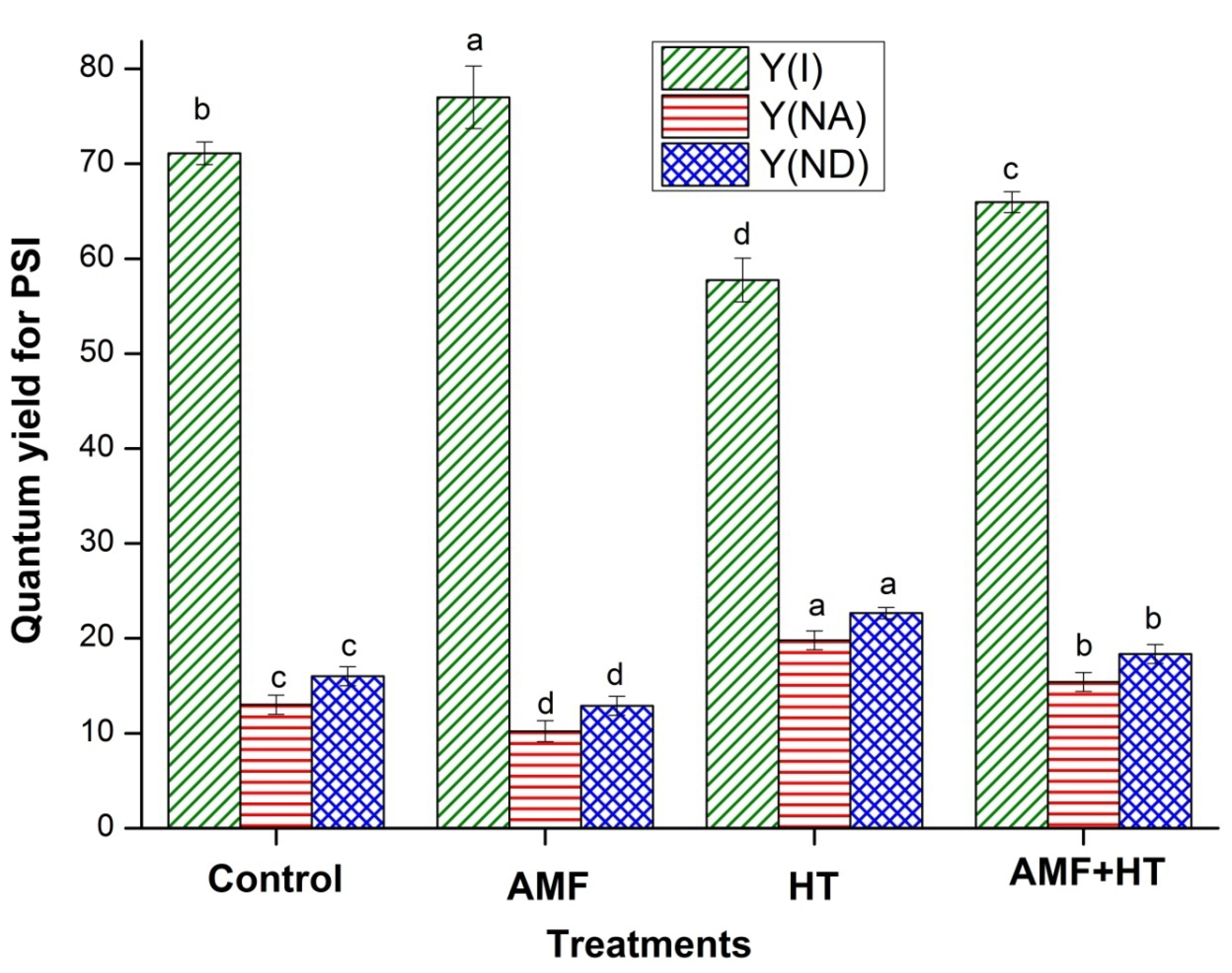
3.2. Chlorophyll (Chl) a Fluorescence Measurements
3.3. Root Colonization and Spore Density
3.4. PLFA Analysis
3.5. MDA Content
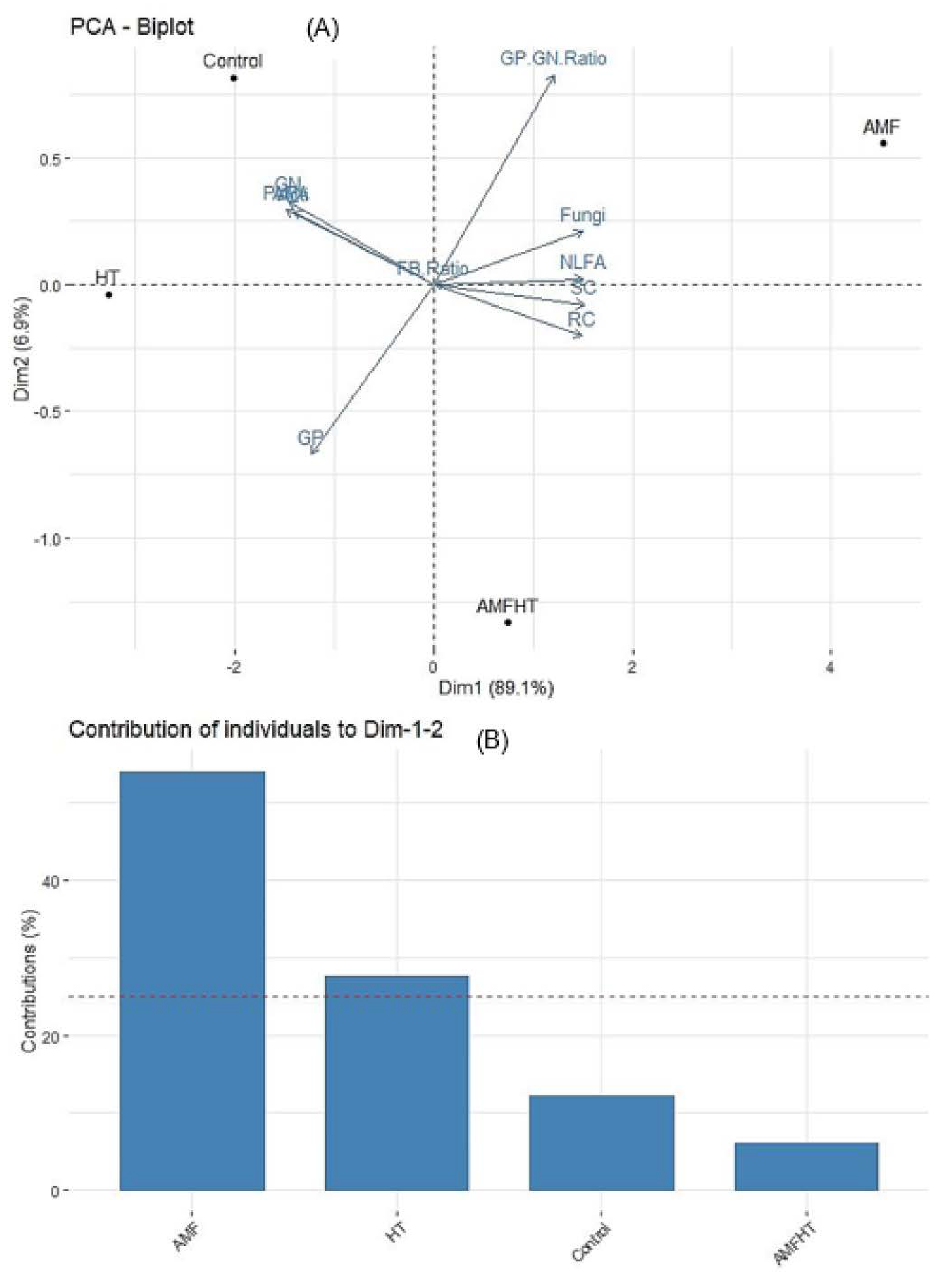
3.6. Principal Component Analysis (PCA)
4. Discussion
4.1. Total Chlorophyll Content
4.2. Chlorophyll a Fluorescence
4.3. Root Colonization and Spore Density
4.4. PLFA
4.5. MDA Content
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Tiwari, Y.K.; Yadav, S.K. High temperature stress tolerance in maize (Zea mays L.): Physiological and molecular mechanisms. J. Plant Biol. 2019, 62, 93–102. [Google Scholar] [CrossRef]
- Hussain, H.A.; Men, S.; Hussain, S.; Chen, Y.; Ali, S.; Sai, Z.; Zhang, K.; Li, Y.; Xu, Q.; Liao, C.; et al. Interactive effects of drought and heat stresses on morpho-physiological attributes, yield, nutrient uptake and oxidative status in maize hybrids. Sci. Rep. 2019, 9, 3890. [Google Scholar] [CrossRef] [Green Version]
- Mathur, S.; Agrawal, D.; Jajoo, A. Photosynthesis: Response to high temperature stress. J. Photochem. Photobiol. B Biol. 2014, 137, 116–126. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Nahar, K.; Alam, M.d.M.; Roychowdhury, R.; Fujita, M. Physiological, biochemical, and molecular mechanisms of heat stress tolerance in plants. Int. J. Mol. Sci. 2013, 14, 9643–9684. [Google Scholar] [CrossRef]
- Jagadish, S.V.K.; Way, D.A.; Sharkey, T.D. Plant heat stress: Concepts directing future research. Plant Cell Environ. 2021, 44, 1992–2005. [Google Scholar] [CrossRef]
- Ruiz-Lozano, J.M.; Aroca, R. Modulation of aquaporin genes by the arbuscular mycorrhizal symbiosis in relation to osmotic stress tolerance. In Symbioses and Stress; Seckbach, J., Grube, M., Eds.; Springer: Dordrecht, The Netherlands, 2010; pp. 357–374. [Google Scholar]
- Smith, S.; Read, D. Mycorrhiza Symbiosis; Academic Press: London, UK, 2008. [Google Scholar]
- Birhane, E.; Sterck, F.; Fetene, M.; Bongers, F.; Kuyper, T. Arbuscular mycorrhizal fungi enhance photosynthesis, water use efficiency, and growth of frankincense seedlings under pulsed water availability conditions. Oecologia 2012, 169, 895–904. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Augé, R.M.; Toler, H.D.; Saxton, A.M. Arbuscular mycorrhizal symbiosis alters stomatal conductance of host plants more under drought than under amply watered conditions: A meta-analysis. Mycorrhiza 2014, 25, 13–24. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Lizarazo, J.C.; Moreno-Fonseca, L.P. Mechanisms for tolerance to water-deficit stress in plants inoculated with arbuscular mycorrhizal fungi. A review. Agron. Colomb. 2016, 34, 179–189. [Google Scholar] [CrossRef]
- Hussain, H.A.; Qingwen, Z.; Hussain, S.; Hongbo, L.; Waqqas, A.; Li, Z. Effects of arbuscular mycorrhizal fungi on maize growth, root colonization, and root exudates varied with inoculum and application method. J. Soil Sci. Plant Nutri. 2021, 21, 2. [Google Scholar]
- Gavito, M.E.; Olsson, P.A.; Rouhier, H.; Medinapeñafiel, A.; Jakobsen, I.; Bago, A. Temperature constraints on the growth and functioning of root organ cultures with arbuscular mycorrhizal fungi. New Phytol. 2005, 168, 179–188. [Google Scholar] [CrossRef] [PubMed]
- Mathur, S.; Tomar, R.S.; Jajoo, A. Arbuscular mycorrhizal fungi (AMF) protects photosynthetic apparatus of wheat under drought stress. Photosynth. Res. 2019, 139, 227–238. [Google Scholar] [CrossRef]
- Alotaibi, M.O.; Saleh, A.M.; Sobrinho, R.L.; Sheteiwy, M.S.; El-Sawah, A.M.; Mohammed, A.E.; Elgawad, H.A. Arbuscular mycorrhizae mitigate aluminum toxicity and regulate proline metabolism in plants grown in acidic soil. J. Fungi 2021, 7, 531. [Google Scholar] [CrossRef]
- Eustis, A.; Murphy, K.M.; Barrios-Masias, F.H. Leaf gas exchange performance of ten Quinoa genotypes under a simulated heatwave. Plants 2020, 9, 81. [Google Scholar] [CrossRef] [Green Version]
- Lennon, J.T.; Jones, S.E. Microbial seed banks: The ecological and evolutionary implications of dormancy. Nat. Rev. Microbiol. 2011, 9, 119–130. [Google Scholar] [CrossRef] [PubMed]
- Rui, J.; Li, J.; Wang, S.; An, J.; Liu, W.; Lin, Q.; Yang, Y.; He, Z.; Li, X. Responses of bacterial communities to simulated climate changes in alpine meadow soil of the Qinghai-Tibet plateau. Appl. Environ. Microbiol. 2015, 81, 6070–6077. [Google Scholar] [CrossRef] [Green Version]
- Olsson, P.A.; Thingstrup, I.; Jakobsen, I.; Bååth, E. Estimation of the biomass of arbuscular mycorrhizal fungi in a linseed field. Soil Biol. Biochem. 1999, 31, 1879–1887. [Google Scholar] [CrossRef]
- Sharma, M.P.; Buyer, J.S. Comparison of biochemical and microscopic methods for quantification of arbuscular mycorrhizal fungi in soil and roots. Appl. Soil Ecol. 2015, 95, 86–89. [Google Scholar] [CrossRef]
- Agnihotri, R.; Bharti, A.; Ramesh, A.; Prakash, A.; Sharma, M.P. Glomalin related protein and C16:1ω5 PLFA associated with AM fungi as potential signatures for assessing the soil C sequestration under contrasting soil management practices. Eur. J. Soil Biol. 2021, 103, 103286. [Google Scholar] [CrossRef]
- Feng, X.; Simpson, M.J. Temperature and substrate controls on microbial phospholipid fatty acid composition during incubation of grassland soils contrasting in organic matter quality. Soil Biol. Biochem. 2009, 41, 804–812. [Google Scholar] [CrossRef]
- Wu, Y.; Yu, X.; Wang, H.; Ding, N.; Xu, J. Does history matter? Temperature effects on soil microbial biomass and community structure based on the phospholipids fatty acid (PLFA) analysis. J. Soils Sediments 2010, 10, 223–230. [Google Scholar] [CrossRef]
- Mathur, S.; Sharma, M.P.; Jajoo, A. Improved photosynthetic efficacy of maize (Zea mays) plants with Arbuscular mycorrhizal fungi (AMF) under high temperature stress. J. Photochem. Photobiol. B Biol. 2018, 180, 149–154. [Google Scholar] [CrossRef] [PubMed]
- Gerdemann, J.; Nicolson, T. Spores of mycorrhizal Endogone species extracted from soil by wet sieving and decanting. Trans. Br. Mycol. Soc. 1963, 46, 235–244. [Google Scholar] [CrossRef]
- Sharma, M.P.; Singh, S.; Sharma, S.K.; Ramesh, A.; Bhatia, V.S. Co-inoculation of resident AM Fungi and soybean rhizobia enhanced nodulation, yield, soil biological parameters and saved fertilizer inputs in vertisols under microcosm and field conditions. Soybean Res. 2016, 14, 39–53. [Google Scholar]
- Zhu, J.; Tremblay, N.; Liang, Y. Comparing SPAD and at LEAF values for chlorophyll assessment in crop species. Can. J. Soil Sci. 2012, 92, 645–648. [Google Scholar] [CrossRef]
- Pfündel, E.; Klughammer, C.; Ulrich, S. Monitoring the effects of reduced PSII antenna size on quantum yields of photosystems I and II using the Dual-PAM-100 measuring system. PAM Appl. Notes 2008, 1, 21–24. [Google Scholar]
- Kramer, D.M.; Avenson, T.J.; Kanazawa, A.; Cruz, J.A.; Ivanov, B.; Edwards, G.E. The relationship between photosynthetic electron transfer and its regulation. In Chlorophyll a Fluorescence: A Signature of Photosynthesis; Papageorgiou, G.C., Govindjee, Eds.; Springer: Dordrecht, The Netherlands, 2004; pp. 251–278. [Google Scholar]
- Klughammer, C.; Schreiber, U. Complementary PSII quantum yields calculated from simple fluorescence parameters measured by PAM fluorometry and the saturation pulse method. PAM Appl. Notes 2008, 1, 27–35. [Google Scholar]
- Hussain, M.I.; Reigosa, M.J. Characterization of xanthophyll pigments, photosynthetic performance, photon energy dissipation, reactive oxygen species generation and carbon isotope discrimination during artemisinin-induced stress in Arabidopsis thaliana. PLoS ONE 2015, 10, e0114826. [Google Scholar]
- Chen, J.; Zhang, H.; Zhang, X.; Tang, M. Arbuscular mycorrhizal symbiosis alleviates salt stress in black locust through improved photosynthesis, water status, and K+/Na+ homeostasis. Front. Plant Sci. 2017, 8, 1739. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, J.; Yan, X.; Sun, S.; Lin, J. The effect of arbuscular mycorrhizal fungi on Photosystem II of the host plant under salt stress: A meta-analysis. Agronomy 2019, 9, 806. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.L.; Qu, W. Experimental Guidance of Plant Physiology; Higher Education Press: Beijing, China, 2004. [Google Scholar]
- Koske, R.E.; Gemma, J.N. A modified procedure for staining roots to detect VA mycorrhizas. Mycol. Res. 1989, 92, 486–488. [Google Scholar] [CrossRef]
- Biermann, B.J.; Lindermann, R.Q. Quantifying vesicular-arbuscular mycorrhizae: Proposed method towards standardization. New Phytol. 1981, 87, 63–67. [Google Scholar] [CrossRef]
- Buyer, J.S.; Sasser, M. High throughput phospholipid fatty acid analysis of soils. Appl. Soil Ecol. 2012, 61, 127–130. [Google Scholar] [CrossRef]
- Buyer, J.S.; Teasdale, J.R.; Roberts, D.P.; Zasada, I.A.; Maul, J.E. Factors affecting soil microbial community structure in tomato cropping systems. Soil Biol. Biochem. 2010, 42, 831–841. [Google Scholar] [CrossRef]
- CoHort/CoStat Software. CoHort Software. 798 Lighthouse Ave., PMB 320; CoHort/CoStat Software: Monterey, CA, USA, 1991. [Google Scholar]
- Lê, S.; Josse, J.; Husson, F. FactoMineR: An R package for multivariate analysis. J. Stat. Softw. 2008, 25, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Wickham, H.; Hester, J.; Chang, W. Devtools: Tools to Make Developing R Packages Easier. R Package Devtools Version 2.3.1. 2019. Available online: https://cran.r-project.org/package=devtools (accessed on 23 July 2020).
- Kassambara, A.; Mundt, F. Factoextra: Extract and Visualize the Results of Multivariate Data Analyses. Package Version 1.0.7 R Package Version. 2020. Available online: https://rpkgs.datanovia.com/factoextra/ (accessed on 23 July 2020).
- Liu, C.Y.; Zhang, F.; Zhang, D.J.; Srivastava, A.K.; Wu, Q.S.; Zou, Y.N. Mycorrhiza stimulates root-hair growth and IAA synthesis and transport in trifoliate orange under drought stress. Sci. Rep. 2018, 8, 1978. [Google Scholar] [CrossRef] [Green Version]
- Mathur, S.; Jajoo, A. Arbuscular mycorrhizal fungi protects maize plants from high temperature stress by regulating photosystem II heterogeneity. Ind. Crops Prod. 2020, 143, 111934. [Google Scholar] [CrossRef]
- He, J.D.; Zou, Y.N.; Wu, Q.S.; Kamil, K. Mycorrhizas enhance drought tolerance of trifoliate orange by enhancing activities and gene expression of antioxidant enzymes. Sci. Hortic. 2020, 262, 108745. [Google Scholar] [CrossRef]
- Augé, R.M. Water relations, drought and vesicular-arbuscular mycorrhizal symbiosis. Mycorrhiza 2001, 11, 3–42. [Google Scholar] [CrossRef]
- Pozo, M.J.; López-Ráez, J.A.; Azcón-Aguilar, C.; García-Garrido, J.M. Phytohormones as integrators of environmental signals in the regulation of mycorrhizal symbioses. New Phytol. 2015, 205, 1431–1436. [Google Scholar] [CrossRef]
- Haneef, I.; Faizan, S.; Perveen, R.; Kausar, S. Role of arbuscular mycorrhizal fungi on growth and photosynthetic pigments in (Coriandrum sativum L.) grown under cadmium stress. World J. Agric. Sci. 2013, 9, 245–250. [Google Scholar]
- Chen, J.; Burke, J.J.; Xin, Z. Chlorophyll fluorescence analysis revealed essential roles of FtsH11 protease in regulation of the adaptive responses of photosynthetic systems to high temperature. BMC Plant Biol. 2018, 18, 11. [Google Scholar] [CrossRef] [Green Version]
- Allakhverdiev, S.I.; Kreslavski, V.D.; Klimov, V.V.; Los, D.A.; Carpentier, R.; Mohanty, P. Heat stress: An overview of molecular responses in photosynthesis. Photosynth. Res. 2008, 98, 541–550. [Google Scholar] [CrossRef]
- Tomar, R.S.; Jajoo, A. Photosystem I (PSI) becomes more tolerant to fluoranthene due to initiation of cyclic electron flow (CEF). Funct. Plant Biol. 2017, 44, 978–984. [Google Scholar] [CrossRef]
- Brestic, M.; Zivcak, M.; Kunderlikova, K.; Sytar, O.; Shao, H.; Kalaji, H.M.; Allakhverdiev, S.I. Low PSI content limits the photoprotection of PSI and PSII in early growth stages of chlorophyll b-deficient wheat mutant lines. Photosynth. Res. 2015, 125, 151–166. [Google Scholar] [CrossRef]
- Oukarroum, A.; Schansker, G.; Strasser, R.J. Drought stress effects on Photosystem I content and Photosystem II thermotolerance analyzed using Chl a fluorescence kinetics in barley varieties differing in their drought tolerance. Physiol. Plant 2009, 137, 188–199. [Google Scholar] [CrossRef] [PubMed]
- Mo, Y.; Wang, Y.; Yang, R.; Zheng, J.; Liu, C.; Li, H.; Ma, J.; Zhang, Y.; Wei, C.; Zhang, X. Regulation of plant growth, photosynthesis, antioxidation and osmosis by an Arbuscular mycorrhizal fungus in Watermelon seedlings under well-watered and drought conditions. Front. Plant Sci. 2016, 7, 644. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shikanai, T. Cyclic electron transport around Photosystem I: Genetic approaches. Ann. Rev. Plant Biol. 2007, 58, 199–217. [Google Scholar] [CrossRef]
- Olsson, P.A.; Bååth, E.; Jakobsen, I. Phosphorus effects on the mycelium and storage structures of an arbuscular mycorrhizal fungus as studied in the soil and roots by analysis of fatty acid signatures. Appl. Environ. Microbiol. 1997, 63, 3531–3538. [Google Scholar] [CrossRef] [Green Version]
- Nichols, P.; Stulp, B.K.; Jones, J.G.; White, D.C. Comparison of fatty acid content and DNA homology of the filamentous gliding bacteria Vitreoscilla, Flexibacter, Filibacter. Arch. Microbiol. 1986, 146, 1–6. [Google Scholar] [CrossRef]
- Bunn, R.; Lekberg, Y.; Zabinski, C. Arbuscular mycorrhizal fungi ameliorate temperature stress in thermophilic plants. Ecology 2009, 90, 1378–1388. [Google Scholar] [CrossRef] [PubMed]
- Heinemeyer, A.; Fitter, A.H. Impact of temperature on the arbuscular mycorrhizal (AM) symbiosis: Growth responses of the host plant and its AM fungal partner. J. Exp. Bot. 2004, 55, 525–534. [Google Scholar] [CrossRef] [Green Version]
- Ali, R.S.; Poll, C.; Kandeler, E. Dynamics of soil respiration and microbial communities: Interactive controls of temperature and substrate quality. Soil Biol. Biochem. 2018, 127, 60–70. [Google Scholar] [CrossRef]
- Bai, Z.; Ma, Q.; Wu, X.; Zhang, Y.; Yu, W. Temperature sensitivity of a PLFA-distinguishable microbial community differs between varying and constant temperature regimes. Geoderma 2017, 308, 54–59. [Google Scholar] [CrossRef]
- Ali, M.B.; Hahn, E.; Paek, K. Effects of temperature on oxidative stress defense systems, lipid peroxidation and lipoxygenase activity in Phalaenopsis. Plant Physiol. Biochem. 2005, 43, 213–223. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.C.; Song, F.B.; Xu, H.W. Influence of arbuscular mycorrhizae on lipid peroxidation and antioxidant enzyme activity of maize plants under temperature stress. Mycorrhiza 2010, 20, 325–332. [Google Scholar] [CrossRef]
- Zhu, X.; Song, F.; Liu, F. Arbuscular mycorrhizal fungi and tolerance of temperature stress in plants. In Arbuscular Mycorrhizas and Stress Tolerance of Plants; Wu, Q.S., Ed.; Springer Nature: Singapore, 2017. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fluorescence Parameters | Description |
|---|---|
| ETR | electron transport rate |
| Fo | minimal fluorescence |
| Fm | maximum fluorescence |
| Pm | maximal change in P700 signal during the quantitative transformation of P700 from the fully reduced to the fully oxidized state |
| qL | fraction of open PSII reaction centers |
| qP | photochemical quenching coefficient used to assess PSII susceptibility to photo-inhibition and reflects the oxidation-reduction state of the primary acceptor (QA) for PSII |
| qN | non-photochemical quenching coefficient, i.e., the fraction of dark-adapted variable fluorescence that is lost upon adaptation to light |
| Y(I) | effective photochemical quantum yield of PSI |
| Y(II) | effective quantum yield of PSII |
| Y(NA) | quantum yield of non-photochemical energy dissipation of reaction centers due to PSI acceptor-side limitation |
| Y(ND) | quantum yield of non-photochemical energy dissipation in reaction centers due to PSI donor-side limitation |
| Y(NO) | quantum yield of non-regulated energy dissipation and the fraction of energy that is passively dissipated in the form of heat and fluorescence mainly due to the closed PSII centers |
| Y(NPQ) | quantum yield of light-induced non-photochemical fluorescence quenching |
| Treatments | MDA Concentration (μM mg−1 FWL) | Total Chl Concentration (SPAD Units) |
|---|---|---|
| Control | 57.0 ± 2.01 | 41 ± 1 |
| AMF | 32.0 * ± 2.0 | 46 * ± 1 |
| HT | 167 *** ± 10.06 | 20 *** ± 1 |
| AMF + HT | 88.0 ** ± 4.01 | 37 ** ± 2 |
| Treatment | 16:1ω5cis PLFA | 16:1ω5cis NLFA | Gram-Negative | Gram-Positive | Fungi | Actinomycetes |
|---|---|---|---|---|---|---|
| Control | 5.96 ± 0.42 a | 10.78 ± 5.17 c | 44.80 ± 4.18 a | 47.30 ± 16.95 a | 1.48 ± 0.35 a | 19.00 ± 5.82 a |
| AMF | 4.44 ± 0.01 c | 35.28 ± 0.64 a | 34.21 ± 3.54 b | 43.77 ± 4.59 a | 1.63 ± 0.00 a | 17.30 ± 1.62 a |
| HT | 6.21 ± 0.06 a | 10.18 ± 0.00 c | 43.50 ± 2.50 ab | 53.93 ± 3.88 a | 1.45 ± 0.26 a | 20.24 ± 3.53 a |
| AMF + HT | 4.97 ± 0.32 b | 20.66 ± 1.54 b | 38.83 ± 5.04 ab | 49.70 ± 6.89 a | 1.52 ± 0.01 a | 17.71 ± 3.19 a |
| LSD (p = 0.05) | 0.50 | 5.11 | 7.39 | 18.13 | 0.41 | 7.23 |
| Main effect | *** | *** | * | ns | ns | ns |
| Treatment | Fungi/Bacteria | Gram-Positive/Gram-Negative |
|---|---|---|
| Control | 0.0767 ± 0.01 a | 1.52 ± 0.09 a |
| AMF | 0.0800 ± 0.01 a | 1.60 ± 0.03 a |
| HT | 0.0800 ± 0.01 a | 1.51 ± 0.08 a |
| AMF + HT | 0.0833 ± 0.01 a | 1.50 ± 0.09 a |
| LSD (p = 0.05) | 0.02 | 0.14 |
| Main effect | ns | ns |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mathur, S.; Agnihotri, R.; Sharma, M.P.; Reddy, V.R.; Jajoo, A. Effect of High-Temperature Stress on Plant Physiological Traits and Mycorrhizal Symbiosis in Maize Plants. J. Fungi 2021, 7, 867. https://doi.org/10.3390/jof7100867
Mathur S, Agnihotri R, Sharma MP, Reddy VR, Jajoo A. Effect of High-Temperature Stress on Plant Physiological Traits and Mycorrhizal Symbiosis in Maize Plants. Journal of Fungi. 2021; 7(10):867. https://doi.org/10.3390/jof7100867
Chicago/Turabian StyleMathur, Sonal, Richa Agnihotri, Mahaveer P. Sharma, Vangimalla R. Reddy, and Anjana Jajoo. 2021. "Effect of High-Temperature Stress on Plant Physiological Traits and Mycorrhizal Symbiosis in Maize Plants" Journal of Fungi 7, no. 10: 867. https://doi.org/10.3390/jof7100867
APA StyleMathur, S., Agnihotri, R., Sharma, M. P., Reddy, V. R., & Jajoo, A. (2021). Effect of High-Temperature Stress on Plant Physiological Traits and Mycorrhizal Symbiosis in Maize Plants. Journal of Fungi, 7(10), 867. https://doi.org/10.3390/jof7100867