

Recent Advances in Heterologous Protein Expression and Natural Product Synthesis by Aspergillus


Abstract

1. Introduction
2. Aspergillus Is an Expression Host for Heterologous Protein Production
2.1. A. niger
2.2. A. oryzae
2.3. A. nidulans
3. Aspergillus Is a Microbial Chassis for Natural Product Synthesis
3.1. A. oryzae Is a Chassis for Natural Product Production
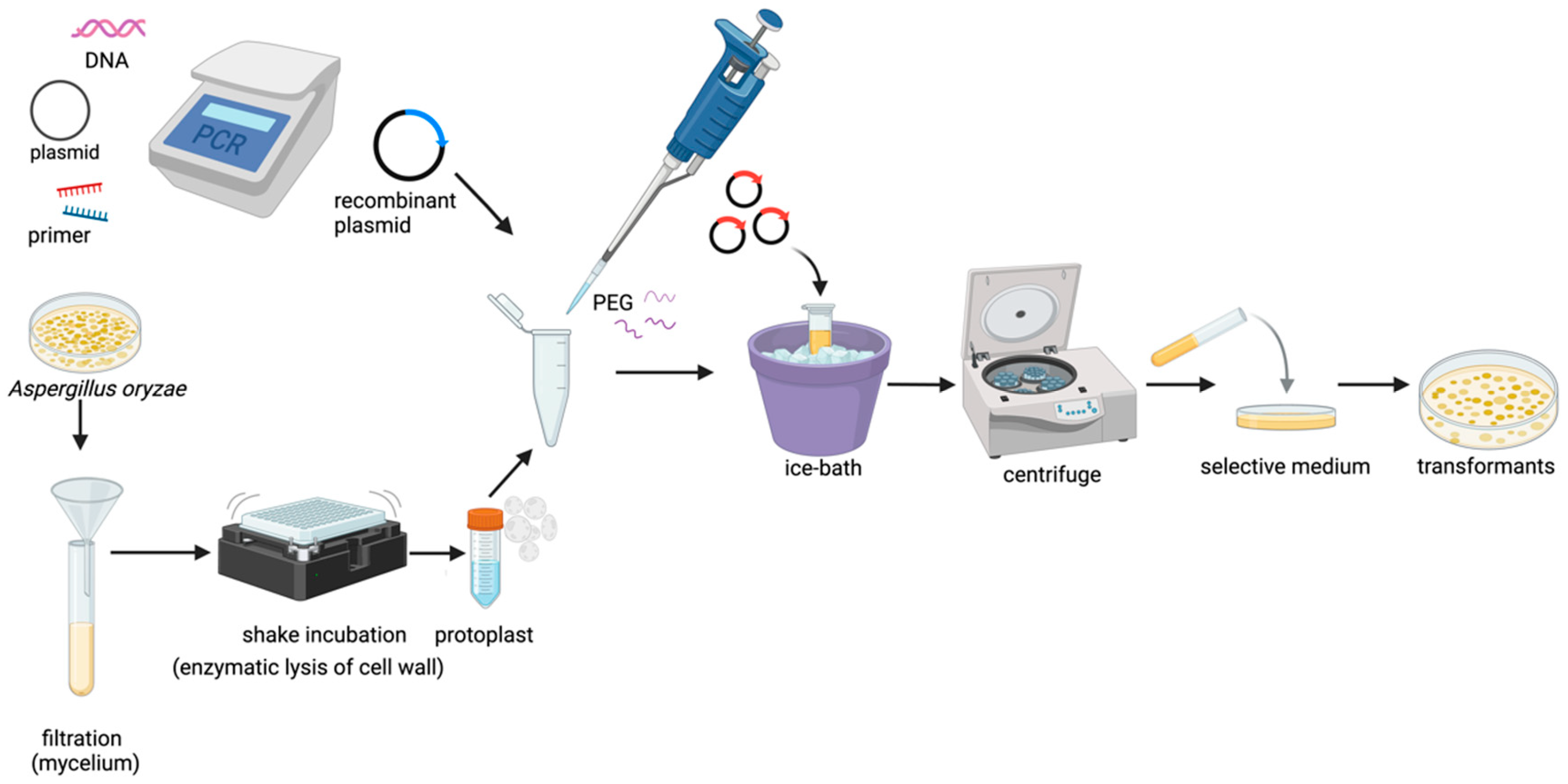
3.1.1. Transformation Methods in A. oryzae
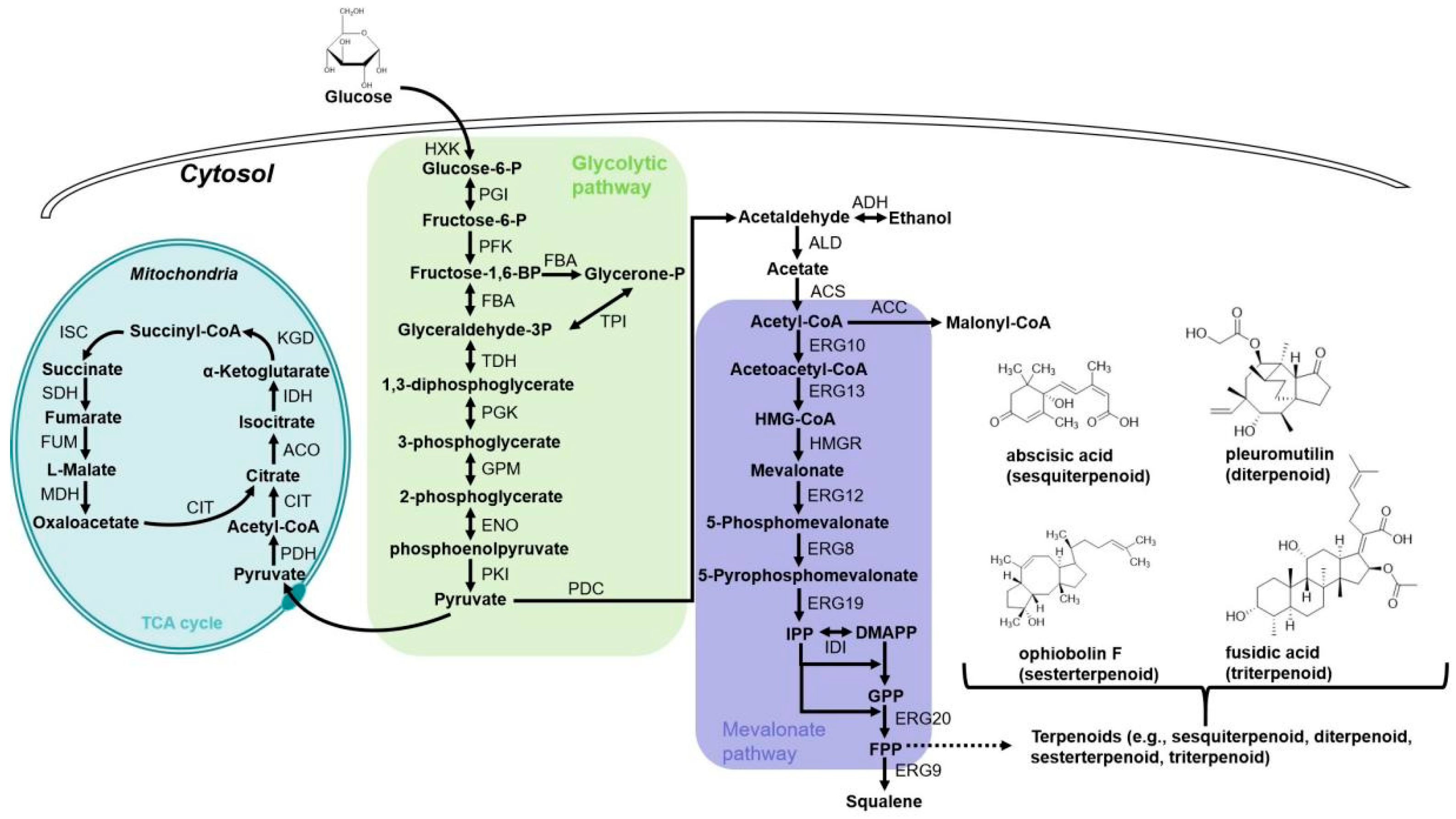
3.1.2. The Production of Terpenoids in A. oryzae
Sesquiterpenoids
Diterpenoids
Sesterterpenoids
Triterpenoids and Steroids

{kind=link}
{kind=link}
{kind=link}
| Natural Products | Structure Type | Gene Source | Heterologous Hosts | References |
|---|---|---|---|---|
| abscisic acid | sesquiterpenoid | Botrytis cinerea SAS56 | A. oryzae NSAR1 | [83] |
| trichobrasilenol | sesquiterpenoid | T. atroviride FKI-3849 | A. oryzae NSAR1 | [84] |
| brasilane A, D and E | sesquiterpenoid | Annulohypoxylon truncatum CBS 140778 | A. oryzae NSAR1 | [85] |
| 1α-hydroxymelleolide | sesquiterpenoid | Armillaria mellea FMC 543 | A. oryzae NSPlD1 | [86] |
| pleuromutilin | diterpenoid | Clitopilus passeckerianus ATCC 34646 | A. oryzae NSAR1 | [21] |
| aphidicolin | diterpenoid | Phoma betae PS-13 | A. oryzae NSAR1 | [90] |
| brassicicene I | diterpenoid | Pseudocercospora fijiensis 10CR-1-24 | A. oryzae NSAR1 | [97] |
| erinacine Q | diterpenoid | Hericiumerinaceusyamabushitake Y2 | A. oryzae NSPlD1 | [67] |
| 20-prenylpaxilline 22-prenylpaxilline | diterpenoid | Tolypocladium inflatum | A. oryzae NSAR1 | [114] |
| cotylenin C, F, I and E | diterpenoid | Talaromyces adpressus | A. oryzae NSAR1 | [115] |
| cycloaraneosene-9-ol-8-one | diterpenoid | Talaromyces adpressus | A. oryzae NSAR1 | [98] |
| ophiobolin F | sesterterpenoid | Aspergillus clavatus NRRL | A. oryzae NSAR1 | [102] |
| astellifadiene | sesterterpenoid | Emericella variecolor NBRC32302 | A. oryzae NSAR1 | [116] |
| quiannulatene quiannulatic acid | sesterterpenoid | Emericella variecolor NBRC32302 | A. oryzae NSAR1 | [99] |
| asperterpenoid A | sesterterpenoid | Talaromyces wortmannii ATCC 26942 | A. oryzae NSAR | [117] |
| asperterpenol A asperterpenol B | sesterterpenoid | Aspergillus calidoustus CBS121601 | A. oryzae NSAR | [104] |
| mangicol J | sesterterpenoid | Fusarium graminearum J1-012 | A. oryzae NSAR1 | [80] |
| variecolin | sesterterpenoid | Aspergillus aculeatus ATCC 16872 | A. oryzae NSAR1 | [106] |
| helvolic acid | triterpenoid | Aspergillus fumigatus Af293 | A. oryzae NSAR1 | [110] |
| fusidic acid | triterpenoid | Acremonium fusidioides ATCC 14700 | A. oryzae NSAR1 | [111] |
| cephalosporin P1 | triterpenoid | Acremonium chrysogenum ATCC 11550 | A. oryzae NSAR1 | [22] |
| polytolypin | triterpenoid | Polytolypa hystricis UAMH7299 | A. oryzae NSAR1 | [112] |
| fuscoatroside | triterpenoid | Humicola fuscoatra NRRL 22980 | A. oryzae NSAR1 | [113] |
3.1.3. A. oryzae as a Chassis for Synthesis of Other Natural Products
Polyketides
Non-Ribosomal Peptides
3.2. A. nidulans as a Chassis for Natural Product Synthesis
3.3. A. niger Is a Chassis for Natural Product Synthesis
4. Application of CRISPR/Cas9-Based Genome Editing Technology in A. oryzae and A. niger
4.1. CRISPR/Cas9-Based Genome Editing Technology for A. niger
4.2. CRISPR/Cas9-Based Genome Editing Technology for A. oryzae
5. Conclusions and Perspectives
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Pan, R.; Yang, X.; Qiu, M.; Jiang, W.; Zhang, W.; Jiang, Y.; Xin, F.; Jiang, M. Construction of Coculture System Containing Escherichia coli with Different Microbial Species for Biochemical Production. ACS Synth. Biol. 2023, 12, 2208–2216. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Wu, X.; Yao, M.; Xiao, W.; Zha, J. Chassis engineering for microbial production of chemicals: From natural microbes to synthetic organisms. Curr. Opin. Biotechnol. 2020, 66, 105–112. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Liu, Y.; Du, G.; Ledesma-Amaro, R.; Liu, L. Microbial Chassis Development for Natural Product Biosynthesis. Trends Biotechnol. 2020, 38, 779–796. [Google Scholar] [CrossRef]
- Knuf, C.; Nielsen, J. Aspergilli: Systems biology and industrial applications. Biotechnol. J. 2012, 7, 1147–1155. [Google Scholar] [CrossRef] [PubMed]
- Sun, Z.; Wu, Y.; Long, S.; Feng, S.; Jia, X.; Hu, Y.; Ma, M.; Liu, J.; Zeng, B. Aspergillus oryzae as a Cell Factory: Research and Applications in Industrial Production. J. Fungi 2024, 10, 248. [Google Scholar] [CrossRef]
- Alberti, F.; Foster, G.D.; Bailey, A.M. Natural products from filamentous fungi and production by heterologous expression. Appl. Microbiol. Biotechnol. 2017, 101, 493–500. [Google Scholar] [CrossRef]
- Van Den Berg, M.; Gidijala, L.; Kiela, J.; Bovenberg, R.; Vander Keli, I. Biosynthesis of active pharmaceuticals: β-lactam biosynthesis in filamentous fungi. Biotechnol. Genet. Eng. Rev. 2010, 27, 1–32. [Google Scholar] [CrossRef]
- Ding, Q.; Ye, C. Microbial cell factories based on filamentous bacteria, yeasts, and fungi. Microb. Cell Fact. 2023, 22, 20. [Google Scholar] [CrossRef]
- Felipe, M.T.D.; Barbosa, R.D.; Bezerra, J.D.P.; de Souza-Motta, C.M. Production of kojic acid by Aspergillus species: Trends and applications. Fungal Biol. Rev. 2023, 45, 100313. [Google Scholar] [CrossRef]
- Chiang, Y.M.; Lin, T.S.; Wang, C.C.C. Total Heterologous Biosynthesis of Fungal Natural Products in Aspergillus nidulans. J. Nat. Prod. 2022, 85, 2484–2518. [Google Scholar] [CrossRef]
- Guo, C.J.; Wang, C.C. Recent advances in genome mining of secondary metabolites in Aspergillus terreus. Front. Microbiol. 2014, 5, 717. [Google Scholar] [CrossRef] [PubMed]
- Meng, X.; Fang, Y.; Ding, M.; Zhang, Y.; Jia, K.; Li, Z.; Collemare, J.; Liu, W. Developing fungal heterologous expression platforms to explore and improve the production of natural products from fungal biodiversity. Biotechnol. Adv. 2022, 54, 107866. [Google Scholar] [CrossRef] [PubMed]
- Cox, R.J. Engineered and total biosynthesis of fungal specialized metabolites. Nat. Rev. Chem. 2024, 8, 61–78. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Men, P.; Tang, S.; Lu, X. Aspergillus terreus as an industrial filamentous fungus for pharmaceutical biotechnology. Curr. Opin. Biotechnol. 2021, 69, 273–280. [Google Scholar] [CrossRef]
- West, T.P. Citric Acid Production by Aspergillus niger Using Solid-State Fermentation of Agricultural Processing Coproducts. Appl. Biosci. 2023, 2, 1–13. [Google Scholar] [CrossRef]
- Daba, G.M.; Mostafa, F.A.; Elkhateeb, W.A. The ancient koji mold (Aspergillus oryzae) as a modern biotechnological tool. Bioresour. Bioprocess. 2021, 8, 52. [Google Scholar] [CrossRef]
- Punt, P.J.; van Biezen, N.; Conesa, A.; Albers, A.; Mangnus, J.; van den Hondel, C. Filamentous fungi as cell factories for heterologous protein production. Trends Biotechnol. 2002, 20, 200–206. [Google Scholar] [CrossRef]
- Huynh, H.H.; Morita, N.; Sakamoto, T.; Katayama, T.; Miyakawa, T.; Tanokura, M.; Chiba, Y.; Shinkura, R.; Maruyama, J.I. Functional production of human antibody by the filamentous fungus Aspergillus oryzae. Fungal Biol. Biotechnol. 2020, 7, 7. [Google Scholar] [CrossRef]
- Fan, J.; Wei, P.L.; Li, Y.; Zhang, S.; Ren, Z.; Li, W.; Yin, W.B. Developing filamentous fungal chassis for natural product production. Bioresour. Technol. 2025, 415, 131703. [Google Scholar] [CrossRef]
- Ntana, F.; Mortensen, U.H.; Sarazin, C.; Figge, R. Aspergillus: A Powerful Protein Production Platform. Catalysts 2020, 10, 1064. [Google Scholar] [CrossRef]
- Bailey, A.M.; Alberti, F.; Kilaru, S.; Collins, C.M.; de Mattos-Shipley, K.; Hartley, A.J.; Hayes, P.; Griffin, A.; Lazarus, C.M.; Cox, R.J.; et al. Identification and manipulation of the pleuromutilin gene cluster from Clitopilus passeckerianus for increased rapid antibiotic production. Sci. Rep. 2016, 6, 25202. [Google Scholar] [CrossRef]
- Cao, Z.Q.; Lv, J.M.; Liu, Q.; Qin, S.Y.; Chen, G.D.; Dai, P.; Zhong, Y.; Gao, H.; Yao, X.S.; Hu, D. Biosynthetic Study of Cephalosporin P(1) Reveals a Multifunctional P450 Enzyme and a Site-Selective Acetyltransferase. ACS Chem. Biol. 2020, 15, 44–51. [Google Scholar] [CrossRef] [PubMed]
- Cairns, T.C.; Nai, C.; Meyer, V. How a fungus shapes biotechnology: 100 years of Aspergillus niger research. Fungal Biol. Biotechnol. 2018, 5, 13. [Google Scholar] [CrossRef] [PubMed]
- Lübeck, M.; Lübeck, P.S. Fungal Cell Factories for Efficient and Sustainable Production of Proteins and Peptides. Microorganisms 2022, 10, 753. [Google Scholar] [CrossRef]
- Gruben, B.S.; Mäkelä, M.R.; Kowalczyk, J.E.; Zhou, M.; Benoit-Gelber, I.; De Vries, R.P. Expression-based clustering of CAZyme-encoding genes of Aspergillus niger. BMC Genom. 2017, 18, 900. [Google Scholar] [CrossRef]
- Fiedler, M.R.M.; Barthel, L.; Kubisch, C.; Nai, C.; Meyer, V. Construction of an improved Aspergillus niger platform for enhanced glucoamylase secretion. Microb. Cell Fact. 2018, 17, 95. [Google Scholar] [CrossRef]
- Zhang, H.; Yan, J.N.; Zhang, H.; Liu, T.Q.; Xu, Y.; Zhang, Y.Y.; Li, J. Effect of gpd box copy numbers in the gpdA promoter of Aspergillus nidulans on its transcription efficiency in Aspergillus niger. FEMS Microbiol. Lett. 2018, 365, fny154. [Google Scholar] [CrossRef]
- Cai, L.N.; Xu, S.N.; Lu, T.; Lin, D.Q.; Yao, S.J. Directed expression of halophilic and acidophilic β-glucosidases by introducing homologous constitutive expression cassettes in marine Aspergillus niger. J. Biotechnol. 2019, 292, 12–22. [Google Scholar] [CrossRef]
- Alazi, E.; Knetsch, T.; Di Falco, M.; Reid, I.D.; Arentshorst, M.; Visser, J.; Tsang, A.; Ram, A.F.J. Inducer-independent production of pectinases in Aspergillus niger by overexpression of the D-galacturonic acid-responsive transcription factor gaaR. Appl. Microbiol. Biotechnol. 2018, 102, 2723–2736. [Google Scholar] [CrossRef]
- Liu, F.; Wang, B.; Ye, Y.; Pan, L. High level expression and characterization of tannase tan7 using Aspergillus niger SH-2 with low-background endogenous secretory proteins as the host. Protein Expr. Purif. 2018, 144, 71–75. [Google Scholar] [CrossRef]
- Zhu, S.Y.; Xu, Y.; Yu, X.W. Improved Homologous Expression of the Acidic Lipase from Aspergillus niger. J. Microbiol. Biotechnol. 2020, 30, 196–205. [Google Scholar] [CrossRef]
- Li, C.; Zhou, J.; Du, G.; Chen, J.; Takahashi, S.; Liu, S. Developing Aspergillus niger as a cell factory for food enzyme production. Biotechnol. Adv. 2020, 44, 107630. [Google Scholar] [CrossRef]
- Archer, D.B.; Jeenes, D.J.; MacKenzie, D.A.; Brightwell, G.; Lambert, N.; Lowe, G.; Radford, S.E.; Dobson, C.M. Hen egg white lysozyme expressed in, and secreted from, Aspergillus niger is correctly processed and folded. Biotechnology 1990, 8, 741–745. [Google Scholar] [CrossRef]
- Ward, P.P.; Piddington, C.S.; Cunningham, G.A.; Zhou, X.; Wyatt, R.D.; Conneely, O.M. A system for production of commercial quantities of human lactoferrin: A broad spectrum natural antibiotic. Biotechnology 1995, 13, 498–503. [Google Scholar] [CrossRef] [PubMed]
- Dunn-Coleman, N.S.; Bloebaum, P.; Berka, R.M.; Bodie, E.; Robinson, N.; Armstrong, G.; Ward, M.; Przetak, M.; Carter, G.L.; LaCost, R.; et al. Commercial levels of chymosin production by Aspergillus. Biotechnology 1991, 9, 976–981. [Google Scholar] [CrossRef] [PubMed]
- Faus, I.; del Moral, C.; Adroer, N.; del Río, J.L.; Patiño, C.; Sisniega, H.; Casas, C.; Bladé, J.; Rubio, V. Secretion of the sweet-tasting protein thaumatin by recombinant strains of Aspergillus niger var. awamori. Appl. Microbiol. Biotechnol. 1998, 49, 393–398. [Google Scholar] [CrossRef] [PubMed]
- Prathumpai, W.; Flitter, S.J.; McIntyre, M.; Nielsen, J. Lipase production by recombinant strains of Aspergillus niger expressing a lipase-encoding gene from Thermomyces lanuginosus. Appl. Microbiol. Biotechnol. 2004, 65, 714–719. [Google Scholar] [CrossRef]
- Chen, X.; Wang, B.; Pan, L. Heterologous expression and characterization of Penicillium citrinum nuclease P1 in Aspergillus niger and its application in the production of nucleotides. Protein Expr. Purif. 2019, 156, 36–43. [Google Scholar] [CrossRef]
- Kamaruddin, N.; Storms, R.; Mahadi, N.M.; Illias, R.M.; Abu Bakar, F.D.; Murad, A.M.A. Reduction of Extracellular Proteases Increased Activity and Stability of Heterologous Protein in Aspergillus niger. Arab. J. Sci. Eng. 2018, 43, 3327–3338. [Google Scholar] [CrossRef]
- Jin, F.J.; Wang, B.T.; Wang, Z.D.; Jin, L.; Han, P. CRISPR/Cas9-Based Genome Editing and Its Application in Aspergillus Species. J. Fungi 2022, 8, 467. [Google Scholar] [CrossRef]
- Dong, L.; Lin, X.; Yu, D.; Huang, L.; Wang, B.; Pan, L. High-level expression of highly active and thermostable trehalase from Myceliophthora thermophila in Aspergillus niger by using the CRISPR/Cas9 tool and its application in ethanol fermentation. J. Ind. Microbiol. Biotechnol. 2020, 47, 133–144. [Google Scholar] [CrossRef]
- Wang, S.; Xue, Y.; Zhang, P.; Yan, Q.; Li, Y.; Jiang, Z. CRISPR/Cas9 System-Mediated Multi-copy Expression of an Alkaline Serine Protease in Aspergillus niger for the Production of XOD-Inhibitory Peptides. J. Agric. Food Chem. 2023, 71, 15194–15203. [Google Scholar] [CrossRef] [PubMed]
- Jeennor, S.; Anantayanon, J.; Panchanawaporn, S.; Chutrakul, C.; Vongsangnak, W.; Laoteng, K. Efficient de novo production of bioactive cordycepin by Aspergillus oryzae using a food-grade expression platform. Microb. Cell Fact. 2023, 22, 253. [Google Scholar] [CrossRef] [PubMed]
- Goldman, G.H.; Kafer, E. Aspergillus nidulans as a model system to characterize the DNA damage response in eukaryotes. Fungal Genet. Biol. 2004, 41, 428–442. [Google Scholar] [CrossRef] [PubMed]
- Todd, R.B.; Davis, M.A.; Hynes, M.J. Genetic manipulation of Aspergillus nidulans: Meiotic progeny for genetic analysis and strain construction. Nat. Protoc. 2007, 2, 811–821. [Google Scholar] [CrossRef]
- Kumar, A. Aspergillus nidulans: A Potential Resource of the Production of the Native and Heterologous Enzymes for Industrial Applications. Int. J. Microbiol. 2020, 2020, 8894215. [Google Scholar] [CrossRef]
- Liu, E.; Li, M.; Abdella, A.; Wilkins, M.R. Development of a cost-effective medium for submerged production of fungal aryl alcohol oxidase using a genetically modified Aspergillus nidulans strain. Bioresour. Technol. 2020, 305, 123038. [Google Scholar] [CrossRef]
- Lopes, A.M.M.; Martins, M.; Goldbeck, R. Heterologous Expression of Lignocellulose-Modifying Enzymes in Microorganisms: Current Status. Mol. Biotechnol. 2021, 63, 184–199. [Google Scholar] [CrossRef]
- Yan, Q.; Han, L.; Liu, Z.; Zhou, S.; Zhou, Z. Stepwise genetic modification for efficient expression of heterologous proteins in Aspergillus nidulans. Appl. Microbiol. Biotechnol. 2023, 107, 6923–6935. [Google Scholar] [CrossRef]
- Gerhardt, J.A.; Rubio, M.V.; Terrasan, C.R.F.; Wassano, N.S.; Rodrigues, A.; Figueiredo, F.L.; Antoniel, E.P.; Contesini, F.J.; Dias, A.H.S.; Mortensen, U.H.; et al. Improving recombinant protein secretion in Aspergillus nidulans by targeting the N-glycosylation machinery. Metab. Eng. Commun. 2025, 20, e00264. [Google Scholar] [CrossRef]
- Fleissner, A.; Dersch, P. Expression and export: Recombinant protein production systems for Aspergillus. Appl. Microbiol. Biotechnol. 2010, 87, 1255–1270. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.; Garrigues, S.; de Vries, R.P. Heterologous protein production in filamentous fungi. Appl. Microbiol. Biotechnol. 2023, 107, 5019–5033. [Google Scholar] [CrossRef]
- Lubertozzi, D.; Keasling, J.D. Developing Aspergillus as a host for heterologous expression. Biotechnol. Adv. 2009, 27, 53–75. [Google Scholar] [CrossRef] [PubMed]
- Lazarus, C.M.; Williams, K.; Bailey, A.M. Reconstructing fungal natural product biosynthetic pathways. Nat. Prod. Rep. 2014, 31, 1339–1347. [Google Scholar] [CrossRef] [PubMed]
- Qiu, S.; Blank, L.M. Recent Advances in Yeast Recombinant Biosynthesis of the Triterpenoid Protopanaxadiol and Glycosylated Derivatives Thereof. J. Agric. Food Chem. 2023, 71, 2197–2210. [Google Scholar] [CrossRef]
- Chiang, Y.M.; Oakley, C.E.; Ahuja, M.; Entwistle, R.; Schultz, A.; Chang, S.L.; Sung, C.T.; Wang, C.C.; Oakley, B.R. An efficient system for heterologous expression of secondary metabolite genes in Aspergillus nidulans. J. Am. Chem. Soc. 2013, 135, 7720–7731. [Google Scholar] [CrossRef]
- Yaegashi, J.; Oakley, B.R.; Wang, C.C. Recent advances in genome mining of secondary metabolite biosynthetic gene clusters and the development of heterologous expression systems in Aspergillus nidulans. J. Ind. Microbiol. Biotechnol. 2014, 41, 433–442. [Google Scholar] [CrossRef]
- Sakai, K.; Kinoshita, H.; Nihira, T. Heterologous expression system in Aspergillus oryzae for fungal biosynthetic gene clusters of secondary metabolites. Appl. Microbiol. Biotechnol. 2012, 93, 2011–2022. [Google Scholar] [CrossRef]
- Oikawa, H. Reconstitution of biosynthetic machinery of fungal natural products in heterologous hosts. Biosci. Biotechnol. Biochem. 2020, 84, 433–444. [Google Scholar] [CrossRef]
- Oikawa, H. Heterologous production of fungal natural products: Reconstitution of biosynthetic gene clusters in model host Aspergillus oryzae. Proc. Jpn. Acad. Ser. B Phys. Biol. Sci. 2020, 96, 420–430. [Google Scholar] [CrossRef]
- Gallagher, R.R.; Patel, J.R.; Interiano, A.L.; Rovner, A.J.; Isaacs, F.J. Multilayered genetic safeguards limit growth of microorganisms to defined environments. Nucleic Acids Res. 2015, 43, 1945–1954. [Google Scholar] [CrossRef]
- Yang, H.; Song, C.; Liu, C.; Wang, P. Synthetic Biology Tools for Engineering Aspergillus oryzae. J. Fungi 2024, 10, 34. [Google Scholar] [CrossRef] [PubMed]
- Matsuda, Y.; Wakimoto, T.; Mori, T.; Awakawa, T.; Abe, I. Complete biosynthetic pathway of anditomin: Nature’s sophisticated synthetic route to a complex fungal meroterpenoid. J. Am. Chem. Soc. 2014, 136, 15326–15336. [Google Scholar] [CrossRef] [PubMed]
- Wei, X.; Chen, X.; Chen, L.; Yan, D.; Wang, W.G.; Matsuda, Y. Heterologous Biosynthesis of Tetrahydroxanthone Dimers: Determination of Key Factors for Selective or Divergent Synthesis. J. Nat. Prod. 2021, 84, 1544–1549. [Google Scholar] [CrossRef] [PubMed]
- Yamane, M.; Minami, A.; Liu, C.; Ozaki, T.; Takeuchi, I.; Tsukagoshi, T.; Tokiwano, T.; Gomi, K.; Oikawa, H. Biosynthetic Machinery of Diterpene Pleuromutilin Isolated from Basidiomycete Fungi. Chembiochem 2017, 18, 2317–2322. [Google Scholar] [CrossRef]
- Zhang, S.L.; Ban, A.; Ebara, N.; Mizutani, O.; Tanaka, M.; Shintani, T.; Gomi, K. Self-excising Cre/mutant lox marker recycling system for multiple gene integrations and consecutive gene deletions in Aspergillus oryzae. J. Biosci. Bioeng. 2017, 123, 403–411. [Google Scholar] [CrossRef]
- Liu, C.; Minami, A.; Ozaki, T.; Wu, J.; Kawagishi, H.; Maruyama, J.I.; Oikawa, H. Efficient Reconstitution of Basidiomycota Diterpene Erinacine Gene Cluster in Ascomycota Host Aspergillus oryzae Based on Genomic DNA Sequences. J. Am. Chem. Soc. 2019, 141, 15519–15523. [Google Scholar] [CrossRef]
- Wei, X.; Matsuyama, T.; Sato, H.; Yan, D.; Chan, P.M.; Miyamoto, K.; Uchiyama, M.; Matsuda, Y. Molecular and Computational Bases for Spirofuranone Formation in Setosusin Biosynthesis. J. Am. Chem. Soc. 2021, 143, 17708–17715. [Google Scholar] [CrossRef]
- Li, Q.H.; Lu, J.C.; Zhang, G.Q.; Zhou, J.W.; Li, J.H.; Du, G.C.; Chen, J. CRISPR/Cas9-Mediated Multiplexed Genome Editing in Aspergillus oryzae. J. Fungi 2023, 9, 109. [Google Scholar] [CrossRef]
- Shataer, D.; Chang, Y.H.; Obul, M.; Aierken, K.; Liu, H.P. An Up-to-date Review on the Classification, Pharmacology, and Production of Terpenes and Terpenoids. Curr. Org. Chem. 2025, 29, 1508–1522. [Google Scholar] [CrossRef]
- Zhang, Y.; Nielsen, J.; Liu, Z. Engineering yeast metabolism for production of terpenoids for use as perfume ingredients, pharmaceuticals and biofuels. FEMS Yeast Res. 2017, 17, fox080. [Google Scholar] [CrossRef]
- Bergman, M.E.; Davis, B.; Phillips, M.A. Medically Useful Plant Terpenoids: Biosynthesis, Occurrence, and Mechanism of Action. Molecules 2019, 24, 3961. [Google Scholar] [CrossRef]
- Zhao, Y.; Pan, H.; Liu, W.; Liu, E.; Pang, Y.; Gao, H.; He, Q.; Liao, W.; Yao, Y.; Zeng, J.; et al. Menthol: An underestimated anticancer agent. Front. Pharmacol. 2023, 14, 1148790. [Google Scholar] [CrossRef]
- Abu Samaan, T.M.; Samec, M.; Liskova, A.; Kubatka, P.; Büsselberg, D. Paclitaxel’s Mechanistic and Clinical Effects on Breast Cancer. Biomolecules 2019, 9, 789. [Google Scholar] [CrossRef]
- Ma, N.; Zhang, Z.; Liao, F.; Jiang, T.; Tu, Y. The birth of artemisinin. Pharmacol. Ther. 2020, 216, 107658. [Google Scholar] [CrossRef]
- Gao, S.; Liu, Z.; Li, H.; Little, P.J.; Liu, P.; Xu, S. Cardiovascular actions and therapeutic potential of tanshinone IIA. Atherosclerosis 2012, 220, 3–10. [Google Scholar] [CrossRef]
- Liao, P.; Hemmerlin, A.; Bach, T.J.; Chye, M.L. The potential of the mevalonate pathway for enhanced isoprenoid production. Biotechnol. Adv. 2016, 34, 697–713. [Google Scholar] [CrossRef] [PubMed]
- Zhao, W.Y.; Yi, J.; Chang, Y.B.; Sun, C.P.; Ma, X.C. Recent studies on terpenoids in Aspergillus fungi: Chemical diversity, biosynthesis, and bioactivity. Phytochemistry 2022, 193, 113011. [Google Scholar] [CrossRef] [PubMed]
- Nagamine, S.; Liu, C.; Nishishita, J.; Kozaki, T.; Sogahata, K.; Sato, Y.; Minami, A.; Ozaki, T.; Schmidt-Dannert, C.; Maruyama, J.I.; et al. Ascomycete Aspergillus oryzae Is an Efficient Expression Host for Production of Basidiomycete Terpenes by Using Genomic DNA Sequences. Appl. Environ. Microbiol. 2019, 85, e00409-19. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Y.J.; Cheng, S.; Bian, G.K.; Yan, P.; Ma, Z.N.; Dai, W.; Chen, R.; Fu, S.; Huang, H.W.; Chi, H.M.; et al. Efficient exploration of terpenoid biosynthetic gene clusters in filamentous fungi. Nat. Catal. 2022, 5, 277. [Google Scholar] [CrossRef]
- Dai, Q.; Zhang, F.L.; Feng, T. Sesquiterpenoids Specially Produced by Fungi: Structures, Biological Activities, Chemical and Biosynthesis (2015–2020). J. Fungi 2021, 7, 1026. [Google Scholar] [CrossRef]
- Chen, D.L.; Wang, B.W.; Sun, Z.C.; Yang, J.S.; Xu, X.D.; Ma, G.X. Natural Nitrogenous Sesquiterpenoids and Their Bioactivity: A Review. Molecules 2020, 25, 2485. [Google Scholar] [CrossRef]
- Takino, J.; Kozaki, T.; Sato, Y.; Liu, C.; Ozaki, T.; Minami, A.; Oikawa, H. Unveiling Biosynthesis of the Phytohormone Abscisic Acid in Fungi: Unprecedented Mechanism of Core Scaffold Formation Catalyzed by an Unusual Sesquiterpene Synthase. J. Am. Chem. Soc. 2018, 140, 12392–12395. [Google Scholar] [CrossRef]
- Murai, K.; Lauterbach, L.; Teramoto, K.; Quan, Z.; Barra, L.; Yamamoto, T.; Nonaka, K.; Shiomi, K.; Nishiyama, M.; Kuzuyama, T.; et al. An Unusual Skeletal Rearrangement in the Biosynthesis of the Sesquiterpene Trichobrasilenol from Trichoderma. Angew. Chem. Int. Ed. Engl. 2019, 58, 15046–15050. [Google Scholar] [CrossRef] [PubMed]
- Feng, J.; Surup, F.; Hauser, M.; Miller, A.; Wennrich, J.P.; Stadler, M.; Cox, R.J.; Kuhnert, E. Biosynthesis of oxygenated brasilane terpene glycosides involves a promiscuous N-acetylglucosamine transferase. Chem. Commun. 2020, 56, 12419–12422. [Google Scholar] [CrossRef] [PubMed]
- Fukaya, M.; Nagamine, S.; Ozaki, T.; Liu, Y.; Ozeki, M.; Matsuyama, T.; Miyamoto, K.; Kawagishi, H.; Uchiyama, M.; Oikawa, H.; et al. Total Biosynthesis of Melleolides from Basidiomycota Fungi: Mechanistic Analysis of the Multifunctional GMC Oxidase Mld7. Angew. Chem. Int. Ed. Engl. 2023, 62, e202308881. [Google Scholar] [CrossRef] [PubMed]
- Han, H.; Zhang, P.; Xie, Z.; Qi, J.; Wang, P.; Li, C.; Xue, Z.; Wu, R.; Liu, C. Functional Characterization of Sesquiterpene Synthases and P450 Enzymes in Flammulina velutipes for Biosynthesis of Spiro [4.5] Decane Terpene. J. Agric. Food Chem. 2024, 72, 9227–9235. [Google Scholar] [CrossRef]
- Rico-Martínez, M.; Medina, F.G.; Marrero, J.G.; Osegueda-Robles, S. Biotransformation of diterpenes. Rsc Adv. 2014, 4, 10627–10647. [Google Scholar] [CrossRef]
- Hu, Z.M.; Liu, X.Y.; Tian, M.; Ma, Y.; Jin, B.L.; Gao, W.; Cui, G.H.; Guo, J.; Huang, L.Q. Recent progress and new perspectives for diterpenoid biosynthesis in medicinal plants. Med. Res. Rev. 2021, 41, 2971–2997. [Google Scholar] [CrossRef]
- Fujii, R.; Minami, A.; Tsukagoshi, T.; Sato, N.; Sahara, T.; Ohgiya, S.; Gomi, K.; Oikawa, H. Total biosynthesis of diterpene aphidicolin, a specific inhibitor of DNA polymerase α: Heterologous expression of four biosynthetic genes in Aspergillus oryzae. Biosci. Biotechnol. Biochem. 2011, 75, 1813–1817. [Google Scholar] [CrossRef]
- Qin, B.; Matsuda, Y.; Mori, T.; Okada, M.; Quan, Z.; Mitsuhashi, T.; Wakimoto, T.; Abe, I. An Unusual Chimeric Diterpene Synthase from Emericella variecolor and Its Functional Conversion into a Sesterterpene Synthase by Domain Swapping. Angew. Chem. Int. Ed. Engl. 2016, 55, 1658–1661. [Google Scholar] [CrossRef]
- Alberti, F.; Khairudin, K.; Venegas, E.R.; Davies, J.A.; Hayes, P.M.; Willis, C.L.; Bailey, A.M.; Foster, G.D. Heterologous expression reveals the biosynthesis of the antibiotic pleuromutilin and generates bioactive semi-synthetic derivatives. Nat. Commun. 2017, 8, 1831. [Google Scholar] [CrossRef]
- Alberti, F.; Khairudin, K.; Davies, J.A.; Sangmalee, S.; Willis, C.L.; Foster, G.D.; Bailey, A.M. Biosynthesis of pleuromutilin congeners using an Aspergillus oryzae expression platform. Chem. Sci. 2023, 14, 3826–3833. [Google Scholar] [CrossRef] [PubMed]
- Schafhauser, T.; Wibberg, D.; Binder, A.; Rückert, C.; Busche, T.; Wohlleben, W.; Kalinowski, J. Genome Assembly and Genetic Traits of the Pleuromutilin-Producer Clitopilus passeckerianus DSM1602. J. Fungi 2022, 8, 862. [Google Scholar] [CrossRef] [PubMed]
- Guo, C.; Dai, H.; Zhang, M.; Liao, H.; Zhang, R.; Chen, B.; Han, J.; Liu, H. Molecular networking assisted discovery and combinatorial biosynthesis of new antimicrobial pleuromutilins. Eur. J. Med. Chem. 2022, 243, 114713. [Google Scholar] [CrossRef]
- Saito, N.; Katayama, T.; Minami, A.; Oikawa, H.; Maruyama, J.I. Versatile filamentous fungal host highly-producing heterologous natural products developed by genome editing-mediated engineering of multiple metabolic pathways. Commun. Biol. 2024, 7, 1263. [Google Scholar] [CrossRef] [PubMed]
- Tazawa, A.; Ye, Y.; Ozaki, T.; Liu, C.; Ogasawara, Y.; Dairi, T.; Higuchi, Y.; Kato, N.; Gomi, K.; Minami, A.; et al. Total Biosynthesis of Brassicicenes: Identification of a Key Enzyme for Skeletal Diversification. Org. Lett. 2018, 20, 6178–6182. [Google Scholar] [CrossRef]
- Xu, Q.; Ren, X.; Hu, L.; Xu, Q.; Zhang, X.; Deng, M.; Ye, Y.; Zhang, Y.; Lu, Y.; Qiao, Y. Uncovering a novel biosynthetic gene cluster for sordarin through genome mining in the fungus Talaromyces adpressus. Bioresour. Bioprocess. 2025, 12, 35. [Google Scholar] [CrossRef]
- Okada, M.; Matsuda, Y.; Mitsuhashi, T.; Hoshino, S.; Mori, T.; Nakagawa, K.; Quan, Z.; Qin, B.; Zhang, H.; Hayashi, F.; et al. Genome-Based Discovery of an Unprecedented Cyclization Mode in Fungal Sesterterpenoid Biosynthesis. J. Am. Chem. Soc. 2016, 138, 10011–10018. [Google Scholar] [CrossRef]
- Li, K.; Gustafson, K.R. Sesterterpenoids: Chemistry, biology, and biosynthesis. Nat. Prod. Rep. 2021, 38, 1251–1281. [Google Scholar] [CrossRef]
- Guo, K.; Liu, Y.; Li, S.H. The untapped potential of plant sesterterpenoids: Chemistry, biological activities and biosynthesis. Nat. Prod. Rep. 2021, 38, 2293–2314. [Google Scholar] [CrossRef] [PubMed]
- Chiba, R.; Minami, A.; Gomi, K.; Oikawa, H. Identification of ophiobolin F synthase by a genome mining approach: A sesterterpene synthase from Aspergillus clavatus. Org. Lett. 2013, 15, 594–597. [Google Scholar] [CrossRef] [PubMed]
- Quan, Z.; Dickschat, J.S. On the mechanism of ophiobolin F synthase and the absolute configuration of its product by isotopic labelling experiments. Org. Biomol. Chem. 2020, 18, 6072–6076. [Google Scholar] [CrossRef]
- Quan, Z.; Dickschat, J.S. Biosynthetic Gene Cluster for Asperterpenols A and B and the Cyclization Mechanism of Asperterpenol A Synthase. Org. Lett. 2020, 22, 7552–7555. [Google Scholar] [CrossRef]
- Qiao, Y.B.; Xu, Q.X.; Huang, Z.J.; Chen, X.; Ren, X.M.; Yuan, W.L.; Guan, Z.H.; Li, P.K.; Li, F.L.; Xiong, C.M.; et al. Genome mining reveals a new cyclopentane-forming sesterterpene synthase with unprecedented stereo-control. Org. Chem. Front. 2022, 9, 5808–5818. [Google Scholar] [CrossRef]
- Yan, D.X.; Arakelyan, J.; Wan, T.; Raina, R.; Chan, T.K.; Ahn, D.; Kushnarev, V.; Cheung, T.K.; Chan, H.C.; Choi, I.; et al. Genomics-driven derivatization of the bioactive fungal sesterterpenoid variecolin: Creation of an unnatural analogue with improved anticancer properties. Acta Pharm. Sin. B 2024, 14, 421–432. [Google Scholar] [CrossRef]
- Qiu, S.; Gilani, M.D.S.; Müller, C.; Zarazua-Navarro, R.M.; Liebal, U.; Eerlings, R.; Blank, L.M. Cultivation optimization promotes ginsenoside and universal triterpenoid production by engineered yeast. New Biotechnol. 2024, 83, 219–230. [Google Scholar] [CrossRef]
- Wang, S.; Meng, D.; Feng, M.; Li, C.; Wang, Y. Efficient Plant Triterpenoids Synthesis in Saccharomyces cerevisiae: From Mechanisms to Engineering Strategies. ACS Synth. Biol. 2024, 13, 1059–1076. [Google Scholar] [CrossRef]
- Li, Y.L.; Wang, J.; Li, L.Y.; Song, W.H.; Li, M.; Hua, X.; Wang, Y.; Yuan, J.F.; Xue, Z.Y. Natural products of pentacyclic triterpenoids: From discovery to heterologous biosynthesis. Nat. Product. Rep. 2023, 40, 1303–1353. [Google Scholar] [CrossRef]
- Lv, J.M.; Hu, D.; Gao, H.; Kushiro, T.; Awakawa, T.; Chen, G.D.; Wang, C.X.; Abe, I.; Yao, X.S. Biosynthesis of helvolic acid and identification of an unusual C-4-demethylation process distinct from sterol biosynthesis. Nat. Commun. 2017, 8, 1644. [Google Scholar] [CrossRef]
- Cao, Z.; Li, S.; Lv, J.; Gao, H.; Chen, G.; Awakawa, T.; Abe, I.; Yao, X.; Hu, D. Biosynthesis of clinically used antibiotic fusidic acid and identification of two short-chain dehydrogenase/reductases with converse stereoselectivity. Acta Pharm. Sin. B 2019, 9, 433–442. [Google Scholar] [CrossRef]
- Li, X.Y.; Lv, J.M.; Cao, Z.Q.; Wang, G.Q.; Lin, F.L.; Chen, G.D.; Qin, S.Y.; Hu, D.; Gao, H.; Yao, X.S. Biosynthetic characterization of the antifungal fernane-type triterpenoid polytolypin for generation of new analogues via combinatorial biosynthesis. Org. Biomol. Chem. 2023, 21, 851–857. [Google Scholar] [CrossRef]
- Cao, Z.Q.; Wang, G.Q.; Luo, R.; Gao, Y.H.; Lv, J.M.; Qin, S.Y.; Chen, G.D.; Awakawa, T.; Bao, X.F.; Mei, Q.H.; et al. Biosynthesis of Enfumafungin-type Antibiotic Reveals an Unusual Enzymatic Fusion Pattern and Unprecedented C-C Bond Cleavage. J. Am. Chem. Soc. 2024, 146, 12723–12733. [Google Scholar] [CrossRef]
- Han, H.; Peng, S.; Wang, Q.; Wang, H.; Wang, P.; Li, C.; Qi, J.; Liu, C. Biochemical characterization of a multiple prenyltransferase from Tolypocladium inflatum. Appl. Microbiol. Biotechnol. 2024, 108, 275. [Google Scholar] [CrossRef] [PubMed]
- Guan, Z.; Yao, N.; Yuan, W.; Li, F.; Xiao, Y.; Rehmutulla, M.; Xie, Y.; Chen, C.; Zhu, H.; Zhou, Y.; et al. Total biosynthesis of cotylenin diterpene glycosides as 14-3-3 protein-protein interaction stabilizers. Chem. Sci. 2025, 16, 867–875. [Google Scholar] [CrossRef] [PubMed]
- Matsuda, Y.; Mitsuhashi, T.; Lee, S.; Hoshino, M.; Mori, T.; Okada, M.; Zhang, H.; Hayashi, F.; Fujita, M.; Abe, I. Astellifadiene: Structure Determination by NMR Spectroscopy and Crystalline Sponge Method, and Elucidation of its Biosynthesis. Angew. Chem. Int. Ed. Engl. 2016, 55, 5785–5788. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.H.; Lv, J.M.; Wang, Q.Z.; Zou, J.; Lu, Y.J.; Wang, Q.L.; Chen, D.N.; Yao, X.S.; Gao, H.; Hu, D. Biosynthesis of an anti-tuberculosis sesterterpenoid asperterpenoid A. Org. Biomol. Chem. 2019, 17, 248–251. [Google Scholar] [CrossRef]
- Hertweck, C. The biosynthetic logic of polyketide diversity. Angew. Chem. Int. Ed. Engl. 2009, 48, 4688–4716. [Google Scholar] [CrossRef]
- Nivina, A.; Yuet, K.P.; Hsu, J.; Khosla, C. Evolution and Diversity of Assembly-Line Polyketide Synthases. Chem. Rev. 2019, 119, 12524–12547. [Google Scholar] [CrossRef]
- Paulsel, T.Q.; Williams, G.J. Current State-of-the-Art Toward Chemoenzymatic Synthesis of Polyketide Natural Products. Chembiochem 2023, 24, e202300386. [Google Scholar] [CrossRef]
- He, Y.; Cox, R.J. The molecular steps of citrinin biosynthesis in fungi. Chem. Sci. 2016, 7, 2119–2127. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, S.; Matsuyama, T.; Ozaki, T.; Takino, J.; Sato, H.; Uchiyama, M.; Minami, A.; Oikawa, H. Elucidation of Late-Stage Biosynthesis of Phomoidride: Proposal of Cyclization Mechanism Affording Characteristic Nine-Membered Ring of Fungal Dimeric Anhydride. J. Am. Chem. Soc. 2022, 144, 20998–21004. [Google Scholar] [CrossRef] [PubMed]
- Han, H.; Yu, C.; Qi, J.; Wang, P.; Zhao, P.; Gong, W.; Xie, C.; Xia, X.; Liu, C. High-efficient production of mushroom polyketide compounds in a platform host Aspergillus oryzae. Microb. Cell Fact. 2023, 22, 60. [Google Scholar] [CrossRef] [PubMed]
- Huang, Z.D.; Zhang, S.X.; Wang, Y.; Song, Z.W.; Wang, W.Y.; Yin, C.P.; Zhang, Y.L. Biosynthesis of Physcion and Identification of an O-Methyltransferase with C6-OH Selectivity in Aspergillus chevalieri BYST01. ACS Chem. Biol. 2025, 20, 1048–1058. [Google Scholar] [CrossRef]
- Schwarzer, D.; Finking, R.; Marahiel, M.A. Nonribosomal peptides: From genes to products. Nat. Prod. Rep. 2003, 20, 275–287. [Google Scholar] [CrossRef]
- Süssmuth, R.D.; Mainz, A. Nonribosomal Peptide Synthesis-Principles and Prospects. Angew. Chem. Int. Ed. Engl. 2017, 56, 3770–3821. [Google Scholar] [CrossRef]
- de Mattos-Shipley, K.M.J.; Greco, C.; Heard, D.M.; Hough, G.; Mulholland, N.P.; Vincent, J.L.; Micklefield, J.; Simpson, T.J.; Willis, C.L.; Cox, R.J.; et al. The cycloaspeptides: Uncovering a new model for methylated nonribosomal peptide biosynthesis. Chem. Sci. 2018, 9, 4109–4117. [Google Scholar] [CrossRef]
- Qi, J.; Han, H.; Sui, D.; Tan, S.; Liu, C.; Wang, P.; Xie, C.; Xia, X.; Gao, J.M.; Liu, C. Efficient production of a cyclic dipeptide (cyclo-TA) using heterologous expression system of filamentous fungus Aspergillus oryzae. Microb. Cell Fact. 2022, 21, 146. [Google Scholar] [CrossRef]
- Katayama, T.; Jiang, Y.; Ozaki, T.; Oikawa, H.; Minami, A.; Maruyama, J.I. Subcellular compartmentalized localization of transmembrane proteins essential for production of fungal cyclic peptide cyclochlorotine. Biosci. Biotechnol. Biochem. 2024, 88, 1279–1288. [Google Scholar] [CrossRef]
- Pinheiro, Â.; Piontkivska, D.; Sequeira, P.; Martins, T.M.; Silva Pereira, C. Aspergillus nidulans . Trends Microbiol. 2023, 31, 212–213. [Google Scholar] [CrossRef]
- Lin, S.Y.; Oakley, C.E.; Jenkinson, C.B.; Chiang, Y.M.; Lee, C.K.; Jones, C.G.; Seidler, P.M.; Nelson, H.M.; Todd, R.B.; Wang, C.C.C.; et al. A heterologous expression platform in Aspergillus nidulans for the elucidation of cryptic secondary metabolism biosynthetic gene clusters: Discovery of the Aspergillus fumigatus sartorypyrone biosynthetic pathway. Chem. Sci. 2023, 14, 11022–11032. [Google Scholar] [CrossRef]
- Hu, J.; Li, H.; Chooi, Y.H. Fungal Dirigent Protein Controls the Stereoselectivity of Multicopper Oxidase-Catalyzed Phenol Coupling in Viriditoxin Biosynthesis. J. Am. Chem. Soc. 2019, 141, 8068–8072. [Google Scholar] [CrossRef]
- Li, H.; Hu, J.; Wei, H.; Solomon, P.S.; Stubbs, K.A.; Chooi, Y.H. Biosynthesis of a Tricyclo [6.2.2.0(2,7)] dodecane System by a Berberine Bridge Enzyme-Like Aldolase. Chemistry 2019, 25, 15062–15066. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Gilchrist, C.L.M.; Phan, C.S.; Lacey, H.J.; Vuong, D.; Moggach, S.A.; Lacey, E.; Piggott, A.M.; Chooi, Y.H. Biosynthesis of a New Benzazepine Alkaloid Nanangelenin A from Aspergillus nanangensis Involves an Unusual l-Kynurenine-Incorporating NRPS Catalyzing Regioselective Lactamization. J. Am. Chem. Soc. 2020, 142, 7145–7152. [Google Scholar] [CrossRef] [PubMed]
- Yuan, G.Y.; Zhang, J.M.; Xu, Q.D.; Zhang, H.R.; Hu, C.; Zou, Y. Biosynthesis of Cosmosporasides Reveals the Assembly Line for Fungal Hybrid Terpenoid Saccharides. Angew. Chem. Int. Ed. Engl. 2023, 62, e202308887. [Google Scholar] [CrossRef] [PubMed]
- He, Q.; Zhang, H.R.; Zou, Y. A Cytochrome P450 Catalyzes Oxidative Coupling Formation of Insecticidal Dimeric Indole Piperazine Alkaloids. Angew. Chem. Int. Ed. Engl. 2024, 63, e202404000. [Google Scholar] [CrossRef]
- Yang, W.; Tian, S.; Du, Y.F.; Zeng, X.L.; Liang, J.J.; Lan, W.J.; Li, H. Genome Mining of the Marine-Derived Fungus Trichoderma erinaceum F1-1 Unearths Bergamotene-Type Sesquiterpenoids. J. Nat. Prod. 2024, 87, 2746–2756. [Google Scholar] [CrossRef]
- Smith, D.J.; Burnham, M.K.; Edwards, J.; Earl, A.J.; Turner, G. Cloning and heterologous expression of the penicillin biosynthetic gene cluster from Penicillum chrysogenum. Biotechnology 1990, 8, 39–41. [Google Scholar] [CrossRef]
- Park, D.; Swayambhu, G.; Pfeifer, B.A. Heterologous biosynthesis as a platform for producing new generation natural products. Curr. Opin. Biotechnol. 2020, 66, 123–130. [Google Scholar] [CrossRef]
- Yeh, H.H.; Chiang, Y.M.; Entwistle, R.; Ahuja, M.; Lee, K.H.; Bruno, K.S.; Wu, T.K.; Oakley, B.R.; Wang, C.C. Molecular genetic analysis reveals that a nonribosomal peptide synthetase-like (NRPS-like) gene in Aspergillus nidulans is responsible for microperfuranone biosynthesis. Appl. Microbiol. Biotechnol. 2012, 96, 739–748. [Google Scholar] [CrossRef]
- Meyer, V.; Fiedler, M.; Nitsche, B.; King, R. The Cell Factory Aspergillus Enters the Big Data Era: Opportunities and Challenges for Optimising Product Formation. Adv. Biochem. Eng. Biotechnol. 2015, 149, 91–132. [Google Scholar] [CrossRef]
- Zaehle, C.; Gressler, M.; Shelest, E.; Geib, E.; Hertweck, C.; Brock, M. Terrein biosynthesis in Aspergillus terreus and its impact on phytotoxicity. Chem. Biol. 2014, 21, 719–731. [Google Scholar] [CrossRef] [PubMed]
- Löhr, N.A.; Eisen, F.; Thiele, W.; Platz, L.; Motter, J.; Hüttel, W.; Gressler, M.; Müller, M.; Hoffmeister, D. Unprecedented Mushroom Polyketide Synthases Produce the Universal Anthraquinone Precursor. Angew. Chem. Int. Ed. Engl. 2022, 61, e202116142. [Google Scholar] [CrossRef] [PubMed]
- Maruyama, J.I. Genome Editing Technology and Its Application Potentials in the Industrial Filamentous Fungus Aspergillus oryzae. J. Fungi 2021, 7, 638. [Google Scholar] [CrossRef] [PubMed]
- Yuan, G.; Deng, S.; Czajka, J.J.; Dai, Z.; Hofstad, B.A.; Kim, J.; Pomraning, K.R. CRISPR-Cas9/Cas12a systems for efficient genome editing and large genomic fragment deletions in Aspergillus niger. Front. Bioeng. Biotechnol. 2024, 12, 1452496. [Google Scholar] [CrossRef]
- Katayama, T.; Tanaka, Y.; Okabe, T.; Nakamura, H.; Fujii, W.; Kitamoto, K.; Maruyama, J. Development of a genome editing technique using the CRISPR/Cas9 system in the industrial filamentous fungus Aspergillus oryzae. Biotechnol. Lett. 2016, 38, 637–642. [Google Scholar] [CrossRef]
- Frisvad, J.C.; Møller, L.L.H.; Larsen, T.O.; Kumar, R.; Arnau, J. Safety of the fungal workhorses of industrial biotechnology: Update on the mycotoxin and secondary metabolite potential of Aspergillus niger, Aspergillus oryzae, and Trichoderma reesei. Appl. Microbiol. Biotechnol. 2018, 102, 9481–9515. [Google Scholar] [CrossRef]
- Kuivanen, J.; Wang, Y.J.; Richard, P. Engineering Aspergillus niger for galactaric acid production: Elimination of galactaric acid catabolism by using RNA sequencing and CRISPR/Cas9. Microb. Cell Fact. 2016, 15, 210. [Google Scholar] [CrossRef]
- Kuivanen, J.; Arvas, M.; Richard, P. Clustered Genes Encoding 2-Keto-l-Gulonate Reductase and l-Idonate 5-Dehydrogenase in the Novel Fungal d-Glucuronic Acid Pathway. Front. Microbiol. 2017, 8, 225. [Google Scholar] [CrossRef]
- Kuivanen, J.; Richard, P. NADPH-dependent 5-keto-D-gluconate reductase is a part of the fungal pathway for D-glucuronate catabolism. FEBS Lett. 2018, 592, 71–77. [Google Scholar] [CrossRef]
- Kuivanen, J.; Korja, V.; Holmström, S.; Richard, P. Development of microtiter plate scale CRISPR/Cas9 transformation method for Aspergillus niger based on in vitro assembled ribonucleoprotein complexes. Fungal Biol. Biotechnol. 2019, 6, 3. [Google Scholar] [CrossRef] [PubMed]
- Sarkari, P.; Marx, H.; Blumhoff, M.L.; Mattanovich, D.; Sauer, M.; Steiger, M.G. An efficient tool for metabolic pathway construction and gene integration for Aspergillus niger. Bioresour. Technol. 2017, 245, 1327–1333. [Google Scholar] [CrossRef] [PubMed]
- Meyer, V.; Wanka, F.; van Gent, J.; Arentshorst, M.; van den Hondel, C.A.; Ram, A.F. Fungal gene expression on demand: An inducible, tunable, and metabolism-independent expression system for Aspergillus niger. Appl. Environ. Microbiol. 2011, 77, 2975–2983. [Google Scholar] [CrossRef] [PubMed]
- Zheng, X.; Cairns, T.; Zheng, P.; Meyer, V.; Sun, J. Protocol for gene characterization in Aspergillus niger using 5S rRNA-CRISPR-Cas9-mediated Tet-on inducible promoter exchange. STAR Protoc. 2022, 3, 101838. [Google Scholar] [CrossRef]
- Cairns, T.C.; Feurstein, C.; Zheng, X.; Zhang, L.H.; Zheng, P.; Sun, J.; Meyer, V. Functional exploration of co-expression networks identifies a nexus for modulating protein and citric acid titres in Aspergillus niger submerged culture. Fungal Biol. Biotechnol. 2019, 6, 18. [Google Scholar] [CrossRef]
- Zhang, L.; Zheng, X.; Cairns, T.C.; Zhang, Z.; Wang, D.; Zheng, P.; Sun, J. Disruption or reduced expression of the orotidine-5′-decarboxylase gene pyrG increases citric acid production: A new discovery during recyclable genome editing in Aspergillus niger. Microb. Cell Fact. 2020, 19, 76. [Google Scholar] [CrossRef]
- Li, Y.; Li, C.; Fu, Y.; Zhang, Q.; Ma, J.; Zhou, J.; Li, J.; Du, G.; Liu, S. A CRISPR/Cas9-based visual toolkit enabling multiplex integration at specific genomic loci in Aspergillus niger. Synth. Syst. Biotechnol. 2024, 9, 209–216. [Google Scholar] [CrossRef]
- Nakamura, H.; Katayama, T.; Okabe, T.; Iwashita, K.; Fujii, W.; Kitamoto, K.; Maruyama, J.I. Highly efficient gene targeting in Aspergillus oryzae industrial strains under ligD mutation introduced by genome editing: Strain-specific differences in the effects of deleting EcdR, the negative regulator of sclerotia formation. J. Gen. Appl. Microbiol. 2017, 63, 172–178. [Google Scholar] [CrossRef]
- Katayama, T.; Nakamura, H.; Zhang, Y.; Pascal, A.; Fujii, W.; Maruyama, J.I. Forced Recycling of an AMA1-Based Genome-Editing Plasmid Allows for Efficient Multiple Gene Deletion/Integration in the Industrial Filamentous Fungus Aspergillus oryzae. Appl. Environ. Microbiol. 2019, 85, e01896-18. [Google Scholar] [CrossRef]
- Maini Rekdal, V.; van der Luijt, C.R.B.; Chen, Y.; Kakumanu, R.; Baidoo, E.E.K.; Petzold, C.J.; Cruz-Morales, P.; Keasling, J.D. Edible mycelium bioengineered for enhanced nutritional value and sensory appeal using a modular synthetic biology toolkit. Nat. Commun. 2024, 15, 2099. [Google Scholar] [CrossRef]
- Mózsik, L.; Iacovelli, R.; Bovenberg, R.A.L.; Driessen, A.J.M. Transcriptional Activation of Biosynthetic Gene Clusters in Filamentous Fungi. Front. Bioeng. Biotechnol. 2022, 10, 901037. [Google Scholar] [CrossRef] [PubMed]
- Wösten, H.A.B. Filamentous fungi for the production of enzymes, chemicals and materials. Curr. Opin. Biotechnol. 2019, 59, 65–70. [Google Scholar] [CrossRef] [PubMed]
- Park, H.S.; Jun, S.C.; Han, K.H.; Hong, S.B.; Yu, J.H. Diversity, Application, and Synthetic Biology of Industrially Important Aspergillus Fungi. Adv. Appl. Microbiol. 2017, 100, 161–202. [Google Scholar] [CrossRef]
- Feng, J.; Hauser, M.; Cox, R.J.; Skellam, E. Engineering Aspergillus oryzae for the Heterologous Expression of a Bacterial Modular Polyketide Synthase. J. Fungi 2021, 7, 1085. [Google Scholar] [CrossRef] [PubMed]
- Kotani, A.; Ozaki, T.; Takino, J.; Mochizuki, S.; Akimitsu, K.; Minami, A.; Oikawa, H. Heterologous expression of a polyketide synthase ACRTS2 in Aspergillus oryzae produces host-selective ACR toxins: Coproduction of minor metabolites. Biosci. Biotechnol. Biochem. 2022, 86, 287–293. [Google Scholar] [CrossRef]
- Jin, F.J.; Hu, S.; Wang, B.T.; Jin, L. Advances in Genetic Engineering Technology and Its Application in the Industrial Fungus Aspergillus oryzae. Front. Microbiol. 2021, 12, 644404. [Google Scholar] [CrossRef]
- Panchanawaporn, S.; Chutrakul, C.; Jeennor, S.; Anantayanon, J.; Rattanaphan, N.; Laoteng, K. Potential of Aspergillus oryzae as a biosynthetic platform for indigoidine, a non-ribosomal peptide pigment with antioxidant activity. PLoS ONE 2022, 17, e0270359. [Google Scholar] [CrossRef]
- Takahashi, T.; Masuda, T.; Koyama, Y. Enhanced gene targeting frequency in ku70 and ku80 disruption mutants of Aspergillus sojae and Aspergillus oryzae. Mol. Genet. Genom. 2006, 275, 460–470. [Google Scholar] [CrossRef]
- Mizutani, O.; Arazoe, T.; Toshida, K.; Hayashi, R.; Ohsato, S.; Sakuma, T.; Yamamoto, T.; Kuwata, S.; Yamada, O. Detailed analysis of targeted gene mutations caused by the Platinum-Fungal TALENs in Aspergillus oryzae RIB40 strain and a ligD disruptant. J. Biosci. Bioeng. 2017, 123, 287–293. [Google Scholar] [CrossRef]
- Son, Y.E.; Park, H.S. Genetic Manipulation and Transformation Methods for Aspergillus spp. Mycobiology 2021, 49, 95–104. [Google Scholar] [CrossRef]
- Liu, R.; Chen, L.; Jiang, Y.; Zhou, Z.; Zou, G. Efficient genome editing in filamentous fungus Trichoderma reesei using the CRISPR/Cas9 system. Cell Discov. 2015, 1, 15007. [Google Scholar] [CrossRef]
- Nodvig, C.S.; Nielsen, J.B.; Kogle, M.E.; Mortensen, U.H. A CRISPR-Cas9 System for Genetic Engineering of Filamentous Fungi. PLoS ONE 2015, 10, e0133085. [Google Scholar] [CrossRef]
- Matsu-Ura, T.; Baek, M.; Kwon, J.; Hong, C. Efficient gene editing in Neurospora crassa with CRISPR technology. Fungal Biol. Biotechnol. 2015, 2, 4. [Google Scholar] [CrossRef]
- Jiang, C.M.; Lv, G.B.; Tu, Y.Y.; Cheng, X.J.; Duan, Y.T.; Zeng, B.; He, B. Applications of CRISPR/Cas9 in the Synthesis of Secondary Metabolites in Filamentous Fungi. Front. Microbiol. 2021, 12, 638096. [Google Scholar] [CrossRef] [PubMed]
- Shen, Q.; Ruan, H.; Zhang, H.; Wu, T.; Zhu, K.; Han, W.; Dong, R.; Ming, T.; Qi, H.; Zhang, Y. Utilization of CRISPR-Cas genome editing technology in filamentous fungi: Function and advancement potentiality. Front. Microbiol. 2024, 15, 1375120. [Google Scholar] [CrossRef]
- Wang, D.; Jin, S.; Lu, Q.; Chen, Y. Advances and Challenges in CRISPR/Cas-Based Fungal Genome Engineering for Secondary Metabolite Production: A Review. J. Fungi 2023, 9, 362. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Yang, D.; Zhang, J.; Xu, J.; Chen, Y.E. Recent Advances in Improving Gene-Editing Specificity through CRISPR-Cas9 Nuclease Engineering. Cells 2022, 11, 2186. [Google Scholar] [CrossRef] [PubMed]
- Sakekar, A.A.; Gaikwad, S.R.; Punekar, N.S. Protein expression and secretion by filamentous fungi. J. Biosci. 2021, 46, 5. [Google Scholar] [CrossRef]
- He, Y.; Wang, B.; Chen, W.P.; Cox, R.J.; He, J.R.; Chen, F.S. Recent advances in reconstructing microbial secondary metabolites biosynthesis in Aspergillus spp. Biotechnol. Adv. 2018, 36, 739–783. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sheng, Y.; Qiu, S.; Deng, Y.; Zeng, B. Recent Advances in Heterologous Protein Expression and Natural Product Synthesis by Aspergillus. J. Fungi 2025, 11, 534. https://doi.org/10.3390/jof11070534
Sheng Y, Qiu S, Deng Y, Zeng B. Recent Advances in Heterologous Protein Expression and Natural Product Synthesis by Aspergillus. Journal of Fungi. 2025; 11(7):534. https://doi.org/10.3390/jof11070534
Chicago/Turabian StyleSheng, Yuyang, Shangkun Qiu, Yaoming Deng, and Bin Zeng. 2025. "Recent Advances in Heterologous Protein Expression and Natural Product Synthesis by Aspergillus" Journal of Fungi 11, no. 7: 534. https://doi.org/10.3390/jof11070534
APA StyleSheng, Y., Qiu, S., Deng, Y., & Zeng, B. (2025). Recent Advances in Heterologous Protein Expression and Natural Product Synthesis by Aspergillus. Journal of Fungi, 11(7), 534. https://doi.org/10.3390/jof11070534