
Sexual Dimorphic Innate Immune Response to a Viral–Bacterial Respiratory Disease Challenge in Beef Calves


Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Design
2.2. Health and Nasal Lesion Scores
2.3. Serum Analysis
2.4. Statistical Analysis
3. Results
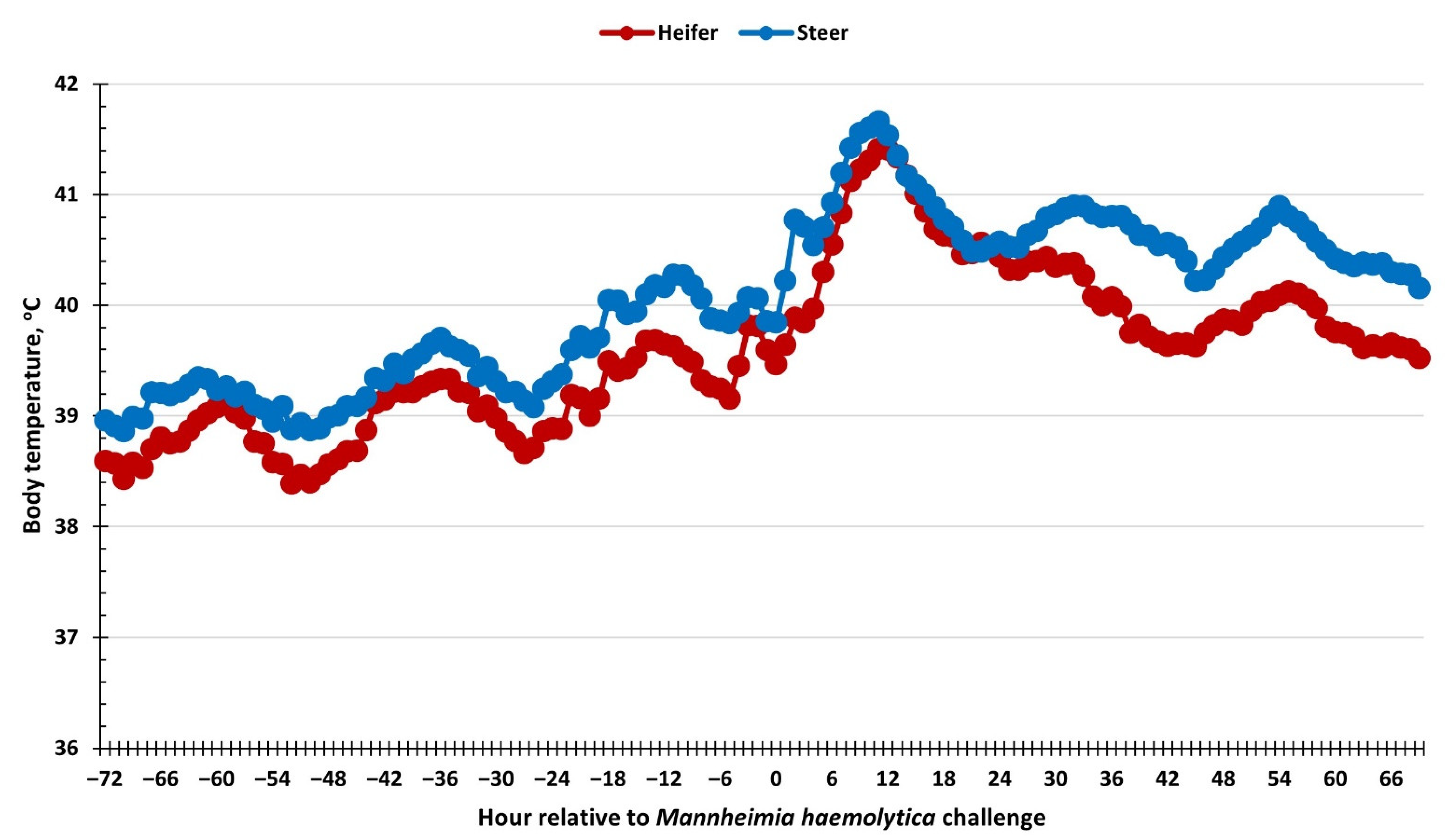
3.1. Health and Nasal Lesion Scores, Body Temperature, and Water Intake
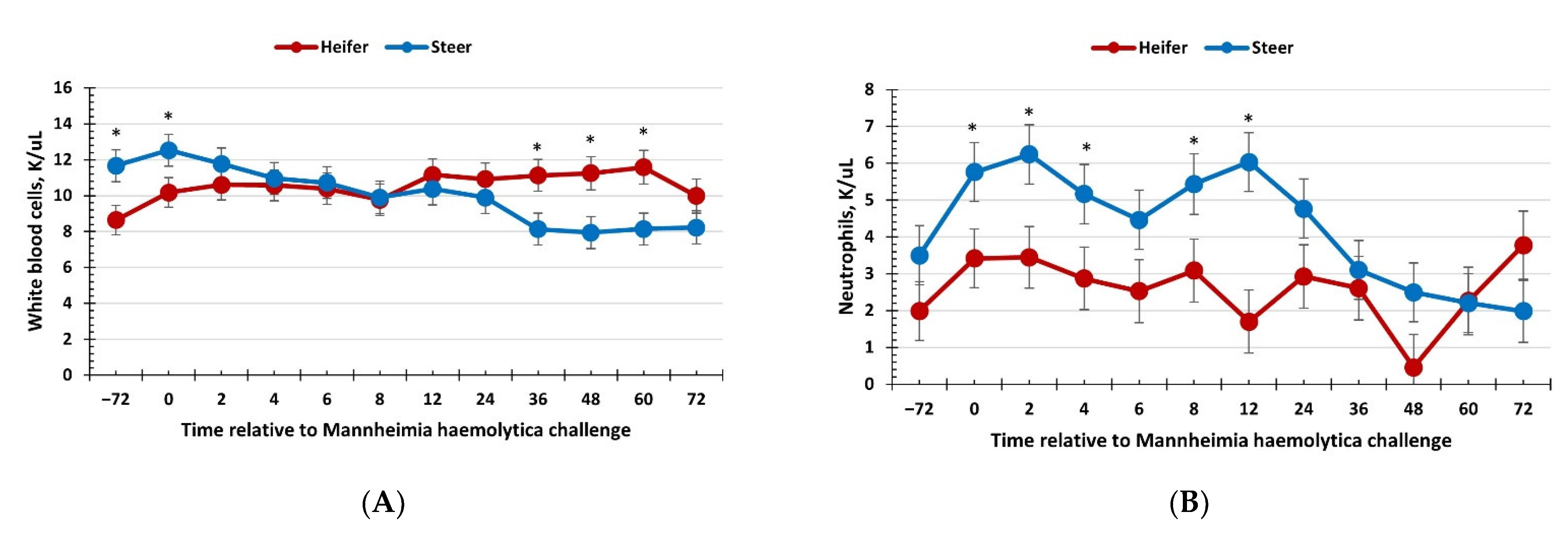
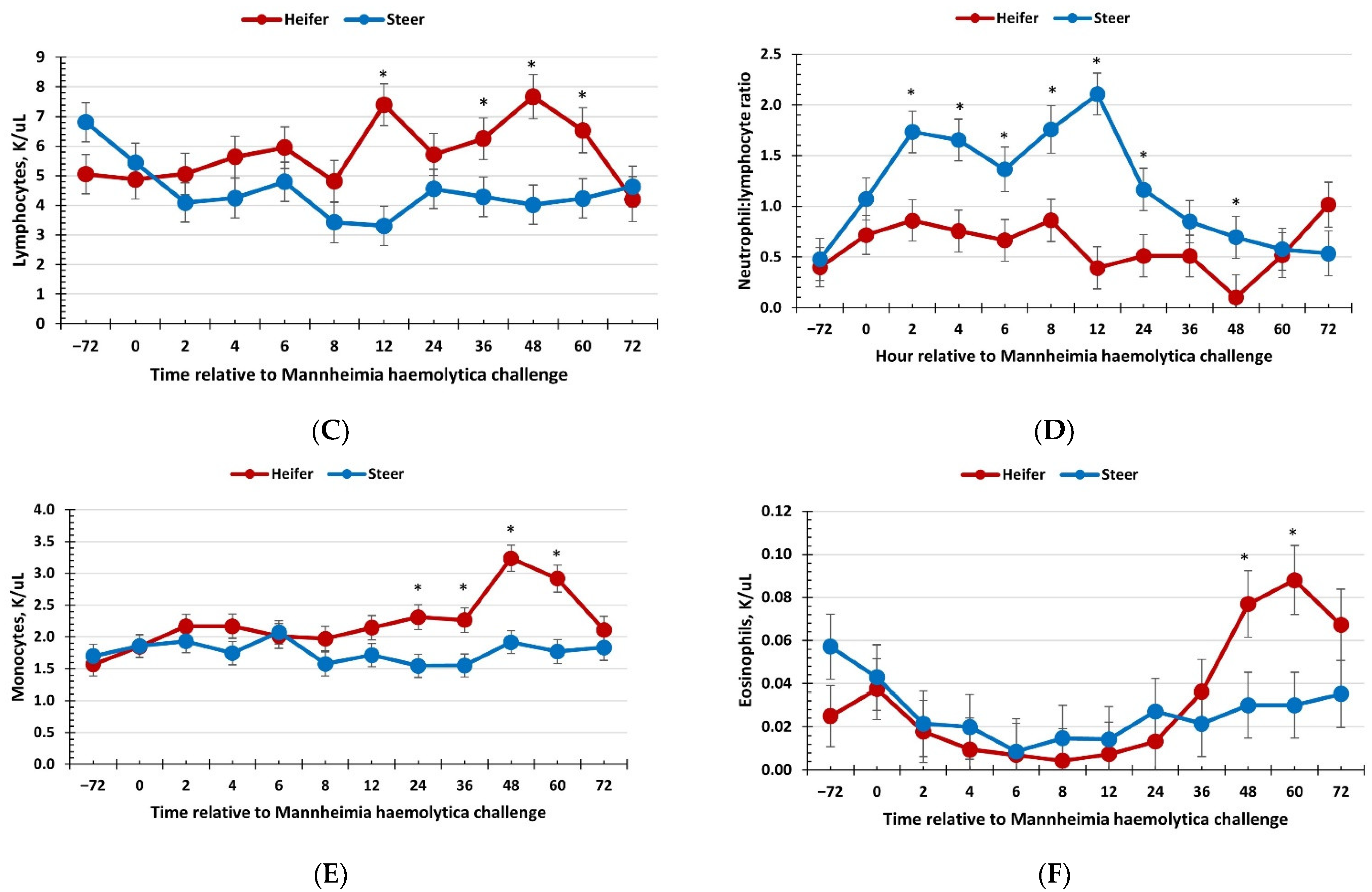
3.2. Hematology
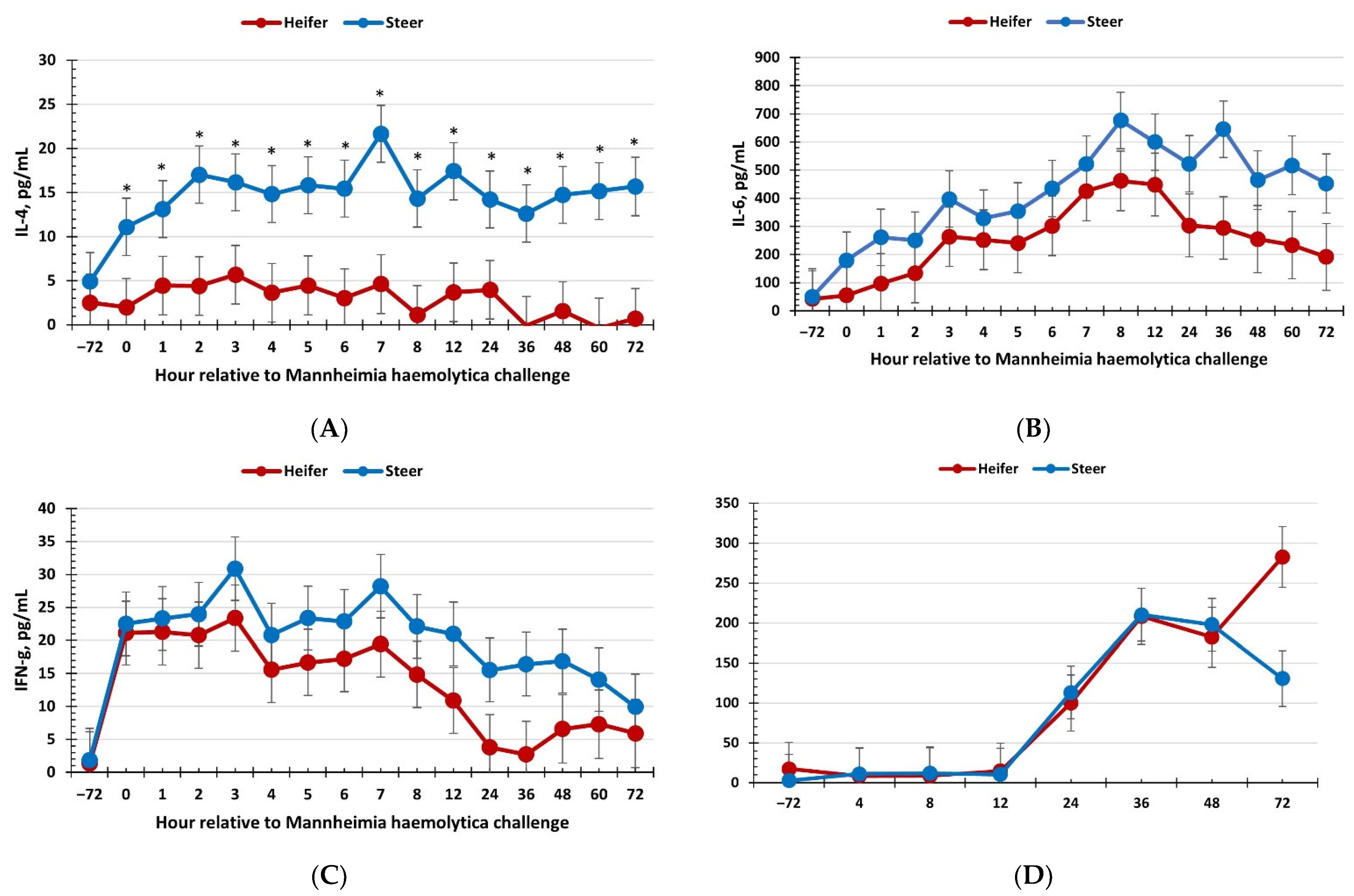
3.3. Serum Cytokines and Haptoglobin
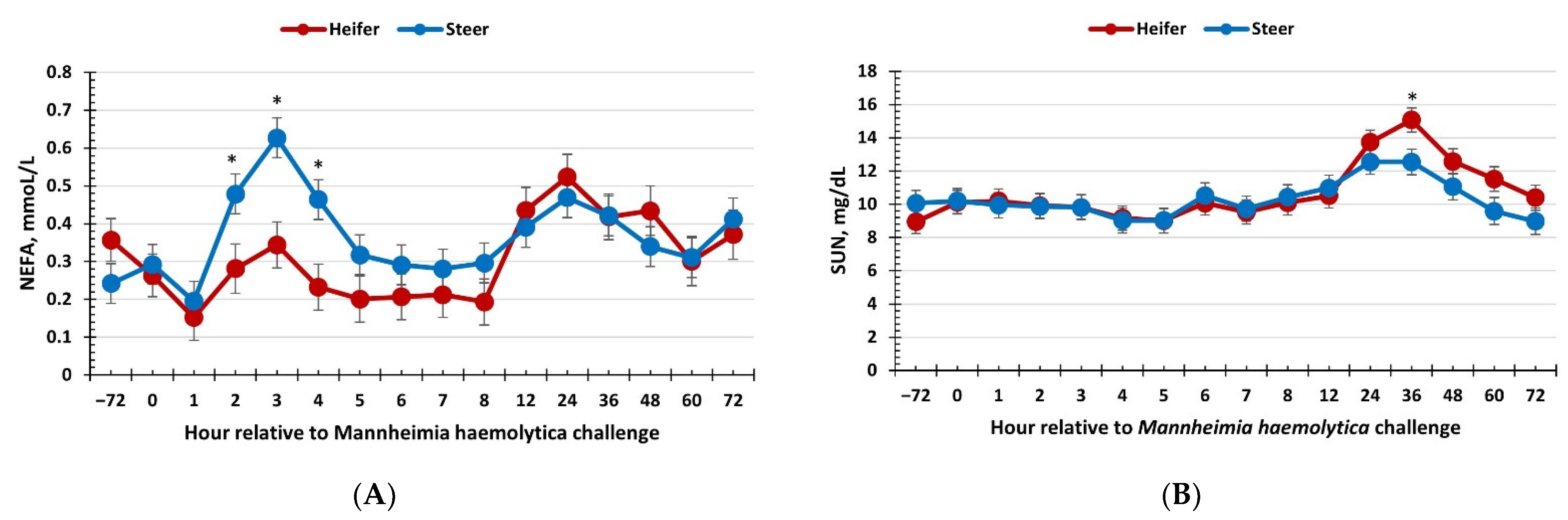
3.4. Serum Glucose, Non-Esterified Fatty Acids, and Urea Nitrogen
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Klein, S.L.; Flanagan, K.L. Sex differences in immune responses. Nat. Rev. Immunol. 2016, 16, 626–638. [Google Scholar] [CrossRef] [PubMed]
- Barker, L.A.; Dazin, P.F.; Levine, J.D.; Green, P.G. Sympathoadrenal-dependent sexually dimorphic effect of nonhabituating stress on in vivo neutrophil recruitment in the rat. Br. J. Pharmacol. 2005, 145, 872–879. [Google Scholar] [CrossRef] [PubMed]
- Canny, B.J.; O’Farrell, K.A.; Clarke, I.J.; Tilbrook, A.J. The influence of sex and gonadectomy on the hypothalamo-pituitary-adrenal axis of the sheep. J. Endocrinol. 1999, 162, 215–225. [Google Scholar] [CrossRef] [PubMed]
- Glucksmann, A. Sexual dimorphism in mammals. Biol. Rev. 1974, 49, 423–475. [Google Scholar] [CrossRef]
- Hulbert, L.E.; Carroll, J.A.; Ballou, M.A.; Burdick, N.C.; Dailey, J.W.; Caldwell, L.C.; Loyd, A.N.; Vann, R.C.; Welsh, T.H.; Randel, R.D. Sexually dimorphic stress and pro-inflammatory cytokine responses to an intravenous corticotropin-releasing hormone challenge of Brahman cattle following transportation. Innate Immun. 2013, 19, 378–387. [Google Scholar] [CrossRef]
- Carroll, J.A.; Burdick Sanchez, N.C.; Hulbert, L.E.; Ballou, M.A.; Dailey, J.W.; Caldwell, L.C.; Vann, R.C.; Welsh, T.H., Jr.; Randel, R.D. Sexually dimorphic innate immunological responses of pre-pubertal Brahman cattle following an intravenous lipopolysaccharide challenge. Vet. Immunol. Immunopathol. 2015, 166, 108–115. [Google Scholar] [CrossRef]
- Burdick Sanchez, N.C.; Broadway, P.R.; Carroll, J.A.; Gart, E.V.; Bryan, L.K.; Lawhon, S.D. Weaned pigs experimentally infected with Salmonella display sexually dimorphic innate immune rsponses without affecting pathogen colonization patterns. Transl. Anim. Sci. 2017, 1, 69–76. [Google Scholar] [CrossRef]
- APHIS. Types and Costs of Respiratory Disease Treatments in U.S. Feedlots; Animal & Plant Health Inspection Service: Riverdale Park, MD, USA, 2013. [Google Scholar]
- Blakebrough-Hall, C.; McMeniman, J.P.; Gonzalez, L.A. An evaluation of the economic effects of bovine respiratory disease on animal performance, carcass traits, and economic outcomes in feedlot cattle defined using four BRD diagnosis methods. J. Anim. Sci. 2020, 98, skaa005. [Google Scholar] [CrossRef]
- Word, A.B.; Broadway, P.R.; Burdick Sanchez, N.C.; Hutcheson, J.P.; Ellis, G.B.; Holland, B.P.; Ballou, M.A.; Carroll, J.A. Acute immunologic and metabolic responses of beef heifers following topical administration of flunixin meglumine at various times relative to bovine herpesvirus 1 and Mannheimia haemolytica challenges. Am. J. Vet. Res. 2020, 81, 243–253. [Google Scholar] [CrossRef]
- Cusack, P.M.; McMeniman, N.P.; Lean, I.J. Feedlot entry characteristics and climate: Their relationship with cattle growth rate, bovine respiratory disease and mortality. Aust. Vet. J. 2007, 85, 311–316. [Google Scholar] [CrossRef]
- Gorden, P.J.; Plummer, P. Control, management, and prevention of bovine respiratory disease in dairy calves and cows. Vet. Clin. N. Am. Food Anim. Pract. 2010, 26, 243–259. [Google Scholar] [CrossRef]
- Muggli-Cockett, N.E.; Cundiff, L.V.; Gregory, K.E. Genetic analysis of bovine respiratory disease in beef calves during the first year of life. J. Anim. Sci. 1992, 70, 2013–2019. [Google Scholar] [CrossRef]
- Reuter, R.R.; Carroll, J.A.; Hulbert, L.E.; Dailey, J.W.; Galyean, M.L. Technical note: Development of a self-contained, indwelling rectal temperature probe for cattle research. J. Anim. Sci. 2010, 88, 3291–3295. [Google Scholar] [CrossRef]
- Burdick, N.C.; Carroll, J.A.; Dailey, J.W.; Randel, R.D.; Falkenberg, S.M.; Schmidt, T.B. Development of a self-contained, indwelling vaginal temperature probe for use in cattle research. J. Therm. Biol. 2012, 37, 339–343. [Google Scholar] [CrossRef]
- Burdick Sanchez, N.C.; Carroll, J.A.; Randel, R.D.; Vann, R.C.; Welsh, T.H. Associations between endotoxin-induced metabolic changes and temperament in Brahman bulls. J. Anim. Physiol. Anim. Nutr. 2014, 98, 178–190. [Google Scholar] [CrossRef]
- Kiser, J.N.; Lawrence, T.E.; Neupane, M.; Seabury, C.M.; Taylor, J.F.; Womack, J.E.; Neibergs, H.L. Rapid communication: Subclinical bovine respiratory disease—Loci and pathogens associated with lung lesions in feedlot cattle. J. Anim. Sci. 2017, 95, 2726–2731. [Google Scholar] [CrossRef]
- Burdick, N.C.; Carroll, J.A.; Hulbert, L.E.; Dailey, J.W.; Ballou, M.A.; Randel, R.D.; Willard, S.T.; Vann, R.C.; Welsh, T.H., Jr. Temperament influences endotoxin-induced changes in rectal temperature, sickness behavior, and plasma epinephrine concentrations in bulls. Innate Immun. 2011, 17, 355–364. [Google Scholar] [CrossRef]
- Carroll, J.A.; Burdick, N.C.; Reuter, R.R.; Chase, C.C., Jr.; Spiers, D.E.; Arthington, J.D.; Coleman, S.W. Differential acute phase immune responses by Angus and Romosinuano steers following an endotoxin challenge. Domest. Anim. Endocrinol. 2011, 41, 163–173. [Google Scholar] [CrossRef]
- Evans, S.S.; Repasky, E.A.; Fisher, D.T. Fever and the thermal regulation of immunity: The immune system feels the heat. Nat. Rev. Immunol. 2015, 15, 335–349. [Google Scholar] [CrossRef]
- Kluger, M.J.; Kozak, W.; Leon, L.R.; Soszynski, D.; Conn, C.A. Cytokines and fever. Neuroimmunomodulation 1995, 2, 216–223. [Google Scholar] [CrossRef]
- Kadioglu, A.; Cuppone, A.M.; Trappetti, C.; List, T.; Spreafico, A.; Pozzi, G.; Andrew, P.W.; Oggioni, M.R. Sex-based differences in susceptibility to respiratory and systemic pneumococcal disease in mice. J. Infect. Dis. 2011, 204, 1971–1979. [Google Scholar] [CrossRef] [PubMed]
- Vernon, R.G. Effects of diet on lipolysis and its regulation. Proc. Nutr. Soc. 1992, 51, 397–408. [Google Scholar] [CrossRef] [PubMed]
- Furman, D.; Hejblum, B.P.; Simon, N.; Jojic, V.; Dekker, C.L.; Thiebaut, R.; Tibshirani, R.J.; Davis, M.M. Systems analysis of sex differences reveals an immunosuppressive role for testosterone in the response to influenza vaccination. Proc. Natl. Acad. Sci. USA 2014, 111, 869–874. [Google Scholar] [CrossRef] [PubMed]
- Abdullah, M.; Chai, P.S.; Chong, M.Y.; Tohit, E.R.M.; Ramasamy, R.; Pei, C.P.; Vidyadaran, S. Gender effect on in vitro lymphocyte subset levels of healthy individuals. Cell. Immunol. 2012, 272, 214–219. [Google Scholar] [CrossRef] [PubMed]
- Furman, D. Sexual dimorphism in immunity: Improving our understanding of vaccine immune responses in men. Expert Rev. Vaccines 2015, 14, 461–471. [Google Scholar] [CrossRef]
- Marriott, I.; Huet-Hudson, Y.M. Sexual dimorphism in innate immune responses to infectious organisms. Immunol. Res. 2006, 34, 177–192. [Google Scholar] [CrossRef]
- Lindholm-Perry, A.K.; Kuehn, L.A.; McDaneld, T.G.; Miles, J.R.; Workman, A.M.; Chitko-McKown, C.G.; Keele, J.W. Complete blood count data and leukocyte expression of cytokine genes and cytokine receptor genes associated with bovine respiratory disease in calves. BMC Res. Notes 2018, 11, 786. [Google Scholar] [CrossRef]
- Cusack, P.M.; McMeniman, N.; Lean, I.J. The medicine and epidemiology of bovine respiratory disease in feedlots. Aust. Vet. J. 2003, 81, 480–487. [Google Scholar] [CrossRef]
- Bielefeldt Ohmann, H.; Babiuk, L.A. Viral-bacterial pneumonia in calves: Effect of bovine herpesvirus-1 on immunologic functions. J. Infect. Dis. 1985, 151, 937–947. [Google Scholar] [CrossRef]
- Cuevas-Gomez, I.; McGee, M.; McCabe, M.; Cormican, P.; O’Riordan, E.; McDaneld, T.; Earley, B. Growth performance and hematological changes of weaned beef calves diagnosed with respiratory disease using respiratory scoring and thoracic ultrasonography. J. Anim. Sci. 2020, 98, 11. [Google Scholar] [CrossRef]
- Singh, K.; Ritchey, J.W.; Confer, A.W. Mannheimia haemolytica: Bacterial-host interactions in bovine Pneumonia. Vet. Pathol. 2011, 48, 338–348. [Google Scholar] [CrossRef]
- Rettew, J.A.; Huet-Hudson, Y.M.; Marriott, I. Testosterone reduces macrophage expression in the mouse of toll-like receptor 4, a trigger for inflammation and innate immunity. Biol. Reprod. 2008, 78, 432–437. [Google Scholar] [CrossRef]
- Leach, R.J.; Chitko-McKown, C.G.; Bennet, G.L.; Jones, S.A.; Kachman, S.D.; Keele, J.W.; Leymaster, K.A.; Thallman, R.M.; Kuehn, L.A. The change in differing leukocyte populations during vaccination to bovine respiratory disease and their correlations with lung scores, health records, and average daily gain. J. Anim. Sci. 2013, 91, 3564–3573. [Google Scholar] [CrossRef]
- Kita, H. Eosinophils: Multifaceted biological properties and roles in health and disease. Immunol. Rev. 2011, 242, 161–177. [Google Scholar] [CrossRef]
- Richeson, J.T.; Pinedo, P.J.; Kegley, E.B.; Powell, J.G.; Gadberry, M.S.; Beck, P.A.; Falkenberg, S.M. Association of hematologic variables and castration status at the time of arrival at a research facility with the risk of bovine respiratory disease in beef calves. J. Am. Vet. Med. Assoc. 2013, 243, 1035–1041. [Google Scholar] [CrossRef]
- Chaudhari, A.A.; Kim, W.H.; Lillehoj, H.S. Interleukin-4 (IL-4) may regulate alternative activation of macrophage-like cells in chickens: A sequential study using novel and specific neutralizing monoclonal antibodies against chicken IL-4. Vet. Immunol. Immunopathol. 2018, 205, 72–82. [Google Scholar] [CrossRef]
- Bluthe, R.M.; Michaud, B.; Poli, V.; Dantzer, R. Role of IL-6 in cytokine-induced sickness behavior: A study with IL-6 deficient mice. Physiol. Behav. 2000, 70, 367–373. [Google Scholar] [CrossRef]
- Dantzer, R.; Bluthé, R.-M.; Aubert, A.; Goodall, G.; Bret-Dibat, J.-L.; Kent, S.; Goujon, E.; Layé, S.; Parnet, P.; Kelley, K.W. Cytokine Actions on Behavior. In Cytokines in the Nervous System; Rothwell, N.J., Ed.; Springer: Boston, MA, USA, 1996; pp. 117–144. [Google Scholar] [CrossRef]
- Harden, L.M.; du Plessis, I.; Poole, S.; Laburn, H.P. Interleukin-6 and leptin mediate lipopolysaccharide-induced fever and sickness behavior. Physiol. Behav. 2006, 89, 146–155. [Google Scholar] [CrossRef]
- Burciaga-Robles, L.O.; Step, D.L.; Krehbiel, C.R.; Holland, B.P.; Richards, C.J.; Montelongo, M.A.; Confer, A.W.; Fulton, R.W. Effects of exposure to calves persistently infected with bovine viral diarrhea virus type 1b and subsequent infection with mannheima haemolytica on clinical signs and immune variables: Model for bovinerespiratory disease via viral and bacterial interaction. J. Anim. Sci. 2010, 88, 2166–2178. [Google Scholar] [CrossRef]
- Aulock, S.V.; Deininger, S.; Draing, C.; Gueinzius, K.; Dehus, O.; Hermann, C. Gender difference in cytokine secretion on immune stimulation with LPS and LTA. J. Interferon Cytokine Res. 2006, 26, 887–892. [Google Scholar] [CrossRef]
- Babiuk, L.A.; van Drunen Littel-van den Hurk, S.; Tikoo, S.K. Immunology of bovine herpesvirus 1 infection. Vet. Microbiol. 1996, 53, 31–42. [Google Scholar] [CrossRef] [PubMed]
- Leite, F.; Kuckleburg, C.; Atapattu, D.; Schultz, R.; Czuprynski, C.J. BHV-1 infection and inflammatory cytokines amplify the interaction of Mannheimia haemolytica leukotoxin with bovine peripheral blood mononuclear cells in vitro. Vet. Immunol. Immunopathol. 2004, 99, 193–202. [Google Scholar] [CrossRef] [PubMed]
- Huzzey, J.M.; Duffield, T.F.; LeBlanc, S.J.; Veira, D.M.; Weary, D.M.; von Keyserlingk, M.A. Short communication: Haptoglobin as an early indicator of metritis. J. Dairy Sci. 2009, 92, 621–625. [Google Scholar] [CrossRef] [PubMed]
- Ackermann, M.R.; Derscheid, R.; Roth, J.A. Innate immunology of bovine respiratory disease. Vet. Clin. N. Am. Food Anim. Pract. 2010, 26, 215–228. [Google Scholar] [CrossRef] [PubMed]
- Idoate, I.; Vander Ley, B.; Schultz, L.; Heller, M. Acute phase proteins in naturally occurring respiratory disease of feedlot cattle. Vet. Immunol. Immunopathol. 2015, 163, 221–226. [Google Scholar] [CrossRef] [PubMed]
- Godson, D.L.; Campos, M.; AttahPoku, S.K.; Redmond, M.J.; Cordeiro, D.M.; Sethi, M.S.; Harland, R.J.; Babiuk, L.A. Serum haptoglobin as an indicator of the acute phase response in bovine respiratory disease. Vet. Immunol. Immunop. 1996, 51, 277–292. [Google Scholar] [CrossRef]
- Burciaga-Robles, L.O.; Holland, B.P.; Step, D.L.; Krehbiel, C.R.; McMillen, G.L.; Richards, C.J.; Sims, L.E.; Jeffers, J.D.; Namjou, K.; McCann, P.J. Evaluation of breath biomarkers and serum haptoglobin concentration for diagnosis of bovine respiratory disease in heifers newly arrived at a feedlot. Am. J. Vet. Res. 2009, 70, 1291–1298. [Google Scholar] [CrossRef]
- Kvidera, S.K.; Horst, E.A.; Abuajamieh, M.; Mayorga, E.J.; Sanz Fernandez, M.V.; Baumgard, L.H. Technical note: A procedure to estimate glucose requirements of an activated immune system in steers. J. Anim. Sci. 2016, 94, 4591–4599. [Google Scholar] [CrossRef]
- Ingvartsen, K.L.; Moyes, K. Nutrition, immune function and health of dairy cattle. Anim. Int. J. Anim. Biosci. 2013, 7 (Suppl. S1), 112–122. [Google Scholar] [CrossRef]
- Aubert, A.; Kelley, K.W.; Dantzer, R. Differential effect of lipopolysaccharide on food hoarding behavior and food consumption in rats. Brain Behav. Immun. 1997, 11, 229–238. [Google Scholar] [CrossRef]
- Wang, A.; Huen, S.C.; Luan, H.H.; Yu, S.; Zhang, C.; Gallezot, J.-D.; Booth, C.J.; Medzhitov, R. Opposing Effects of Fasting Metabolism on Tissue Tolerance in Bacterial and Viral Inflammation. Cell 2016, 166, 1512–1525.e1512. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| p-Value | |||||||
|---|---|---|---|---|---|---|---|
| Variable | Day | Heifer | Steer | SEM | Sex | Time | Sex × Time |
| BoHV-1 titers | |||||||
| Pre-screen | 5.5 | 5.7 | 16.9 | ||||
| d −3 | 6.5 | 4.0 | 16.9 | ||||
| d 0 | 12.7 | 4.6 | 18.2 | ||||
| d 7 | 104.5 | 56.0 | 18.2 | 0.22 | <0.01 | 0.44 | |
| p-Value | ||||||
|---|---|---|---|---|---|---|
| Variable | Heifer | Steer | SEM | Sex | Time | Sex × Time |
| Red blood cells, M/µL | 8.64 | 8.48 | 0.56 | 0.83 | <0.01 | 0.55 |
| Hemoglobin, g/dL | 10.90 | 10.77 | 0.28 | 0.73 | <0.01 | 0.49 |
| Hematocrit, % | 34.29 | 34.39 | 0.96 | 0.93 | <0.01 | 0.48 |
| Platelets, K/µL | 516.84 | 456.21 | 38.13 | 0.27 | <0.01 | 0.36 |
| White blood cells, K/µL | 10.51 | 10.02 | 0.60 | 0.57 | 0.02 | 0.05 |
| Neutrophils, K/µL | 2.56 | 4.26 | 0.45 | 0.02 | <0.01 | 0.03 |
| Lymphocytes, K/µL | 5.76 | 4.48 | 0.35 | 0.02 | 0.02 | <0.01 |
| Neutrophil:Lymphocyte | 0.60 | 1.17 | 0.10 | <0.01 | <0.01 | <0.01 |
| Monocytes, K/µL | 2.23 | 1.77 | 0.09 | <0.01 | <0.01 | 0.01 |
| Eosinophils, K/µL | 0.03 | 0.03 | 0.01 | 0.74 | <0.01 | <0.01 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Burdick Sanchez, N.C.; Broadway, P.R.; Carroll, J.A. Sexual Dimorphic Innate Immune Response to a Viral–Bacterial Respiratory Disease Challenge in Beef Calves. Vet. Sci. 2022, 9, 696. https://doi.org/10.3390/vetsci9120696
Burdick Sanchez NC, Broadway PR, Carroll JA. Sexual Dimorphic Innate Immune Response to a Viral–Bacterial Respiratory Disease Challenge in Beef Calves. Veterinary Sciences. 2022; 9(12):696. https://doi.org/10.3390/vetsci9120696
Chicago/Turabian StyleBurdick Sanchez, Nicole C., Paul R. Broadway, and Jeffery A. Carroll. 2022. "Sexual Dimorphic Innate Immune Response to a Viral–Bacterial Respiratory Disease Challenge in Beef Calves" Veterinary Sciences 9, no. 12: 696. https://doi.org/10.3390/vetsci9120696
APA StyleBurdick Sanchez, N. C., Broadway, P. R., & Carroll, J. A. (2022). Sexual Dimorphic Innate Immune Response to a Viral–Bacterial Respiratory Disease Challenge in Beef Calves. Veterinary Sciences, 9(12), 696. https://doi.org/10.3390/vetsci9120696