
Overexpression of OsCSP41b Enhances Rice Tolerance to Sheath Blight Caused by Rhizoctonia solani
 , and
, and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Materials and Growth Conditions
2.2. Inoculation and Evaluation of Rice ShB Resistance
2.3. Stress and Hormone Treatments
2.4. RNA Extraction and RT-qPCR Analysis
2.5. Phylogenetic Analysis
2.6. Subcellular Localization of OsCSP41b
2.7. Protein Extraction and Western Blot Analysis
2.8. Chlorophyll Content Measurement
2.9. Evaluation of Agronomic Traits in the Field
2.10. RNA-Seq and Data Analysis
3. Results
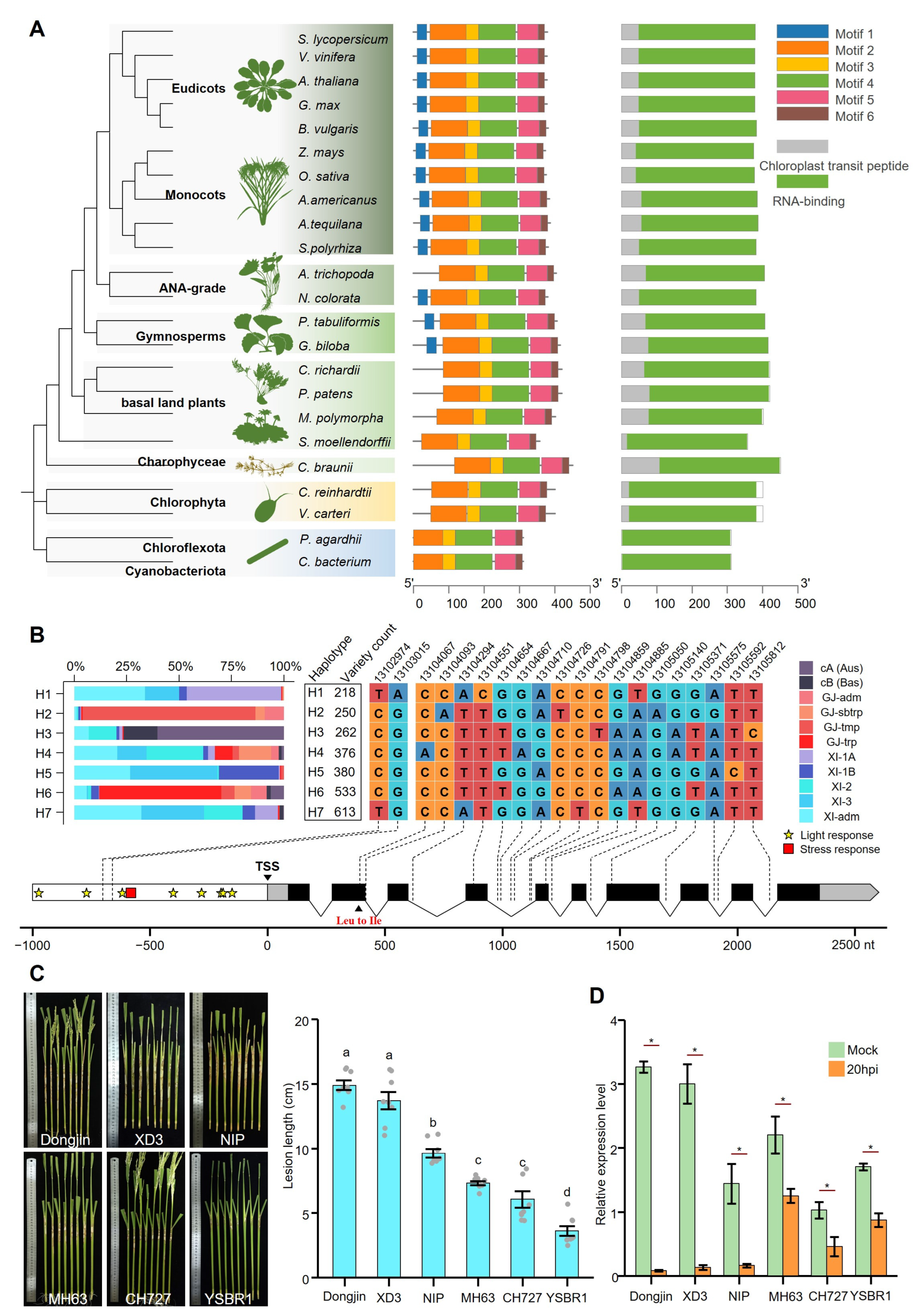
3.1. CSP41b Is Highly Conserved Among Plants and Suppressed Expression by R. solani Infection
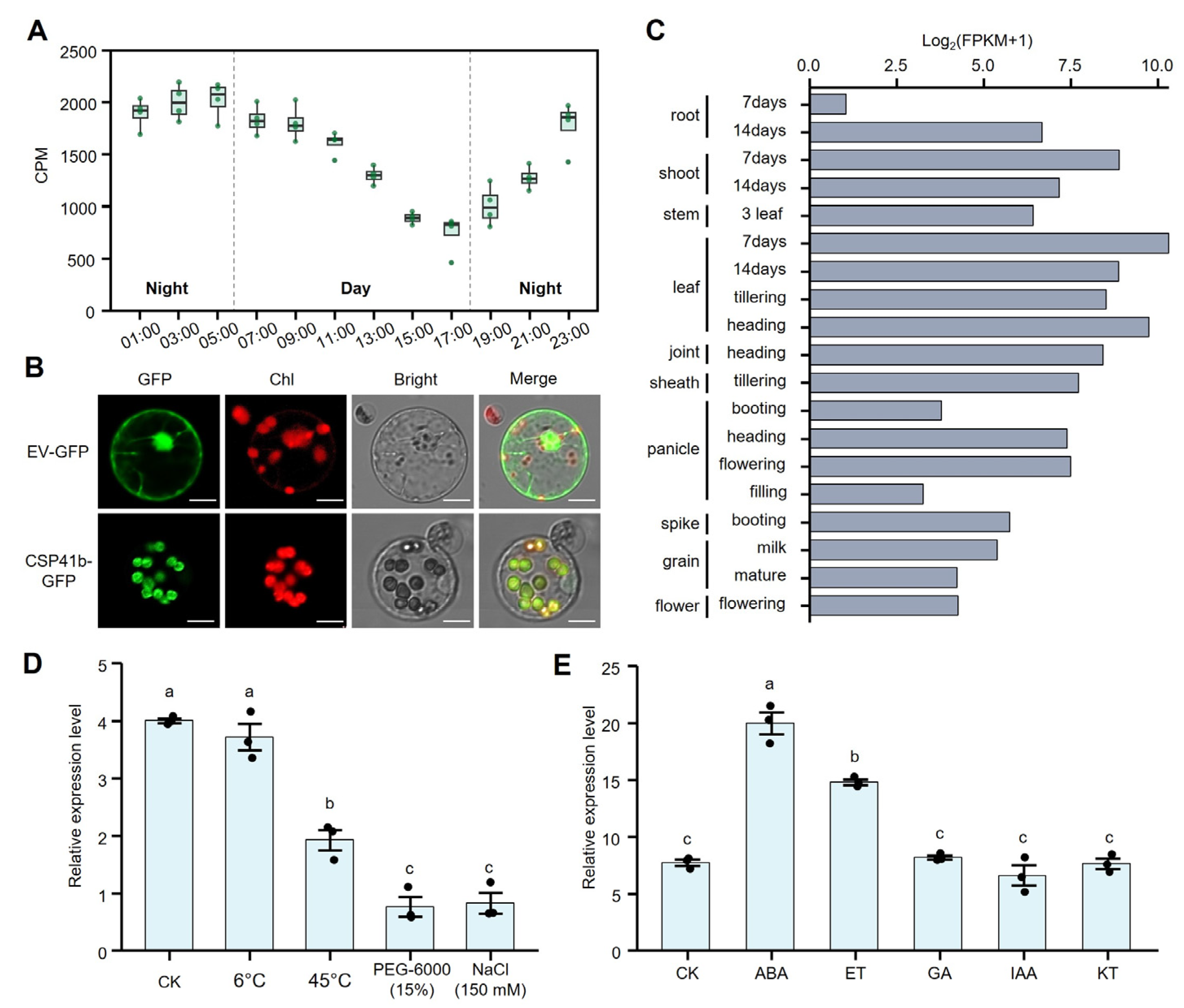
3.2. Expression Pattern and Subcellular Localization of OsCSP41b in Rice
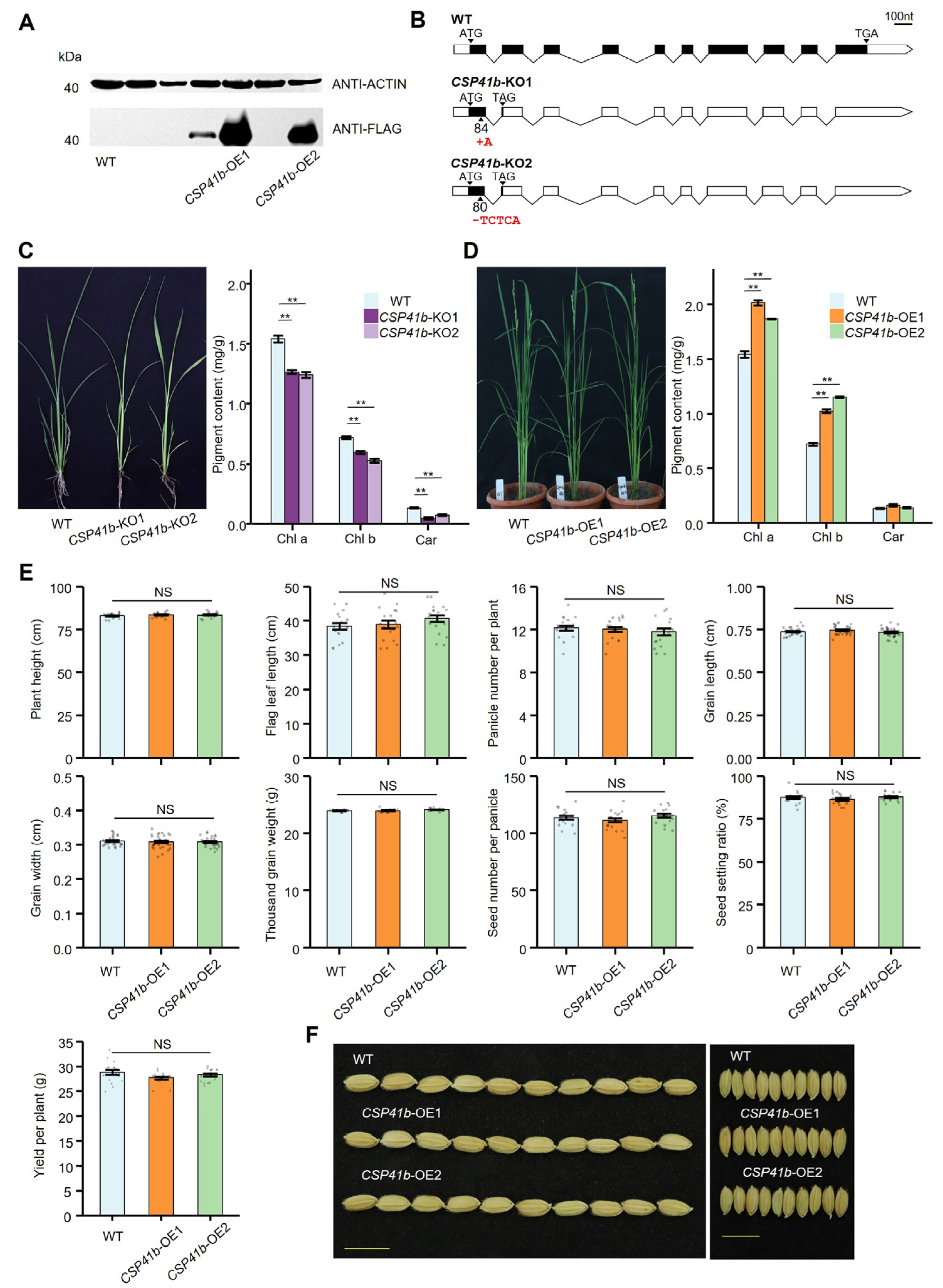
3.3. Knockout of OsCSP41b Significantly Affects Rice Growth While Its Overexpression Displays Comparable Phenotype and Yield-Associated Traits to the Wild Type
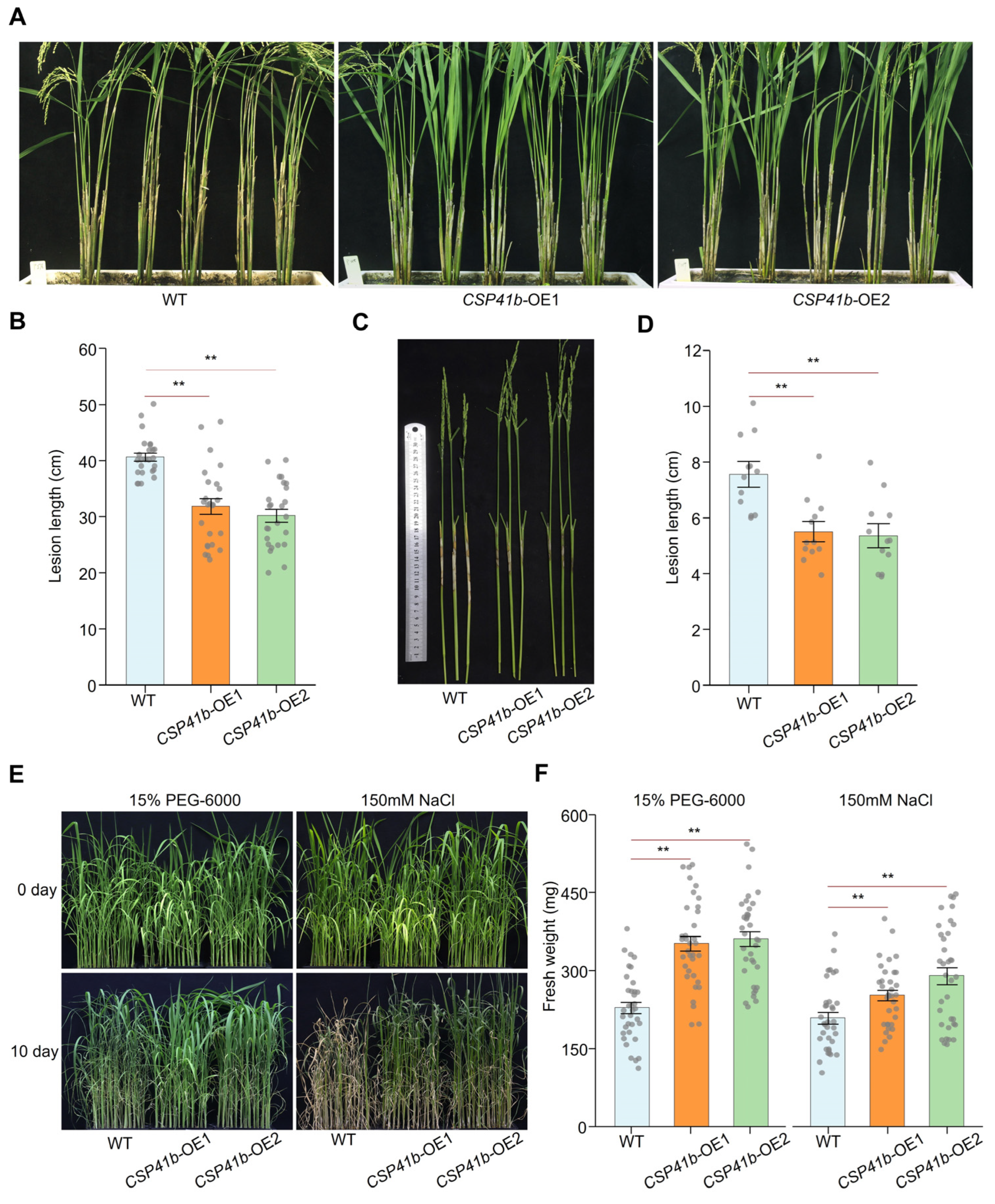
3.4. OsCSP41b Overexpression Enhances Rice Tolerance to R. solani, as Well as to Drought and Salinity
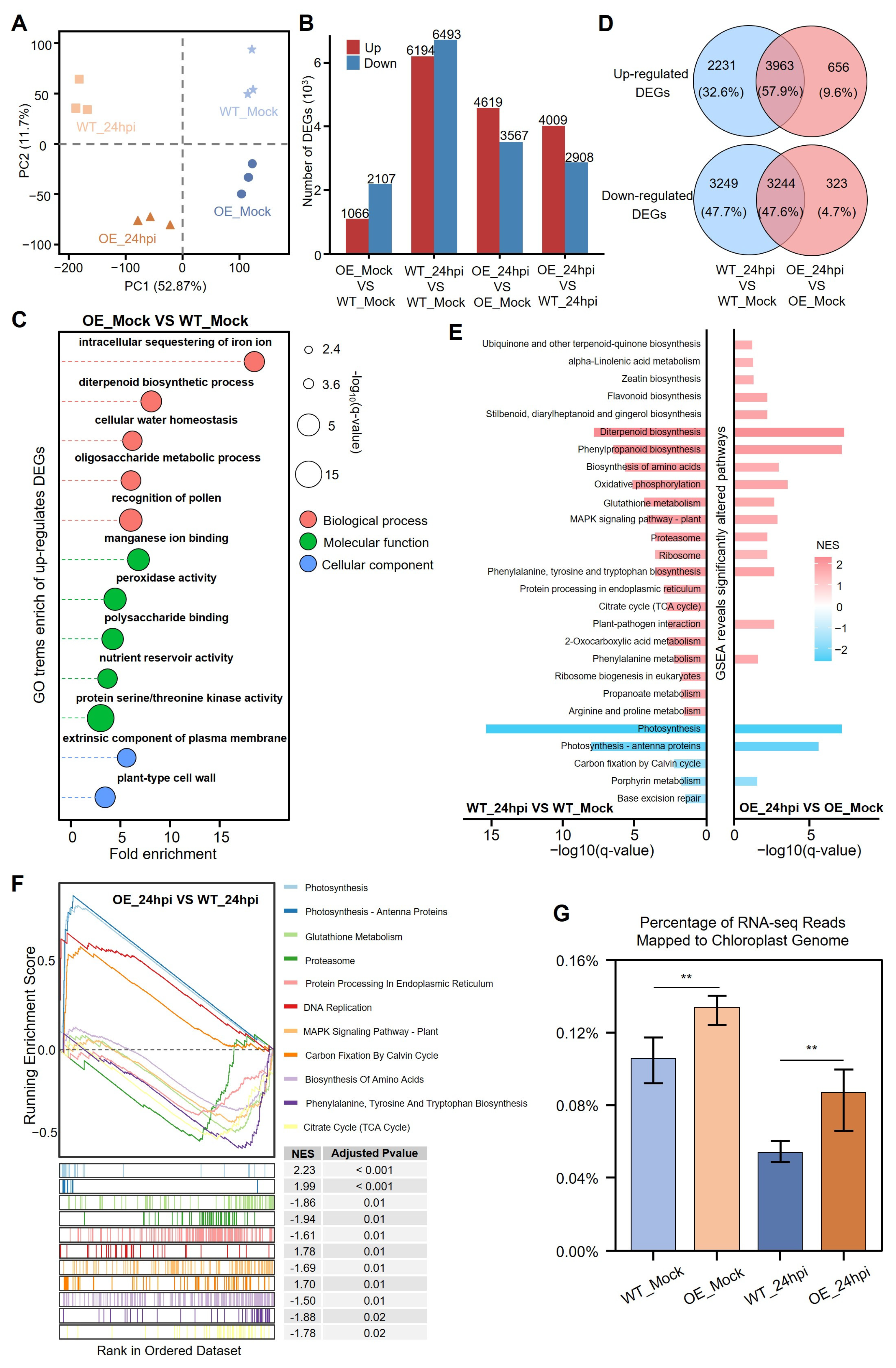
3.5. Chloroplast Transcripts and Photosynthesis-Associated Genes Show More Stabilization in CSP41b-OE Lines than WT After Infection with R. solani
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| ShB | Sheath blight |
| OsCSP41b | Chloroplast stem-loop-binding protein of 41 kDa b |
| WT | Wild type |
| OE | Overexpressing |
| KO | Knockout |
| dpi | Days post-inoculation |
| hpi | Hours post-inoculation |
| PCA | Principal component analysis |
| DEGs | Differentially expressed genes |
| GSEA | Gene set enrichment analysis |
| GO | Gene ontology |
| KEGG | Kyoto encyclopedia of genes and genome |
| QTL | Quantitative trait loci |
| ROS | Reactive oxygen species |
References
- Cao, W.; Zhang, H.; Zhou, Y.; Zhao, J.; Lu, S.; Wang, X.; Chen, X.; Yuan, L.; Guan, H.; Wang, G.; et al. Suppressing Chlorophyll Degradation by Silencing OsNYC3 Improves Rice Resistance to Rhizoctonia Solani, the Causal Agent of Sheath Blight. Plant Biotechnol. J. 2022, 20, 335–349. [Google Scholar] [CrossRef] [PubMed]
- Molla, K.A.; Karmakar, S.; Molla, J.; Bajaj, P.; Varshney, R.K.; Datta, S.K.; Datta, K. Understanding Sheath Blight Resistance in Rice: The Road behind and the Road Ahead. Plant Biotechnol. J. 2019, 18, 895–915. [Google Scholar] [CrossRef] [PubMed]
- Singh, P.; Mazumdar, P.; Harikrishna, J.A.; Babu, S. Sheath Blight of Rice: A Review and Identification of Priorities for Future Research. Planta 2019, 250, 1387–1407. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Ji, Z.; Kou, Y.; Yang, X.; Huang, W.; Sun, Z.; Zuo, S.; Li, Z.; Xiong, Y.; Deng, Y.; et al. Occurrence and Integrated Control of Major Rice Diseases in China. New Plant Protect. 2025, 2, e70004. [Google Scholar] [CrossRef]
- Chen, X.J.; Chen, Y.; Zhang, L.N.; Xu, B.; Zhang, J.H.; Chen, Z.X.; Tong, Y.H.; Zuo, S.M.; Xu, J.Y. Overexpression of OsPGIP1 Enhances Rice Resistance to Sheath Blight. Plant Dis. 2016, 100, 388–395. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Sun, Q.; Zhao, J.; Liu, T.; Du, H.; Shan, W.; Wu, K.; Xue, X.; Yang, C.; Liu, J.; et al. Fine Mapping and Candidate Gene Analysis of qSB12YSB, a Gene Conferring Major Quantitative Resistance to Rice Sheath Blight. Theor. Appl. Genet. 2023, 136, 246. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Xuan, Y.; Yi, J.; Xiao, G.; Yuan, P.; Li, D. Progress in Rice Sheath Blight Resistance Research. Front. Plant Sci. 2023, 14, 1141697. [Google Scholar] [CrossRef] [PubMed]
- Helliwell, E.E.; Wang, Q.; Yang, Y. Transgenic Rice with Inducible Ethylene Production Exhibits Broad-spectrum Disease Resistance to the Fungal Pathogens Magnaporthe Oryzae and Rhizoctonia Solani. Plant Biotechnol. J. 2012, 11, 33–42. [Google Scholar] [CrossRef] [PubMed]
- Xue, X.; Cao, Z.X.; Zhang, X.T.; Wang, Y.; Zhang, Y.F.; Chen, Z.X.; Pan, X.B.; Zuo, S.M. Overexpression of OsOSM1 Enhances Resistance to Rice Sheath Blight. Plant Dis. 2016, 100, 1634–1642. [Google Scholar] [CrossRef] [PubMed]
- Zhu, H.; Zhou, T.; Guan, J.; Li, Z.; Yang, X.; Li, Y.; Sun, J.; Xu, Q.; Xuan, Y.H. Precise Genome Editing of Dense and Erect Panicle 1 Promotes Rice Sheath Blight Resistance and Yield Production in Japonica Rice. Plant Biotechnol. J. 2025, 23, 1832–1846. [Google Scholar] [CrossRef] [PubMed]
- Yuhao, X.; Wenya, X.; Jianhua, Z.; Xiang, X.; Wenlei, C.; Xiaopin, S.; Zhou, W.; Yiwen, W.; Guangda, W.; Zhiming, F.; et al. OsERF7 Negatively Regulates Resistance to Sheath Blight Disease by Inhibiting Phytoalexin Biosynthesis. Rice Sci. 2025, 32, 367–379. [Google Scholar] [CrossRef]
- Xie, W.; Cao, W.; Lu, S.; Zhao, J.; Shi, X.; Yue, X.; Wang, G.; Feng, Z.; Hu, K.; Chen, Z.; et al. Knockout of Transcription Factor OsERF65 Enhances ROS Scavenging Ability and Confers Resistance to Rice Sheath Blight. Mol. Plant Pathol. 2023, 24, 1535–1551. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Tan, J.; Wu, Z.; VandenLangenberg, K.; Wehner, T.C.; Wen, C.; Zheng, X.; Owens, K.; Thornton, A.; Bang, H.H.; et al. STAYGREEN, STAY HEALTHY: A Loss-of-susceptibility Mutation in the STAYGREEN Gene Provides Durable, Broad-spectrum Disease Resistances for over 50 Years of US Cucumber Production. New Phytol. 2018, 221, 415–430. [Google Scholar] [CrossRef] [PubMed]
- Xie, W.; Xue, X.; Wang, Y.; Zhang, G.; Zhao, J.; Zhang, H.; Wang, G.; Li, L.; Wang, Y.; Shan, W.; et al. Natural Mutation in Stay-Green (OsSGR) Confers Enhanced Resistance to Rice Sheath Blight through Elevating Cytokinin Content. Plant Biotechnol. J. 2024, 23, 807–823. [Google Scholar] [CrossRef] [PubMed]
- de Torres Zabala, M.; Littlejohn, G.; Jayaraman, S.; Studholme, D.; Bailey, T.; Lawson, T.; Tillich, M.; Licht, D.; Bölter, B.; Delfino, L.; et al. Chloroplasts Play a Central Role in Plant Defence and Are Targeted by Pathogen Effectors. Nat. Plants 2015, 1, 15074. [Google Scholar] [CrossRef] [PubMed]
- Akhtar, S.S.; Mekureyaw, M.F.; Pandey, C.; Roitsch, T. Role of Cytokinins for Interactions of Plants with Microbial Pathogens and Pest Insects. Front. Plant Sci. 2020, 10, 1777. [Google Scholar] [CrossRef] [PubMed]
- Bürger, M.; Chory, J. Stressed Out About Hormones: How Plants Orchestrate Immunity. Cell Host Microbe 2019, 26, 163–172. [Google Scholar] [CrossRef] [PubMed]
- Pitsili, E.; Phukan, U.J.; Coll, N.S. Cell Death in Plant Immunity. Cold Spring Harb. Perspect. Biol. 2019, 12, a036483. [Google Scholar] [CrossRef] [PubMed]
- Mei, J.; Li, F.; Liu, X.; Hu, G.; Fu, Y.; Liu, W. Newly Identified CSP41b Gene Localized in Chloroplasts Affects Leaf Color in Rice. Plant Sci. 2016, 256, 39–45. [Google Scholar] [CrossRef] [PubMed]
- Ariga, H.; Tanaka, T.; Ono, H.; Sakata, Y.; Hayashi, T.; Taji, T. CSP41b, a Protein Identified via FOX Hunting Using Eutrema Salsugineum cDNAs, Improves Heat and Salinity Stress Tolerance in Transgenic Arabidopsis Thaliana. Biochem. Biophys. Res. Commun. 2015, 464, 318–323. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.J.; Zhang, Q. Optimising the Tissue Culture Conditions for High Efficiency Transformation of Indica Rice. Plant Cell Rep. 2004, 23, 540–547. [Google Scholar] [CrossRef] [PubMed]
- Willocquet, L.; Lore, J.S.; Srinivasachary, S.; Savary, S. Quantification of the Components of Resistance to Rice Sheath Blight Using a Detached Tiller Test Under Controlled Conditions. Plant Dis. 2011, 95, 1507–1515. [Google Scholar] [CrossRef] [PubMed]
- Luo, Y.; Fang, B.; Wang, W.; Yang, Y.; Rao, L.; Zhang, C. Genome-Wide Analysis of the Rice J-Protein Family: Identification, Genomic Organization, and Expression Profiles under Multiple Stresses. 3 Biotech 2019, 9, 358. [Google Scholar] [CrossRef] [PubMed]
- Zhong, Y.; Luo, Y.; Sun, J.; Qin, X.; Gan, P.; Zhou, Z.; Qian, Y.; Zhao, R.; Zhao, Z.; Cai, W.; et al. Pan-Transcriptomic Analysis Reveals Alternative Splicing Control of Cold Tolerance in Rice. Plant Cell 2024, 36, 2117–2139. [Google Scholar] [CrossRef] [PubMed]
- Mei, W.; Chen, W.; Wang, Y.; Liu, Z.; Dong, Y.; Zhang, G.; Deng, H.; Liu, X.; Lu, X.; Wang, F.; et al. Exogenous Kinetin Modulates ROS Homeostasis to Affect Heat Tolerance in Rice Seedlings. Int. J. Mol. Sci. 2023, 24, 6252. [Google Scholar] [CrossRef] [PubMed]
- Lichtenthaler, H.K. Chlorophylls and Carotenoids: Pigments of Photosynthetic Biomembranes. Methods Enzymol. 1987, 148, 350–382. [Google Scholar] [CrossRef]
- Wang, W.; Mauleon, R.; Hu, Z.; Chebotarov, D.; Tai, S.; Wu, Z.; Li, M.; Zheng, T.; Fuentes, R.R.; Zhang, F.; et al. Genomic Variation in 3,010 Diverse Accessions of Asian Cultivated Rice. Nature 2018, 557, 43–49. [Google Scholar] [CrossRef] [PubMed]
- Leung, C.C.; Tarté, D.A.; Oliver, L.S.; Wang, Q.; Gendron, J.M. Systematic Characterization of Photoperiodic Gene Expression Patterns Reveals Diverse Seasonal Transcriptional Systems in Arabidopsis. PLoS Biol. 2023, 21, e3002283. [Google Scholar] [CrossRef] [PubMed]
- Hashida, Y.; Tezuka, A.; Nomura, Y.; Kamitani, M.; Kashima, M.; Kurita, Y.; Nagano, A.J. Fillable and Unfillable Gaps in Plant Transcriptome under Field and Controlled Environments. Plant Cell Environ. 2022, 45, 2410–2427. [Google Scholar] [CrossRef] [PubMed]
- Peng, H.; Wang, K.; Chen, Z.; Cao, Y.; Gao, Q.; Li, Y.; Li, X.; Lu, H.; Du, H.; Lu, M.; et al. MBKbase for Rice: An Integrated Omics Knowledgebase for Molecular Breeding in Rice. Nucleic Acids Res. 2019, 48, D1085–D1092. [Google Scholar] [CrossRef] [PubMed]
- Prasad, A.; Sharma, S.; Prasad, M. Suppressing Plant Defense: Scavenge the ROS! Physiol. Plant. 2023, 175, e13942. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Wang, D.-D.; Fang, X.; Chen, X.-Y.; Mao, Y.-B. Plant Specialized Metabolism Regulated by Jasmonate Signaling. Plant Cell Physiol. 2019, 60, 2638–2647. [Google Scholar] [CrossRef] [PubMed]
- Jiang, L.; Zhang, X.; Zhao, Y.; Zhu, H.; Fu, Q.; Lu, X.; Huang, W.; Yang, X.; Zhou, X.; Wu, L.; et al. Phytoalexin Sakuranetin Attenuates Endocytosis and Enhances Resistance to Rice Blast. Nat. Commun. 2024, 15, 3437. [Google Scholar] [CrossRef] [PubMed]
- Kunova, A.; Pinna, C.; Ghosh, S.; Dozio, D.; Pizzatti, C.; Princiotto, S.; Cortesi, P.; Dallavalle, S.; Pinto, A. Stilbenoids as Antifungals to Counteract Rice Blast Pathogen Pyricularia Oryzae. ACS Agric. Sci. Technol. 2023, 4, 43–50. [Google Scholar] [CrossRef]
- Huang, F.; Jiang, Y.; Chen, T.; Li, H.; Fu, M.; Wang, Y.; Xu, Y.; Li, Y.; Zhou, Z.; Jia, L.; et al. New Data and New Features of the FunRiceGenes (Functionally Characterized Rice Genes) Database: 2021 Update. Rice 2022, 15, 23. [Google Scholar] [CrossRef] [PubMed]
- Mao, B.; Liu, X.; Hu, D.; Li, D. Co-Expression of RCH10 and AGLU1 Confers Rice Resistance to Fungal Sheath Blight Rhizoctonia Solani and Blast Magnorpathe Oryzae and Reveals Impact on Seed Germination. World J. Microbiol. Biotechnol. 2013, 30, 1229–1238. [Google Scholar] [CrossRef] [PubMed]
- Shah, J.M.; Singh, R.; Veluthambi, K. Transgenic Rice Lines Constitutively Co-Expressing tlp-D34 and Chi11 Display Enhancement of Sheath Blight Resistance. Biol. Plant. 2013, 57, 351–358. [Google Scholar] [CrossRef]
- Zvereva, A.S.; Klingenbrunner, M.; Teige, M. Calcium Signaling: An Emerging Player in Plant Antiviral Defense. J. Exp. Bot. 2024, 75, 1265–1273. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Gong, P.; Lu, R.; Lozano-Durán, R.; Zhou, X.; Li, F. Chloroplast Immunity: A Cornerstone of Plant Defense. Mol. Plant 2024, 17, 686–688. [Google Scholar] [CrossRef] [PubMed]
- Jung, S.; Woo, J.; Park, E. Talk to Your Neighbors in an Emergency: Stromule-Mediated Chloroplast-Nucleus Communication in Plant Immunity. Curr. Opin. Plant. Biol. 2024, 79, 102529. [Google Scholar] [CrossRef] [PubMed]
- Rui, L.; Yang, S.-Q.; Zhou, X.-H.; Wang, W. The Important Role of Chloroplasts in Plant Immunity. Plant. Commun. 2025, 101420. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Herva, J.J.; González-Melendi, P.; Cuartas-Lanza, R.; Antúnez-Lamas, M.; Río-Alvarez, I.; Li, Z.; López-Torrejón, G.; Díaz, I.; Del Pozo, J.C.; Chakravarthy, S.; et al. A Bacterial Cysteine Protease Effector Protein Interferes with Photosynthesis to Suppress Plant Innate Immune Responses. Cell Microbiol. 2012, 14, 669–681. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Guo, Z.; Hu, J.; Chen, Y.; Chen, F.; Chen, W.; Wang, W.; Ye, B.; Yang, Z.; Li, G.; et al. The Multifunctional Ascorbate Peroxidase Moapx1 Secreted by Magnaporthe oryzae Mediates the Suppression of Rice Immunity. Plant Cell 2025, 37, koaf146. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, S.; Kanwar, P.; Jha, G. Alterations in Rice Chloroplast Integrity, Photosynthesis and Metabolome Associated with Pathogenesis of Rhizoctonia Solani. Sci. Rep. 2017, 7, 41610. [Google Scholar] [CrossRef] [PubMed]
- Cao, B.; Wang, J.; Ma, J.; Hai, Y.; Wang, X.; Fu, Z.; Xiang, Z.; Wang, Y.; Zhang, L.; Wang, J.; et al. Large-Scale Screening and Function Analysis of Rhizoctonia solani Effectors Targeting Rice Chloroplasts. J. Agric. Food Chem. 2024, 72, 24336–24346. [Google Scholar] [CrossRef] [PubMed]
- Qi, Y.; Armbruster, U.; Schmitz-Linneweber, C.; Delannoy, E.; de Longevialle, A.F.; Rühle, T.; Small, I.; Jahns, P.; Leister, D. Arabidopsis CSP41 Proteins Form Multimeric Complexes That Bind and Stabilize Distinct Plastid Transcripts. J. Exp. Bot. 2011, 63, 1251–1270. [Google Scholar] [CrossRef] [PubMed]
- Bollenbach, T.J.; Sharwood, R.E.; Gutierrez, R.; Lerbs-Mache, S.; Stern, D.B. The RNA-Binding Proteins CSP41a and CSP41b May Regulate Transcription and Translation of Chloroplast-Encoded RNAs in Arabidopsis. Plant Mol. Biol. 2008, 69, 541–552. [Google Scholar] [CrossRef] [PubMed]
- Beligni, M.V.; Mayfield, S.P. Arabidopsis Thaliana Mutants Reveal a Role for CSP41a and CSP41b, Two Ribosome-Associated Endonucleases, in Chloroplast Ribosomal RNA Metabolism. Plant Mol. Biol. 2008, 67, 389–401. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Zhu, P.; Yu, X.; Xu, J.; Liu, G. Physiological and Molecular Mechanisms of Rice Tolerance to Salt and Drought Stress: Advances and Future Directions. Int. J. Mol. Sci. 2024, 25, 9404. [Google Scholar] [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, J.; Zhang, Y.; Liu, T.; Wang, G.; Ju, R.; Sun, Q.; Chen, Q.; Xiong, Y.; Zhai, P.; Xie, W.; et al. Overexpression of OsCSP41b Enhances Rice Tolerance to Sheath Blight Caused by Rhizoctonia solani. J. Fungi 2025, 11, 548. https://doi.org/10.3390/jof11080548
Zhao J, Zhang Y, Liu T, Wang G, Ju R, Sun Q, Chen Q, Xiong Y, Zhai P, Xie W, et al. Overexpression of OsCSP41b Enhances Rice Tolerance to Sheath Blight Caused by Rhizoctonia solani. Journal of Fungi. 2025; 11(8):548. https://doi.org/10.3390/jof11080548
Chicago/Turabian StyleZhao, Jianhua, Yan Zhang, Taixuan Liu, Guangda Wang, Ran Ju, Quanyi Sun, Qi Chen, Yixuan Xiong, Penfei Zhai, Wenya Xie, and et al. 2025. "Overexpression of OsCSP41b Enhances Rice Tolerance to Sheath Blight Caused by Rhizoctonia solani" Journal of Fungi 11, no. 8: 548. https://doi.org/10.3390/jof11080548
APA StyleZhao, J., Zhang, Y., Liu, T., Wang, G., Ju, R., Sun, Q., Chen, Q., Xiong, Y., Zhai, P., Xie, W., Feng, Z., Chen, Z., Hu, K., & Zuo, S. (2025). Overexpression of OsCSP41b Enhances Rice Tolerance to Sheath Blight Caused by Rhizoctonia solani. Journal of Fungi, 11(8), 548. https://doi.org/10.3390/jof11080548