
Flies as Vectors and Potential Sentinels for Bacterial Pathogens and Antimicrobial Resistance: A Review



Abstract
1. Introduction
2. Flies as Vectors of Bacterial Pathogens
2.1. External Carriage of Bacterial Pathogens
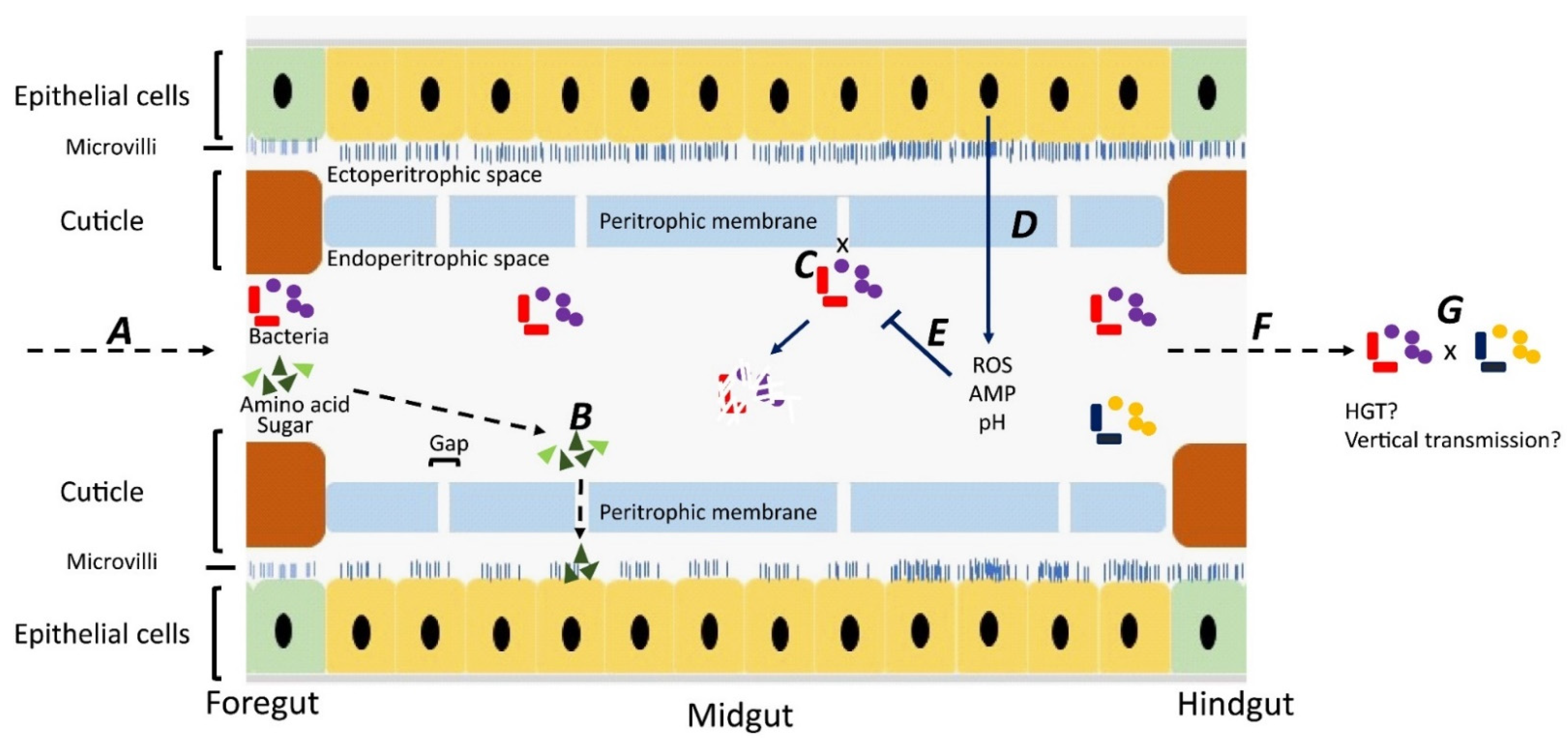
2.2. Internal Carriage of Bacterial Pathogens
3. Transmission of Antimicrobial Resistance by Flies
3.1. Horizontal Transmission
3.2. Vertical Transmission
4. Potential of Flies to Be Sentinels for Antimicrobial Resistance
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Yarger, A.M.; Fox, J.L. Dipteran halteres: Perspectives on function and integration for a unique sensory organ. Integr. Comp. Biol. 2016, 56, 865–876. [Google Scholar] [CrossRef]
- Nayduch, D.; Burrus, R.G. Flourishing in filth: House fly—Microbe interactions across life history. Ann. Entomol. Soc. Am. 2017, 110, 6–18. [Google Scholar] [CrossRef]
- Khamesipour, F.; Lankarani, K.B.; Honarvar, B.; Kwenti, T.E. A systematic review of human pathogens carried by the housefly (Musca domestica L.). BMC Public Health 2018, 18, 1049. [Google Scholar] [CrossRef]
- Ommi, D. Molecular detection and antimicrobial resistance of Aeromonas from houseflies (Musca domestica) in Iran. Rev. MVZ Córdoba 2015, 20, 4929–4936. [Google Scholar] [CrossRef][Green Version]
- de Jonge, N.; Michaelsen, T.Y.; Ejbye-Ernst, R.; Jensen, A.; Nielsen, M.E.; Bahrndorff, S.; Nielsen, J.L. Housefly (Musca domestica L.) associated microbiota across different life stages. Sci Rep. 2020, 10, 7842. [Google Scholar] [CrossRef]
- Cortinhas, L.B.; Martins Mendonça, P.; Braga, M.V.; Queiroz, M.M.C. Ultrastructure of the immature stages of Musca domestica (Diptera: Muscidae: Muscinae). J. Med. Entomol. 2020, 57, 1712–1721. [Google Scholar] [CrossRef]
- Stafford, K. Fly Management Handbook: A Guide to Biology, Dispersal, and Management of the House Fly and Related Flies for Farmers, Municipalities, and Public Health Officials; The Connecticut Agricultural Experiment Station: New Haven, CT, USA, 2008. [Google Scholar]
- Onwugamba, F.C.; Fitzgerald, J.R.; Rochon, K.; Guardabassi, L.; Alabi, A.; Kühne, S.; Grobusch, M.P.; Schaumburg, F. The role of filth ‘flies’ in the spread of antimicrobial resistance. Travel Med. Infect. Dis. 2018, 22, 8–17. [Google Scholar] [CrossRef]
- Förster, M.; Klimpel, S.; Mehlhorn, H.; Sievert, K.; Messler, S.; Pfeffer, K. Pilot study on synanthropic flies (e.g. Musca, Sarcophaga, Calliphora, Fannia, Lucilia, Stomoxys) as vectors of pathogenic microorganisms. Parasitol. Res. 2007, 101, 243–246. [Google Scholar] [CrossRef]
- Chaiwong, T.; Srivoramas, T.; Sukontason, K.; Sanford, M.R.; Moophayak, K.; Sukontason, K.L. Survey of the synanthropic flies associated with human habitations in ubon ratchathani province of northeast Thailand. J. Parasitol. Res. 2012, 2012, 613132. [Google Scholar] [CrossRef]
- Murvosh, C.M.; Thaggard, C.W. Ecological studies of the house fly. Ann. Entomol. Soc. Am. 1966, 59, 533–547. [Google Scholar] [CrossRef]
- Balaraman, V.; Drolet, B.S.; Mitzel, D.N.; Wilson, W.C.; Owens, J.; Gaudreault, N.N.; Meekins, D.A.; Bold, D.; Trujillo, J.D.; Noronha, L.E.; et al. Mechanical transmission of SARS-CoV-2 by house flies. Parasites Vectors 2021, 14, 214. [Google Scholar] [CrossRef]
- Fotedar, R.; Banerjee, U.; Samantray, S.J.C. Vector potential of hospital houseflies with special reference to Klebsiella species. Epidemiol. Infect. 1992, 109, 143–147. [Google Scholar]
- Mawak, J.D.; Olukose, O.J. Vector potential of houseflies (Musca domestica) for pathogenic organisms in Jos, Nigeria. J. Pest Dis. Vector Manag. 2006, 7, 418–423. [Google Scholar]
- Hemmatinezhad, B.; Ommi, D.; Hafshejani, T.T.; Khamesipour, F. Molecular detection and antimicrobial resistance of Pseudomonas aeruginosa from houseflies (Musca domestica) in Iran. J. Venom. Anim. Toxins Incl. Trop. Dis. 2015, 21, 18. [Google Scholar] [CrossRef][Green Version]
- Gill, C.; Bahrndorff, S.; Lowenberger, C. Campylobacter jejuni in Musca domestica: An examination of survival and transmission potential in light of the innate immune responses of the house flies. Insect Sci. 2017, 24, 584–598. [Google Scholar] [CrossRef]
- Satish, S.; Saksham, C.; Ther, S.V.; Rakesh, S.; Ravi, S.; Central Poultry Diagnostic Laboratory (Phoenix Group). Isolation and identification of enterobacterial species from Musca domestica in broiler farms of Madhya Pradesh. Vet. Pract. 2013, 14, 239–241. [Google Scholar]
- Bahrndorff, S.; de Jonge, N.; Skovgård, H.; Nielsen, J.L. Bacterial communities associated with houseflies (Musca domestica L.) sampled within and between farms. PLoS ONE 2017, 12, e0169753. [Google Scholar] [CrossRef]
- Rahuma, N.; Ghenghesh, K.S.; Ben Aissa, R.; Elamaari, A. Carriage by the housefly (Musca domestica) of multiple-antibiotic-resistant bacteria that are potentially pathogenic to humans, in hospital and other urban environments in Misurata, Libya. Ann. Trop. Med. Parasitol. 2005, 99, 795–802. [Google Scholar] [CrossRef]
- Sulaiman, S.; Othman, M.Z.; Aziz, A.H. Isolations of enteric pathogens from synanthropic flies trapped in downtown Kuala Lumpur. J. Vector Ecol. 2000, 25, 90–93. [Google Scholar]
- Graczyk, T.K.; Knight, R.; Gilman, R.H.; Cranfield, M.R. The role of non-biting flies in the epidemiology of human infectious diseases. Microbes Infect. 2001, 3, 231–235. [Google Scholar] [CrossRef]
- Wiktorczyk-Kapischke, N.; Skowron, K.; Kwiecińska-Piróg, J.; Białucha, A.; Wałecka-Zacharska, E.; Grudlewska-Buda, K.; Kraszewska, Z.; Gospodarek-Komkowska, E. Flies as a potential vector of selected alert pathogens in a hospital environment. Int. J. Environ. Health Res. 2021, 1–20. [Google Scholar] [CrossRef]
- De Jesús, A.J.; Olsen, A.R.; Bryce, J.R.; Whiting, R.C. Quantitative contamination and transfer of Escherichia coli from foods by houseflies, Musca domestica L. (Diptera: Muscidae). Int. J. Food Microbiol. 2004, 93, 259–262. [Google Scholar] [CrossRef]
- Graham, J.P.; Price, L.B.; Evans, S.L.; Graczyk, T.K.; Silbergeld, E.K. Antibiotic resistant enterococci and staphylococci isolated from flies collected near confined poultry feeding operations. Sci. Total Environ. 2009, 407, 2701–2710. [Google Scholar] [CrossRef]
- Ranjbar, R.; Izadi, M.; Hafshejani, T.T.; Khamesipour, F. Molecular detection and antimicrobial resistance of Klebsiella pneumoniae from house flies (Musca domestica) in kitchens, farms, hospitals and slaughterhouses. J. Infect. Public Health 2016, 9, 499–505. [Google Scholar] [CrossRef]
- Wang, Y.-C.; Chang, Y.-C.; Chuang, H.-L.; Chiu, C.-C.; Yeh, K.-S.; Chang, C.-C.; Hsuan, S.-L.; Lin, W.-H.; Chen, T.-H. Transmission of Salmonella between swine farms by the housefly (Musca domestica). J. Food Prot. 2011, 74, 1012–1016. [Google Scholar] [CrossRef]
- Neupane, S.; White, K.; Thomson, J.L.; Zurek, L.; Nayduch, D. Environmental and sex effects on bacterial carriage by adult house flies (Musca domestica L.). Insects 2020, 11, 401. [Google Scholar] [CrossRef]
- Joyner, C.; Mills, M.K.; Nayduch, D. Pseudomonas aeruginosa in Musca domestica L.: Temporospatial examination of bacteria population dynamics and house fly antimicrobial responses. PLoS ONE 2013, 8, e79224. [Google Scholar] [CrossRef]
- Chifanzwa, R.; Nayduch, D. Dose-dependent effects on replication and persistence of Salmonella enterica serovar Typhimurium in house flies (Diptera: Muscidae). J. Med. Entomol. 2018, 55, 225–229. [Google Scholar] [CrossRef]
- Nayduch, D.; Honko, A.; Noblet, G.P.; Stutzenberger, F. Detection of Aeromonas caviae in the common housefly Musca domestica by culture and polymerase chain reaction. Epidemiol. Infect. 2001, 127, 561–566. [Google Scholar] [CrossRef]
- Nayduch, D.; Noblet, G.P.; Stutzenberger, F.J. Vector potential of houseflies for the bacterium Aeromonas caviae. Med. Vet. Entomol. 2002, 16, 193–198. [Google Scholar] [CrossRef]
- Doud, C.W.; Zurek, L. Enterococcus faecalis OG1RF:pMV158 survives and proliferates in the house fly digestive tract. J. Med. Entomol. 2012, 49, 150–155. [Google Scholar] [CrossRef] [PubMed]
- Kelkenberg, M.; Odman-Naresh, J.; Muthukrishnan, S.; Merzendorfer, H. Chitin is a necessary component to maintain the barrier function of the peritrophic matrix in the insect midgut. Insect Biochem. Mol. Biol. 2015, 56, 21–28. [Google Scholar] [CrossRef] [PubMed]
- Lemaitre, B.; Miguel-Aliaga, I. The digestive tract of Drosophila melanogaster. Annu. Rev. Genet. 2013, 47, 377–404. [Google Scholar] [CrossRef] [PubMed]
- Lehane, M.J.; Msangi, A.R. Lectin and peritrophic membrane development in the gut of Glossina m.morsitans and a discussion of their role in protecting the fly against trypanosome infection. Med. Vet. Entomol. 1991, 5, 495–501. [Google Scholar] [CrossRef] [PubMed]
- Lehane, M.J. Peritrophic matrix structure and function. Annu. Rev. Entomol. 1997, 42, 525–550. [Google Scholar] [CrossRef]
- Kumar, N.H.; Nayduch, D. Dose-dependent fate of GFP-expressing Escherichia coli in the alimentary canal of adult house flies. Med. Vet. Entomol. 2016, 30, 218–228. [Google Scholar] [CrossRef]
- Greenberg, B.; Kowalski, J.A.; Klowden, M.J. Factors affecting the transmission of salmonella by flies: Natural resistance to colonization and bacterial interference. Infect. Immun. 1970, 2, 800–809. [Google Scholar] [CrossRef]
- Rochon, K.; Lysyk, T.J.; Selinger, L.B. Retention of Escherichia coli by house fly and stable fly (Diptera: Muscidae) during pupal metamorphosis and eclosion. J. Med. Entomol. 2005, 42, 397–403. [Google Scholar] [CrossRef][Green Version]
- Mramba, F.; Broce, A.B.; Zurek, L. Vector competence of stable flies, Stomoxys calcitrans L. (Diptera: Muscidae), for Enterobacter sakazakii. J. Vector Ecol. 2007, 32, 134–139. [Google Scholar] [CrossRef]
- Pava-Ripoll, M.; Pearson, R.E.; Miller, A.K.; Ziobro, G.C. Prevalence and relative risk of Cronobacter spp., Salmonella spp., and Listeria monocytogenes associated with the body surfaces and guts of individual filth flies. Appl. Environ. Microbiol. 2012, 78, 7891–7902. [Google Scholar] [CrossRef]
- Gwenzi, W.; Chaukura, N.; Muisa-Zikali, N.; Teta, C.; Musvuugwa, T.; Rzymski, P.; Abia, A.L.K. Insects, rodents, and pets as reservoirs, vectors, and sentinels of antimicrobial resistance. Antibiotics 2021, 10, 68. [Google Scholar] [CrossRef] [PubMed]
- Fukuda, A.; Usui, M.; Okubo, T.; Tamura, Y. Horizontal transfer of plasmid-mediated cephalosporin resistance genes in the intestine of houseflies (Musca domestica). Microb. Drug Resist. 2016, 22, 336–341. [Google Scholar] [CrossRef] [PubMed]
- Akhtar, M.; Hirt, H.; Zurek, L. Horizontal transfer of the tetracycline resistance gene tetM mediated by pCF10 among Enterococcus faecalis in the house fly (Musca domestica L.) alimentary canal. Microb. Ecol. 2009, 58, 509–518. [Google Scholar] [CrossRef] [PubMed]
- Petridis, M.; Bagdasarian, M.; Waldor, M.K.; Walker, E. Horizontal transfer of Shiga toxin and antibiotic resistance genes among Escherichia coli strains in house fly (Diptera: Muscidae) gut. J. Med. Entomol. 2006, 43, 288–295. [Google Scholar] [CrossRef]
- Zurek, L.; Ghosh, A. Insects represent a link between food animal farms and the urban environment for antibiotic resistance traits. Appl. Environ. Microbiol. 2014, 80, 3562–3567. [Google Scholar] [CrossRef]
- Pava-Ripoll, M.; Pearson, R.E.; Miller, A.K.; Tall, B.D.; Keys, C.E.; Ziobro, G.C. Ingested Salmonella enterica, Cronobacter sakazakii, Escherichia coli O157:H7, and Listeria monocytogenes: Transmission dynamics from adult house flies to their eggs and first filial (F1) generation adults. BMC Microbiol. 2015, 15, 150. [Google Scholar] [CrossRef]
- Fukuda, A.; Usui, M.; Okamura, M.; Dong-Liang, H.; Tamura, Y. Role of flies in the maintenance of antimicrobial resistance in farm environments. Microb. Drug Resist. 2019, 25, 127–132. [Google Scholar] [CrossRef]
- Hoelzer, K.; Wong, N.; Thomas, J.; Talkington, K.; Jungman, E.; Coukell, A. Antimicrobial drug use in food-producing animals and associated human health risks: What, and how strong, is the evidence? BMC Vet. Res. 2017, 13, 211. [Google Scholar] [CrossRef]
- Dallal, M.M.S.; Motalebi, S.; Asl, H.M.; Yazdi, M.K.S.; Rahimi Forushani, A. Antimicrobial investigation on the multi-state outbreak of salmonellosis and shigellosis in Iran. Med. J. Islam. Repub. Iran 2020, 34, 49. [Google Scholar] [CrossRef]
- Spellberg, B.; Blaser, M.; Guidos, R.J.; Boucher, H.W.; Bradley, J.S.; Eisenstein, B.I.; Gerding, D.; Lynfield, R.; Reller, L.B.; Rex, J.; et al. Combating antimicrobial resistance: Policy recommendations to save lives. Clin. Infect. Dis. 2011, 52 (Suppl. 5), S397–S428. [Google Scholar] [CrossRef]
- Tacconelli, E.; Carrara, E.; Savoldi, A.; Harbarth, S.; Mendelson, M.; Monnet, D.L.; Pulcini, C.; Kahlmeter, G.; Kluytmans, J.; Carmeli, Y.; et al. Discovery, research, and development of new antibiotics: The WHO priority list of antibiotic-resistant bacteria and tuberculosis. Lancet Infect. Dis. 2018, 18, 318–327. [Google Scholar] [CrossRef]
- Mather, A.E.; Reeve, R.; Mellor, D.J.; Matthews, L.; Reid-Smith, R.J.; Dutil, L.; Haydon, D.T.; Reid, S.W. Detection of rare antimicrobial resistance profiles by active and passive surveillance approaches. PLoS ONE 2016, 11, e0158515. [Google Scholar] [CrossRef] [PubMed]
- Cheah, A.L.Y.; Cheng, A.C.; Spelman, D.; Nation, R.L.; Kong, D.C.M.; McBryde, E.S. Mathematical modelling of vancomycin-resistant enterococci transmission during passive surveillance and active surveillance with contact isolation highlights the need to identify and address the source of acquisition. BMC Infect. Dis. 2018, 18, 511. [Google Scholar] [CrossRef] [PubMed]
- Kadykalo, S.V.; Anderson, M.E.C.; Alsop, J.E. Passive surveillance of antimicrobial resistance in Salmonella and Escherichia coli isolates from Ontario livestock, 2007–2015. Can. Vet. J. 2018, 59, 617–622. [Google Scholar]
- Rempel, O.R.; Laupland, K.B. Surveillance for antimicrobial resistant organisms: Potential sources and magnitude of bias. Epidemiol. Infect. 2009, 137, 1665–1673. [Google Scholar] [CrossRef]
- Agarwal, R.; Mohapatra, S.; Rath, G.P.; Kapil, A. Active surveillance of health care associated infections in neurosurgical patients. J. Clin. Diagn. Res. 2017, 11, DC01–DC04. [Google Scholar] [CrossRef]
- Huijbers, P.M.; Blaak, H.; de Jong, M.C.; Graat, E.A.; Vandenbroucke-Grauls, C.M.; de Roda Husman, A.M. Role of the environment in the transmission of antimicrobial resistance to humans: A review. Environ. Sci. Technol. 2015, 49, 11993–12004. [Google Scholar] [CrossRef]
- Barro, N.; Aly, S.; Tidiane, O.C.; Sababénédjo, T.A. Carriage of bacteria by proboscises, legs, and feces of two species of flies in street food vending sites in Ouagadougou, Burkina Faso. J. Food Prot. 2006, 69, 2007–2010. [Google Scholar] [CrossRef]
- Schaumburg, F.; Onwugamba, F.C.; Akulenko, R.; Peters, G.; Mellmann, A.; Köck, R.; Becker, K. A geospatial analysis of flies and the spread of antimicrobial resistant bacteria. Int. J. Med. Microbiol. 2016, 306, 566–571. [Google Scholar] [CrossRef]
- Gupta, A.K.; Nayduch, D.; Verma, P.; Shah, B.; Ghate, H.V.; Patole, M.S.; Shouche, Y.S. Phylogenetic characterization of bacteria in the gut of house flies (Musca domestica L.). FEMS Microbiol. Ecol. 2012, 79, 581–593. [Google Scholar] [CrossRef]
- Sproston, E.L.; Ogden, I.D.; MacRae, M.; Forbes, K.J.; Dallas, J.F.; Sheppard, S.K.; Cody, A.; Colles, F.; Wilson, M.J.; Strachan, N.J. Multi-locus sequence types of Campylobacter carried by flies and slugs acquired from local ruminant faeces. J. Appl. Microbiol. 2010, 109, 829–838. [Google Scholar] [CrossRef] [PubMed]
- Pava-Ripoll, M.; Pearson, R.E.; Miller, A.K.; Ziobro, G.C. Detection of foodborne bacterial pathogens from individual filth flies. J. Vis. Exp. 2015, 96, e52372. [Google Scholar] [CrossRef] [PubMed]
- Fukushima, H.; Ito, Y.; Saito, K.; Tsubokura, M.; Otsuki, K. Role of the fly in the transport of Yersinia enterocolitica. Appl. Environ. Microbiol. 1979, 38, 1009–1010. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Yao, Y.; Wei, P.; Song, C.; Wan, S.; Yang, S.; Zhu, G.M.; Liu, H.M. Housefly Phormicin inhibits Staphylococcus aureus and MRSA by disrupting biofilm formation and altering gene expression in vitro and in vivo. Int. J. Biol. Macromol. 2021, 167, 1424–1434. [Google Scholar] [CrossRef] [PubMed]
- Keen, J.E.; Wittum, T.E.; Dunn, J.R.; Bono, J.L.; Durso, L.M. Shiga-toxigenic Escherichia coli O157 in agricultural fair livestock, United States. Emerg. Infect. Dis. 2006, 12, 780–786. [Google Scholar] [CrossRef] [PubMed]
- Holt, P.S.; Geden, C.J.; Moore, R.W.; Gast, R.K. Isolation of Salmonella enterica serovar Enteritidis from houseflies (Musca domestica) found in rooms containing Salmonella serovar Enteritidis-challenged hens. Appl. Environ. Microbiol. 2007, 73, 6030–6035. [Google Scholar] [CrossRef]
- Punyadi, P.; Thongngen, P.; Kiddee, A.; Assawatheptawee, K.; Tansawai, U.; Bunchu, N.; Niumsup, P.R. Prevalence of bla(CTX-M) and emergence of bla(CTX-M-5)-carrying Escherichia coli in Chrysomya megacephala (Diptera: Calliphoridae), Northern Thailand. Microb. Drug Resist. 2021, 27, 698–705. [Google Scholar] [CrossRef]
- Ngbede, E.O.; Poudel, A.; Kalalah, A.; Yang, Y.; Adekanmbi, F.; Adikwu, A.A.; Adamu, A.M.; Mamfe, L.M.; Daniel, S.T.; Useh, N.M.; et al. Identification of mobile colistin resistance genes (mcr-1.1, mcr-5 and mcr-8.1) in Enterobacteriaceae and Alcaligenes faecalis of human and animal origin, Nigeria. Int. J. Antimicrob. Agents 2020, 56, 106108. [Google Scholar] [CrossRef]
- Rosef, O.; Kapperud, G. House flies (Musca domestica) as possible vectors of Campylobacter fetus subsp. jejuni. Appl. Environ. Microbiol. 1983, 45, 381–383. [Google Scholar] [CrossRef]
- Macovei, L.; Zurek, L. Influx of enterococci and associated antibiotic resistance and virulence genes from ready-to-eat food to the human digestive tract. Appl. Environ. Microbiol. 2007, 73, 6740–6747. [Google Scholar] [CrossRef]
- Macovei, L.; Miles, B.; Zurek, L. Potential of houseflies to contaminate ready-to-eat food with antibiotic-resistant enterococci. J. Food Prot. 2008, 71, 435–439. [Google Scholar] [CrossRef] [PubMed]
- Poudel, A.; Hathcock, T.; Butaye, P.; Kang, Y.; Price, S.; Macklin, K.; Walz, P.; Cattley, R.; Kalalah, A.; Adekanmbi, F.; et al. Multidrug-resistant Escherichia coli, Klebsiella pneumoniae and Staphylococcus spp. in houseflies and blowflies from farms and their environmental settings. Int. J. Environ. Res. Public Health 2019, 16, 3583. [Google Scholar] [CrossRef] [PubMed]
- Poudel, A.; Kang, Y.; Mandal, R.K.; Kalalah, A.; Butaye, P.; Hathcock, T.; Kelly, P.; Walz, P.; Macklin, K.; Cattley, R.; et al. Comparison of microbiota, antimicrobial resistance genes and mobile genetic elements in flies and the feces of sympatric animals. FEMS Microbiol. Ecol. 2020, 96, fiaa027. [Google Scholar] [CrossRef]
- Bouamamaa, L.; Sorlozano, A.; Laglaoui, A.; Lebbadi, M.; Aarab, A.; Gutierrez, J. Antibiotic resistance patterns of bacterial strains isolated from Periplaneta americana and Musca domestica in Tangier, Morocco. J. Infect. Dev. Ctries. 2010, 4, 194–201. [Google Scholar] [CrossRef][Green Version]
- Usui, M.; Iwasa, T.; Fukuda, A.; Sato, T.; Okubo, T.; Tamura, Y. The role of flies in spreading the extended-spectrum β-lactamase gene from cattle. Microb. Drug Resist. 2013, 19, 415–420. [Google Scholar] [CrossRef] [PubMed]
- Blaak, H.; Hamidjaja, R.A.; van Hoek, A.H.; de Heer, L.; de Roda Husman, A.M.; Schets, F.M. Detection of extended-spectrum beta-lactamase (ESBL)-producing Escherichia coli on flies at poultry farms. Appl. Environ. Microbiol. 2014, 80, 239–246. [Google Scholar] [CrossRef] [PubMed]
- Solà-Ginés, M.; González-López, J.J.; Cameron-Veas, K.; Piedra-Carrasco, N.; Cerdà-Cuéllar, M.; Migura-Garcia, L. Houseflies (Musca domestica) as Vectors for Extended-Spectrum β-Lactamase-Producing Escherichia coli on Spanish Broiler Farms. Appl. Environ. Microbiol. 2015, 81, 3604–3611. [Google Scholar] [CrossRef] [PubMed]
- Songe, M.M.; Hang’ombe, B.M.; Knight-Jones, T.J.; Grace, D. Antimicrobial resistant enteropathogenic Escherichia coli and Salmonella spp. in houseflies infesting fish in food markets in Zambia. Int. J. Environ. Res. Public Health 2016, 14, 21. [Google Scholar] [CrossRef]
- Guenther, S.; Falgenhauer, L.; Semmler, T.; Imirzalioglu, C.; Chakraborty, T.; Roesler, U.; Roschanski, N. Environmental emission of multiresistant Escherichia coli carrying the colistin resistance gene mcr-1 from German swine farms. J. Antimicrob. Chemother. 2017, 72, 1289–1292. [Google Scholar] [CrossRef]
- Wadaskar, B.; Kolhe, R.; Waskar, V.; Pawade, M.; Kundu, K. Isolation of Escherichia coli and Salmonella species in flies trapped at animal farm premises. J. Entomol. Zool. Stud. 2019, 7, 198–201. [Google Scholar]
- Carramaschi, I.N.; Lopes, J.C.O.; Leite, J.A.; Carneiro, M.T.; Barbosa, R.R.; Boas, M.H.V.; Rangel, K.; Chagas, T.P.G.; Queiroz, M.M.; Zahner, V. Surveillance of antimicrobial resistant bacteria in flies (Diptera) in Rio de Janeiro city. Acta Trop. 2021, 220, 105962. [Google Scholar] [CrossRef] [PubMed]
- Alves, T.D.S.; Lara, G.H.B.; Maluta, R.P.; Ribeiro, M.G.; Leite, D.D.S. Carrier flies of multidrug-resistant Escherichia coli as potential dissemination agent in dairy farm environment. Sci. Total Environ. 2018, 633, 1345–1351. [Google Scholar] [CrossRef] [PubMed]
- Sobur, A.; Haque, Z.F.; Sabuj, A.A.; Ievy, S.; Rahman, A.T.; El Zowalaty, M.E.; Rahman, T. Molecular detection of multidrug and colistin-resistant Escherichia coli isolated from house flies in various environmental settings. Future Microbiol. 2019, 14, 847–858. [Google Scholar] [CrossRef] [PubMed]
- Tufa, T.B.; Fuchs, A.; Wienemann, T.; Eggers, Y.; Abdissa, S.; Schneider, M.; Jensen, B.O.; Bode, J.G.; Pfeffer, K.; Häussinger, D.; et al. Carriage of ESBL-producing Gram-negative bacteria by flies captured in a hospital and its suburban surroundings in Ethiopia. Antimicrob. Resist. Infect. Control 2020, 9, 175. [Google Scholar] [CrossRef]
- Yang, Q.E.; Tansawai, U.; Andrey, D.O.; Wang, S.; Wang, Y.; Sands, K.; Kiddee, A.; Assawatheptawee, K.; Bunchu, N.; Hassan, B.; et al. Environmental dissemination of mcr-1 positive Enterobacteriaceae by Chrysomya spp. (common blowfly): An increasing public health risk. Environ. Int. 2019, 122, 281–290. [Google Scholar] [CrossRef]
- Heiden, S.E.; Kurz, M.S.E.; Bohnert, J.; Bayingana, C.; Ndoli, J.M.; Sendegeya, A.; Gahutu, J.B.; Eger, E.; Mockenhaupt, F.P.; Schaufler, K. Flies from a tertiary hospital in Rwanda carry multidrug-resistant Gram-negative pathogens including extended-spectrum beta-lactamase-producing E. coli sequence type 131. Antimicrob. Resist. Infect. Control 2020, 9, 34. [Google Scholar] [CrossRef]
- Wetzker, W.; Pfeifer, Y.; Wolke, S.; Haselbeck, A.; Leistner, R.; Kola, A.; Gastmeier, P.; Salm, F. Extended-spectrum Beta-Lactamase (ESBL)-producing Escherichia coli isolated from flies in the urban center of Berlin, Germany. Int. J. Environ. Res. Public Health 2019, 16, 1530. [Google Scholar] [CrossRef]
- Odetoyin, B.; Adeola, B.; Olaniran, O. Frequency and antimicrobial resistance patterns of bacterial species isolated from the body surface of the housefly (Musca domestica) in Akure, Ondo state, Nigeria. J. Arthropod Borne Dis. 2020, 14, 88–96. [Google Scholar] [CrossRef]
- Pileggi, M.T.; Chase, J.R.; Shu, R.; Teng, L.; Jeong, K.C.; Kaufman, P.E.; Wong, A.C.N. Prevalence of field-collected house flies and stable flies with bacteria displaying cefotaxime and multidrug resistance. J. Med. Entomol. 2021, 58, 921–928. [Google Scholar] [CrossRef]


{kind=link}
| Country and Sampling Period | Collection Sites | Flies | Bacteria | Resistant Gene | Resistant Antibiotics | Ref |
|---|---|---|---|---|---|---|
| Norway, 1983 | Chicken and pig farms | Houseflies | Campylobacter jejuni, Campylobacter coli | N/A * | NAL | [70] |
| Libya, 2005 | Hospital, streets, abattoir | Houseflies | Pseudomonas, Staphylococcus aureus | N/A | AMC, AN, CPH, D, K, NAL | [19] |
| USA, 2007 | Chicken salad meal, chicken sandwich with French fries, carrot cake | Houseflies | Enterococcus casseliflavus, Enterococcus faecium, Enterococcus faecalis, Enterococcus hirae | N/A | TET, ERY, K | [71] |
| USA, 2008 | Cattle feedlot, contaminated fast food | Houseflies | Enterococcus faecalis | N/A | TET, CIP, ERY | [72] |
| USA, 2009 | Poultry farm | Houseflies, Blow flies, Bottle flies | Enterococci. Staphylococcus aureus | erm(B), erm(A), msr(C), msr(A/B) | N/A | [24] |
| Morocco, 2010 | Houses, garbage heaps, open defecating grounds | Houseflies | Enterococcus, Escherichia coli, Klebsiella, Providencia, Staphylococcus | N/A | AMG, Carbapenems | [75] |
| Taiwan, 2011 | Pig farm | Houseflies | Salmonella | N/A | AMP, AMC, C, CIP, STREP, TET, NAL | [26] |
| Japan, 2013 | Cattle barn | Houseflies, Stable flies | Escherichia coli | blaCTX-M-15 | [76] | |
| Netherlands, 2014 | Poultry farm | Houseflies, Lesser houseflies, Stable flies, Blow flies | Escherichia coli | blaTEM-52 | N/A | [77] |
| Spain, 2015 | Poultry farm | Houseflies Lesser house fliesStable flies | Escherichia coli | blaCTX-M-1, blaCTX-M-14, blaCTX-M-9. | N/A | [78] |
| Zambia, 2016 | Fish | Houseflies | Escherichia coli Salmonella | blaTEM, blaSHV | N/A | [79] |
| Germany, 2017 | Pig farm | Stable flies | Escherichia coli | Colistin | [80] | |
| Brazil, 2018 | Cattle farm | Houseflies | Escherichia coli | blaTEM, blaCTX-M | N/A | [83] |
| USA, 2019 | Dairy unit, dog kennel, poultry farm, beef cattle unit, urban trash facility and urban downtown | Houseflies Stable flies | Escherichia coli, Staphylococcus Klebsiella pneumoniae, | blaCMY-2, blaCTXM-1 | N/A | [73] |
| Bangladesh, 2019 | Hospital, livestock | Houseflies | Escherichia coli, Salmonella | N/A | AMG, ERY, OTC, TET | [84] |
| India, 2019 | Livestock | Houseflies Blow flies | Escherichia coli, Salmonella | N/A | AMX, AMP, ATM, CTX, IPM | [81] |
| Germany, 2019 | Hospital, household, zoo, streets | Houseflies Blow flies Flesh flies | Acinetobacter, Citrobacter, Enterobacter, Escherichia coli, Klebsiella, Pseudomonas aeruginosa, Raoultella | N/A | AMP, AMX, CEP, GEN, STREP, TET | [87,88] |
| Ethiopia, 2020 | Hospital, market | Houseflies | Klebsiella, Proteus | N/A | C, GEN, SXT | [85] |
| Nigeria, 2020 | Slaughterhouse trash, hospital | Houseflies | Proteus, Salmonella Pseudomonas aeruginosa, | N/A | AMX, AUG, GEN, STREP, SXT, TET | [89] |
| Thailand, 2021 | Livestock | Blow flies | Enterobacteriaceae Escherichia coli | N/A | AMP, CEP, STREP | [68] |
| Brazil, 2021 | Trash | Houseflies Blow flies | Enterobacter, Escherichia coli, Klebsiella, Raoultella, Serratia, | N/A | C, CTX, GEN, MEM, SXT, TET | [82] |
| USA, 2021 | Livestock facilities | Houseflies Stable flies | Bacillus, Commensalibacter, Enterococcus, Kytococcus, Oceanobacillus, Ochrobactrum | N/A | CTX | [90] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yin, J.-H.; Kelly, P.J.; Wang, C. Flies as Vectors and Potential Sentinels for Bacterial Pathogens and Antimicrobial Resistance: A Review. Vet. Sci. 2022, 9, 300. https://doi.org/10.3390/vetsci9060300
Yin J-H, Kelly PJ, Wang C. Flies as Vectors and Potential Sentinels for Bacterial Pathogens and Antimicrobial Resistance: A Review. Veterinary Sciences. 2022; 9(6):300. https://doi.org/10.3390/vetsci9060300
Chicago/Turabian StyleYin, Ji-Hang, Patrick John Kelly, and Chengming Wang. 2022. "Flies as Vectors and Potential Sentinels for Bacterial Pathogens and Antimicrobial Resistance: A Review" Veterinary Sciences 9, no. 6: 300. https://doi.org/10.3390/vetsci9060300
APA StyleYin, J.-H., Kelly, P. J., & Wang, C. (2022). Flies as Vectors and Potential Sentinels for Bacterial Pathogens and Antimicrobial Resistance: A Review. Veterinary Sciences, 9(6), 300. https://doi.org/10.3390/vetsci9060300