Ligninolytic Enzyme Production and Decolorization Capacity of Synthetic Dyes by Saprotrophic White Rot, Brown Rot, and Litter Decomposing Basidiomycetes


Abstract
1. Introduction
2. Materials and Methods
2.1. Organisms
2.2. Decolorization Assay
2.3. Ligninolytic Enzyme Assays
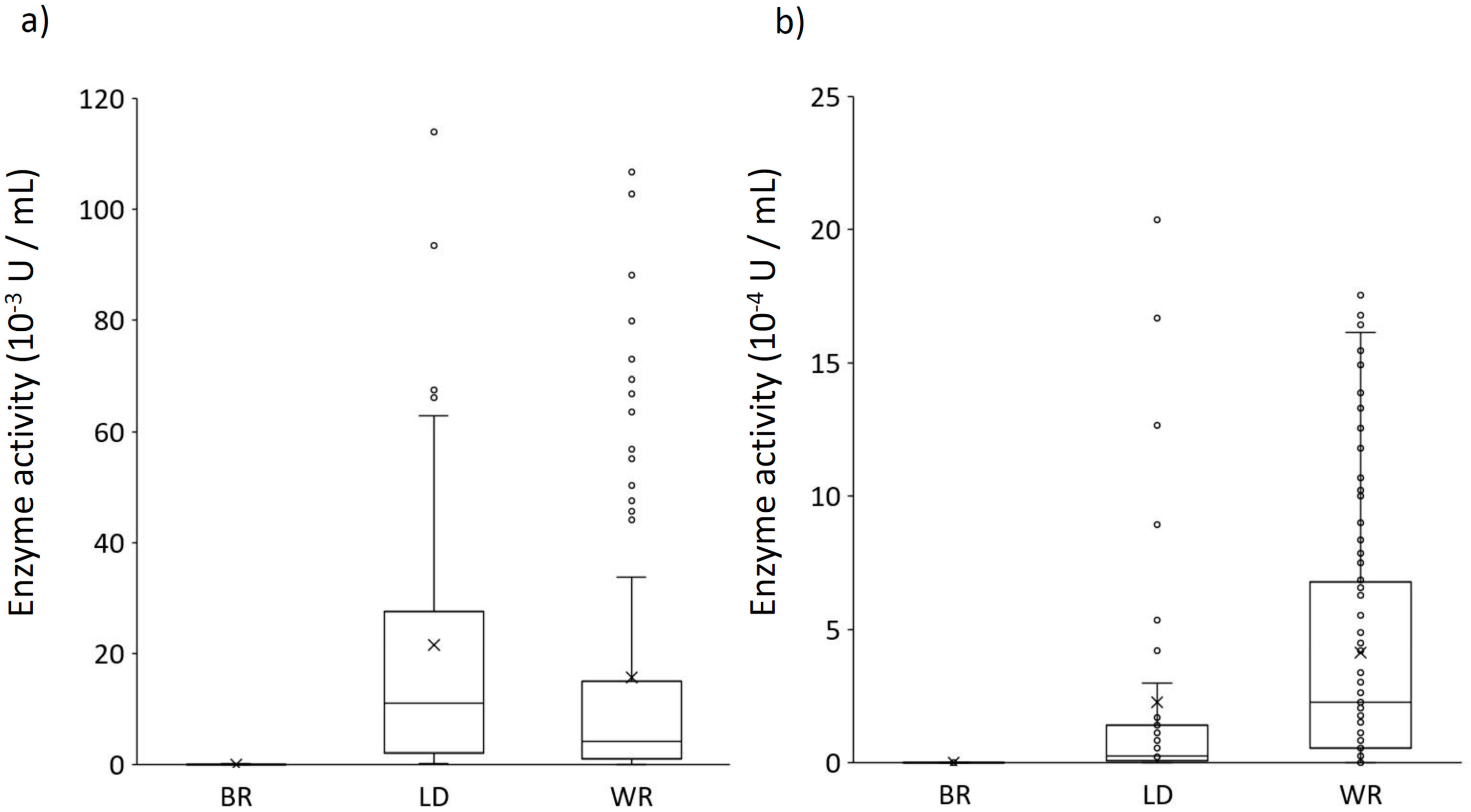
3. Results
3.1. Agar–Plate Screening for Orange G and RBBR Decolorization
3.2. Growth on Agar Plates Containing Dyes
3.3. Ligninolytic Enzyme Assays
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Lee, A.H.; Lee, H.; Heo, Y.M.; Lim, Y.W.; Kim, C.-M.; Kim, G.-H.; Chang, W.; Kim, J.-J. A proposed stepwise screening framework for the selection of polycyclic aromatic hydrocarbon (PAH)-degrading white rot fungi. Bioprocess Biosyst. Eng. 2020, 43, 767–783. [Google Scholar] [CrossRef] [PubMed]
- Sen, S.K.; Raut, S.; Bandyopadhyay, P.; Raut, S. Fungal decolouration and degradation of azo dyes: A review. Fungal Biol. Rev. 2016, 30, 112–133. [Google Scholar] [CrossRef]
- Baldrián, P. Chapter 2 Enzymes of Saprotrophic Basidiomycetes; Elsevier: Amsterdam, The Netherlands, 2008; Volume 28, pp. 19–41. [Google Scholar]
- Kjøller, A.; Struwe, S. Fungal Communities, Succession, Enzymes, and Decomposition. In Enzymes in the Environment; Marcel Dekker: New York, NY, USA, 2003; pp. 267–284. [Google Scholar]
- Van Der Wal, A.; Geydan, T.D.; Kuyper, T.W.; De Boer, W. A thready affair: Linking fungal diversity and community dynamics to terrestrial decomposition processes. FEMS Microbiol. Rev. 2013, 37, 477–494. [Google Scholar] [CrossRef] [PubMed]
- Baldrian, P.; Valášková, V. Degradation of cellulose by basidiomycetous fungi. FEMS Microbiol. Rev. 2008, 32, 501–521. [Google Scholar] [CrossRef]
- Hofrichter, M.; Ullrich, R.; Pecyna, M.J.; Liers, C.; Lundell, T. New and classic families of secreted fungal heme peroxidases. Appl. Microbiol. Biotechnol. 2010, 87, 871–897. [Google Scholar] [CrossRef] [PubMed]
- Martínez, M.J.; Speranza, M.; Ruiz-Dueñas, F.J.; Ferreira, P.; Camarero, S.; Guillénb, F.; Martínez, M.J.; Gutiérrez, A.; Del Río, J.C. Biodegradation of lignocellulosics: Microbial, chemical, and enzymatic aspects of the fungal attack of lignin. Int. Microbiol. 2005, 8, 195–204. [Google Scholar]
- Osono, T.; Takeda, H. Comparison of litter decomposing ability among diverse fungi in a cool temperate deciduous forest in Japan. In Mycologia; Bennett, J.W., Ed.; Taylor & Francis: London, UK, 2002; Volume 94, pp. 421–427. Available online: https://www.tandfonline.com/doi/abs/10.1080/15572536.2003.11833207 (accessed on 31 January 2017).
- Osono, T.; Takeda, H. Fungal decomposition of Abies needle and Betula leaf litter. Mycologia 2006, 98, 172–179. [Google Scholar] [CrossRef]
- Batista-García, R.A.; Kumar, V.V.; Ariste, A.; Tovar-Herrera, O.E.; Savary, O.; Peidro-Guzmán, H.; González-Abradelo, D.; Jackson, S.A.; Dobson, A.D.; Sánchez-Carbente, M.D.R.; et al. Simple screening protocol for identification of potential mycoremediation tools for the elimination of polycyclic aromatic hydrocarbons and phenols from hyperalkalophile industrial effluents. J. Environ. Manag. 2017, 198, 1–11. [Google Scholar] [CrossRef]
- Kamal, S.; Asgher, M.; Khalil-ur-Rehman, Q.; Zahir, Z.A. Hyperproduction of laccase by Pleurotus ostreatus IBL-02 during decolorization of Drimarene Brulliant Red K-4BL. Fresen Environ. Bull. 2011, 20, 1478–1486. [Google Scholar]
- Pointing, S. Feasibility of bioremediation by white-rot fungi. Appl. Microbiol. Biotechnol. 2001, 57, 20–33. [Google Scholar] [CrossRef]
- Steffen, K.T.; Cajthaml, T.; Šnajdr, J.; Baldrian, P. Differential degradation of oak (Quercus petraea) leaf litter by litter-decomposing basidiomycetes. Res. Microbiol. 2007, 158, 447–455. [Google Scholar] [CrossRef] [PubMed]
- Moreira, M.; Mielgo, I.; Feijoo, G.; Lema, J. Evaluation of different fungal strains in the decolourisation of synthetic dyes. Biotechnol. Lett. 2000, 22, 1499–1503. [Google Scholar] [CrossRef]
- Shin, K.; Oh, I.; Kim, C. Production and Purification of Remazol Brilliant Blue R Decolorizing Peroxidase from the Culture Filtrate of Pleurotus ostreatus. Appl. Environ. Microbiol. 1997, 63, 1744–1748. [Google Scholar] [CrossRef] [PubMed]
- Kapich, A.N.; A Jensen, K.; Hammel, K.E. Peroxyl radicals are potential agents of lignin biodegradation. FEBS Lett. 1999, 461, 115–119. [Google Scholar] [CrossRef]
- Tanaka, H.; Itakura, S.; Enoke, A. Hydroxyl Radical Generation by an Extracellular Low-Molecular-Weight Substance and Phenol Oxidase Activity During Wood Degradation by the White-Rot Basidiomycete Phanerochaete chrysosporium. J. Biotechnol. 1999, 53, 21–28. [Google Scholar] [CrossRef]
- Mehmood, R.T.; Asad, M.J.; Asgher, M.; Hadri, S.H.; Gulfraz, M.; Wu, J.D.; Bhatti, M.I.; Zaman, N.; Ahmed, D. First Report of using Response Surface Methodology for the Biodegradation of Single Azo Disperse Dyes by Indigenous Daedalea dickinsii-IEBL-2. Romanian Biotechnol. Lett. 2020, 25, 1236–1245. [Google Scholar] [CrossRef]
- Bibi, I.; Javed, S.; Ata, S.; Majid, F.; Kamal, S.; Sultan, M.; Jilani, K.; Umair, M.; Khan, M.I.; Iqbal, M.; et al. Biodegradation of synthetic orange G dye by Plearotus sojar-caju with Punica granatum peal as natural mediator. Biocatal. Agric. Biotechnol. 2019, 22, 101420. [Google Scholar] [CrossRef]
- Iark, D.; Buzzo, A.J.D.R.; Garcia, J.A.A.; Côrrea, V.G.; Helm, C.V.; Corrêa, R.C.G.; Peralta, R.A.; Moreira, R.D.F.P.M.; Bracht, A.; Peralta, R.M. Enzymatic degradation and detoxification of azo dye Congo red by a new laccase from Oudemansiella canarii. Bioresour. Technol. 2019, 289, 121655. [Google Scholar] [CrossRef]
- Rao, R.G.; Ravichandran, A.; Kandalam, G.; Kumar, S.A.; Swaraj, S.; Sridhar, M. Screening of wild basidiomycetes and evaluation of the biodegradation potential of dyes and lignin by manganese peroxidases. BioResources 2019, 14, 6558–6576. [Google Scholar]
- Pandey, R.K.; Tewari, S.; Tewari, L. Lignolytic mushroom Lenzites elegans WDP2: Laccase production, characterization, and bioremediation of synthetic dyes. Ecotoxicol. Environ. Saf. 2018, 158, 50–58. [Google Scholar] [CrossRef]
- Legerská, B.; Chmelová, D.; Ondrejovič, M. Decolourization and detoxification of monoazo dyes by laccase from the white-rot fungus Trametes versicolor. J. Biotechnol. 2018, 285, 84–90. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Sánchez, J.; Membrillo-Venegas, I.; Martínez-Trujillo, A.; García-Rivero, A. Decolorization of reactive black 5 by immobilized Trametes versicolor. Rev. Mex. Ing. Quim. 2018, 17, 107–121. [Google Scholar] [CrossRef]
- Yehia, R.S.; Rodriguez-Couto, S. Discoloration of the azo dye Congo Red by manganese-dependent peroxidase from Pleurotus sajor caju. Appl. Biochem. Microbiol. 2017, 53, 222–229. [Google Scholar] [CrossRef]
- Bosco, F.; Mollea, C.; Ruggeri, B. Decolorization of Congo Red by Phanerochaete chrysosporium: The role of biosorption and biodegradation. Environ. Technol. 2017, 38, 2581–2588. [Google Scholar] [CrossRef] [PubMed]
- Sing, N.N.; Husaini, A.; Zulkharnain, A.; Roslan, H.A. Decolourisation Capabilities of Ligninolytic Enzymes Produced by Marasmius cladophyllus UMAS MS8 on Remazol Brilliant Blue R and Other Azo Dyes. BioMed Res. Int. 2017, 2017, 1–8. [Google Scholar] [CrossRef]
- Sayahi, E.; Ladhari, N.; Mechichi, T.; Sakli, F. Azo dyes decolourization by the laccase fromTrametes trogii. J. Text. Inst. 2016, 107, 1478–1482. [Google Scholar] [CrossRef]
- Permpornsakul, P.; Prasongsuk, S.; Lotrakul, P.; Eveleigh, D.; Kobayashi, D.; Imai, T.; Punnapayak, H. Biological Treatment of Reactive Black 5 by Resupinate White Rot Fungus Phanerochaete sordida PBU 0057. Pol. J. Environ. Stud. 2016, 25, 1167–1176. [Google Scholar] [CrossRef]
- Qin, X.; Zhang, J.; Zhang, X.; Yang, Y. Induction, Purification and Characterization of a Novel Manganese Peroxidase from Irpex lacteus CD2 and Its Application in the Decolorization of Different Types of Dye. PLOS ONE 2014, 9, e113282. [Google Scholar] [CrossRef]
- Knop, D.; Ben-Ari, J.; Salame, T.M.; Levinson, D.; Yarden, O.; Hadar, Y. Mn2+-deficiency reveals a key role for the Pleurotus ostreatus versatile peroxidase (VP4) in oxidation of aromatic compounds. Appl. Microbiol. Biotechnol. 2014, 98, 6795–6804. [Google Scholar] [CrossRef]
- Daâssi, D.; Rodriguez-Couto, S.; Nasri, M.; Mechichi, T. Biodegradation of textile dyes by immobilized laccase form Coriolopsis galica into Ca-alginate beads. Int. Biodeter. Biodegr. 2014, 90, 71–78. [Google Scholar] [CrossRef]
- Bao, S.; Teng, Z.; Ding, S. Heterologous expression and characterization of a novel laccase isoenzyme with dyes decolorization potential from Coprinus comatus. Mol. Biol. Rep. 2012, 40, 1927–1936. [Google Scholar] [CrossRef] [PubMed]
- Neto, S.L.M.; Mussatto, S.I.; Machado, K.M.G.; Milagres, A.M.F. Decolorization of salt-alkaline effluent with industrial reactive dyes by laccase-producing basidiomycetes strains. Lett. Appl. Microbiol. 2013, 56, 283–290. [Google Scholar] [CrossRef] [PubMed]
- Salame, T.M.; Knop, D.; Levinson, D.; Mabjeesh, S.J.; Yarden, O.; Hadar, Y. Release of Pleurotus ostreatus Versatile-Peroxidase from Mn2+ Repression Enhances Anthropogenic and Natural Substrate Degradation. PLOS ONE 2012, 7, e52446. [Google Scholar] [CrossRef] [PubMed]
- Pakshirajan, K.; Jaiswal, S.; Das, R.K. Biodecolourization of azo dyes using Phanerochaete chrysosporium: Effect of culture conditions and enzyme activities. J. Sci. Ind. Res. 2011, 70, 987–991. [Google Scholar]
- Bhattacharya, S.; Das, A.; G, M.; K, V.; J, S. Mycoremediation of Congo red dye by filamentous fungi. Braz. J. Microbiol. 2011, 42, 1526–1536. [Google Scholar] [CrossRef] [PubMed]
- Diwaniyan, S.; Kharb, D.; Raghukumar, C.; Kuhad, R.C. Decolorization of Synthetic Dyes and Textile Effluents by Basidiomycetous Fungi. Water Air Soil Pollut. 2009, 210, 409–419. [Google Scholar] [CrossRef]
- Gomes, E.; Aguiar, A.P.; Carvalho, C.C.; Bonfá, M.R.B.; Silva, R.D.; Boscolo, M. Ligninases production by Basidiomycetes strains on lignocellulosic agricultural residues and their application in the decolorization of synthetic dyes. Braz. J. Microbiol. 2009, 40, 31–39. [Google Scholar] [CrossRef]
- Chhabra, M.; Mishra, S.; Sreekrishnan, T.R. Mediator-assisted Decolorization and Detoxification of Textile Dyes/Dye Mixture by Cyathus bulleri Laccase. Appl. Biochem. Biotechnol. 2008, 151, 587–598. [Google Scholar] [CrossRef]
- Junghanns, C.; Krauss, G.; Schlosser, D. Potential of aquatic fungi derived from diverse freshwater environments to decolourise synthetic azo and anthraquinone dyes. Bioresour. Technol. 2008, 99, 1225–1235. [Google Scholar] [CrossRef]
- Šušla, M.; Svobodová, K. Effect of various synthetic dyes on the production of manganese-dependent peroxidase isoenzymes by immobilized Irpex lacteus. World J. Microbiol. Biotechnol. 2007, 24, 225–230. [Google Scholar] [CrossRef]
- Abrahão, M.C.; Gugliotta, A.D.M.; Da Silva, R.; Fujieda, R.J.Y.; Boscolo, M.; Gomes, E. Ligninolytic activity from newly isolated basidiomycete strains and effect of these enzymes on the azo dye orange II decolourisation. Ann. Microbiol. 2008, 58, 427–432. [Google Scholar] [CrossRef]
- Kokol, V.; Doliška, A.; Eichlerová, I.; Baldrian, P.; Nerud, F. Decolorization of textile dyes by whole cultures of Ischnoderma resinosum and by purified laccase and Mn-peroxidase. Enzym. Microb. Technol. 2007, 40, 1673–1677. [Google Scholar] [CrossRef]
- Qin, P.; Wu, Y.; Adil, B.; Wang, J.; Gu, Y.; Yu, X.; Zhao, K.; Zhang, X.; Ma, M.; Chen, Q.; et al. Optimization of Laccase from Ganoderma lucidum Decolorizing Remazol Brilliant Blue R and Glac1 as Main Laccase-Contributing Gene. Molecules 2019, 24, 3914. [Google Scholar] [CrossRef] [PubMed]
- Anita, S.H.; Sari, F.P.; Yanto, D.H.Y. Decolorization of Synthetic Dyes by Ligninolytic Enzymes from Trametes hirsuta D7. Makara J. Sci. 2019, 23, 44–50. [Google Scholar] [CrossRef]
- Siddiqui, Y.; Surendran, A.; Fishal, E.M.M. Inhibition of Lignin Degrading Enzymes of Ganoderma spp.: An Alternative Control of Basal Stem Rot Disease of Oil Palm. Int. J. Agric. Biol. 2019, 3, 523–530. [Google Scholar]
- Zhuo, R.; Zhang, J.; Yu, H.; Ma, F.; Zhang, X. The roles of Pleurotus ostreatus HAUCC 162 laccase isoenzymes in decolorization of synthetic dyes and the transformation pathways. Chemosphere 2019, 234, 733–745. [Google Scholar] [CrossRef]
- Pype, R.; Flahaut, S.; Debaste, F. On the importance of mechanisms analysis in the degradation of micropollutants by laccases: The case of Remazol Brilliant Blue R. Environ. Technol. Innov. 2019, 14, 100324. [Google Scholar] [CrossRef]
- Chicatto, J.A.; Rainert, K.T.; Gonçalves, M.J.; Helm, C.V.; Altmajer-Vaz, D.; Tavares, L.B.B. Decolorization of textile industry wastewater in solid state fermentation with Peach-Palm (Bactris gasipaes) residue. Braz. J. Biol. 2018, 78, 718–727. [Google Scholar] [CrossRef]
- Cardoso, B.K.; Linde, G.A.; Colauto, N.B.; Valle, J.S.D. Panus strigellus laccase decolorizes anthraquinone, azo, and triphenylmethane dyes. Biocatal. Agric. Biotechnol. 2018, 16, 558–563. [Google Scholar] [CrossRef]
- Isanapong, J.; Mataraj, S. Application of ligninolytic enzyme ofLentinus polychrouson synthetic dye decolorization. IOP Conf. Ser. Earth Environ. Sci. 2018, 185, 012004. [Google Scholar] [CrossRef]
- Garrido-Bazán, V.; Téllez-Téllez, M.; Herrera-Estrella, A.; Díaz-Godínez, G.; Nava-Galicia, S.; Villalobos-López, M.Á.; Arroyo-Becerra, A.; Bibbins-Martínez, M. Effect of textile dyes on activity and differential regulation of laccase genes from Pleurotus ostreatus grown in submerged fermentation. AMB Express 2016, 6, 1–9. [Google Scholar] [CrossRef]
- Lu, R.; Ma, L.; He, F.; Yu, D.; Fan, R.; Zhang, Y.; Long, Z.; Zhang, X.; Yang, Y. White-rot fungus Ganoderma sp.En3 had a strong ability to decolorize and tolerate the anthraquinone, indigo and triphenylmethane dye with high concentrations. Bioprocess Biosyst. Eng. 2015, 39, 381–390. [Google Scholar] [CrossRef] [PubMed]
- Sumandono, T.; Saragih, H.; Migirin; Watanabe, T.; Amirta, R. Decolorization of Remazol Brilliant Blue R by New Isolated White Rot Fungus Collected from Tropical Rain Forest in East Kalimantan and its Ligninolytic Enzymes Activity. Procedia Environ. Sci. 2015, 28, 45–51. [Google Scholar] [CrossRef]
- Grandes-Blanco, A.I.; Díaz-Godínez, G.; Téllez-Téllez, M.; Delgado-Macuil, R.; Rojas-López, M.; Bibbins-Martínez, M.D. LIGNINOLYTIC ACTIVITY PATTERNS OFPleurotus ostreatusOBTAINED BY SUBMERGED FERMENTATION IN PRESENCE OF 2,6-DIMETHOXYPHENOL AND REMAZOL BRILLIANT BLUE R DYE. Prep. Biochem. Biotechnol. 2013, 43, 468–480. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.-C.; Wu, P.-H.; Su, Y.-C.; Wen, T.-N.; Wei, Y.-S.; Wang, N.-C.; Hsu, C.-A.; Wang, A.H.-J.; Shyur, L.-F. Biochemical characterization of a novel laccase from the basidiomycete fungus Cerrena sp. WR1. Protein Eng. Des. Sel. 2012, 25, 761–769. [Google Scholar] [CrossRef] [PubMed]
- Sun, S. Decolourization characterizations of crude enzymes from a novel basidiomycete Mycena purpureofusca. Afr. J. Microbiol. Res. 2012, 6, 3501–3509. [Google Scholar] [CrossRef]
- Bibi, I.; Bhatti, H.N. Enhanced Biodecolorization of Reactive Dyes by Basidiomycetes Under Static Conditions. Appl. Biochem. Biotechnol. 2012, 166, 2078–2090. [Google Scholar] [CrossRef]
- Hadibarata, T.; Yusoff, A.R.M.; Kristanti, R.A. Decolorization and Metabolism of Anthraquionone-Type Dye by Laccase of White-Rot Fungi Polyporus sp. S133. Water Air Soil Pollut. 2012, 223, 933–941. [Google Scholar] [CrossRef]
- Zilly, A.; Coelho-Moreira, J.D.S.; Bracht, A.; De Souza, C.G.M.; Carvajal, A.E.; Koehnlein, E.A.; Peralta, R.M. Influence of NaCl and Na2SO4 on the kinetics and dye decolorization ability of crude laccase from Ganoderma lucidum. Int. Biodeterior. Biodegradation 2011, 65, 340–344. [Google Scholar] [CrossRef]
- Wang, Z.-X.; Cai, Y.; Liao, X.; Tao, G.-J.; Li, Y.-Y.; Zhang, F.; Zhang, D.-B. Purification and characterization of two thermostable laccases with high cold adapted characteristics from Pycnoporus sp. SYBC-L1. Process. Biochem. 2010, 45, 1720–1729. [Google Scholar] [CrossRef]
- Neifar, M.; Jaouani, A.; Ellouze-Ghorbel, R.; Ellouze-Chaabouni, S. Purification, characterization and decolourization ability of Fomes fomentarius laccase produced in solid medium. J. Mol. Catal. B Enzym. 2010, 64, 68–74. [Google Scholar] [CrossRef]
- Kanwal, N.; Asgher, M.; Bhatti, H.N.; Sheikh, M.A. Ligninase synthesis and decolorization of drimarine blue k2rl by Pleurotus ostreatus IBL-02 in carbon and nitrogen sufficient shake flask medium. Fresen. Environ. Bull. 2010, 19, 63–68. [Google Scholar]
- Neto, S.L.M.; Matheus, D.R.; Machado, K.M.G. Influence of pH on the growth, laccase activity and RBBR decolorization by tropical basidiomycetes. Braz. Arch. Biol. Technol. 2009, 52, 1075–1082. [Google Scholar] [CrossRef]
- Lu, L.; Zhao, M.; Zhang, B.-B.; Yu, S.-Y.; Bian, X.-J.; Wang, W.; Wang, Y. Purification and characterization of laccase from Pycnoporus sanguineus and decolorization of an anthraquinone dye by the enzyme. Appl. Microbiol. Biotechnol. 2007, 74, 1232–1239. [Google Scholar] [CrossRef]
- Palmieri, G.; Cennamo, G.; Sannia, G. Remazol Brilliant Blue R decolourisation by the fungus Pleurotus ostreatus and its oxidative enzymatic system. Enzym. Microb. Technol. 2005, 36, 17–24. [Google Scholar] [CrossRef]
- Soares, G.M.; Amorim, M.; Hrdina, R.; Costa-Ferreira, M. Studies on the biotransformation of novel disazo dyes by laccase. Process. Biochem. 2002, 37, 581–587. [Google Scholar] [CrossRef]
- Chagas, E.P.; Durrant, L.R. Decolorization of azo dyes by Phanerochaete chrysosporium and Pleurotus sajorcaju. Enzym. Microb. Technol. 2001, 29, 473–477. [Google Scholar] [CrossRef]
- Eichlerová, I.; Homolka, L.; Žifčáková, L.; Lisá, L.; Dobiášová, P.; Baldrian, P. Enzymatic systems involved in decomposition reflects the ecology and taxonomy of saprotrophic fungi. Fungal Ecol. 2015, 13, 10–22. [Google Scholar] [CrossRef]
- Tien, M.; Kirk, T.K. Lignin peroxidase of Phanerochaete chrysosporium. In Method Enzymol; Wood, W.A., Kellogg, T.S., Eds.; Academic Press: New York, NY, USA, 1988; Volume 161, pp. 238–248. Available online: https://www.sciencedirect.com/science/article/pii/0076687988610251 (accessed on 7 January 2004).
- Bourbonnais, R.; Paice, M.G. Oxidation of non-phenolic substrates. An expanded role for laccase in lignin biodegradation. FEBS Lett. 1990, 267, 99–102. [Google Scholar] [CrossRef]
- Ngo, T.; Lenhoff, H.M. A sensitive and versatile chromogenic assay for peroxidase and peroxidase-coupled reactions. Anal. Biochem. 1980, 105, 389–397. [Google Scholar] [CrossRef]
- Daniel, G.; Volc, J.; Kubatova, E. Pyranose Oxidase, a Major Source of H2O2 during Wood Degradation by Phanerochaete chrysosporium, Trametes versicolor, and Oudemansiella mucida. Appl. Environ. Microbiol. 1994, 60, 2524–2532. [Google Scholar] [CrossRef] [PubMed]
- Jarosz-Wilkołazka, A.; Rdest-Kochmańska, J.; Malarczyk, E.; Wardas, W.; Leonowicz, A. Fungi and their ability to decolorize azo and anthraquinonic dyes. Enzyme. Microbiol. Technol. 2002, 30, 566–572. [Google Scholar] [CrossRef]
- Novotný, Č.; Rawal, B.; Bhatt, M.; Patel, M.; Šašek, V.; Molitoris, H.P. Capacity of Irpex lacteus and Pleurotus ostreatus for decolorization of chemically different dyes. J. Biotechnol. 2001, 89, 113–122. [Google Scholar] [CrossRef]
- Casieri, L.; Anastasi, A.; Prigione, V.; Varese, G.C. Survey of ectomycorrhizal, litter-degrading, and wood-degrading Basidiomycetes for dye decolorization and ligninolytic enzyme activity. Antonie van Leeuwenhoek 2010, 98, 483–504. [Google Scholar] [CrossRef] [PubMed]
- Chander, M.; Arora, D. Evaluation of some white-rot fungi for their potential to decolourise industrial dyes. Dye Pigment. 2007, 72, 192–198. [Google Scholar] [CrossRef]
- Chander, M.; Arora, D.S.; Bath, H.K. Biodecolourisation of some industrial dyes by white-rot fungi. J. Ind. Microbiol. Biotechnol. 2004, 31, 94–97. [Google Scholar] [CrossRef]
- Eichlerová, I.; Homolka, L.; Nerud, F. Synthetic dye decolorization capacity of white rot fungus Dichomitus squalens. Bioresour. Technol. 2006, 97, 2153–2159. [Google Scholar] [CrossRef]
- Fu, Y.; Viraraghavan, T. Fungal decolorization of dye wastewaters: A review. Bioresour. Technol. 2001, 79, 251–262. [Google Scholar] [CrossRef]
- Paszczynski, A.; Pasti-Grigsby, M.B.; Goszczynski, S.; Crawford, R.L. Mineralization of sulfonated azo dyes and sulfanilic acid by Phanerochaete chrysosporium and Streptomyces chromofuscus. Appl. Environ. Microbiol. 1992, 58, 3598–3604. [Google Scholar] [CrossRef]
- Spadaro, J.T.; Gold, M.H.; Renganathan, V. Degradation of azo dyes by the lignin-degrading fungus Phanerochaete chrysosporium. Appl. Environ. Microbiol. 1992, 58, 2397–2401. [Google Scholar] [CrossRef]
- Swamy, J.; Ramsay, J. The evaluation of white rot fungi in the decoloration of textile dyes. Enzym. Microb. Technol. 1999, 24, 130–137. [Google Scholar] [CrossRef]
- Dias, A.; Bezerra, R.M.; Lemos, P.M.; Pereira, A.N. In vivo and laccase-catalyzed decolourization of xenobiotic azo dyes by a basidiomycetous fungus: Characterization of its ligninolytic system. World J. Microb. Biot. 2003, 19, 969–975. [Google Scholar] [CrossRef]
- Kunjadia, P.D.; Sanghvi, G.V.; Kunjadia, A.P.; Mukhopadhyay, P.N.; Dave, G.S. Role of ligninolytic enzymes of white rot fungi (Pleurotus spp.) grown with azo dyes. SpringerPlus 2016, 5, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Nyanhongo, G.; Gomes, J.; Gübitz, G.; Zvauya, R.; Read, J.; Steiner, W. Decolorization of textile dyes by laccases from a newly isolated strain of Trametes modesta. Water Res. 2002, 36, 1449–1456. [Google Scholar] [CrossRef]
- Harazono, K.; Watanabe, Y.; Nakamura, K. Decolorization of azo dye by white-rot basidiomycete Phanerochaete sordida and by its manganese peroxidase. J. Biosci. Bioeng. 2003, 5, 455–459. [Google Scholar] [CrossRef]
- Kariminiaae-Hamedaani, H.-R.; Sakurai, A.; Sakakibara, M. Decolorization of synthetic dyes by a new manganese peroxidase-producing white rot fungus. Dye. Pigment. 2007, 72, 157–162. [Google Scholar] [CrossRef]
- Moreira, M.T.; Palma, C.; Mielgo, I.; Feijoo, G.; Lema, J. In vitro degradation of a polymeric dye (Poly R-478) by manganese peroxidase. Biotechnol. Bioeng. 2001, 75, 362–368. [Google Scholar] [CrossRef]
- Wesenberg, D. White-rot fungi and their enzymes for the treatment of industrial dye effluents. Biotechnol. Adv. 2003, 22, 161–187. [Google Scholar] [CrossRef]
- Kotterman, M.; A Wasseveld, R.; A Field, J. Hydrogen Peroxide Production as a Limiting Factor in Xenobiotic Compound Oxidation by Nitrogen-Sufficient Cultures of Bjerkandera sp. Strain BOS55 Overproducing Peroxidases. Appl. Environ. Microbiol. 1996, 62, 880–885. [Google Scholar] [CrossRef]
- Reina, R.; Liers, C.; García-Romera, I.; Aranda, E. Enzymatic mechanisms and detoxification of dry olive-mill residue by Cyclocybe aegerita, Mycetinis alliaceus and Chondrostereum purpureum. Int. Biodeterior. Biodegradation 2017, 117, 89–96. [Google Scholar] [CrossRef]
- Heinfling, A.; Bergbauer, M.; Szewzyk, U. Biodegradation of azo and phthalocyanine dyes by Trametes versicolor and Bjerkandera adusta. Appl. Microbiol. Biotechnol. 1997, 48, 261–266. [Google Scholar] [CrossRef]
- Sodaneath, H.; Lee, J.-I.; Yang, S.-O.; Jung, H.; Ryu, H.W.; Cho, K.-S. Decolorization of textile dyes in an air-lift bioreactor inoculated with Bjerkandera adusta OBR105. J. Environ. Sci. Heal. Part A 2017, 52, 1099–1111. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Organism | Dye Decolorized | Enzyme Studied | Reference |
|---|---|---|---|
| Daedalea dickinsii | Disperse violet-63 Disperse orange-30 | Laccase, MnP | [19] |
| Pleurotus sajor-caju | Orange G | Laccase | [20] |
| Oudemansiella canarii | Congo red | Laccase | [21] |
| Clitopilus scyphoides, Ganoderma resinaceum, Schizophyllum sp. | Reactive Black 5 | MnP | [22] |
| Lenzites elegans | Congo Red | Laccase | [23] |
| Trametes versicolor | Orange G, Acid Orange 6 | Laccase | [24] |
| Trametes versicolor | Reactive Black 5 | Laccase | [25] |
| Pleurotus sajor caju | Congo Red | MnP | [26] |
| Phanerochaete chrysosporium | Congo Red | MnP | [27] |
| Marasmius cladophyllus | Orange G | Laccase | [28,29] |
| Kongo Red | |||
| Trametes trogii | Reactive Black 5, Reactive Violet 5 | Laccase | |
| Phanerochaete sordida | Reactive Black 5 | Laccase | [30] |
| Irpex lacteus | Remazol Brilliant Violet 5R, Direct Red 5B | MnP | [31] |
| Pleurotus ostreatus | Orange II, Reactive Black 5 (RB5), (DMP), Phenol Red | MnP | [32] |
| Coriolopsis gallica | Reactive Black 5, Bismark Brown R | Laccase | [33] |
| Coprinus comatus | Reactive Dark Blue KR, Reactive orange 1, Reactive Red X-3B, Congo red | Laccase | [34] |
| Hygrocybe sp., Lentinus bertieri, Lentinus villosus, Peniophora cinerea, Pleurotus flabellatus, Pleurotus ostreatus, Psilocybe castanella, Pycnoporus sanguineus, Rigidoporus microsporus, Trametes villosa, | Cibacron Red | Laccase | [35] |
| Pleurotus ostratus | Orange II, Reactive Black 5, Amaranth | MnP | [36] |
| Phanerochaete chrysosporium | Direct Red 80, Mordant -9 Blue | MnP | [37] |
| Pleurotus ostreatus | Congo Red | Laccase | [38] |
| Pleurotus ostreatus | Drimarene Brilliant Red K-4BL | Laccase | [12] |
| Phanerochaete. chrysosporium, Pycnoporus cinnabarinus | Congo Red, Xylidine Ponceau 2R | Laccase, MnP | [39] |
| Coriolopsis byrsina, Lentinus strigellus, Lentinus sp., Pycnoporus sanguineus, Phellinus rimosus | Orange II | Laccase | [40] |
| Cyathus bulleri | Reactive Orange 1 | Laccase | [41] |
| Coriolopsis polyzona, Hypholoma fasciculare, Pycnoporus sanguineus, Stropharia rugosoannulata, Trametes versicolor | Acid Red 299, Direct Blue 1, Direct Red 28, Disperse Red 1, Disperse Yellow 3, Reactive Black 5, Reactive Red 4, Reactive Yellow 81 | Laccase | [42] |
| Irpex lacteus | Reactive Orange 16 | MnP | [43] |
| Datronia caperata, Polyporus tenuiculus, Pycnoporus sanguineus, Hexagonia hirta | Orange II | Laccase | [44] |
| Ischnoderma resinosum | Reactive Black 5, Reactive Red 22, Reactive Yellow 15 | Laccase, MnP | [45] |
| Organism | Dye Decolorized | Enzyme Studied | References |
|---|---|---|---|
| Microporus vernicipes, Peniophora incarnata, Perenniporia subacida, Phanerochaete sordida, Phlebia acerina, Phlebia radiata. | RBBR | Laccase, MnP | [1] |
| Ganoderma lucidum | RBBR | Laccase | [46] |
| Trametes hirsuta | RBBR, Acid Blue 129 | Laccase, MnP | [47] |
| Ganoderma boninense, Ganoderma.miniatocinctum, Ganoderma zonatum, Ganoderma tornatum | RBBR | Laccase, MnP | [48] |
| Pleurotus ostreatus | RBBR, Bromfenol Blue | Laccase | [49] |
| Trametes versicolor | RBBR | Laccase | [50] |
| Clitopilus scyphoides, Ganoderma resinaceum, Schizophyllum sp. | RBBR | MnP | [22] |
| Ganoderma lucidum | RBBR | Laccase, MnP | [51] |
| Panus strigellus | RBBR, Reactive Blue 220 | Laccase | [52] |
| Lentinus polychrous | RBBR | Laccase | [53] |
| Trametes hirsuta, Phanerochaete chrysosporium, Pleurotus ostreatus | RBBR | Laccase, MnP | [11] |
| Marasmius cladophyllus | RBBR | Laccase | [28] |
| Pleurotus ostreatus | RBBR | Laccase | [54] |
| Ganoderma sp. | RBBR | Laccase | [55] |
| Phanerochaete chrysosporium, Ceriporiopsis subvermispora | RBBR | Laccase, MnP | [56] |
| Irpex lacteus | RBBR | MnP | [31] |
| Coriolopsis gallica | RBBR | Laccase | [33] |
| Coprinus comatus | RBBR, Reactive Brilliant Blue X-BR, Reactive Brilliant Blue K-GR, Reactive Brilliant Blue K-3R | Laccase | [34] |
| Pleurotus ostreatus | RBBR | Laccase, MnP | [57] |
| Cerrena sp. | RBBR | Laccase | [58] |
| Mycena purpureofusca | RBBR | Laccase | [59] |
| Trametes hirsuta, Pycnoporus sp., Irpex sp. | RBBR | Laccase | [60] |
| Polyporus sp. | RBBR | Laccase | [61] |
| Ganoderma lucidum | RBBR | Laccase | [62] |
| Pycnoporus sp. | RBBR | Laccase | [63] |
| Fomes fomentarius | RBBR | Laccase | [64] |
| Pleurotus ostreatus | Drimarene Blue K2RL | Laccase, MnP | [65] |
| Lentinus crinitus, Psilocybe castanella | RBBR | Laccase | [66] |
| Coriolopsis byrsina, Lentinus strigellus, Lentinus sp., Pycnoporus sanguineus, Phellinus rimosus | RBBR, Cibacron Blue 3GA | Laccase | [40] |
| Coriolopsis polyzona,Hypholoma fasciculare, Pycnoporus sanguineus, Stropharia rugosoannulata, Trametes versicolor | Acid Blue 62, Disperse Blue 1, Reactive Blue 19 | Laccase | [42] |
| Irpex lacteus | MnP | [43] | |
| Ischnoderma resinosum | Reactive Blue 19 | Laccase, MnP | [45] |
| Pycnoporus sanguineus | RBBR | Laccase | [67] |
| Pleurotus ostreatus | RBBR | Laccase | [68] |
| Ecophysiological Classification | Distribution of Taxonomical Groups in the Studied Set (n) | ||||
|---|---|---|---|---|---|
| Orders | Families | Genera | Species | Strains | |
| White rot | 4 | 13 | 33 | 52 | 105 |
| Brown rot | 2 | 3 | 4 | 4 | 7 |
| Litter decomposers | 1 | 7 | 15 | 21 | 38 |
| CCBAS Number | Genus | Species | Family | Order | Ecology | Lacc 1 | MnP 2 | Orange G | RBBR | ||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Decolorization 3 | Growth Rate 4 | Decolorization 3 | Growth Rate 4 | ||||||||
| 521 | Abortiporus | biennis | Meruliaceae | Polyporales | WR | 102.73 | 0.94 | 5 | 5 | 5 | 5 |
| 498 | Abortiporus | biennis | Meruliaceae | Polyporales | WR | 88.25 | 0.61 | 4 | 4 | 4 | 4 |
| 301 | Agaricus | arvensis | Agaricaceae | Agaricales | LD | 8.23 | 0.28 | 0 | 3 | 0 | 2 |
| 643 | Agrocybe | smithii | Strophariaceae | Agaricales | LD | 24.74 | 0.09 | 2 | 2 | 3 | 2 |
| 803 | Agrocybe | smithii | Strophariaceae | Agaricales | LD | 42.96 | 0.09 | 3 | 4 | 3 | 2 |
| 641 | Agrocybe | praecox | Strophariaceae | Agaricales | LD | 12.71 | 0.56 | 1 | 1 | 2 | 1 |
| 543 | Antrodia | heteromorpha | Fomitopsidaceae | Polyporales | BR | 0.14 | 0.00 | 0 | 5 | 0 | 4 |
| 542 | Antrodia | heteromorpha | Fomitopsidaceae | Polyporales | BR | 0.20 | 0.00 | 0 | 5 | 0 | 4 |
| 706 | Antrodia | heteromorpha | Fomitopsidaceae | Polyporales | BR | 0.04 | 0.00 | 0 | 4 | 0 | 3 |
| 331 | Armillaria | gemina | Physalacriaceae | Agaricales | WR | 5.50 | 1.50 | 0 | 1 | 1 | 1 |
| 330 | Armillaria | calvescens | Physalacriaceae | Agaricales | WR | 4.87 | 2.72 | 0 | 1 | 1 | 1 |
| 833 | Armillaria | calvescens | Physalacriaceae | Agaricales | WR | 6.86 | 0.84 | 0 | 1 | 1 | 1 |
| 558 | Ceriporia | camaresiana | Phanerochaetaceae | Agaricales | WR | 4.27 | 0.56 | 1 | 3 | 2 | 3 |
| 678 | Armillaria | sinapina | Physalacriaceae | Agaricales | WR | 3.11 | 0.09 | 0 | 1 | 0 | 1 |
| 344 | Clitopilus | passeckerianus | Entolomataceae | Agaricales | LD | 36.03 | 12.67 | 3 | 4 | 2 | 2 |
| 775 | Clitopilus | passeckerianus | Entolomataceae | Agaricales | LD | 66.15 | 8.94 | 4 | 4 | 2 | 4 |
| 775 | Clitopilus | passeckerianus | Entolomataceae | Agaricales | LD | 62.80 | 4.22 | 3 | 3 | 4 | 4 |
| 356 | Coprinellus | bisporus | Psathyrellaceae | Agaricales | LD | 13.24 | 0.84 | 0 | 2 | 1 | 1 |
| 357 | Coprinellus | bisporus | Psathyrellaceae | Agaricales | LD | 13.67 | 1.41 | 0 | 3 | 1 | 1 |
| 359 | Coprinellus | bisporus | Psathyrellaceae | Agaricales | LD | 10.25 | 0.00 | 0 | 3 | 1 | 1 |
| 358 | Coprinellus | bisporus | Psathyrellaceae | Agaricales | LD | 12.22 | 0.00 | 0 | 3 | 1 | 1 |
| 305 | Cyclocybe | aegerita | Strophariaceae | Agaricales | LD | 1.50 | 0.75 | 0 | 2 | 3 | 2 |
| 312 | Cyclocybe | aegerita | Strophariaceae | Agaricales | LD | 0.45 | 0.28 | 0 | 3 | 3 | 4 |
| 496 | Cyclocybe | aegerita | Strophariaceae | Agaricales | LD | 0.19 | 0.01 | 0 | 2 | 2 | 3 |
| 645 | Cyclocybe | erebia | Strophariaceae | Agaricales | LD | 67.50 | 0.94 | 3 | 4 | 3 | 3 |
| 811 | Cyclocybe | erebia | Strophariaceae | Agaricales | LD | 113.90 | 1.41 | 5 | 5 | 5 | 5 |
| 530 | Daedaleopsis | confragosa | Polyporaceae | Polyporales | WR | 6.28 | 0.08 | 4 | 5 | 4 | 4 |
| 795 | Daedaleopsis | confragosa | Polyporaceae | Polyporales | WR | 4.18 | 0.04 | 3 | 5 | 3 | 4 |
| 423 | Fayodia | gracilipes | Tricholomataceae | Agaricales | LD | 1.26 | 0.09 | 0 | 1 | 1 | 1 |
| 805 | Fayodia | gracilipes | Tricholomataceae | Agaricales | LD | 3.06 | 0.28 | 0 | 1 | 1 | 1 |
| 455 | Flammula | alnicola | Strophariaceae | Agaricales | WR | 17.18 | 4.88 | 3 | 4 | 2 | 1 |
| 836 | Flammulina | velutipes | Physalacriaceae | Agaricales | WR | 0.23 | 0.33 | 0 | 4 | 0 | 4 |
| 365 | Flammulina | velutipes | Physalacriaceae | Agaricales | WR | 0.93 | 0.56 | 0 | 5 | 0 | 5 |
| 262 | Fomitiporia | mediterranea | Hymenochaetaceae | Hymenochaetales | WR | 10.51 | 16.42 | 3 | 3 | 3 | 3 |
| 565 | Ganoderma | applanatum | Ganodermataceae | Polyporales | WR | 8.55 | 5.01 | 0 | 2 | 1 | 1 |
| 555 | Ganoderma | applanatum | Ganodermataceae | Polyporales | WR | 5.38 | 2.06 | 0 | 2 | 3 | 3 |
| 556 | Ganoderma | applanatum | Ganodermataceae | Polyporales | WR | 12.97 | 6.56 | 0 | 3 | 2 | 1 |
| 707 | Ganoderma | applanatum | Ganodermataceae | Polyporales | WR | 9.14 | 6.76 | 0 | 2 | 1 | 1 |
| 705 | Ganoderma | applanatum | Ganodermataceae | Polyporales | WR | 16.61 | 7.04 | 1 | 2 | 2 | 2 |
| 746 | Ganoderma | lucidum | Ganodermataceae | Polyporales | WR | 0.28 | 1.03 | 0 | 3 | 0 | 2 |
| 522 | Hapalopilus | croceus | Polyporaceae | Polyporales | WR | 4.50 | 0.47 | 0 | 3 | 0 | 1 |
| 663 | Hericium | coralloides | Hericiaceae | Russulales | WR | 10.59 | 1.03 | 1 | 1 | 1 | 1 |
| 548 | Hericium | coralloides | Hericiaceae | Russulales | WR | 44.04 | 0.72 | 2 | 3 | 2 | 2 |
| 654 | Hericium | erinaceus | Hericiaceae | Russulales | WR | 21.08 | 17.55 | 0 | 1 | 0 | 1 |
| 374 | Hohenbuehelia | auriscalpium | Pleurotaceae | Agaricales | WR | 69.41 | 0.19 | 0 | 2 | 1 | 2 |
| 373 | Hohenbuehelia | auriscalpium | Pleurotaceae | Agaricales | WR | 33.43 | 0.00 | 0 | 1 | 0 | 1 |
| 381 | Hypholoma | fasciculare | Strophariaceae | Agaricales | WR | 16.16 | 0.19 | 1 | 3 | 3 | 4 |
| 362 | Hypholoma | fasciculare | Strophariaceae | Agaricales | WR | 10.39 | 0.11 | 0 | 1 | 0 | 1 |
| 362 | Hypholoma | fasciculare | Strophariaceae | Agaricales | WR | 11.47 | 0.08 | 0 | 2 | 2 | 1 |
| 703 | Inocutis | dryophila | Hymenochaetaceae | Hymenochaetales | WR | 1.04 | 4.88 | 3 | 4 | 3 | 3 |
| 559 | Inonotus | obliquus | Hymenochaetaceae | Hymenochaetales | WR | 4.87 | 16.79 | 0 | 4 | 3 | 3 |
| 694 | Irpex | lacteus | Meruliaceae | Polyporales | WR | 0.15 | 0.20 | 0 | 1 | 0 | 1 |
| 369 | Irpex | lacteus | Meruliaceae | Polyporales | WR | 0.31 | 0.19 | 0 | 2 | 0 | 2 |
| 553 | Ischnoderma | benzoinum | Fomitopsidaceae | Polyporales | WR | 79.88 | 8.35 | 5 | 5 | 5 | 5 |
| 656 | Ischnoderma | benzoinum | Fomitopsidaceae | Polyporales | WR | 55.13 | 3.19 | 4 | 5 | 5 | 5 |
| 561 | Laetiporus | sulphureus | Fomitopsidaceae | Polyporales | BR | 0.14 | 0.00 | 0 | 4 | 0 | 4 |
| 681 | Laetiporus | sulphureus | Fomitopsidaceae | Polyporales | BR | 0.10 | 0.00 | 0 | 2 | 0 | 2 |
| 389 | Lentinula | edodes | Omphalotaceae | Agaricales | WR | 2.69 | 2.63 | 2 | 4 | 3 | 3 |
| 390 | Lentinula | edodes | Omphalotaceae | Agaricales | WR | 2.08 | 1.05 | 2 | 3 | 2 | 3 |
| 391 | Lentinula | edodes | Omphalotaceae | Agaricales | WR | 3.46 | 7.51 | 5 | 5 | 4 | 4 |
| 724 | Lentinula | edodes | Omphalotaceae | Agaricales | WR | 5.43 | 2.75 | 3 | 4 | 3 | 3 |
| 590 | Lenzites | tricolor | Polyporaceae | Polyporales | WR | 3.39 | 0.00 | 5 | 5 | 4 | 4 |
| 797 | Lepista | irina | Tricholomataceae | Agaricales | LD | 4.13 | 0.28 | 0 | 1 | 0 | 1 |
| 838 | Lepista | irina | Tricholomataceae | Agaricales | LD | 1.08 | 0.75 | 0 | 1 | 0 | 1 |
| 394 | Lepista | nuda | Tricholomataceae | Agaricales | LD | 0.48 | 0.09 | 0 | 2 | 0 | 1 |
| 761 | Lepista | sordida | Tricholomataceae | Agaricales | LD | 7.25 | 3.00 | 0 | 1 | 1 | 1 |
| 761 | Lepista | sordida | Tricholomataceae | Agaricales | LD | 9.23 | 5.35 | 0 | 3 | 1 | 2 |
| 842 | Leucoagaricus | bresadolae | Agaricaceae | Agaricales | LD | 56.05 | 1.13 | 0 | 4 | 3 | 4 |
| 802 | Leucoagaricus | bresadolae | Agaricaceae | Agaricales | LD | 93.47 | 1.41 | 0 | 3 | 2 | 3 |
| 405 | Lycoperdon | perlatum | Agaricaceae | Agaricales | LD | 1.67 | 0.28 | 3 | 2 | 2 | 1 |
| 516 | Lycoperdon | perlatum | Agaricaceae | Agaricales | LD | 2.18 | 1.69 | 5 | 5 | 5 | 5 |
| 439 | Mucidula | mucida | Physalacriaceae | Agaricales | WR | 12.93 | 5.72 | 4 | 5 | 4 | 5 |
| 816 | Mycena | crocata | Mycenaceae | Agaricales | LD | 1.89 | 0.00 | 0 | 2 | 1 | 1 |
| 817 | Mycena | polygramma | Mycenaceae | Agaricales | LD | 12.66 | 0.23 | 0 | 1 | 0 | 1 |
| 419 | Mycena | polygramma | Mycenaceae | Agaricales | LD | 15.54 | 0.00 | 0 | 1 | 0 | 1 |
| 520 | Mycena | polygramma | Mycenaceae | Agaricales | LD | 10.93 | 0.84 | 0 | 1 | 0 | 1 |
| 623 | Mycetinis | alliaceus | Omphalotaceae | Agaricales | LD | 0.65 | 16.7 | 0 | 2 | 0 | 2 |
| 343 | Omphalina | mutila | Tricholomataceae | Agaricales | LD | 17.69 | 20.36 | 0 | 4 | 4 | 4 |
| 388 | Omphalotus | japonicus | Omphalotaceae | Agaricales | WR | 0.39 | 0.84 | 0 | 3 | 0 | 2 |
| 708 | Onnia | tomentosa | Hymenochaetaceae | Hymenochaetales | WR | 6.09 | 14.92 | 0 | 3 | 0 | 2 |
| 616 | Oxyporus | latemarginatus | Schizoporaceae | Hymenochaetales | WR | 3.70 | 12.57 | 3 | 5 | 5 | 5 |
| 810 | Oxyporus | latemarginatus | Schizoporaceae | Hymenochaetales | WR | 1.55 | 7.69 | 5 | 5 | 5 | 5 |
| 615 | Oxyporus | latemarginatus | Schizoporaceae | Hymenochaetales | WR | 1.16 | 10.23 | 3 | 4 | 4 | 4 |
| 276 | Phellinus | hartigii | Hymenochaetaceae | Hymenochaetales | WR | 0.32 | 0.00 | 0 | 5 | 0 | 3 |
| 575 | Phellinus | igniarius | Hymenochaetaceae | Hymenochaetales | WR | 0.45 | 0.94 | 0 | 3 | 0 | 3 |
| 577 | Phellinus | igniarius | Hymenochaetaceae | Hymenochaetales | WR | 0.23 | 2.20 | 0 | 1 | 0 | 1 |
| 269 | Phellinus | igniarius | Hymenochaetaceae | Hymenochaetales | WR | 0.51 | 6.29 | 0 | 5 | 5 | 5 |
| 657 | Phellinus | igniarius | Hymenochaetaceae | Hymenochaetales | WR | 1.45 | 2.35 | 0 | 3 | 2 | 3 |
| 758 | Phellinus | igniarius | Hymenochaetaceae | Hymenochaetales | WR | 2.93 | 0.84 | 0 | 1 | 1 | 1 |
| 274 | Phellinus | igniarius | Hymenochaetaceae | Hymenochaetales | WR | 4.50 | 1.97 | 0 | 1 | 1 | 1 |
| 265 | Phelinus | pomaceus | Hymenochaetaceae | Hymenochaetales | WR | 1.16 | 13.32 | 3 | 3 | 1 | 1 |
| 587 | Phellinus | robustus | Hymenochaetaceae | Hymenochaetales | WR | 8.71 | 16.14 | 0 | 3 | 3 | 2 |
| 715 | Phlebia | chrysocreas | Meruliaceae | Polyporales | WR | 7.93 | 0.00 | 0 | 1 | 0 | 1 |
| 846 | Pholiota | adiposa | Strophariaceae | Agaricales | WR | 3.57 | 10.04 | 3 | 4 | 2 | 3 |
| 847 | Pholiota | adiposa | Strophariaceae | Agaricales | WR | 1.09 | 2.68 | 2 | 1 | 2 | 1 |
| 683 | Pholiota | adiposa | Strophariaceae | Agaricales | WR | 1.95 | 4.41 | 3 | 3 | 2 | 3 |
| 780 | Pholiota | aurivella | Strophariaceae | Agaricales | WR | 56.08 | 10.70 | 4 | 4 | 3 | 3 |
| 849 | Pholiota | aurivella | Strophariaceae | Agaricales | WR | 45.58 | 9.05 | 3 | 3 | 3 | 2 |
| 450 | Pleurotus | calyptratus | Pleurotaceae | Agaricales | WR | 1.21 | 0.28 | 0 | 3 | 2 | 1 |
| 691 | Pleurotus | citrinopileatus | Pleurotaceae | Agaricales | WR | 5.50 | 6.85 | 0 | 4 | 0 | 1 |
| 564 | Pleurotus | cornucopiae | Pleurotaceae | Agaricales | WR | 2.16 | 0.38 | 0 | 3 | 1 | 4 |
| 464 | Pleurotus | cornucopiae | Pleurotaceae | Agaricales | WR | 3.08 | 1.13 | 2 | 2 | 3 | 3 |
| 466 | Pleurotus | cystidiosus | Pleurotaceae | Agaricales | WR | 13.3 | 9.19 | 0 | 2 | 2 | 3 |
| 461 | Pleurotus | djamor | Pleurotaceae | Agaricales | WR | 31.08 | 1.78 | 4 | 4 | 3 | 3 |
| 666 | Pleurotus | djamor | Pleurotaceae | Agaricales | WR | 88.86 | 4.50 | 5 | 5 | 5 | 5 |
| 468 | Pleurotus | dryinus | Pleurotaceae | Agaricales | WR | 14.04 | 13.89 | 0 | 1 | 1 | 1 |
| 372 | Pleurotus | eryngii | Pleurotaceae | Agaricales | WR | 1.24 | 2.35 | 0 | 3 | 0 | 1 |
| 754 | Pleurotus | eryngii | Pleurotaceae | Agaricales | WR | 1.08 | 1.03 | 0 | 1 | 0 | 1 |
| 408 | Pleurotus | eryngii | Pleurotaceae | Agaricales | WR | 2.40 | 3.38 | 0 | 3 | 1 | 3 |
| 354 | Pleurotus | eryngii | Pleurotaceae | Agaricales | WR | 0.29 | 0.19 | 0 | 1 | 0 | 1 |
| 544 | Pleurotus | eryngii | Pleurotaceae | Agaricales | WR | 1.38 | 4.22 | 0 | 1 | 2 | 2 |
| 819 | Pleurotus | eryngii | Pleurotaceae | Agaricales | WR | 1.45 | 4.32 | 0 | 3 | 2 | 3 |
| 625 | Pleurotus | eryngii | Pleurotaceae | Agaricales | WR | 1.01 | 0.28 | 0 | 2 | 0 | 2 |
| 830 | Pleurotus | eryngii | Pleurotaceae | Agaricales | WR | 0.23 | 2.25 | 0 | 1 | 0 | 1 |
| 843 | Pleurotus | eryngii | Pleurotaceae | Agaricales | WR | 4.42 | 3.50 | 0 | 3 | 3 | 3 |
| 692 | Pleurotus | ostreatus | Pleurotaceae | Agaricales | WR | 66.78 | 1.13 | 4 | 4 | 4 | 4 |
| 462 | Pleurotus | ostreatus | Pleurotaceae | Agaricales | WR | 106.68 | 6.94 | 5 | 5 | 3 | 3 |
| 473 | Pleurotus | ostreatus | Pleurotaceae | Agaricales | WR | 72.95 | 6.94 | 3 | 4 | 5 | 5 |
| 766 | Pleurotus | ostreatus | Pleurotaceae | Agaricales | WR | 50.37 | 2.28 | 2 | 3 | 3 | 3 |
| 479 | Pleurotus | pulmonarius | Pleurotaceae | Agaricales | WR | 47.49 | 1.03 | 5 | 5 | 3 | 3 |
| 589 | Polyporus | brumalis | Polyporaceae | Polyporales | WR | 0.03 | 15.48 | 0 | 2 | 1 | 1 |
| 818 | Polyporus | brumalis | Polyporaceae | Polyporales | WR | 0.01 | 10.00 | 0 | 2 | 0 | 2 |
| 591 | Polyporus | ciliatus | Polyporaceae | Polyporales | WR | 8.27 | 2.72 | 0 | 1 | 2 | 1 |
| 598 | Polyporus | lepideus | Polyporaceae | Polyporales | WR | 2.29 | 1.16 | 3 | 2 | 3 | 2 |
| 608 | Polyporus | lepideus | Polyporaceae | Polyporales | WR | 4.69 | 0.14 | 5 | 5 | 5 | 5 |
| 534 | Polyporus | lepideus | Polyporaceae | Polyporales | WR | 3.04 | 0.28 | 3 | 2 | 3 | 2 |
| 676 | Polyporus | squamosus | Polyporaceae | Polyporales | WR | 0.37 | 1.29 | 0 | 2 | 0 | 2 |
| 261 | Porodaedalea | pini | Hymenochaetaceae | Hymenochaetales | WR | 1.27 | 0.56 | 3 | 3 | 3 | 3 |
| 735 | Porodaedalea | pini | Hymenochaetaceae | Hymenochaetales | WR | 0.91 | 1.26 | 3 | 3 | 2 | 2 |
| 261 | Porodaedalea | pini | Hymenochaetaceae | Hymenochaetales | WR | 1.25 | 0.38 | 2 | 2 | 2 | 2 |
| 492 | Psilocybe | subaeruginosa | Strophariaceae | Agaricales | LD | 10.20 | 0.19 | 0 | 1 | 1 | 1 |
| 488 | Psilocybe | subaeruginosa | Strophariaceae | Agaricales | LD | 14.44 | 0.28 | 0 | 1 | 1 | 1 |
| 595 | Pycnoporus | sanguineus | Polyporaceae | Polyporales | WR | 33.77 | 3.03 | 4 | 5 | 5 | 5 |
| 596 | Pycnoporus | sanguineus | Polyporaceae | Polyporales | WR | 56.92 | 9.01 | 5 | 5 | 5 | 5 |
| 702 | Rhodocollybia | butyracea | Omphalotaceae | Agaricales | LD | 54.6 | 0.28 | 0 | 1 | 2 | 1 |
| 349 | Rhodocollybia | maculata | Omphalotaceae | Agaricales | LD | 4.39 | 0.19 | 0 | 1 | 0 | 1 |
| 110 | Serpula | himantioides | Serpulaceae | Boletales | BR | 0.00 | 0.00 | 0 | 5 | 0 | 5 |
| 752 | Schizophyllum | commune | Schizophyllaceae | Agaricales | WR | 0.14 | 0.47 | 0 | 5 | 0 | 4 |
| 658 | Sparassis | crispa | Sparassidaceae | Polyporales | BR | 0.00 | 0.00 | 0 | 1 | 0 | 1 |
| 524 | Stereum | gausapatum | Stereaceae | Russulales | WR | 0.39 | 0.19 | 0 | 3 | 0 | 3 |
| 610 | Trametes | hirsuta | Polyporaceae | Polyporales | WR | 24.23 | 5.54 | 5 | 5 | 5 | 5 |
| 611 | Trametes | versicolor | Polyporaceae | Polyporales | WR | 2.85 | 7.88 | 5 | 5 | 3 | 4 |
| 614 | Trametes | versicolor | Polyporaceae | Polyporales | WR | 25.05 | 6.38 | 5 | 5 | 5 | 5 |
| 528 | Trametes | versicolor | Polyporaceae | Polyporales | WR | 47.84 | 11.82 | 4 | 5 | 4 | 5 |
| 704 | Trametes | versicolor | Polyporaceae | Polyporales | WR | 3.56 | 7.60 | 4 | 5 | 4 | 5 |
| 612 | Trametes | versicolor | Polyporaceae | Polyporales | WR | 63.54 | 1.60 | 5 | 5 | 5 | 5 |
| 750 | Tricholoma | sejunctum | Tricholomataceae | Agaricales | LD | 11.24 | 0.00 | 5 | 5 | 5 | 5 |
| 673 | Tyromyces | chioneus | Polyporaceae | Polyporales | WR | 0.54 | 4.32 | 0 | 2 | 2 | 2 |
| 267 | Tyromyces | chioneus | Polyporaceae | Polyporales | WR | 0.85 | 3.57 | 2 | 3 | 2 | 3 |
| 277 | Tyromyces | chinoeus | Polyporaceae | Polyporales | WR | 0.12 | 1.13 | 0 | 1 | 0 | 1 |
| Number of Strains (n) | ||||||||
|---|---|---|---|---|---|---|---|---|
| Order | n | Family | n | High Ligninolytic Enzyme Activity and Decolorization Capacity | ||||
| Laccase | MnP | Orange G | RBBR | Orange G and RBBR | ||||
| Agaricales | 85 | Agaricaceae | 5 | 2 a | 0 b | 2/1 c | 2/1 c | 1/1 c |
| Agaricales | Entolomataceae | 3 | 3 | 3 | 3/0 | 1/0 | 1/0 | |
| Agaricales | Mycenaceae | 4 | 0 | 0 | 0/0 | 0/0 | 0/0 | |
| Agaricales | Omphalotaceae | 8 | 1 | 2 | 2/1 | 3/0 | 2/0 | |
| Agaricales | Phanerochaeteceae | 1 | 0 | 0 | 0/0 | 0/0 | 0/0 | |
| Agaricales | Physalacriaceae | 7 | 0 | 1 | 1/0 | 1/0 | 1/0 | |
| Agaricales | Pleurotaceae | 24 | 9 | 10 | 6/3 | 9/2 | 6/1 | |
| Agaricales | Psathyrellaceae | 4 | 0 | 0 | 0/0 | 0/0 | 0/0 | |
| Agaricales | Schizophyllaceae | 1 | 0 | 0 | 0/0 | 0/0 | 0/0 | |
| Agaricales | Strophariaceae | 19 | 5 | 5 | 8/1 | 9/1 | 5/1 | |
| Agaricales | Tricholomataceae | 9 | 0 | 2 | 1/1 | 2/1 | 1/1 | |
| Hymenochaetales | 19 | Hymenochaetaceae | 16 | 0 | 7 | 5/0 | 3/0 | 3/0 |
| Hymenochaetales | Schizoporaceae | 3 | 0 | 3 | 3/1 | 3/2 | 3/1 | |
| Polyporales | 41 | Fomitopsidaceae | 7 | 2 | 2 | 2/1 | 2/2 | 2/1 |
| Polyporales | Ganodermataceae | 6 | 0 | 4 | 0/0 | 0/0 | 0/0 | |
| Polyporales | Meruliaceae | 5 | 2 | 0 | 2/1 | 2/1 | 2/1 | |
| Polyporales | Polyporaceae | 22 | 4 | 11 | 14/7 | 14/6 | 14/5 | |
| Polyporales | Sparassidaceae | 1 | 0 | 0 | 0/0 | 0/0 | 0/0 | |
| Russulales | 4 | Hericiaceae | 3 | 1 | 1 | 0/0 | 0/0 | 0/0 |
| Russulales | Stereaceae | 1 | 0 | 0 | 0/0 | 0/0 | 0/0 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Eichlerová, I.; Baldrian, P. Ligninolytic Enzyme Production and Decolorization Capacity of Synthetic Dyes by Saprotrophic White Rot, Brown Rot, and Litter Decomposing Basidiomycetes. J. Fungi 2020, 6, 301. https://doi.org/10.3390/jof6040301
Eichlerová I, Baldrian P. Ligninolytic Enzyme Production and Decolorization Capacity of Synthetic Dyes by Saprotrophic White Rot, Brown Rot, and Litter Decomposing Basidiomycetes. Journal of Fungi. 2020; 6(4):301. https://doi.org/10.3390/jof6040301
Chicago/Turabian StyleEichlerová, Ivana, and Petr Baldrian. 2020. "Ligninolytic Enzyme Production and Decolorization Capacity of Synthetic Dyes by Saprotrophic White Rot, Brown Rot, and Litter Decomposing Basidiomycetes" Journal of Fungi 6, no. 4: 301. https://doi.org/10.3390/jof6040301
APA StyleEichlerová, I., & Baldrian, P. (2020). Ligninolytic Enzyme Production and Decolorization Capacity of Synthetic Dyes by Saprotrophic White Rot, Brown Rot, and Litter Decomposing Basidiomycetes. Journal of Fungi, 6(4), 301. https://doi.org/10.3390/jof6040301